Neuromodulation of Cortical Targets in Freezing of Gait
Gonzalo J. Revuelta, Daniel Lench, Carla Silva‐Batista, Marian L. Dale, Martina Mancini

TL;DR
This paper explores how noninvasive brain stimulation can help understand and treat freezing of gait in Parkinson's Disease by targeting specific brain regions.
Contribution
The paper introduces a systematic framework for interpreting network changes in freezing of gait using neuromodulation.
Findings
Noninvasive neuromodulation can alter cortical networks involved in freezing of gait behavior.
Different stimulation parameters can reveal adaptive or maladaptive contributions of cortical targets.
Multimodal outcomes are needed to better understand the relationship between network changes and gait behavior.
Abstract
Freezing of gait (FOG) is a disabling feature of Parkinson's Disease (PD) with unclear underlying pathophysiology. Evidence from multimodal neuroimaging studies suggests that complex interactions between cortical and subcortical areas may occur in FOG. While noninvasive neuromodulation techniques, such as transcranial magnetic stimulation (TMS), can effectively modulate large‐scale networks involved in FOG, the development of noninvasive neuromodulation interventions is limited by an incomplete understanding of the interactions between underlying network disruptions and FOG behavior. Recent studies have brought into question whether observed network changes in FOG are truly causal or secondary, and if secondary, are they adaptive, maladaptive, or not related? Although these questions go beyond correlative analyses, neuromodulation approaches provide an opportunity to systematically…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
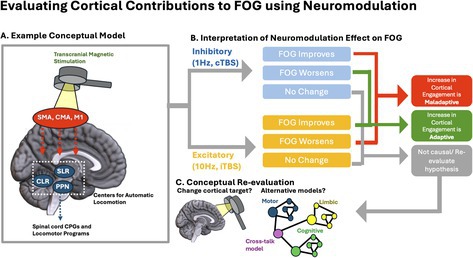 Figure 1
Figure 1| Excitatory studies that improve FOG | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| TMS | |||||||||
| Modality | Design |
| Number of sessions | Dose (total pulses/current) | Target | Outcome measures | Significant results | Durability | |
| Kim et al., 2015 (Kim et al. | 10 Hz rTMS vs. Sham | RCT | 17 PD with FOG | 5 sessions | 5000 pulses | Primary Motor Cortex (M1) leg area |
180 Turns FOGQ TUG UPDRS |
1.3‐step improvement in the 180° turn test. 1.6‐point improvement in FOG‐Q. 2‐s improvement in TUG. 1.4‐point improvement in UPDRS‐III. | All significant improvements were maintained 1 week after intervention. |
| Mi et al. 2019 (Mi et al. | 10 Hz rTMS vs. Sham | RCT | 30 PD with FOG | 10 sessions | 10,000 pulses | SMA |
FOGQ UPDRS Turn duration |
Immediate 1.6‐point improvement in FOG‐Q after intervention 2.13‐point improvement in FOG‐Q at 4 weeks post‐intervention | Improvements in FOGQ persisted at least 4 weeks after intervention. |
| Potvin‐Desrochers et al. 2023 (Potvin‐Desrochers et al. | iTBS, cTBS, Sham | RCT | 14 PD with FOG | 4 sessions | Not Specified | PPC |
Noninstrumented FOG‐provoking task |
Decrease in the number of freezing episodes from a median of 6 to 4 episodes Reduction in percent time frozen during the task from 14% to 10%
No improvements | Not assessed. |
| Lee et al. 2014 (Lee et al. | 10 Hz rTMS vs. Sham | RCT, Crossover | 20 PD with FOG | Single session | 1000 pulses |
SMA M1‐LL DLPFC On dominant hemisphere |
TUG Turn time and Step number UPDRS FOGQ |
M1‐LL stimulation reduced TUG time from 52.7 ± 13.0 s to 38.0 ± 5.8 s. DLPFC stimulation reduced TUG time from 47.3 ± 9.7 s to 42.0 ± 8.6 s
| Not assessed |
| tDCS | |||||||||
| Dagan et al. 2018 (Dagan et al. | Anodal tDCS vs. Sham | RCT, Crossover | 20 PD with FOG | 3 sessions (20 min each):
M1 only M1 + DLPFC Sham | 2 mA continuously over 20 min |
M1 Left DLPFC |
FOG provoking task TUG Stroop test |
Significant reduction in FOG episodes during simultaneous M1 + Left DLPFC tDCS compared to M1 only or sham, 88% of participants (15 out of 17) showed improvement in FOG. Exact amount of reduction not specified | Not assessed. |
| Valentino et al. 2014 (Valentino et al. | Anodal tDCS vs. Sham |
RCT, Crossover | 10 with FOG | 5 consecutive daily sessions | 2 mA continuously over 20 min | M1 |
Stand Walk Sit (SWS) FOGQ Gait & Fall Questionnaire UPDRS |
Significant reduction in the number and duration of FOG episodes. Reduction in the number of steps and time needed to complete the SWS.
Significant improvement | 1 month |
|
| |||||||||
| TMS | |||||||||
| Brugger et al. 2020 (Brugger et al. | iTBS vs. Sham | RCT | 23 (12 PD with FOG, and 11 PD without FOG) | Not Specified | Not Specified | BL SMC |
3D gait analysis (stride time, stride length) FOGQ UPDRS FAB MMSE |
iTBS over the SMC resulted in a relative worsening of gait initiation parameters in PD patients with FOG. Numerical values were not reported | Not assessed. |
| Tard et al. 2016 (Tard et al. | iTBS vs. Sham |
RCT, Crossover | 15 PD with FOG | 2 sessions (one iTBS, one sham) | 600 pulses over 3 min |
Percent time with FOG 3D analysis | No improvement | Not assessed. | |
| Rektorova et al. 2007 (Rektorova et al. | 10 Hz rTMS vs. Sham | RCT | 6 PD with FOG | 5 Consecutive daily sessions | 6750 pulses |
M1 L DLPFC |
Video analysis UPDRS Neuropsychological Battery | No improvement | Terminated due to patient withdrawal. |
| tDCS | |||||||||
| Lu et al. 2018 (Lu et al. | Anodal tDCS vs. Sham |
RCT, Crossover | 10 PD with FOG | 2 sessions (one tdcs one sham) | 1 mA continuously for 10 min | SMA |
APAs | No improvement | Not assessed. |
| Combined TMS/tDCS | |||||||||
| Chang et al. 2017 (Chang et al. | 10 Hz rTMS and Anodal tDCS vs. Sham | RCT | 32 PD patients (16 dual‐mode stimulation, 16 rTMS only) | 5 sessions |
rTMS: 5000 pulses total tDCS: 1 mA for 20 min |
rTMS: M1‐LL tDCS: Left DLPFC |
FOGQ 180 Turn UPDRS TUG | No improvements | Limited to 1 week after intervention. |
|
| |||||||||
| TMS | |||||||||
| Degan et al. 2017 (Dagan et al. | 10 Hz rTMS vs. Sham | RCT | 9 PD with FOG | 16 sessions | 33,600 pulses | mPFC |
FOG provoking task NFOGQ UPDRS Spatiotemporals (gait speed, stride length, step time, swing time) |
Significant reduction in FOG severity Large effect sizes reported (0.90 after intensive phase, 0.82 after maintenance phase) | Not assessed. |
| tDCS | |||||||||
| Lee et al. 2021 (Lee and Kim | Anodal tDCS + Visual Cueing vs. Sham | RCT | 30 PD patients (FOG was not a requirement; exact number with FOG not provided) | 20 sessions | 2 mA continuously for 20 min | SMA |
FOGQ Functional Gait Assessment (FGA) Cadence UPDRS Spatiotemporal gait parameters (measured using GAITRite system) |
No improvements Improvement in gait and cadence | Limited to 2 weeks after intervention ended |
| Manor et al. 2021 (Manor et al. | tDCS vs. Sham | RCT | 77 PD with FOG | 15 sessions | Not specified |
Anodal tDCS: Left DLPFC Cathodal tDCS: BL M1‐LL |
FOG Provoking task UPDRS nFOGQ |
No overall improvement in FOG symptoms Subset of participants with mild FOG severity showed improvement | Increased daily living step counts observed after 2 weeks of stimulation and at 10‐week follow‐up |
|
| |||||||||
| Lench et al. 2021 (Lench et al. | 1 Hz rTMS + Gait Training vs. Sham + Gait Training | RCT | 20 PD with FOG | 10 sessions | 12,000 | SMA |
nFOGQ Spatiotemporals rs‐fMRI UPDRS |
No statistically significant improvement in nFOG‐Q scores between active and sham groups Active rTMS reduced SMA connectivity to regions involved in executive control and attention | 3 months |
- —National Institute of Neurological Disorders and Stroke10.13039/100000065
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurological disorders and treatments · Transcranial Magnetic Stimulation Studies · Functional Brain Connectivity Studies
Background
1
Under the prevailing current pathophysiological model of Parkinson's disease (PD), degeneration of dopaminergic neurons in the substantia nigra pars compacta produces marked dopaminergic denervation of the posterior putamen. This region is critical for habitual motor control; thus, its dysfunction results in impaired movement automaticity and increased reliance on goal‐directed, attention‐demanding motor control (Redgrave et al. 2010). Imaging studies support this model: healthy individuals consistently recruit the posterior putamen during automatic movements, whereas individuals with PD show a shift toward activation of more rostral striatal regions and frontoparietal cortical regions when performing the same tasks. As a result, individuals with PD rely on executive resources to regulate gait, a process normally executed automatically (Wu et al. 2015).
Within this framework, freezing of gait (FOG) can be viewed as a loss of automatic control of locomotion that must be compensated by volitional control until the system becomes overwhelmed, for example, during dual‐tasking, high cognitive load, or heightened limbic drive (Lewis and Shine 2016; Georgiades et al. 2019; Shine, Matar, Ward, Bolitho, et al. 2013). Multiple conceptual models have been proposed to explain this breakdown (Nutt et al. 2011), including the interference model, in which competing cognitive–motor demands overload limited executive control capacity (exemplified above), and the reduced capacity model, which posits deficits across multiple domains/networks (Lewis and Shine 2016; Georgiades et al. 2019; Shine, Matar, Ward, Bolitho, et al. 2013). Here, we focus on how neuromodulation of cortical targets can be used to address fundamental questions about the FOG pathophysiology and to guide future therapeutic development.
In order to interpret the network changes that have been reported in FOG (e.g., altered frontoparietal activation, disrupted cortico‐striatal and cortico‐brainstem coupling) it is critical to understand if such changes are compensatory adaptations (either effective or ineffective) or truly maladaptive responses that contribute to FOG severity. Furthermore, behavioral associations to imaging findings remain unclear, as it is unknown whether the observed imaging findings are primary (causal) drivers of FOG or secondary consequences of gait breakdown. To address these knowledge gaps, neuromodulation strategies have been utilized to either strengthen or weaken cortical–subcortical functional connections, network synchrony, coherence, or coupling, testing varied hypotheses and conceptual models. It is important to recognize that the interactions that occur within these networks are complex; however, evidence so far suggests the presence of systems‐level dysfunction in FOG and neuromodulation interventions currently available can effect network changes leading to measurable behavioral effects.
Definitions
2
Gait Automaticity
2.1
The ability of the central nervous system to control walking with minimal use of executive control (which demands attention) (Clark 2015). Proxies of loss of gait automaticity in PD include increased prefrontal cortex (PFC) activity during walking, higher dual‐task cost of gait, and greater gait variability. In fact, people with FOG usually show higher PFC activity (Vitorio et al. 2020; Bardakan et al. 2022), higher dual‐task cost of walking (de Souza Fortaleza et al. 2017; Monaghan et al. 2023), and greater gait variability (Vitorio et al. 2020; Hausdorff, Balash, and Giladi 2003; Nanhoe‐Mahabier et al. 2011) compared to people without FOG.
Volitional Control
2.2
Motor tasks that are deliberate and attention dependent require significant executive resources.
Adaptive Network Responses
2.3
Any measurable network change that occurs as a result of FOG behavior can be considered adaptive. Adaptive changes are understood to be secondary, as opposed to primary or causal. Adaptive changes can be effective, which would imply that interventions to strengthen these changes can improve FOG behavior; ineffective, which would imply that interventions to increase or decrease these observed changes are unlikely to have any effect on FOG behavior.
Maladaptive Network Responses
2.4
Measurable network changes that occur as a result of FOG can also worsen FOG. Maladaptive network responses can be viewed as bad habits that need to be broken in order to facilitate rehabilitation or a positive behavioral change. Neuromodulation interventions targeting maladaptive changes would aim to reverse these changes in order to improve FOG behavior.
Electrophysiologcal and Imaging Evidence for Maladaptive Network Response in FOG
3
Highly automatized motor skills which have been learned over time (including locomotion) require little to no volitional control. In fact, volitional control may be disruptive in the context of such behaviors. For example, skilled athletes frequently experience performance deterioration when they “overthink” a well learned skill, an analogy relevant to PD and FOG. Conceptually, gait becomes increasingly attention dependent, requiring recruitment of cognitive resources that were not needed when automaticity was intact. Although this shift may initially represent a beneficial adaptation, excessive cognitive monitoring of gait may ultimately become maladaptive in a manner similar to overfocused motor control in skilled athletes.
At the circuit‐level, overreliance on cortical structures involved in volitional control of gait may interfere with brainstem and basal ganglia locomotor circuits that normally drive automatic gait. Anatomical studies demonstrate direct projections from the premotor and supplementary motor area (SMA) to subcortical locomotor centers, including components of the mesencephalic locomotor region (MLR) (Matsumura et al. 2000). Electrophysiological recordings in humans have shown that the pedunculopontine nucleus (PPN), a key structure within the MLR, interacts with the SMA as evidenced by changes in beta coherence prior to movement initiation (Tsang et al. 2010). While some degree of cortical‐brainstem coupling is likely necessary for adaptive modulation of gait, several studies report excessive coupling or abnormally increased SMA–PPN connectivity in individuals with FOG (Lench et al. 2020; Lench et al. 2021; Fling et al. 2014).
Several neuroimaging studies (fMRI) implicate frontal and frontostriatal dysfunction in FOG (Shine, Matar, Ward, Bolitho, et al. 2013; Gilat et al. 2017; Shine, Matar, Ward, Frank, et al. 2013; Ehgoetz Martens et al. 2018). FOG appears to arise when the balance between automatic motor control and cognitively mediated control processes destabilizes (Vandenbossche et al. 2012) through impaired integration of cognitive control networks (e.g., executive and attentional networks) (Shine, Matar, Ward, Frank, et al. 2013; Shine, Moustafa, Matar, Frank, and Lewis 2013). Virtual reality fMRI gait paradigm consistently shows heightened frontal and parietal activation combined with reduced activity in subcortical regions during FOG episodes (Shine, Matar, Ward, Bolitho, et al. 2013; Gilat et al. 2017; Shine, Matar, Ward, Frank, et al. 2013). In addition, FOG episodes (off medication) are associated with reduced synchrony across corticostriatal pathways (Ehgoetz Martens et al. 2018).
Increasingly complex models have considered how multi‐domain networks can contribute to FOG behavior. The “cross‐talk” model of FOG, for example, suggests that excessive communication and conflict between cognitive, motor, and limbic networks can lead to FOG. Neural networks, which are usually segregated in healthy controls, can become over‐integrated during pathological “freezing” states. This integration appears to increase during times in which PD patients are in their OFF medication state (Shine et al. 2019). However, neurotransmitters other than dopamine are also likely involved in changes in pathological network integration. For example, the noradrenergic system is thought to drive the integration of networks by increasing neural gain across the entire brain. Furthermore, these topological features of networks are dynamic and fluctuate over time, which aligns with the episodic and paroxysmal nature of FOG (Shine et al. 2016). Overall, this conceptual framework provides a plausible explanation of how high levels of anxiety, arousal, and cognitive load may influence cortical networks to disrupt the effectiveness of automatic gait circuits to function properly (Taylor et al. 2022). At this time, causal studies are needed to determine the precise extent to which network integration is maladaptive or adaptive. Furthermore, it remains unknown what levels of network integration or segregation are optimal for gait performance.
A major limitation of fMRI studies in investigating cortical contributions to gait is that subjects cannot ambulate in the scanner environment. Mobile neuroimaging methods, such as functional near infrared spectroscopy (fNIRS), have been used to study cortical activation, more commonly PFC activation, during walking and turning in people with PD with and without FOG (Vitorio et al. 2020; Ranchet et al. 2020; Hamacher et al. 2015; Herold et al. 2017; Maidan et al. 2016; Silva‐Batista et al. 2024; Belluscio et al. 2019). Gait during dual‐task conditions and turning in place have been the main triggers for evoking FOG in the laboratory setting (Conde et al. 2023). Using fNIRS, several reports show increased PFC activity during dual‐task walking (Vitorio et al. 2020), 360° turning in place (Belluscio et al. 2019), and immediately before and during FOG episodes triggered by an unanticipated 180° turning while walking (Maidan et al. 2015). Recent work expanding beyond the PFC demonstrated that freezers also show elevated activation in SMA, premotor cortex, and posterior parietal cortex during gait‐related tasks (turning, stopping while walking). However, during actual FOG episodes, cortical activation shows a distinct and abnormal pattern compared to voluntary stopping: PFC activity decreases markedly while SMA, premotor cortex, and posterior parietal cortex do not show a significant change, suggesting that no cortical region effectively “steps in” during the freeze. This pattern suggests that FOG may be driven by a disruption or imbalance within the stopping‐related network, particularly between SMA and PFC, leading to a failure of coordinated cortical–subcortical control at the moment of freezing.
Taken together, these fMRI and fNIRS findings suggest that people with FOG demonstrated increased activity in fronto‐parietal regions in an attempt to compensate for striatal dysfunction leading to reduced automaticity of gait and increased stride‐to‐stride variability (Hausdorff, Schaafsma, et al. 2003; Gilat et al. 2013; Plotnik et al. 2005). Reliance on these cognitive control networks aligns with evidence that people with PD increasingly depend on executive resources and external cues in order to generate effective locomotion by switching to goal‐directed behavior (Redgrave et al. 2010; Gilat et al. 2021). However, this compensatory strategy may become inefficient in two cases: (1) during challenging conditions that further rely on PFC, such as when multi‐tasking or during turns, when the load on the frontal networks may become unsustainable (overload), thus leading to FOG (Shine, Matar, Ward, Frank, et al. 2013; Vandenbossche et al. 2012; Shine, Moustafa, Matar, Frank, and Lewis 2013) or (2) the degeneration of the compensatory cortical areas that occurs with disease progression. A recent review suggests that compensatory recruitment of cognitive and sensory networks initially supports gait, but deteriorates over time, leading to more frequent breakdown of gait and FOG (Tosserams et al. 2025). While studies show correlations between worsening FOG and greater cortical control over gait (Belluscio et al. 2019; Maidan et al. 2015; Shine et al. 2014), it remains unclear if this is due to worsening FOG, which engages greater cortical areas, or truly a maladaptive process that contributes to worsening FOG severity. To understand this relationship, manipulating cortical targets systematically via neuromodulation approaches can be highly informative.
Review of Neuromodulation Interventions Designed to Improve Adaptive Responses in FOG
4
A comparative analysis of the available evidence on neuromodulation of cortical control is challenging, given the high number of variables, including neuromodulation modality, study design, target, and choice of FOG outcome measures. We sought to represent neuromodulation modalities that addressed FOG specifically in idiopathic PD and excluded open‐label studies as it is difficult to draw conclusions from these findings. The objective of this summary was not to evaluate clinical evidence toward efficacy, but rather to elucidate the effects of experimental manipulation of relevant networks on FOG severity.
Overall, improvements reported from excitatory repetitive transcranial magnetic stimulation (rTMS) or transcranial direct current stimulation (tDCS) approaches were modest, with up to 2 points improvement in the Freezing of Gait Questionnaire (FOGQ), reduction of two FOG episodes, 4% reduction of percent time frozen in FOG provoking tasks, and one study showed up to a 15‐s reduction in TUG time in the active group but no significant improvement in FOGQ. Several excitatory studies did not show any efficacy or were equivocal (see Table 1).
Review of Neuromodulation Interventions Designed to Diminish Maladaptive Response in FOG
5
While the vast majority of neuromodulation studies for FOG have been performed to increase cortical control, only two studies used rTMS to reduce cortical control of gait (Lench et al. 2021). One comparative study of cTBS and iTBS reported a small improvement in the iTBS (excitatory) group (2 freezing episodes or 4% reduction) and no improvement in the inhibitory group (Potvin‐Desrochers et al. 2023). The other study (Lench et al. 2021) (conducted by authors DL and GJR) tested the framework that hyper‐connectivity of cortical brain regions to automatic gait centers could be dampened by using an inhibitory form of stimulation. In this study, 20 participants with FOG were randomized into either a sham or active TMS treatment over 10 days. 1 Hz rTMS was administered at a suprathreshold level (110% rMT) over the SMA and combined with daily gait training sessions. These 20‐min training sessions were used to promote gait automaticity by having participants focus on a cognitive task while maintaining a constant gait velocity. This study found a 4.8‐point reduction on the New Freezing of Gait Questionnaire FOG‐Q (nFOG‐Q) following the active TMS intervention and a modest 1.8‐point reduction following sham TMS intervention which included gait training. Although pre‐ to post‐changes in the active group were significant, between‐group differences were not significant in this small sample. Reductions in SMA connectivity were found with several brain regions, including the anterior cingulate cortex (ACC), medial PFC, and angular gyrus in the active group. This study provides direct evidence that a reduction in cortical control of gait can lead to an improvement in FOG.
Taken together, there is reasonable evidence that reducing cortical control of gait through noninvasive forms of stimulation may in fact alleviate FOG. Future studies that compare stimulation protocols and perform more standardized interventions to facilitate gait automaticity will be needed to provide more conclusive evidence.
Conclusion
6
Here we explore the role of cortical (volitional) control of locomotion and present a conceptual framework in which modulating cortical input into subcortical locomotor centers may improve FOG. Given the development of noninvasive neuromodulation for cortical targets, we propose that neuromodulation interventions for FOG can address two important knowledge gaps: (1) is cortical activation, involving PFC, premotor, and SMA, an ineffective adaptation or a maladaptive response? and (2) should these changes be enhanced or inhibited to improve FOG? Currently, the vast majority of the literature in this field has aimed to increase cortical control of locomotion via neuromodulation and rehabilitation approaches with largely equivocal, negative, or subclinical effects. We propose that under this framework, cortical inhibitory strategies, either in isolation or combined with approaches to improve gait automaticity, deserve systematic investigation.
Although it is difficult to compare efficacy across studies due to differences in modalities, targets, design, and particularly the lack of standardized outcome measures, the FOGQ and nFOGQ have been most widely used, allowing for a more direct comparison of the magnitude of effects of multiple studies. The nFOGQ, however, has been criticized due to having a large minimum detectable change (Hulzinga et al. 2020), which is greater than any of the reported differences. It is important to note that the reported improvements in the limited number of inhibitory studies are at least as large as or greater in magnitude than the excitatory studies; however, future studies should include objective measures of FOG severity.
Interpreting neuromodulation effects is further complicated by the fact that inhibitory or excitatory strategies do not always result in the intended effect, particularly with more advanced or accelerated approaches like theta burst stimulation (TBS) (McCalley et al. 2021). As our understanding of the individual variability of FOG grows, and the variability of the effects of noninvasive neuromodulation interventions, future studies may pursue personalized interventions based on individual‐level findings that guide network‐specific interventions.
It is important to highlight that only one study (Hausdorff, Balash, and Giladi 2003) investigated the mechanistic effects of the intervention. This allows for accurate interpretation of the behavioral effects of the intervention. Clinical improvement (or lack thereof) of an investigational therapy can be a result of a variety of factors including: target, dose, modality, selected outcomes and directionality of the intervention. Therefore a null effect is not informative unless the study is designed to detect target engagement and the network effect of the intervention. If the network effect was normalized and there was no behavioral effect, this finding is much more meaningful as it provides critical data to clarify the relationship between the network change and FOG behavior itself. Thus far, there is evidence from one study that the SMA target can be engaged, and SMA connections can be decreased with rTMS, resulting in a positive effect on FOG behavior. There is also evidence of positive and null effects with excitatory neuromodulation interventions on a variety of targets, which are difficult to interpret with limited data on target engagement and network effects.
While current neuromodulation studies have primarily focused on either enhancing or inhibiting cortical control, we acknowledge that these do not encompass other more complex models that underlie FOG, such as the “cross‐talk” model. Despite this gap in research, we believe that modern neuromodulation techniques may also have the potential to interrogate the impact of cortical network features that have not been traditionally evaluated in a causal manner, such as network integration. To date, most cortical neuromodulation approaches for FOG have been focused on rTMS and tDCS. However, other neuromodulation approaches, like transcranial alternating current stimulation (tACS), which can entrain neural oscillations across the brain, have the potential to modulate the synchrony of neural activity across spatially distributed networks (Bachinger et al. 2017). Additionally, transauricular vagus nerve stimulation (taVNS) can transiently modulate noradrenergic brain centers such as the locus ceruleus, which have previously been shown to globally drive network integration and segregation (Wienke et al. 2023). These noninvasive neuromodulation approaches, paired with the advancement of more dynamic neuroimaging techniques, are a promising avenue to elucidate the adaptive or maladaptive role of cortical network integration and “cross‐talk” in FOG.
As the field advances toward accelerated stimulation paradigms, individualized targeting, and combined neuromodulation‐rehabilitation approaches, it will be critical to evaluate both behavioral outcomes of FOG and the underlying network effects. We encourage future studies to include neuroimaging and neurophysiological outcomes along with standardized behavioral outcomes for FOG. Such mechanistic–behavioral coupling will be essential for determining which cortical processes should be suppressed, enhanced, or rebalanced to meaningfully improve FOG.
Author Contributions
Gonzalo J. Revuelta: conceptualization, data curation, formal analysis, funding acquisition, methodology, writing – original draft, writing – review and editing. Daniel Lench: conceptualization, writing – review and editing. Carla Silva‐Batista: writing – review and editing. Marian L. Dale: conceptualization, writing – review and editing. Martina Mancini: conceptualization, writing – review and editing.
Funding
This work was supported by the National Institute of Neurological Disorders and Stroke, (1R01NS131396, R00NS131447).
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bachinger, M. , V. Zerbi , M. Moisa , et al. 2017. “Concurrent TACS‐FMRI Reveals Causal Influence of Power Synchronized Neural Activity on Resting State FMRI Connectivity.” Journal of Neuroscience 37, no. 18: 4766–4777. 10.1523/JNEUROSCI.1756-16.2017.28385876 PMC 6596494 · doi ↗ · pubmed ↗
- 2Bardakan, M. M. , G. R. Fink , L. Zapparoli , G. Bottini , E. Paulesu , and P. H. Weiss . 2022. “Imaging the Neural Underpinnings of Freezing of Gait in Parkinson's Disease.” Neuro Image: Clinical 35: 103123. 10.1016/j.nicl.2022.103123.35917720 PMC 9421505 · doi ↗ · pubmed ↗
- 3Belluscio, V. , S. Stuart , E. Bergamini , G. Vannozzi , and M. Mancini . 2019. “The Association Between Prefrontal Cortex Activity and Turning Behavior in People With and Without Freezing of Gait.” Neuroscience 416: 168–176. 10.1016/j.neuroscience.2019.07.024.31330231 PMC 7778469 · doi ↗ · pubmed ↗
- 4Brugger, F. , R. Wegener , J. Walch , et al. 2020. “Altered Activation and Connectivity of the Supplementary Motor Cortex at Motor Initiation in Parkinson's Disease Patients With Freezing.” Clinical Neurophysiology 131, no. 9: 2171–2180. 10.1016/j.clinph.2020.05.023.32683125 · doi ↗ · pubmed ↗
- 5Chang, W. H. , M. S. Kim , E. Park , et al. 2017. “Effect of Dual‐Mode and Dual‐Site Noninvasive Brain Stimulation on Freezing of Gait in Patients With Parkinson Disease.” Archives of Physical Medicine and Rehabilitation 98, no. 7: 1283–1290. 10.1016/j.apmr.2017.01.011.28193533 · doi ↗ · pubmed ↗
- 6Clark, D. J. 2015. “Automaticity of Walking: Functional Significance, Mechanisms, Measurement and Rehabilitation Strategies.” Frontiers in Human Neuroscience 9: 246. 10.3389/fnhum.2015.00246.25999838 PMC 4419715 · doi ↗ · pubmed ↗
- 7Conde, C. I. , C. Lang , C. R. Baumann , C. A. Easthope , W. R. Taylor , and D. K. Ravi . 2023. “Triggers for Freezing of Gait in Individuals With Parkinson's Disease: A Systematic Review.” Frontiers in Neurology 14: 1326300. 10.3389/fneur.2023.1326300.38187152 PMC 10771308 · doi ↗ · pubmed ↗
- 8Dagan, M. , T. Herman , R. Harrison , et al. 2018. “Multitarget Transcranial Direct Current Stimulation for Freezing of Gait in Parkinson's Disease.” Movement Disorders 33, no. 4: 642–646. 10.1002/mds.27300.29436740 PMC 5964604 · doi ↗ · pubmed ↗