The single and combined effects of deltamethrin and polyethylene microplastics on the development and biochemical responses of Xenopus laevis in early life stages
Duygu Ozhan Turhan, Cihan Anıl Benli, Muhittin Yurekli, Abbas Güngördü

TL;DR
This study examines how deltamethrin and polyethylene microplastics affect Xenopus laevis embryos and tadpoles, finding combined effects that vary depending on the life stage and biomarker.
Contribution
The study introduces a detailed assessment of single and combined toxicity of deltamethrin and microplastics in amphibians, highlighting life stage-dependent responses.
Findings
Deltamethrin caused embryotoxicity with LC50 of 68.0 µg/L and EC50 of 7.58 µg/L.
Combined exposures showed additive and antagonistic effects on enzymatic biomarkers.
Tadpoles showed inhibited enzyme activity at high microplastic concentrations.
Abstract
The co-occurrence of microplastics and pesticides in freshwater ecosystems raises concerns for aquatic organisms. However, their combined effects remain poorly understood. In this study, the single and combined effects of deltamethrin (DEL) and polyethylene microplastics (PE-MPs) were investigated in the early life stages of Xenopus laevis. Embryonic developmental toxicity was evaluated under standardized Frog Embryo Teratogenesis Assay–Xenopus (FETAX) conditions in embryos, and sublethal biochemical responses were also assessed in embryos and tadpoles. In the first phase, embryos were exposed for 96 h to a concentration range of DEL (3.125–6,400 µg/L) and PE-MPs (50–1,000 mg/L) to estimate the median lethal concentration (LC50) and the median effective concentration for malformation (EC50), as well as the teratogenic index (TI). In the second phase, assays were performed at DEL 2.72…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
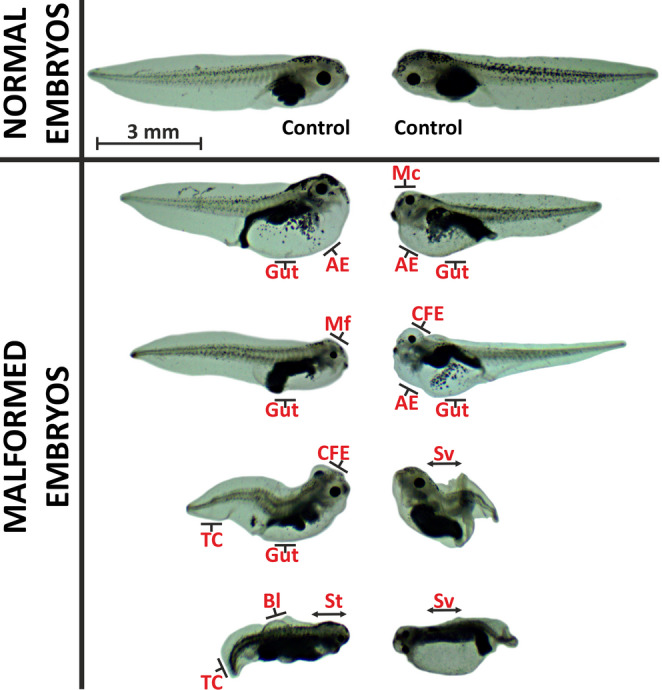 Figure 1
Figure 1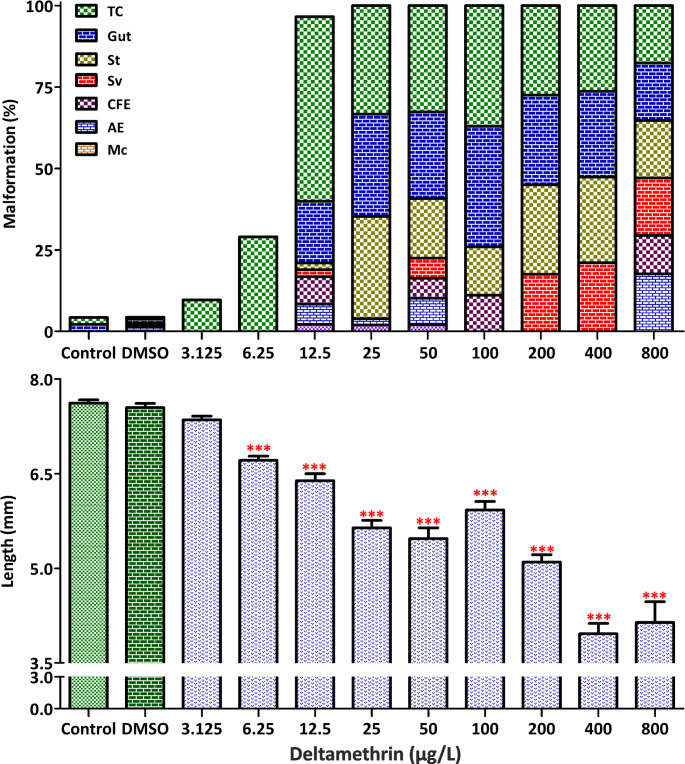 Figure 2
Figure 2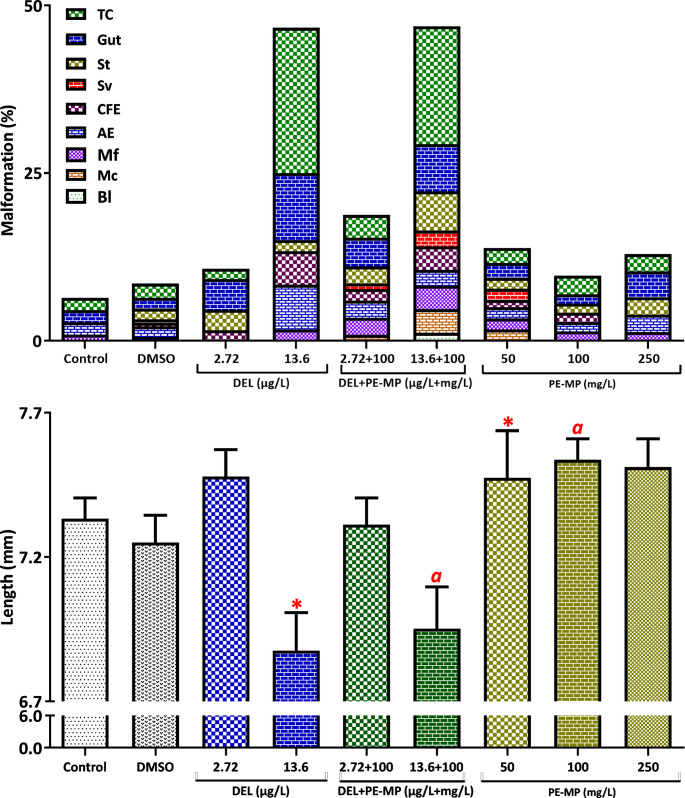 Figure 3
Figure 3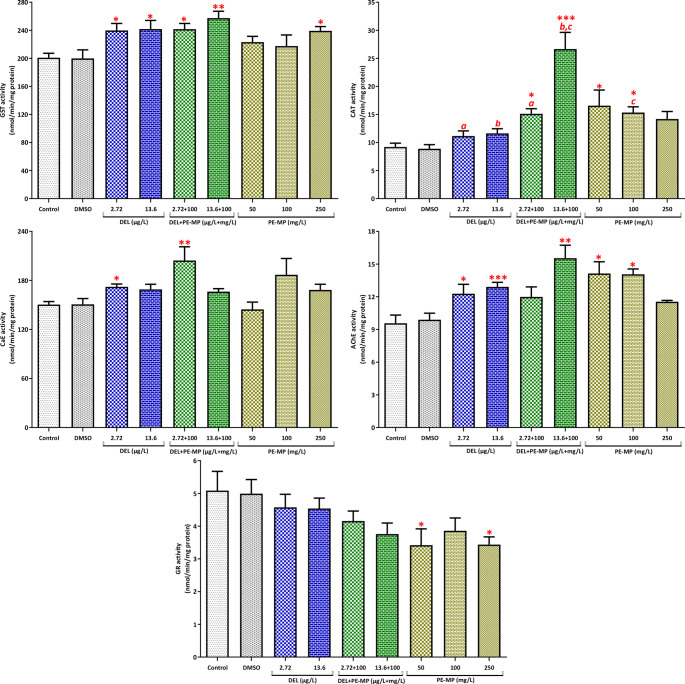 Figure 4
Figure 4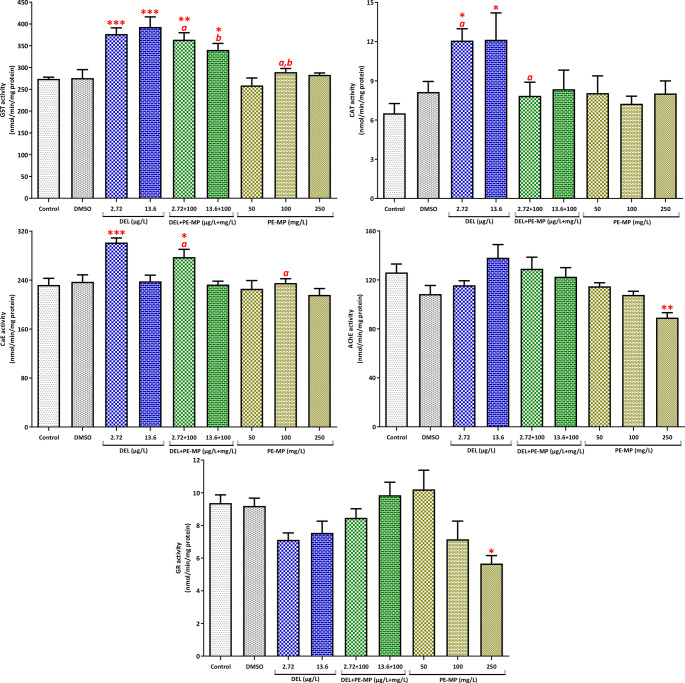 Figure 5
Figure 5- —Inonu University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicroplastics and Plastic Pollution · Marine Biology and Environmental Chemistry · Effects and risks of endocrine disrupting chemicals
Introduction
The term “plastic” refers to a broad class of synthetic polymers derived from petroleum sources, including polyethylene (PE), polystyrene, polypropylene, polyvinyl chloride, and nylon (Anderson et al. 2016). These materials are widely used due to their durability, low density, and low thermal conductivity (Meng et al. 2020). Since their industrial introduction in the 1940s, global plastic production has increased steadily, reaching approximately 400 million tons in 2022, with further growth projected by 2050 (PlasticsEurope 2023). PE is the most prevalent polymer, accounting for about 30% of global plastic demand, with annual production exceeding 140 million tons (Anderson et al. 2016; Houssini et al. 2025).
Plastic waste is pervasive in marine, freshwater, and terrestrial environments, where it degrades via mechanical, photochemical, and biological processes. These processes result in the formation of microplastics (MPs; particles < 5 mm) and nanoplastics (NPs; particles < 100 nm) (Cole et al. 2011). MPs can originate as primary particles, which are released directly from products such as cosmetics, textiles, and tire wear, or as secondary particles, which are formed by the fragmentation of larger plastic debris (Fang et al. 2019). PE-MPs are among the most frequently detected MPs in aquatic environments and display variability in size, shape, and color (Zhang et al. 2018). Due to their persistence and bioavailability, MPs can enter aquatic food webs and may adversely affect ecosystem integrity and potentially increase human exposure (Hasan et al. 2024; Tatlı et al. 2025; Timaná Morales et al. 2025).
Synthetic pyrethroids are a widely used class of insecticides applied in agriculture, veterinary medicine, public health, and domestic settings (El-Demerdash 2007). These compounds are chemically classified as Type I (non-cyano) or Type II (alpha-cyano) pyrethroids (Soderlund 2020). Their neurotoxic mode of action involves modulating sodium, potassium, GABA, and calcium channels, which disrupts nerve transmission and causes paralysis in target pests and non-target aquatic organisms (Ren et al. 2016; Yang et al. 2020). Pyrethroids are highly toxic to fish, amphibians, and other aquatic invertebrates, even at low environmental concentrations, while showing comparatively lower toxicity to birds and mammals (Brander et al. 2009; Hong et al. 2021; Ranatunga et al. 2023).
Deltamethrin (DEL), a type II pyrethroid, is used globally for crop protection, livestock farming, and vector control, including programs targeting disease vectors, such as mosquitoes (Lu et al. 2019; Yadav et al. 2001). Environmental monitoring reports have detected DEL concentrations ranging from a few micrograms per liter in surface waters to milligrams per liter near industrial discharges (Rodrigues et al. 2023). Due to its hydrophobic nature and low water solubility, deltamethrin tends to adsorb onto sediments, organic matter, and microplastic particles, which can affect its environmental fate and toxicity (Horton et al. 2018). Microplastics may act as vectors for hydrophobic organic pollutants, including pesticides, potentially modifying their transport, bioavailability, and toxicity (Trevisan et al. 2019; Wen et al. 2018; Xu et al. 2021). PE-MPs and DEL have been reported to co-occur in freshwater ecosystems (Luo et al. 2021; Soares et al. 2024), yet their combined toxicological effects on aquatic vertebrates remain insufficiently characterized.
Amphibians are particularly vulnerable to chemical pollution due to their permeable skin and life cycles that span both aquatic and terrestrial habitats (Fernandez et al. 2020). Xenopus laevis is a well-established ecotoxicological model widely used in developmental toxicity assays, including the Frog Embryo Teratogenesis Assay*–Xenopus* (FETAX; ASTM 2003), the Amphibian Metamorphosis Assay (AMA; OECD 2009), and the Xenopus Eleutheroembryo Thyroid Assay (OECD 2019). Both pesticides and microplastics are increasingly recognized as emerging contaminants in freshwater ecosystems that can co-occur and exert combined adverse effects on aquatic biota.
Recent studies have reported that microplastics can modify the bioavailability and toxicity of various environmental pollutants through adsorption or co-transport mechanisms (Sunny et al. 2025). For instance, co-exposure of MPs with metals, pharmaceuticals, or pesticides has been shown to induce oxidative stress, genotoxicity, and developmental alterations in amphibians and fish (Rahman et al. 2024; Subaramaniyam et al. 2023; Tang 2025; Zhang et al. 2023). Although mixture toxicity studies involving MPs and pesticides in amphibians remain limited, emerging evidence suggests that MPs can alter contaminant uptake and detoxification capacity – leading to species- and stage-specific responses (Tang 2025; Zhang et al. 2023).
In this context, the present study aimed to evaluate the lethal and sublethal effects of DEL and PE-MPs, both individually and in combination, on the early life stages of X. laevis. A three-tiered experimental design was employed. First, the FETAX assay was used to evaluate mortality, malformations, and developmental delays. Second, the interactive effects of sublethal DEL concentrations (LC_50_/25 and LC_50_/5) alone and in combination with 100 mg/L PE-MPs were evaluated. Third, key enzymatic biomarkers involved in detoxification, oxidative stress, and neurotoxicity, including glutathione S-transferase (GST), glutathione reductase (GR), catalase (CAT), carboxylesterase (CaE), and acetylcholinesterase (AChE), were measured in embryos and tadpoles to elucidate physiological responses. This integrated approach provides novel insights into the complex interactions between pesticide and microplastic mixtures, enhancing our understanding of the ecological risks these contaminants pose to amphibian populations in freshwater environments.
Materials and methods
Chemicals and reagents
Virgin spherical polyethylene (PE) particles (40 μm diameter, 434272, Sigma-Aldrich, USA) and deltamethrin (DEL; Boston BioProducts, BDA-275575, USA) were tested in this study. Morphological verification of the PE-MPs was performed using light microscopy, as the particles were virgin, commercially standardized, and pre-characterized by the supplier. Particle diameters were measured using calibrated ImageFocus software, and descriptive parameters (mean ± SD, median, min–max, D10, D90) were summarized (Supplementary Table S1). Representative frequency histograms and images illustrating morphology and dispersion/aggregation behavior in the medium were provided (Supplementary Figures S3–S4). Additional reagents for biochemical assays, including 1-chloro-2,4-dinitrobenzene (CDNB), 5,5′-dithio-bis(2-nitrobenzoic acid) (DTNB), p-nitrophenyl acetate (PNPA), acetylthiocholine iodide (ACTI), reduced glutathione (GSH), and bovine serum albumin (BSA), were obtained from Sigma-Aldrich. Oxidized glutathione (GSSG) and NADPH were sourced from MP Biomedicals (USA).
Test organisms
Adult Xenopus laevis were obtained from the Aquatic Vertebrates Experimental Unit at Inonu University and maintained at 23 ± 1 °C with a 12 h photoperiod. The FETAX solution used as the control was prepared according to ASTM Type I water standards (ASTM 2003), with the specified salts dissolved in one liter of distilled water. Adult frogs were acclimated for two days prior to hormonally induced breeding using human chorionic gonadotropin (hCG; Chorulon^®^, MSD, Germany; 600 IU for males and 500 IU for females). The fertilized eggs were then transferred to an aerated FETAX solution for exposure. Embryos at Nieuwkoop and Faber stages 8–11 were used for toxicity testing and biochemical assays. Tadpoles were maintained until stage 46 for subsequent biochemical analyses. All animal procedures were approved by the Inonu University Local Ethics Committee for Animal Experiments (Protocol No. 13505) and conducted in accordance with ASTM E1439-98 (ASTM 2003).
FETAX assay for single substances
The FETAX assay was conducted to determine the LC_50_, EC_50_, the minimum concentration to inhibit growth (MCIG), and the teratogenic index (TI) for DEL and PE-MP. DEL concentrations (3.125–6,400 µg/L) were based on preliminary range-finding and literature to cover both environmentally relevant and high-end exposure conditions, as reported in surface waters and near agricultural runoff (Rodrigues et al. 2023; Shi et al. 2024). This range ensured adequate coverage for accurate LC_50_ and TI derivation.
The PE-MP concentrations (50–1,000 mg/L) were selected based on studies of freshwater fish that reported oxidative stress at similar levels (de Souza Freire et al. 2023; Lee et al. 2023), despite the lack of X. laevis–specific data and were used as high-end laboratory concentrations to support mechanistic assessment of sublethal responses and potential thresholds.
DEL stock solution was dissolved in dimethyl sulfoxide (DMSO) and diluted in FETAX; the final DMSO concentration did not exceed 0.006% (v/v). PE-MP solutions were prepared directly in FETAX. The pH, conductivity, and temperature of the FETAX solution used in the study were 7.78 ± 0.04, 1.54 ± 0.01 mS/cm, and 22 ± 0.2 °C, respectively. Embryos (stages 8–11) were exposed in 24-well plates (four embryos per well). Each treatment used 32 embryos (eight wells), while controls used 48 embryos (12 wells). Solutions were replaced every 48 h, and the embryos were examined every 24 h throughout the 96 h exposure period. The surviving embryos were euthanized with MS222 (200 mg/L), fixed in 3% formalin, and photographed for morphological assessment. Lengths were measured using Euromex ImageFocus 4.0 software.
FETAX assay for mixtures
Mixture tests used two DEL concentrations (LC_50_/25: 2.72 µg/L and LC_50_/5: 13.6 µg/L) selected as sublethal, monitoring-relevant concentrations for mechanistic assessment, allowing for sufficient survival and sublethal endpoint assessment (Rodrigues et al. 2023; Shi et al. 2024). The PE-MP concentrations (50, 100, and 250 mg/L) included levels that were previously shown to cause oxidative stress. Mixture groups included each DEL concentration with 100 mg/L PE-MP. The methods mirrored those used for single-substance testing.
Biochemical analyses
Biochemical assessments were performed on X. laevis embryos (stages 8–11) and tadpoles (stage 46) exposed to PE-MPs (50, 100, or 250 mg/L), DEL (2.72–13.6 µg/L), or mixtures of the two (100 mg/L PE-MPs + 2.72 µg/L or 13.6 µg/L DEL). The concentrations were selected based on developmental toxicity results and environmentally relevant sublethal levels. Fifteen individuals per replicate (five replicates per group) were exposed to a 10 mL test solution in 25 mL plastic containers. The solutions were renewed every 48 h, and the embryos were examined at 24 h intervals throughout the 96 h exposure period. Survivors were euthanized with buffered MS222 (200 mg/L), transferred to microcentrifuge tubes on ice, and stored at -80 °C until biochemical analysis.
The samples were homogenized on ice in a 0.1 M potassium phosphate buffer solution (pH 7.4) containing 150 mM KCl, 1 mM EDTA, and 0.05 mM DTT. The homogenates were then centrifuged at 16,000 g for 20 min at 4 °C. The resulting supernatants were used immediately for enzyme assays. Enzyme activities were measured for GST, GR, CAT, CaE, and AChE spectrophotometrically using established methods with slight modifications (Güngördü and Turhan 2024). GST activity was assessed by CDNB conjugation at 344 nm (Habig et al. 1974). GR activity was monitored via DTNB reduction at 405 nm (Stephensen et al. 2000). CAT activity was determined by measuring H₂O₂ decomposition at 240 nm (Aebi 1974). CaE activity was measured using a PNPA substrate adapted for microplate readers (Santhoshkumar and Shivanandappa 1999). AChE activity was quantified using the ACTI substrate by adapting the method of Ellman et al. (1961) for microplates (Ozmen et al. 1998). Total protein content was measured using the Bradford assay (Bradford 1976) to normalize the enzyme activities.
High-performance liquid chromatography (HPLC) analysis
Before exposure, DEL concentrations in the test media were verified using HPLC (1100 System, Agilent Technologies, USA) for biochemical assays. Analyses were conducted with a C18 reversed-phase column (5 μm, 4.6 × 250 mm) at 25 °C using a mobile phase of acetonitrile and distilled water (80:20, v/v) (Mat Hussin et al. 2021) at a flow rate of 1.0 mL/min. The injection volume was 10 µL, and detection was performed at 230 nm. Quantification was based on a calibration curve (1.56–400 µg/L, R² = 0.994). Representative chromatograms and measured DEL concentrations are available in Supplementary Material (Table S2 and Figures S1).
Statistical analysis and calculations
The 96 h LC_50_ and EC_50_ values for DEL were calculated using Finney’s probit analysis with the U.S. EPA Probit Analysis Program (version 1.5). The TI was determined as the ratio of LC_50_ to EC_50_.
All statistical analyses were performed using GraphPad Prism 8 (GraphPad Software, USA). Normality and homogeneity of variances were assessed using Kolmogorov–Smirnov and Bartlett’s tests, respectively. Parametric data were analyzed using a one-way ANOVA followed by a Dunnett’s post hoc test to compare treatments with controls. Nonparametric data were analyzed using the Kruskal–Wallis test, followed by the Dunn multiple comparison test. Statistical significance was set at p < 0.05.
In addition, pairwise comparisons among treatment groups (e.g., between single and mixture exposures) were performed using independent t-tests for normally distributed data or Mann–Whitney U tests for nonparametric data. These additional analyses allowed for a more detailed assessment of specific differences between DEL, PE-MP, and DEL + PE-MP groups at corresponding concentrations.
Statistical power and sample size justification were assessed a priori; detailed power analysis is provided in the Supplementary Materials.
To evaluate the interaction between DEL and PE-MPs in mixtures, we calculated the interaction index (II) following Peluso et al. (2024): II = (M + Co)/(A1 + A2).
M is the mean biomarker response in the mixture group; A1 and A2 are the means of the single-exposure groups (DEL and PE-MPs); and Co is the control mean. For biomarkers that are increased relative to the control group, an II value greater than 1 indicates synergism and an II value less than 1 indicates antagonism. For biomarkers that are decreased relative to the control group, an II value greater than 1 indicates antagonism and an II value less than 1 indicates synergism. Values of II between 0.95 and 1.05 were considered additive. Only mixtures that caused statistically significant differences in biomarkers or in which at least one treatment differed significantly from the control were considered for II analysis.
Results
Toxicity of individual DEL and PE-MP exposures
According to the FETAX assay results, the 72 h and 96 h LC_50_ values for DEL were 2,035 µg/L and 68 µg/L, respectively (Table 1). The 96 h TI was calculated as 8.97, with an EC_50_ of 7.58 µg/L and an MCIG of 3.125 µg/L. Among the surviving embryos exposed to DEL, 67% exhibited malformations. The types of malformations and abbreviations used are defined in Fig. 1. The most frequent malformation types included tail curvature (65%), gut abnormalities (43%), and growth retardation (31%) (Fig. 2; see also Table S3 and Figure S2 in the Supplementary Material for details). Growth inhibition became statistically significant at concentrations ≥ 6.25 µg/L (p < 0.001).
Table 1LC_50_, EC_50_, TI and MCIG values of deltamethrin for X. laevis embryosConc. (µg/L)Conc. #LC_50_ (95% CI) (µg/L)EC_50_ (95% CI)(µg/L)TI(LC_50_/EC_50_)MCIG(µg/L)72 h96 h3.125–6400122035(1397–3230)68.00(48.95–94.90)7.58(7.02–8.17)8.973.125LC_50_ = median lethal concentration; EC_50_ = median effective concentration for malformation; TI = teratogenic index, calculated as LC_50_/EC_50_; MCIG = minimum concentration to inhibit growth. Data are expressed with 95% confidence intervals
In contrast, exposure to PE-MPs alone resulted in ≤ 3% mortality and a total malformation rate of ≤ 4% across all tested concentrations. PE-MPs were verified by light microscopy; the particles were virgin, commercially standardized, and pre-characterized by the supplier. The mean particle diameter was ~ 41 μm (range 17–77 μm), and the particles appeared spherical and relatively smooth under light microscopy (Supplementary Table S1, Figures S3–S4). Consequently, LC_50_, EC_50_, and TI values could not be determined. No concentration-dependent increase in malformation frequency or growth inhibition was observed in PE-MP-treated embryos (see also Table S4 and Figure S5in the Supplementary Material for details).
Fig. 1. Types of malformations observed in X. laevis embryos exposed to the test substances. Malformations: Sv, severe (multiple, severe malformations); St, stunted growth; Gut, gut abnormalities (improper gut coiling and long, loosely coiled gut); AE, abdominal edema; CFE, craniofacial edema (face, head, and eye edema); Mc, microcephaly; Mf, microphthalmia; TC, tail curvature; Bl, blister
Fig. 2. Malformations (%) and body length in embryos exposed to deltamethrin (DEL) concentrations for 96 h under FETAX (see also Table S3). Malformations as defined in Fig. 1. Bars show mean ± SE (n = 8 wells × 4 embryos/well). *p < 0.001 compared with control (one-way ANOVA with Dunnett’s or Kruskal–Wallis with Dunn’s, as appropriate)
Toxicity of DEL and PE-MP mixtures
Mixture tests combined two sublethal DEL concentrations (2.72 and 13.6 µg/L) with three PE-MP concentrations (50, 100, and 250 mg/L). Fixed-combination groups were also tested using 100 mg/L PE-MP with each DEL concentration. Embryo viability remained high across all groups, ranging from 88% to 98% (see also Table S5 in the Supplementary Material for details.) Malformations were observed in 29% of embryos treated with DEL alone and in 33% of embryos exposed to mixtures. Significant growth inhibition occurred only in the 13.6 µg/L DEL group (p < 0.05), while the corresponding mixture group exhibited a similar but non-significant trend (Fig. 3; see also Table S5 and Figure S6 in the Supplementary Material for details).
Embryos exposed to DEL alone or to the DEL/PE-MP mixtures exhibited a significant increase in GST activity compared to the control group (p < 0.05, Fig. 4; see also Table S6 in the Supplementary Material for details). However, pairwise comparisons showed no significant difference between the single and mixture groups for GST activity. Exposure to DEL alone did not significantly alter CAT activity (p > 0.05). However, exposure to the DEL/PE-MP mixtures or PE-MP alone (50 and 100 mg/L) significantly increased CAT activity. Pairwise tests showed that CAT in the 2.72 µg/L DEL + 100 mg/L PE-MP mixture was significantly higher than in 2.72 µg/L DEL alone, and CAT in the 13.6 µg/L DEL + 100 mg/L PE-MP mixture was higher than both 13.6 µg/L DEL and 100 mg/L PE-MP. Exposure to DEL alone increased AChE activity, and the increase was significant at 13.6 µg/L of DEL, both alone and in combination with PE-MP (p < 0.05). CaE activity increased significantly (p < 0.05) in embryos exposed to 2.72 µg/L DEL alone and in those exposed to the mixture of 2.72 µg/L DEL with PE-MP. Interaction index (II) analysis revealed that the low DEL mixture (2.72 µg/L) produced additive effects on GST and CaE and antagonistic effects on CAT and AChE in embryos. At the higher DEL level (13.6 µg/L), GST responses remained additive, CAT exhibited a synergistic pattern, and AChE showed antagonism (Table 2).
Fig. 3. Malformations (%) and body length in embryos exposed to sublethal DEL and polyethylene microplastics (PE-MPs) alone and as DEL/PE-MP mixtures (96 h, FETAX; see also Table S5). Malformations as defined in Fig. 1. Data are mean ± SE. *p < 0.05 compared with control. Letters denote significant pairwise differences between treatment groups (p < 0.05; t-test or Mann–Whitney U, as applicable)
Fig. 4. Biomarker responses (GST, GR, CAT, CaE, AChE) in embryos (stages 8–11; 96 h exposure) (see also Table S6). Values are mean ± SE of n = 5 biological replicates (each replicate = pool of 15 embryos). *, **, *** indicate p < 0.05, p < 0.01, p < 0.001 compared with control (one-way ANOVA/Dunnett’s or Kruskal–Wallis/Dunn’s). Letters denote significant pairwise differences between treatment groups (p < 0.05; t-test or Mann–Whitney U)
Table 2. Interaction indices (II) for biochemical biomarkers in embryos and tadpolesEmbryosTadpolesDELPE-MPDELPE-MPDELPE-MPDELPE-MP Biomarkers 2.72 µg/L100 mg/L13.6 µg/L100 mg/L2.72 µg/L100 mg/L13.6 µg/L100 mg/L GST 0.97 ^Ad^ 1.00 ^Ad^ 0.96 ^Ad^ 0.90 ^An^
GR –––– CAT 0.92 ^An^ 1.33 ^Si^ 0.74 ^An^ 0.77 ^An^
CaE 0.99 ^Ad^ –0.95 ^Ad^ – AChE 0.82 ^An^ 0.93 ^An^ ––^Ad^: Additive effect, ^Si^: Synergistic effect, ^An^: Antagonistic effect
Similar biomarker trends were observed in tadpoles. DEL alone and DEL/PE-MP mixtures significantly increased GST activity (Fig. 5; see also Table S7 in the Supplementary Material for details). However, CAT activity was elevated only in the groups exposed to DEL alone, which differs from the embryonic pattern. CaE activity increased significantly in tadpoles exposed to 2.72 µg/L DEL (p < 0.05), and pairwise comparisons showed a significant difference between the DEL (2.72 µg/L) + PE-MP (100 mg/L) mixture and the single PE-MP (100 mg/L) group, suggesting that DEL modified the microplastic-induced response. PE-MP alone at 250 mg/L significantly inhibited AChE and GR activity. Tadpole analysis indicated additive interactions for GST and CaE and antagonistic interactions for CAT at the lower DEL concentration (Table 2). At the higher DEL concentration, both GST and CAT exhibited antagonistic responses.
Fig. 5. Biomarker responses (GST, GR, CAT, CaE, AChE) in tadpoles (stage 46; 96 h exposure) (see also Table S7). Values are mean ± SE of n = 5 biological replicates (each replicate = pool of 15 tadpoles). *, **, *** indicate p < 0.05, p < 0.01, p < 0.001 compared with control (one-way ANOVA/Dunnett’s or Kruskal–Wallis/Dunn’s). Letters denote significant pairwise differences between treatment groups (p < 0.05; t-test or Mann–Whitney U)
Discussion
The primary aim of this study was to evaluate the lethality and teratogenicity of DEL and PE-MPs in X. laevis embryos individually, and to assess their combined effects in the second phase. The tested PE-MP concentrations did not cause significant increases in mortality or malformations compared to the control group. This observation is consistent with their relatively inert morphology and size distribution, as characterized in this study. Light microscopy images also showed partial aggregation of PE-MPs in FETAX medium, which may influence their interaction with DEL and its apparent bioavailability (Supplementary Figure S4). This finding is consistent with previous research showing no lethality or phenotypic changes in X. laevis embryos exposed to microplastics at concentrations of up to 100 mg/L in FETAX assays (Bonfanti et al. 2021).
It is worth noting that the PE-MP concentrations used in this study (50–250 mg/L) exceed typical environmental levels, which are usually within the µg/L to low mg/L range. However, these levels were selected to simulate worst-case exposure conditions, as is common in laboratory ecotoxicology, and to reveal potential biochemical and developmental thresholds that could manifest under chronic or hotspot contamination scenarios. Accordingly, these concentrations are intended for mechanistic assessment and should not be interpreted as environmentally representative exposures.
Embryos exposed to DEL exhibited no lethality during the initial 48 h. However, LC_50_ values decreased significantly at 72 and 96 h (2,035 and 68 µg/L, respectively), indicating time-dependent toxicity associated with critical developmental stages, such as organogenesis and neurophysiological activation. This aligns with reports of pyrethroid-induced neurodevelopmental disruption (Andersen et al. 2022). Reported 96 h LC₅₀ values for DEL vary widely across species and life stages. X. laevis tadpoles exhibit values of 190 µg/L (pure) and 6.26 µg/L (commercial formulation) (Aydin-Sinan et al. 2012; Channing 1998), Bufo arenarum tadpoles exhibit values of approximately 4.4–4.5 µg/L (Salibián 1992), and juvenile fish as low as 1.5–1.9 µg/L (Srivastav et al. 1997; Vijayavel and Balasubramanian 2007). The intermediate LC_50_ observed in the present study in embryos underscores the influence of exposure conditions and formulations on toxicity and the sensitivity of different developmental stages.
The FETAX assay has been widely used to evaluate the toxic and teratogenic effects of environmental pollutants on amphibian embryos (Arancio et al. 2019; Cardoso-Vera et al. 2017; Menegola et al. 2024). This assay can quantify the teratogenicity index (TI), where values ≥ 1.5 indicate teratogenic potential according to ASTM standards. In this study, DEL was classified as teratogenic with a TI of 8.97, whereas PE-MP was not. Embryos exposed to DEL exhibited three main malformations: tail curvature, intestinal abnormalities, and stunted growth. Similar malformations have been reported in fish embryos exposed to DEL, indicating conserved developmental effects across vertebrate species (Kuder and Gundala 2018). Tail malformations in X. laevis embryos exposed to various pesticides have been linked to muscle overload and prolonged contractions (Yu et al. 2013).
Biochemically, exposure to DEL alone or in combination with PE-MP increased AChE activity in embryos. The lack of AChE inhibition is expected because AChE is not a direct target of DEL (Ray and Fry 2006). Pyrethroids act by binding to voltage-gated sodium channels, which causes prolonged nerve stimulation and neurotoxicity rather than directly inhibiting AChE (He et al. 2008). CaE activity significantly increased in embryos exposed to a low DEL/PE-MP mixture and in tadpoles exposed to DEL alone or in combination with PE-MP. CaEs play a role in detoxifying ester-containing pesticides, such as pyrethroids, and are sensitive biomarkers of environmental pesticide exposure (Wheelock et al. 2005; Yang et al. 2020). The increased CaE activity likely reflects phase I biotransformation and detoxification processes (Satoh and Hosokawa 2006).
GST, a key phase II detoxification enzyme and a widely used biomarker of xenobiotic exposure, was elevated in embryos and tadpoles following exposure to DEL alone or DEL/PE-MP mixtures (Anila et al. 2021; Lajmanovich et al. 2019; Pimpão et al. 2007; Tierbach et al. 2018). GST induction may represent an adaptive response that enables organisms to counteract oxidative and chemical stress (Liu et al. 2006).
CAT activity increased in embryos exposed to DEL/PE-MP mixtures or PE-MP alone; however, in tadpoles, CAT elevation occurred only with DEL exposure. DEL toxicity is often associated with oxidative stress due to the generation of reactive oxygen and nitrogen species, which disrupt metabolism and cellular defenses (Hafsi et al. 2024; Lu et al. 2019). The elevated GST and CAT activities, combined with the observed malformations, suggest that considerable oxidative stress may contribute to teratogenic effects. Differences in antioxidant enzyme responses between embryos and tadpoles may be explained by developmental changes in glutathione metabolism.
During early embryogenesis of X. laevis, antioxidant defense primarily depends on catalase (CAT) and superoxide dismutase (SOD) activities, while the glutathione-dependent system (including GST and GR) becomes more prominent after stage 35/36 as differentiation progresses (Rizzo et al. 2007). Accordingly, the moderate increase of phase I (CaE) and phase II (GST) enzyme activities observed in tadpoles compared with embryos may reflect the higher metabolic capacity and detoxification potential that develops during post-embryonic stages of X. laevis, rather than a fully adaptive compensatory response (Angelucci et al. 2002; Menon and Rozman 2007; Prokić et al. 2019). This stage-dependent antioxidant shift may involve transcriptional control of redox-responsive genes. In X. laevis, activation of the Nrf2 transcription factor under oxidative stress up-regulates glutathione S-transferase (GST) and aldo-keto reductase (AKR) isoforms, indicating that Nrf2-mediated regulation contributes to enhanced detoxification capacity during post-embryonic development (Malik and Storey 2009).
Only the highest PE-MP concentration (250 mg/L) significantly inhibited GR and AChE activities in tadpoles. AChE inhibition leads to acetylcholine accumulation, causing neurotoxicity and potentially lethality (Kaushal et al. 2021; Storck et al. 2025). GR maintains the critical glutathione redox balance for managing oxidative stress (van der Oost et al. 2003). Thus, inhibition of these enzymes reflects the sublethal toxic effects of PE-MP.
Analysis of the sublethal effects of DEL and PE-MP mixtures revealed mostly additive or antagonistic interactions across biomarkers. However, pairwise comparisons indicated that CAT activity in embryos exposed to DEL + PE-MP mixtures was significantly higher than in either single exposure, supporting an endpoint-specific synergistic pattern in CAT activity. Similarly, in tadpoles, a significant pairwise difference in CaE activity between the DEL (2.72 µg/L) + PE-MP (100 mg/L) mixture and the single PE-MP (100 mg/L) group suggested that DEL modified the microplastic-induced detoxification response. These results suggest that microplastics may modify the apparent toxicity of hydrophobic compounds like DEL, particularly through oxidative and detoxification-related pathways.
Similar mixture-dependent effects have been described in recent studies involving microplastics and other pollutants. For instance, Ye et al. (2025) demonstrated that co-exposure to polyethylene microplastics and bisphenol A induced synergistic endocrine disruption and cellular toxicity in zebrafish and MLTC-1 cell models, while Kim et al. (2024) reported developmental abnormalities and oxidative stress in zebrafish embryos exposed to mixtures of microplastics and plastic additives. Together with our data, these studies indicate that microplastics can modulate DEL bioavailability and redox responses, highlighting their capacity to alter oxidative and detoxification mechanisms in aquatic vertebrates.
Although the present experimental design ensured adequate power to detect medium-to-large effects, it may have been insufficient to capture subtle biochemical changes, particularly for variable biomarkers such as GR, CAT, and AChE. Embryonic assays showed slightly higher variability (CV up to ~ 26%) than tadpole assays, which may have increased the likelihood of Type II errors at low concentrations. Therefore, non-significant outcomes at the lowest doses should be interpreted with caution and supported by trends and effect-size considerations rather than p-values alone. In addition, higher-resolution imaging and more advanced microplastic characterization (e.g., polymer verification and surface/morphology analyses) would strengthen future studies.
These results highlight the importance of considering life stage and chemical combinations when assessing environmental risk, as mixture interactions can mask early cellular damage and lead to delayed toxicity (Burić et al. 2023). Combined exposure to MPs and pesticides has been shown to impair metamorphosis, reduce growth and fecundity, and alter behavioral responses in amphibians and fish (Rahman et al. 2024; Relyea 2009; Subaramaniyam et al. 2023). Such sublethal effects, even in the absence of acute mortality, may threaten amphibian population stability and disrupt freshwater ecosystem balance (Mann et al. 2009; Peluso et al. 2024).
Conclusions
The PE-MP concentrations tested in this study (50–250 mg/L in mixtures; 50–1,000 mg/L in single exposures) did not induce significant lethality, malformations, or developmental delays in X. laevis embryos, indicating no observable acute developmental effects under the tested conditions. In contrast, even low concentrations of DEL caused significant embryotoxic and teratogenic effects, supporting its potency as a developmental toxicant. Sublethal DEL exposure also modulated the activities of several enzymes involved in the oxidative stress response and detoxification, such as GST and CAT. Distinct biochemical patterns were observed between embryonic and larval stages. Mixture effects were predominantly additive or antagonistic, with synergy limited to specific endpoints (e.g., CAT in embryos). Our results underscore the importance of considering pollutant mixtures in ecotoxicological risk assessments, as microplastics can modify the effects of co-occurring toxicants in ways that may be biologically relevant depending on life stage and endpoint. Given the growing likelihood of pesticide–microplastic co-occurrence in natural waters, such interactions warrant further investigation to better inform ecological risk assessments.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1ASTM (2003) American Society for Testing and Materials, Standard guide for conducting the Frog Embryo Teragonesis Assay-Xenopus (FETAX), E 1439-98. In: ASTM Standards on Biological Effects and Environmental Fate. Vol. 11.05. Philadelphia, PA, 447–457
- 2OECD (2019) Test Guideline No. 248: Xenopus Eleutheroembryo Thyroid Assay (XETA), Sect. 2: Effects on Biotic Systems. OECD Publishing
- 3Plastics Europe (2023) Plastics Europe launches Plastics – the fast Facts 2023: https://plasticseurope.org/media/plastics-europe-launches-the-plastics-the-fast-facts-2023/
- 4Ren Q, Zhang T, Li S et al (2016) Integrative characterization of toxic response of zebra fish (Danio rerio) to deltamethrin based on A Ch E activity and behavior strength. Bio Med Res Int 2016:7309184 10.1155/2016/7309184
- 5Storck TR, Ames J, Qualhato G et al (2025) Differential biochemical responses of Cyprinus Carpio after dietary and waterborne exposure to microplastics from polyethylene-based biodegradable and conventional bags. 10.1007/s 10646-025-02937-1. Ecotoxicology