Monitoring of Cd and GSH contents and Bn-OASTL expression in transgenic tobacco seedlings in response to Cd stress
Xiaolan He, Jianwei Wang, Wenxu Li, Xinhong Chen, Naser Anjum, Debasis Chakrabarty, Debasis Chakrabarty

TL;DR
Researchers studied a gene from oilseed rape in tobacco plants to see if it helps with cadmium stress, but found it doesn't improve tolerance or accumulation.
Contribution
The study isolates and expresses a novel OASTL gene from Brassica napus in tobacco to investigate its role in cadmium stress response.
Findings
BnaOASTL gene expression is upregulated in transgenic tobacco under cadmium stress.
Overexpression of BnaOASTL does not enhance cadmium tolerance or accumulation in tobacco.
Glutathione levels remain unchanged in transgenic plants compared to wild-type.
Abstract
Cadmium (Cd) pollution threatens agricultural productivity and food safety. O-acetylserine(thiol)lyase (OASTL) genes have been tied to plant responses to heavy metal stress, yet their roles in heterologous systems, particularly in Cd accumulation and tolerance, remain unclear. Here, we isolated a novel OASTL gene, BnaOASTL, from the high-Cd-accumulating oilseed rape cultivar Brassica napus “Nanyou 868” and expressed it in tobacco (Nicotiana benthamiana). Transgenic lines were exposed to Cd stress, and Cd content, glutathione (GSH) level, and BnaOASTL expression were evaluated. The full-length BnaOASTL cDNA (969 bp) encoded a cytoplasmic/nuclear protein of 322 amino acids. Under Cd stress, Bn-OASTL expression was significantly upregulated in transgenic plants, particularly in roots. However, compared with wild-type, transgenic lines showed no improvement in Cd tolerance or accumulation…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1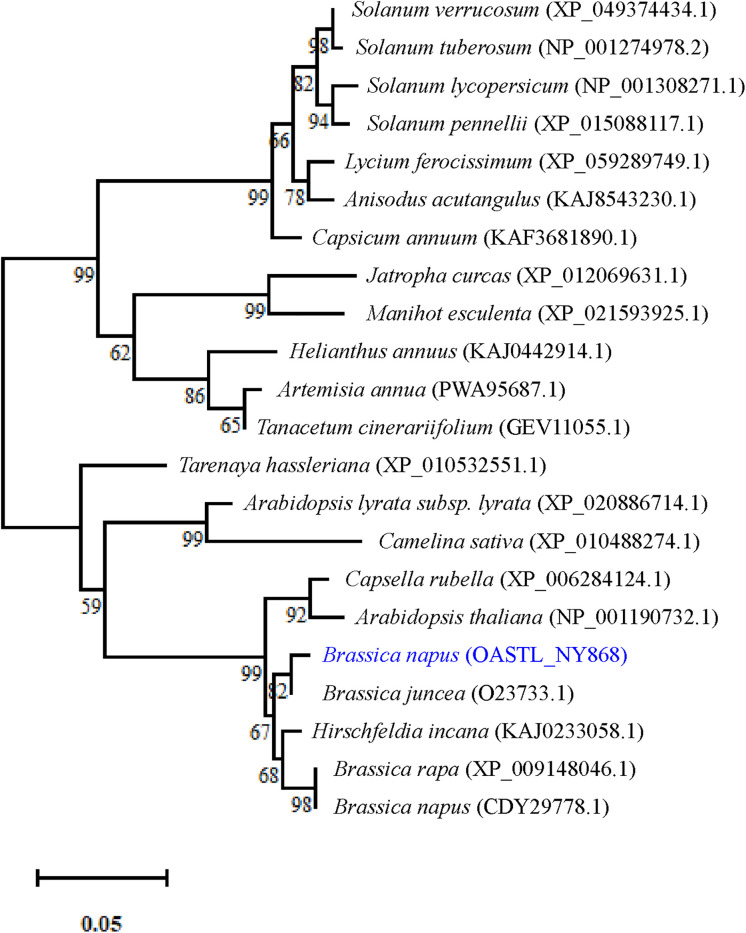 Fig 2
Fig 2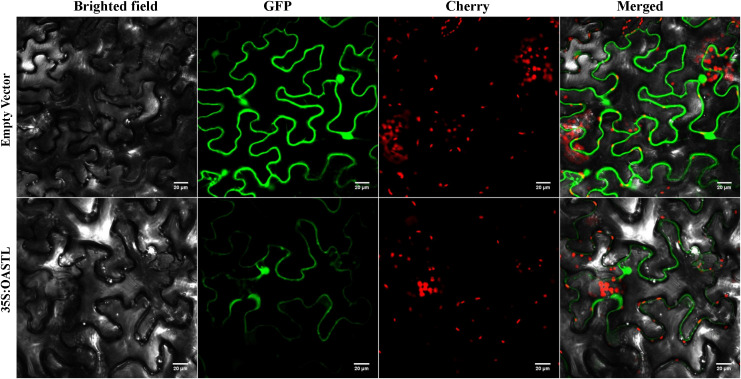 Fig 3
Fig 3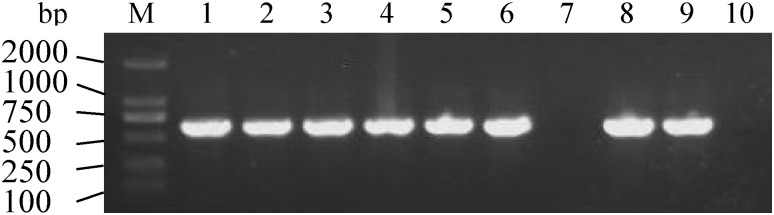 Fig 4
Fig 4 Fig 5
Fig 5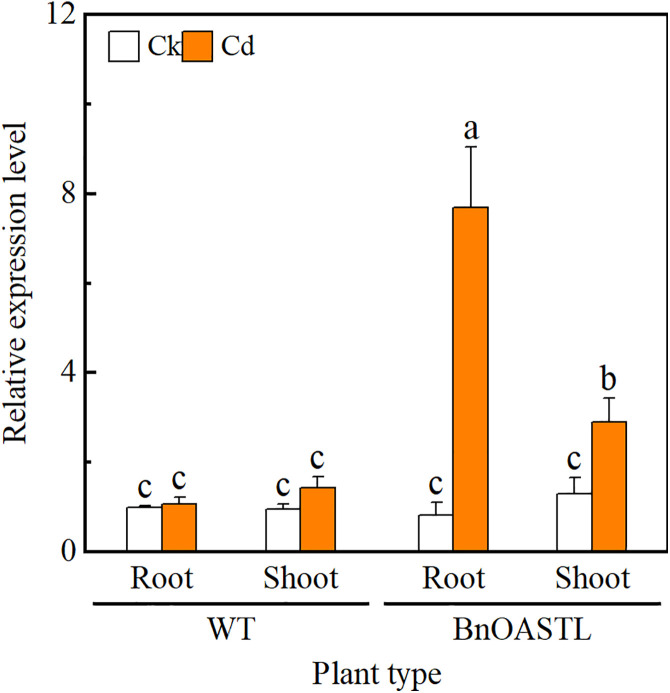 Fig 6
Fig 6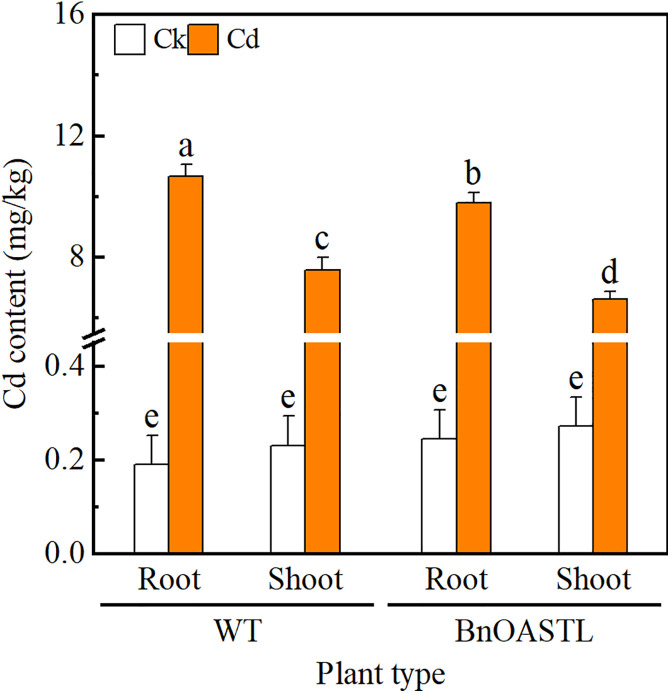 Fig 7
Fig 7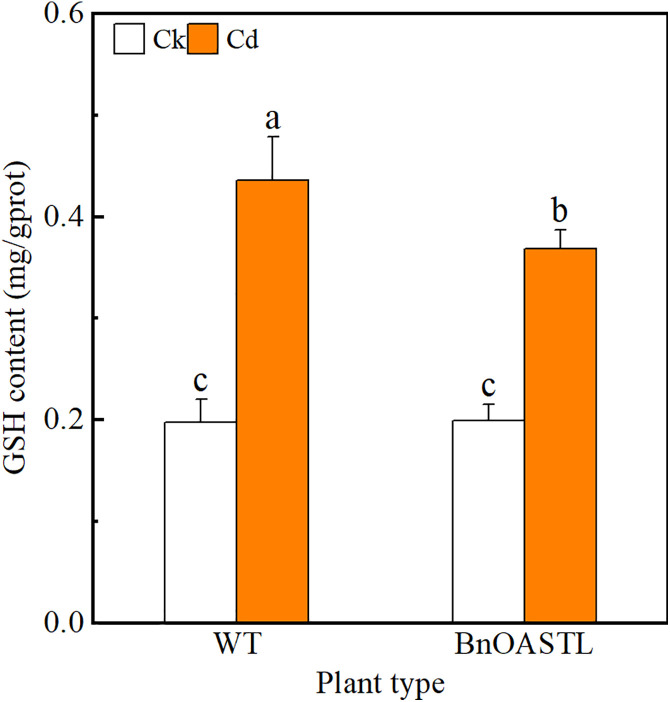 Fig 8
Fig 8- —Doctoral Development of Kaili University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Photosynthetic Processes and Mechanisms · Plant Gene Expression Analysis
1. Introduction
Cadmium (Cd) contamination in farmland soils is a pressing environmental problem, threatening crop production and human health due to its toxicity and high mobility in plants [1,2]. Mitigation strategies such as phytoremediation and the development of Cd-safe crops depend on understanding the molecular pathways governing Cd uptake, transport, and detoxification.
The O-acetylserine(thiol)lyase (OASTL) family is central to cysteine (Cys) biosynthesis, which underpins production of glutathione (GSH), a key metabolite in heavy metal detoxification [3–5]. In Arabidopsis thaliana, depletion of AtOASTL-A1 reduces intracellular Cys and GSH contents, increasing Cd sensitivity [6]. Since the first OASTL was identified in A. thaliana [7–10], homologs have been isolated from spinach [11], Brassica juncea L. [12], vetch (Vicia sativa L.) [13], Glycine max (L.) [4], Leucaena leucocephala [14], Sorghum bicolor [3], Solanum lycopersicum L. [15], and Cardamine hupingshanensis [16]. Functional analyses have confirmed that some OASTL genes enhance heavy metal resistance when overexpressed in transgenic plants [4].
OASTL exists in multiple isoforms localized to the cytoplasm, mitochondria, and chloroplasts [5,17]. These proteins are highly conserved across species, particularly in the diphosphate (PLP)-binding site (PXXSVKDR), substrate-binding site (TSGNT), and serine acetyltransferase (SAT) interaction site (KPGPHK) [15,18–20], underscoring their essential metabolic role. Their involvement in Cd stress is well documented. For example, overexpression of GmOASTL4 in tobacco enhances Cd tolerance [4]. Conversely, disruption of AtOASTL-A1, a key factor in the final step in Cys biosynthesis [9], reduces GSH levels, increases oxidative stress, and heightens Cd sensitivity [5,6,21], highlighting the link between OASTL activity and Cd detoxification.
Despite extensive studies in other species, OASTL genes from high-Cd-accumulating Brassica napus genotypes remain poorly characterized. Recent work examined OASTL in selenium metabolism in Cardamine hupingshanensis [16] and in tomato under heavy metal stress [3], but the function of OASTL from Cd-accumulating rapeseed cultivars and its role in heterologous systems has not been thoroughly investigated. It is not yet known whether overexpression can confer Cd tolerance or alter accumulation patterns in a heterologous system. Here, we cloned the BnaOASTL gene from the high-Cd-accumulating oilseed rape cultivar “Nanyou 868” and evaluated its role in Cd stress response through heterologous expression in tobacco. This work provides functional insight into BnaOASTL in a heterologous context and contributes to understanding the molecular basis of Cd response mechanisms, with potential implications for developing Cd-tolerant crops via molecular breeding.
2. Materials and methods
2.1. Plants and treatment
The high Cd-accumulating B. napus cultivar ‘Nanyou 868’ (2n = 4x = 38, AACC) [22] was used as the gene source. Nicotiana benthamiana was selected for transgenic plant (TP) development. Surface-sterilized seeds were placed on half-strength Murashige and Skoog (MS) medium for germination at 25°C under a 12 h/12 h light/dark cycle in a greenhouse. For Cd treatment, five-leaf-stage seedlings were exposed to 5 mg·L^−1^ CdCl_2_·5H_2_O for 72 h, after which leaves were harvested, snap-frozen in liquid nitrogen, and kept at −80°C.
2.2. Cloning of BnaOASTL cDNA
Total RNA was extracted using the RNeasy Plant Mini Kit (Qiagen, Germany, Cat. No. 74904) and converted into cDNA using SuperScript III Reverse Transcriptase (Invitrogen USA, Cat. No. 18080093). Two OASTL fragments were amplified with primers designed specifically for OASTL sequence (accession no. GQ996586.1), namely, N6-OASTL-1 (DPsen): cagtGGTCTCacaacatggcatctcgaattgctaaag, N6-OASTL-1 (DPantisen): cagtGGTCTCaattccatacagcttaacgttag; N6-OASTL-2 (DPsen): cagtGGTCTCagaatggagccaattgaaagtg, and N6-OASTL-2 (DPantisen): cagtGGTCTCatacaagcctggaaggtcattgattc using KOD FX Neo (Toyobo, Japan, Cat. No. KFX-201) on a T100 Thermal Cycler (Bio-Rad, USA). Each reaction contained 1.0 µL cDNA (50 ng), 1.0 µL each primer (10 µM), 5 μL buffer, 10 µL dNTP mix, 1 µL KOD, and 31 µL ddH₂O. Amplification conditions were 94°C for 5 min; 30 cycles of 30 s at 94°C, 45 s at 55°C, and 58 s at 72°C, and 10 min at 72°C. The products were purified and cloned into a custom vector via the “Golden Gate” method and subsequently sequenced [23].
2.3. Identification and characterization of BnaOASTL
The open reading frame (ORF) of BnaOASTL was identified using NCBI ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder/). Subcellular localization was predicted with PSORT (http://psort.ims.u-tokyo.ac.jp/). The isoelectric point (pI) and molecular weight (MW), and other physicochemical properties were obtained using the Expert Protein Analysis System (ExPASy) (http://cn.expasy.org/). Multiple sequences were aligned using ClustalX and visualized in GenDoc. A phylogenetic tree was generated in MEGA7.0 utilizing the neighbor-joining (NJ) method.
2.4. Vector construction and tobacco transformation
The BnaOASTL ORF was cloned into vector pBWA(V)HS-ccdb-osgfp (BioRun, Wuhan, China) via Golden Gate assembly (Fig 1). Sequence-verified plasmids were introduced into Agrobacterium tumefaciens strain EHA105. Transgenic tobacco plants were established via the leaf-disc method and screened on MS medium with 20 mg/L hygromycin B.
Schematic of the recombinant vector.RB and LB, right and left T-DNA borders; 35S, CaMV 35S poly A; HYG, hygromycin B; osgfp, the BnaOASTL-GFP fusion gene; Tnos, nopaline synthase terminator.
2.5. Subcellular localization
pBWA(V)HS-BnaOASTL-osgfp and empty control vectors were introduced into A. tumefaciens strain GV3101 and transiently expressed in N. benthamiana leaves via agroinfiltration. GFP signals were observed 48 h post-infiltration under a confocal microscope (OLYMPUS IX71, Japan) at 488 nm excitation and 510 nm emission.
2.6. Molecular characterization of transgenic plants
DNA from T_3_ transgenic and control plants (untransformed and vector control) was extracted by the CTAB method and screened for BnaOASTL transgene using PCR with primers FP (5′-ttcatttggagagaacacgggggac-3′) and RP (5′-gttctcaaactgttggagcatg-3′) in triplicate with a 58 °C annealing temperature. Products were resolved on a 1.0% agarose gel, yielding a ~ 500 bp fragment from the positive plants.
2.7. Cd tolerance in transgenic plants
Cd stress was applied following [24] with slight modifications. T_3_ transgenic and wild-type (WT) tobacco seeds were germinated on 1/2 MS medium at 25°C with a 16 h/8 h light/ dark cycle for 10 days. Five-leaf-stage T_3_ seedlings were transferred to 9-cm pots containing perlite and vermiculite (1:1) and treated with 5 mg· kg^−1^ Cd (CdCl_2_·2.5 H_2_O) in basal nutrient solution (pH 5.5) or 400 ml distilled water. Each treatment included three replicates, with one plant per pot, and seedling growth was monitored for one week.
2.8. Measurement of Cd and GCH content
After 20 days of Cd exposure, roots were immersed in 20 mmol·L^−1^ Na_2_-EDTA for 30 min, then washed three times with deionized water to remove surface-bound Cd. Samples were then separated into roots and shoots. Half of each sample was immediately frozen in liquid nitrogen for fresh tissue analysis, while the remainder was oven-dried at 75°C for Cd determination. Cd content was measured following the Chinese National Standard HJ786–2016 in China [25,26]. Dried roots and shoots were weighed, pulverized, decomposed at 550°C for 8 h, and digested in 30% HNO_3_. Cd concentrations were measured using flame atomic absorption spectrometry (Shimadzu AA-6300, Japan). Data represent the mean of three replicates. GSH was extracted from shoots of Cd-treated and control plants and quantified using a reduced glutathione (GSH) kit (Visible light spectrophotometer, Shanghai Jinghua 721).
2.9. Quantitative real-time PCR (qRT-PCR)
qRT-PCR was executed on a StepOnePLUS Real-Time PCR system (Applied Biosystems, USA) using 2 × SG Green qPCR Mix with ROX (SinoGene, China, Cat. No. SG011) using primers QRT-BnaOASTL-F (5’-ACCCTGCCAACCCAAAGATA-3’) and QRT-BnaOASTL-R (5’-ACCACCAGTACCAATCCCAG-3’). Actin served as the internal control and amplified using Actin-F (5’-tttcctggcattgcagatcg-3’) and Actin-R (5’-tccagacactgtacttgcgt-3’). Reactions (20 µL) included 2 µL cDNA, 0.4 µL of each primer (10 µM), 10 µL 2 × SG Green Mix, and 7.2 µL ddH₂O. Cycling was run at 95°C for 3 min, then 40 cycles of 95°C for 10 s and 60°C for 30 s. Relative expression was quantified using the 2^-ΔΔCt^ method with three biological and three technical replicates.
2.10. Statistical analysis
All results are shown as mean ± standard deviation (SD) ≥3 independent replicates. Data were compared using one-way ANOVA with Tukey’s HSD post hoc test (SPSS 22.0, IBM, USA), with P < 0.05 considered significant.
3. Results
3.1. Sequence and phylogenetic analysis of BnaOASTL
Gene structure analysis revealed that the BnaOASTL ORF is 969 bp long and encodes a 322-amino acid protein (S1 Fig). The mature protein has an estimated molecular weight of 33.9 kDa, a calculated pI of 5.50, and three conserved domains: TSGNT (substrate-binding site), KPGPHK (SAT1-binding site), and PXXSVKDR (PLP-binding site) (S2 Fig). Alignment with cysteine synthases from Brassica rapa, Raphanus sativus, Hirschfeldia incana, and Eutrema salsugineum showed amino acid sequence identities of 99.69%, 98.45%, 98.14%, and 97.52%, respectively. Phylogenetic analysis of the BnaOASTL protein with other 21 representative species (Fig 2) divided the 22 OASTL proteins into two subcategories. Subcategory I includes XP_049374434.1, NP_001274978.2, NP_001308271.1, XP_015088117.1, XP_059289749.1, KAJ8543230.1, KAF3681890.1, XP_012069631.1, XP_021593925.1, KAJ0233058.1, PWA95687.1 and GEV11055.1. Subcategory II includes XP_010532551.1, XP_020886714.1, XP_010488274.1, XP_006284124.1, NP_001190732.1, OASTL_NY868, O23733.1, KAJ0442914.1, XP_009148046.1 and CDY29778.1, with OASTL_NY868 closely related to O23733.1. Subcellular localization analysis indicated that BnaOASTL is cytoplasmic and nuclear, lacks transmembrane domains, and belongs to the PLP-dependent, β-substituted alanine synthase superfamily.
Phylogenetic tree of OASTL proteins from B. napus and other species based on amino acid sequences.Bootstrap values (>50%) from 1000 replicates are demonstrated at branch nodes. The scale bar represents a distance of 0.05. The B. napus OASTL sequence is marked with a blue symbol. Other sequences were obtained from XP_049374434.1, NP_001274978.2, NP_001308271.1, XP_015088117.1, XP_059289749.1, KAJ8543230.1, KAF3681890.1, PWA95687.1, GEV11055.1, KAJ0442914.1, XP_012069631.1, XP_021593925.1, OASTL_NY868, O23733.1, XP_009148046.1, CDY29778.1, KAJ0233058.1, XP_006284124.1, NP_001190732.1, XP_020886714.1, XP_010488274.1 and XP_010532551.1.
3.2. Subcellular localization of BnaOASTL
BnaOASTL-GFP was predominantly localized to the cytoplasm and nucleus (Fig 3), suggesting that BnaOASTL likely performs its functions in these subcellular compartments.
Subcellular localization of BnaOASTL-GFP in tobacco epidermal cells.Scale bar = 10 µm.
3.3. Molecular characterization of transgenic lines
Eight independent transgenic tobacco lines expressing the BnaOASTL gene were generated. Plants transformed with empty vector pBWA(V)HS-osgfp served as controls. Integration of the BnaOASTL in T3 plants was validated by PCR using gene-specific primers (Fig 4).
Confirmation of BnaOASTL integration in tobacco lines by PCR, showing amplified BnaOASTL bands from 10 tobacco lines.M, DL2000 DNA marker; 1–6 and 8: transgenic lines; 9, positive control (plasmid); and 10, negative control (WT).
3.4. Phenotypic response to Cd stress
Tobacco plants were treated with 5 mg· kg^−1^ Cd (CdCl_2_·2.5 H_2_O). After 7 days, transgenic and WT plants showed similar phenotypes under Cd stress, except for the slower growth of WT (Fig 5).
Images of WT and transgenic plants before (a–f) and after (g–l) cadmium treatment.
Panels a and g represent WT plants, and panels b–f and h–l represent transgenic lines.
3.5. BnaOASTL expression under Cd stress
qRT-PCR results (Fig 6) indicated that Cd stress significantly elevates BnaOASTL transcript abundance in roots and shoots, with levels in roots substantially exceeding those in shoots.
BnaOASTL expression by qRT-PCR in roots and leaves of seedlings after 24 h treatment with 0 or 5 mg· kg−1 Cd.Data are shown as means ± SD (n = 3) with different letters indicating significant differences (P < 0.05, one-way ANOVA, Tukey’s test).
3.6. Cd and GSH contents
The expression of BnaOASTL in transgenic lines was further investigated by analyzing Cd contents in T3 transgenic and WT tobacco plants before and during Cd stress (Fig 7). Under both control and stress conditions, Cd levels were similar between BnaOASTL overexpression lines and wild-type plants. Similarly, GSH contents showed no significant differences between transgenic lines and WT under either normal or Cd-stress conditions (Fig 8).
Cadmium accumulation in shoots and roots of transgenic (T1-T4) and WT plants under 0 or 5 mg· kg−1 Cd (CdCl2·2.5 H2O) treatment.Values represent mean ± SD (n = 4), with different letters indicating significant differences (P < 0.05).
Glutathione content in shoots.Data are mean ± SD; no significant differences were found.
4. Discussion
4.1. Sequence and structure analysis of BnaOASTL
Different OASTL isoenzymes perform diverse functions [16], and several family members contribute to plant responses to heavy metals and oxidative stress [5]. Recently, OASTLs have gained attention for their role in Cd stress responses [3]. In this study, a putative OASTL gene, designated BnaOASTL, was isolated from the high Cd-accumulating B. napus L. (‘Nanyou 868’). Sequence analysis unveiled that BnaOASTL cDNA contains a 969 bp ORF encoding a 322-amino acid protein with a predicted MW of 33.9 kDa and a PI of 5.50. These features are consistent with OASTL proteins from Arabidopsis, Sorghum, and Cardamine hupingshanensis, which typically range from 305 to 433 amino acids and exhibit acidic values (<7) characteristic of cytoplasmic OASTLs [3,16]. The deduced BnaOASTL protein contains three conserved domains, i.e., TSGNT (substrate-binding site), KPGPHK (SAT1-binding site), and PXXSVKDR (PLP-binding site), indicating it likely uses pyridoxal monohydrate 5’-phosphate as a cofactor and belongs to the PLP-dependent β-substituted alanine synthase superfamily. The subcellular localization analysis predicts that BnaOASTL may exist in the cytoplasm, chloroplast, and mitochondrion. Confocal imaging of BnaOASTL-GFP in tobacco confirmed predominant localization in the cytoplasm and nucleus, consistent with previous reports highlighting cytoplasmic OASTL function [5,16]. These findings indicate that BnsOASTL belongs to the OASTLA1 type. Multiple sequence alignment unveiled highly conserved C-termini but more variable N-termini among OASTL proteins, suggesting both conserved function and potential for regulatory diversity. Phylogenetic analysis showed that BnaOASTL is most related to OASTL from B. cretica and B. rapa, confirming it as a genuine member of the OASTL family and representing a novel variant within this group.
4.2. Expression of BnaOASTL in transgenic lines in response to Cd
Morphological and physiological responses are key indicators of metal toxicity. To assess the functional role of BnaOASTL in Cd stress, transgenic tobacco overexpressing BnaOASTL was generated. Under Cd stress, no significant phenotypic differences, such as chlorosis or leaf abscission, were observed between transgenic and wild-type seedlings, suggesting that tobacco inherently tolerates Cd at the applied concentration. qRT-PCR unveiled that BnaOASTL expression was dramatically induced in roots but only slightly in shoots of transgenic plants under Cd stress, consistent with patterns observed in other species [3,4]. This upregulation indicates that BnaOASTL is responsive to Cd stress and may participate in Cd-responsive pathways. However, despite the elevated transcript levels, transgenic plants did not exhibit enhanced Cd tolerance or accumulation compared to WT. Moreover, glutathione (GSH) content remained unchanged. These results suggest that overexpression of BnaOASTL alone is insufficient to alter Cd detoxification or accumulation in tobacco. This lack of phenotypic effect may be explained by several factors: (1) functional redundancy and compartmentalization of OASTL isoforms in different subcellular compartments (cytosol, chloroplasts, mitochondria), limiting changes in overall cysteine or GSH pools [15,18–20]; (2) post-translational regulation of OASTL via interaction with serine acetyltransferase (SAT) and feedback inhibition, which may prevent increased cysteine production without concomitant pathway activation; (3) species-specific context, as tobacco may lack regulatory elements or interacting partners present in B. napus, and (4) the complex of Cd detoxification networks, including phytochelatin synthesis, metal transporters, and the antioxidant systems [27], which cannot be significantly altered by overexpression of a single gene. These findings highlight the complexity of Cd stress responses and suggest that BnaOASTL likely functions within a broader metabolic and regulatory network rather than acting in isolation. However, these hypothesis needs a more in-depth study.
5. Conclusions
In summary, this study provides functional characterization of BnaOASTL from a high-Cd-accumulating rapeseed cultivar in a heterologous system. Although its overexpression in tobacco did not enhance Cd tolerance or accumulation, the gene’s response to Cd stress underscores its involvement in Cd-responsive pathways. These findings highlight the complexity of Cd stress responses and suggest that BnaOASTL may function within a broader regulatory network.
Supporting information
S1 FigThe complete cDNA sequence of BnaOASTL and its deduced protein sequence, with amino acids shown in one-letter code below the nucleotides and the asterisk denoting stop codon.(TIF)
S2 FigSequence alignment of BnaOASTL with OASTL proteins from various species.Identical residues are marked with asterisks, conserved substitutions with colons, and semiconserved substitutions with periods. The conserved domains: TSGNT (substrate-binding site), KPGPHK (SAT1-binding site), and PXXSVKDR (PLP-binding site), are boxed. The BnaOASTL sequence was obtained from B. napus, and other sequences from XP_018470163.1, ACX70136.1, KAJ0233058.1, KAJ4871408.1, XP_006414628.1, XP_020874249.1, CAA58893.1, XP_006284124.1, and XP_009107806.1.(TIF)
S1 Raw imageRaw gel images.(PDF)
S1 DataRaw data for fig 6.(XLSX)
S2 DataRaw data for fig 7.(XLSX)
S3 DataRaw data for fig 8.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wu L, Yu Y, Hu H, Tao Y, Song P, Li D, et al. New SFT 2-like Vesicle Transport Protein (SFT 2L) Enhances Cadmium Tolerance and Reduces Cadmium Accumulation in Common Wheat Grains. J Agric Food Chem. 2022;70(18):5526–40. doi: 10.1021/acs.jafc.1c 08021 35484643 · doi ↗ · pubmed ↗
- 2Yu W, Deng S, Chen X, Cheng Y, Li Z, Wu J, et al. Pc NRAMP 1 Enhances Cadmium Uptake and Accumulation in Populus × canescens. Int J Mol Sci. 2022;23(14):7593. doi: 10.3390/ijms 23147593 35886940 PMC 9316961 · doi ↗ · pubmed ↗
- 3Akbudak MA, Filiz E, Uylas S. Identification of O-acetylserine(thiol)lyase (OASTL) genes in sorghum (Sorghum bicolor) and gene expression analysis under cadmium stress. Mol Biol Rep. 2019;46(1):343–54. doi: 10.1007/s 11033-018-4477-0 30443823 · doi ↗ · pubmed ↗
- 4Ning H, Zhang C, Yao Y, Yu D. Overexpression of a soybean O-acetylserine (thiol) lyase-encoding gene Gm OASTL 4 in tobacco increases cysteine levels and enhances tolerance to cadmium stress. Biotechnol Lett. 2010;32(4):557–64. doi: 10.1007/s 10529-009-0178-z 19953300 · doi ↗ · pubmed ↗
- 5Wang C, Zheng L, Tang Z, Sun S, Ma JF, Huang X-Y, et al. OASTL-A 1 functions as a cytosolic cysteine synthase and affects arsenic tolerance in rice. J Exp Bot. 2020;71(12):3678–89. doi: 10.1093/jxb/eraa 113 32129444 · doi ↗ · pubmed ↗
- 6Shirzadian-Khorramabad R, Jing H-C, Everts GE, Schippers JHM, Hille J, Dijkwel PP. A mutation in the cytosolic O-acetylserine (thiol) lyase induces a genome-dependent early leaf death phenotype in Arabidopsis. BMC Plant Biol. 2010;10:80. doi: 10.1186/1471-2229-10-80 20429919 PMC 2890954 · doi ↗ · pubmed ↗
- 7Alvarez C, Calo L, Romero LC, García I, Gotor C. An O-acetylserine(thiol)lyase homolog with L-cysteine desulfhydrase activity regulates cysteine homeostasis in Arabidopsis. Plant Physiol. 2010;152(2):656–69. doi: 10.1104/pp.109.147975 19955263 PMC 2815857 · doi ↗ · pubmed ↗
- 8Bermúdez MA, Páez-Ochoa MA, Gotor C, Romero LC. Arabidopsis S-sulfocysteine synthase activity is essential for chloroplast function and long-day light-dependent redox control. Plant Cell. 2010;22(2):403–16. doi: 10.1105/tpc.109.071985 20179139 PMC 2845405 · doi ↗ · pubmed ↗