Spx inhibits expression of the SwrA•DegU master flagellar activator in Bacillus subtilis
Abigail E. Jackson, Stephen G. Olney, Ayushi Mishra, Daniel B. Kearns

TL;DR
The study reveals that Spx limits flagellar gene expression in Bacillus subtilis by repressing key activators, and ClpX protease helps control this by degrading Spx.
Contribution
The novel finding is that Spx represses flagellar gene transcription, and ClpX protease regulates this repression by degrading Spx.
Findings
Spx represses transcription of degU and swrA genes, which are key for flagellar biosynthesis.
ClpX protease degrades Spx, relieving its repression and promoting swarming motility.
Spx limits DegU-mediated positive feedback to prevent excessive flagellar biosynthesis under stress.
Abstract
Flagella are trans-envelope nanomachines expressed from genes organized in a complex regulatory hierarchy governed at the highest level by transcription factors called master activators. The master activator of flagellar biosynthesis in Bacillus subtilis is a hybrid of SwrA•DegU that is required to increase flagellar density to swarm over solid surfaces. Here, we show that the ClpX unfoldase subunit of the ClpP protease is required for swarming motility, and that cells mutated for ClpX fail to swarm due to restricted levels of both SwrA and DegU. Suppressor mutations were found that increased expression of the fla/che operon under SwrA•DegU control, and mutation of the LonA protease elevates the levels of SwrA protein, while mutation of the global transcriptional regulator Spx increases transcription of both the degU and swrA genes. We conclude that ClpX promotes swarming motility via…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
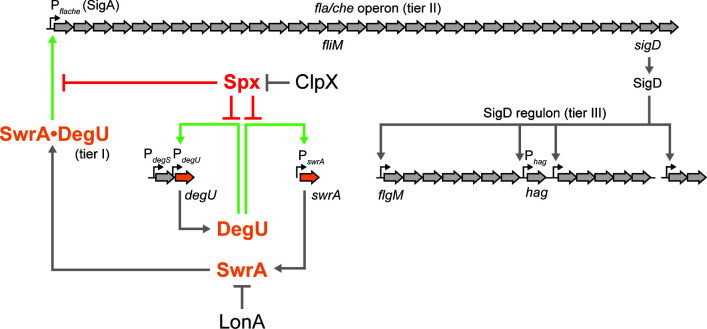 Fig 1
Fig 1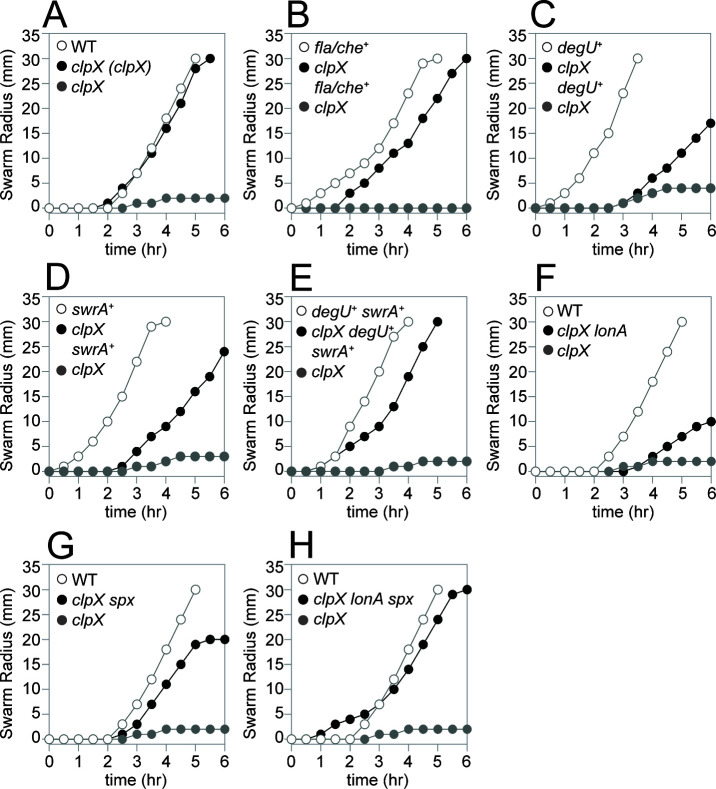 Fig 2
Fig 2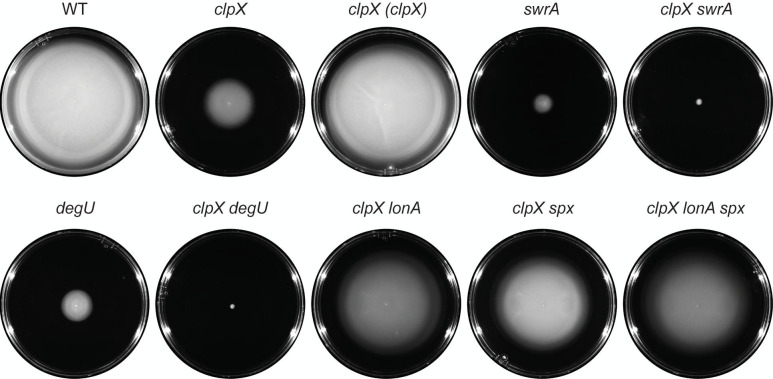 Fig 3
Fig 3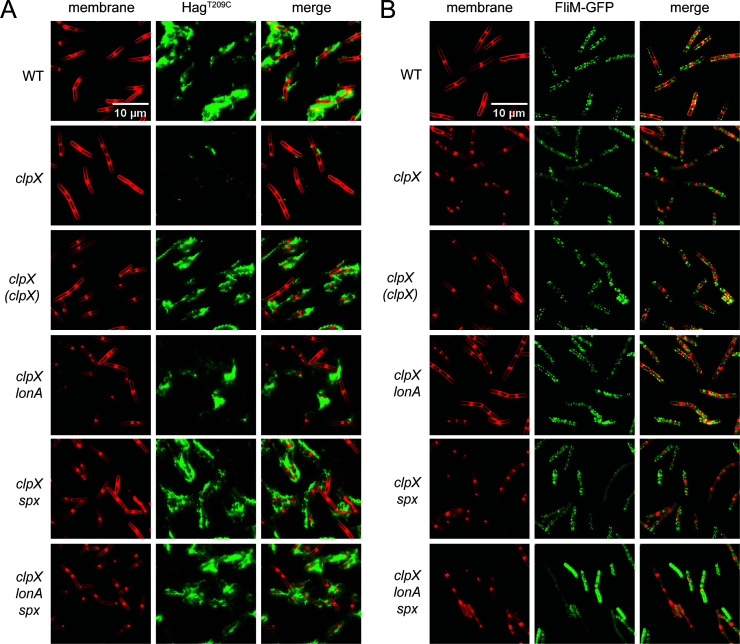 Fig 4
Fig 4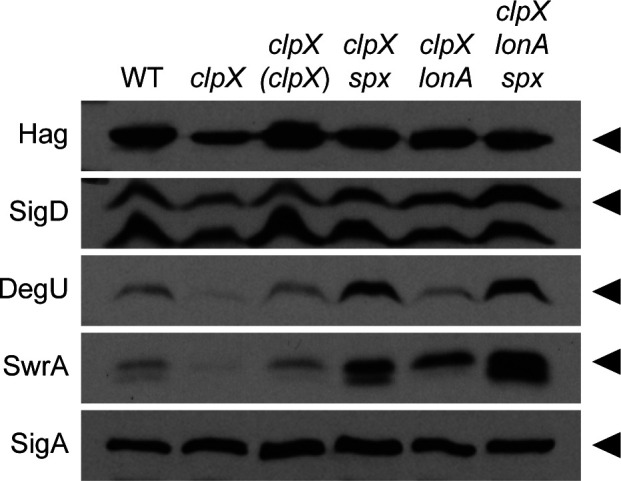 Fig 5
Fig 5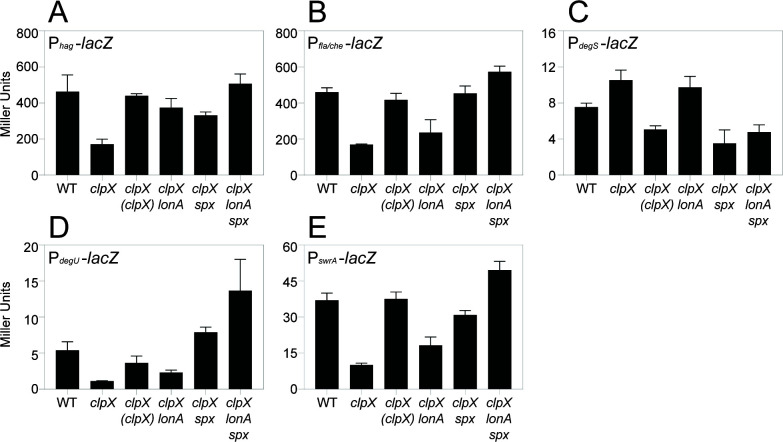 Fig 6
Fig 6| Strain | Genotype |
|---|---|
| NCIB3610 | Wild type |
| DB10 | |
| DB17 | |
| DB20 | |
| DB22 | |
| DB23 | |
| DB24 | |
| DB26 | |
| DB28 | |
| DB53 | |
| DB54 | |
| DB61 | Δ |
| DB62 | |
| DB63 | Δ |
| DB64 | |
| DB67 | |
| DB68 | |
| DB69 | |
| DB70 | |
| DB71 | |
| DB117 | |
| DB470 | |
| DB548 | |
| DB559 | |
| DB644 | |
| DB806 | |
| DB807 | |
| DB808 | |
| DB809 | |
| DB810 | |
| DB811 | |
| DB812 | |
| DB813 | |
| DB847 | |
| DB848 | |
| DB1246 | |
| DB1304 | |
| DB1351 | Δ |
| DB1371 | Δ |
| DB1372 | Δ |
| DB1373 | Δ |
| DB1391 | Δ |
| DB1527 | |
| DB1528 | |
| DB1529 | |
| DB1530 | |
| DB1531 | |
| DB2379 | |
| DB3069 | |
| DB3070 | |
| DB3071 | |
| DB3250 | |
| DK1042 |
|
| DK2399 | |
| DK5457 | |
| DK6563 | |
| DK6613 | |
| DK6647 | |
| DK7584 | |
| DK9386 | |
| DK9387 | |
| DK9388 | |
| DK9389 | |
| DK9390 | |
| DK9391 | |
| DK9392 | |
| DK9393 | |
| DK9447 | |
| DK9529 | |
| DK9597 | |
| DK9609 | |
| DK9618 | |
| DK9744 | |
| DK9746 | |
| DK9748 | |
| DK9749 | |
| DK9750 | |
| DK9751 | |
| DK9753 | |
| DK9842 | |
| DK9843 | |
| DK9865 | |
| DK9925 | |
| DK9934 | |
| DK9981 | Δ |
| DK9984 | Δ |
| DK9985 | Δ |
| DS2231 | |
| DS6239 | Δ |
| DS9446 | Δ |
| Allele | Class | Gene/region | Mutation |
|---|---|---|---|
|
| Ia | P | TA |
|
| Ib | Upstream of P |
|
|
| II |
| AAAAAAA (7) → AAAAAA (6) |
|
| II |
| AAAAAAA (7) → AAAAAAAA (8) |
|
| II |
| Insertion of 23 nucleotides |
|
| IIIa |
| Deletion of base pairs 202-205 |
|
| IIIa | P | Deletion of P |
|
| IIIa | P | Deletion of P |
|
| IIIb |
| Deletion (includes |
|
| IIIb |
| Deletion (includes |
|
| IIIc | P | T |
|
| IIIc | P | T |
|
| IIIc | P | T |
|
| IIIc | P | T |
|
| IIId | P | Internal deletion |
|
| IIIe |
| V2F |
|
| IIIe |
| Y5C |
|
| IIIe |
| T49I |
|
| IIIe |
| Y80H |
|
| IIIe |
| P93L |
|
| IIIe |
| F113C |
|
| IIIe |
| R119H |
|
| IIIf |
| ΩG53 |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Genetics and Biotechnology · Bacterial biofilms and quorum sensing · Bacillus and Francisella bacterial research
INTRODUCTION
Ancestral strains of Bacillus subtilis exhibit two forms of flagellar-mediated motility, called swimming and swarming (1). Swimming motility occurs in a liquid environment and involves the movement of individuals in three dimensions, while swarming motility occurs on a solid surface and cells move as groups in two dimensions (2). While both swimming and swarming require the same flagellar structural genes, swarming has additional genetic requirements. For example, swarming requires the production of surfactin, a secreted surfactant that reduces surface tension and creates a thin layer of water within which to swarm (3, 4). Swarming cells are also hyperflagellated compared to swimming cells and require enhanced activity of the SwrA•DegU hybrid master regulator of flagellar biosynthesis to increase flagellar gene expression (Fig. 1) (5–8). Commonly used domesticated laboratory strains lack swarming motility due to mutations in sfp and swrA that abolish surfactin production and hyperflagellation, respectively (3–5, 9, 10). To better understand swarming motility, we identify and characterize other genes that alter swarming behavior in the ancestral strain of B. subtilis.
A cartoon diagram of the flagellar regulon of Bacillus subtilis. The flagellar regulon of B. subtilis is organized in three tiers. Tier I is the SwrA•DegU hybrid master activator that activates expression of the sigma A-dependent Pfla/che promoter. Tier II is the fla/che operon that contains 32 genes dedicated to the assembly of the flagellar basal body, rod, and hook, as well as genes for chemotaxis signal transduction and the alternative sigma factor SigD. Tier III is the SigD regulon (shown abbreviated) containing proteins involved in flagellar filament assembly and flagellar rotation (among other products). Block arrows indicate genes, and the names of genes mentioned in text are indicated in italics below their corresponding coding region. Small bent arrows indicate promoters, and the names of the promoters mentioned in text are indicated above the promoter location. Thick arrows indicate activation; T bars indicate inhibition. Light green arrows indicate DegU-dependent transcriptional activation; red T-bars indicate Spx-dependent inhibition.
One mutation that enhances swarming motility is a mutation of the gene lonA, which encodes the cytoplasmic protease LonA (11). LonA degrades SwrA during growth in liquid media and requires the adaptor protein SmiA, which both primes SwrA for degradation and delivers it to the LonA protease (7, 12, 13). When cells are inoculated on a surface conducive to swarming, SwrA levels increase seemingly due to the inactivation of proteolysis (7). In turn, a complex of SwrA•DegU binds to and activates the promoter of the fla/che operon, flagellar biosynthesis is activated, flagellar density on the cell surface increases, and swarming commences (6, 8, 14–16). How exactly surface contact antagonizes SwrA proteolysis is unknown, but the inhibitory role of swarming motility is specific to LonA, as mutation of other cytoplasmic proteolysis systems in the cell does not similarly enhance swarming motility (7). Indeed, ClpX, one of several unfoldase partners for the cytoplasmic protease ClpP (17, 18), promotes motility such that both swimming and swarming are abolished when it is mutated (7, 19).
Previous work indicated that mutation of ClpX abolished swimming motility in laboratory strains indirectly through the global transcription factor Spx (19). Spx is restricted to very low levels in the cell by ClpX-mediated proteolysis but accumulates to a high level when ClpX is mutated (20, 21). Spx is an unusual transcription factor in that it acts as a monomer to both activate and repress gene expression (22, 23). Spx activates transcription by interacting with the alpha subunit of RNA polymerase and the vegetative sigma factor SigA and is thought to enhance binding of the RNAP holoenzyme to certain promoters based on a short sequence of DNA upstream of the −35 element (24–28). Spx is also thought to inhibit transcription by binding to the alpha subunit and neutralizing interaction with other transcriptional activators, but the need for a DNA-binding sequence in this case is unclear (29, 30). Spx controls a wide variety of processes in B. subtilis, including activation of the oxidative and/or disulfide stress response pathway (24, 31–33), activation of cell division (34), inhibition of surfactin gene expression (22, 29), inhibition of sulfur uptake (35), and the inhibition of competence for DNA import (20, 36). Relatively few promoters that are directly antagonized by Spx have been identified, and the target responsible for motility inhibition is unknown.
Here, we show that mutation of ClpX abolishes swarming motility in B. subtilis due to a defect in flagellar number. We track the motility defect to a decrease in the level of transcription of the fla/che operon, a result consistent with previous reports in laboratory strains (19). Furthermore, we find that in the absence of ClpX, protein levels of the master motility regulators DegU and SwrA are reduced, suggesting that the defect occurs upstream of fla/che operon transcription. Suppressors that restored swarming to cells mutated for clpX had additional mutations in either lonA or spx. While both classes of suppressors increased the expression of the degU and swrA genes, we infer that LonA primarily acts by restricting SwrA protein accumulation and that Spx primarily acts by reducing transcription of DegU-activated genes. Spx levels rise under cell envelope stress, and we suggest that Spx-mediated repression of motility may serve to limit stress associated with flagellar biosynthesis.
MATERIALS AND METHODS
Growth conditions
B. subtilis strains were grown in lysogeny (LB) broth (10 g tryptone, 5 g yeast extract, 5 g NaCl per L) or on LB plates fortified with 1.5% Bacto agar at 37°C. When appropriate, antibiotics were included at the following concentrations: 5 µg/mL chloramphenicol (cat), 5 µg/mL kanamycin (kan), 100 µg/mL spectinomycin (spec), 10 µg/mL tetracycline (tet), or 1 µg/mL erythromycin plus 25 µg/mL lincomycin (mls).
SPP1 phage transduction
To 200 µL of dense culture grown in TY broth (LB broth supplemented after autoclaving with 10 mM MgSO_4_ and 100 µM MnSO_4_), serial dilutions of SPP1 phage stock were added. To each mixture, 3 mL molten TY top agar (TY supplemented with 0.5% agar) was added, the mixture was poured atop fresh TY plates (TY fortified with 1.5% agar), and the plates were incubated at 30°C overnight. Top agar from a plate containing near-confluent plaques was harvested by scraping into a 15 mL conical tube with 5 mL of TY broth, vortexed, and centrifuged at 6,500 × g for 5 min. The supernatant was passed through a 0.2 µm syringe filter and stored at 4°C.
Recipient cells for transduction were grown to stationary phase in 3 mL TY broth at 37°C. One milliliter of culture was mixed with 25 µL of SPP1 donor phage stock made above. Nine milliliters of TY broth was added to the mixture and allowed to stand at 37°C for 20 min. The transduction mixture was then centrifuged at 6,000 × g for 5 min, the supernatant was discarded, and the pellet was resuspended in the remaining volume. Cell suspension (~100 µL) was then plated on LB plates fortified with the appropriate antibiotic for selection and 10 mM sodium citrate if the antibiotic was chloramphenicol, kanamycin, or spectinomycin.
Strain construction
All constructs were either first introduced into the domesticated Bacillus subtilis strain PY79 by natural competence and then transferred to the undomesticated Bacillus subtilis 3610 background using SPP1-mediated generalized phage transduction (37) or transformed directly into DK1042 (38). All plasmids used in this study are listed in Table S1. All primers used in this study are listed in Table S2. All strains used in this study are listed in Table 1.
Antibiotic replacement constructs
The spx::spec allele was a generous gift from John Helmann (Cornell University).
The lonA::tet insertion-deletion allele was generated using a modified “Gibson” isothermal assembly protocol (39). Briefly, the region upstream of lonA was PCR amplified using the primer pair 7838/7839, and the region downstream of lonA was PCR amplified using the primer pair 7840/7841. Primers 7839 and 7840 contained a short sequence (23 bp) of homology to the tetracycline resistance gene. DNA containing a tetracycline resistance gene was amplified using universal primers 3250/3251 and pDG1515 as a template (40). The three DNA fragments were combined at equimolar amounts to a total volume of 5 µL and added to a 10 µL aliquot of 2× prepared master mix (see below). The reaction was incubated for 60 min at 50°C. The completed reaction was then PCR amplified using primers 7838/7841 to amplify the assembled product. The amplified product was transformed into competent cells of PY79 and then transferred to the 3610 background using SPP1-mediated generalized transduction and selection on tetracycline. The 5× isothermal assembly reaction buffer (500 mM Tris-HCl pH 7.5, 50 mM MgCl_2_, 1 mM of each dNTP [New England BioLabs], 50 mM DTT [Bio-Rad], 312.5 mM PEG-8000 [Fisher Scientific], and 5 mM NAD^+^ [New England BioLabs]) was aliquoted and stored at −80°C. The 2× assembly master mixture was made by combining the prepared 5× isothermal assembly reaction buffer with T5 exonuclease diluted 1:5 with 5× reaction buffer (New England BioLabs) (0.01 units/µL), Phusion DNA polymerase (New England BioLabs) (0.033 units/µL), and Taq DNA ligase (New England BioLabs) (5,328 units/µL). The master mix was aliquoted as 10 µL and stored at −80°C.
In-frame deletions
To generate the ΔclpX in-frame markerless deletion construct, the region upstream of clpX was PCR amplified using the primer pairs 6333/6334 and 6335/6336 using 3610 chromosomal DNA as a template. The PCR products were then assembled by isothermal assembly, and the resulting assembly was digested with BamHI/KpnI and cloned into the BamHI/KpnI sites of pMiniMAD2 which carries a temperature-sensitive origin of replication and an mls resistance cassette (41) to generate pSO18. The plasmid pSO18 was purified from recA^+^ TG1 Escherichia coli and introduced to DK1042 by transformation at the restrictive temperature for plasmid replication (37°C) using mls resistance as a selection. To evict the plasmid, the strain was incubated in 3 mL LB broth at a permissive temperature for plasmid replication (22°C) for 14 h, then serially diluted and plated on LB agar at 37°C. Individual colonies were patched on LB plates and LB plates containing mls to identify mls-sensitive colonies that had evicted the plasmid (42). Chromosomal DNA from colonies that had excised the plasmid was purified and screened by PCR using primers 6333/6336 to determine which isolate had retained the ΔclpX allele.
Complementation constructs
To generate the PclpX-clpX complementation construct pDP538, a PCR product containing the clpX coding region plus 434 base pairs of upstream sequence was amplified from B. subtilis 3610 chromosomal DNA using the primer pair 7410/7411, digested with XhoI and NheI, and cloned into the XhoI and NheI sites of pKM86, containing a polylinker and tetracycline resistance cassette between two arms of ycgO (generous gift of Dr. David Rudner, Harvard Medical School).
Inducible constructs
The Physpank-degU construct was a generous gift of Dr. Nicola Stanley-Wall (University of Dundee).
Transcriptional reporter constructs
To generate the β-galactosidase (lacZ) reporter constructs pAEJ5, pAEJ9, and pAEJ15, PCR products containing the following promoters were amplified from B. subtilis 3610 or DK1042 chromosomal DNA using the primers indicated in parentheses: PdegS (7657/7658) and PdegU (7944/7945). Each PCR product was digested with EcoRI and either BamHI or HindIII and cloned independently into the EcoRI and either BamHI or HindIII sites of plasmid pDG268, which carries a chloramphenicol resistance marker and a polylinker upstream of the lacZ gene between two arms of the amyE gene (43).
Swarm expansion assay
Cells were grown overnight at room temperature in LB broth, back-diluted and grown to mid-log phase at 37°C in LB broth and 1 mM IPTG (if applicable), and resuspended to an OD_600_ of 10 in MQ H_2_O containing 0.5% India ink. Freshly prepared LB plates containing 0.7% Bacto agar (25 mL/plate) and 1 mM IPTG (if applicable) were dried for 10 min in a laminar flow hood, centrally inoculated with 10 µL of the cell suspension, dried for another 10 min, and incubated at 37°C for 6 h. Each strain was done in technical triplicate. The India ink demarcates the origin of the inoculation, and the swarm radius was measured in millimeters relative to this origin. For consistency, an axis was drawn on the back of the plate, and swarm radii measurements were taken along this axis.
Swim assay
Swim agar plates containing 25 mL LB fortified with 0.3% Bacto agar were prepared fresh. Each swim plate was toothpick-inoculated from a colony grown overnight on a plate and photographed for motility after 12 h of incubation at 37°C. Plates were visualized with a camera.
Microscopy
Fluorescence microscopy was performed with a Nikon 80i microscope with a phase-contrast objective Nikon Plan Apo 100× and an Excite 120 metal halide lamp. FM4-64 was visualized with a C-FL HYQ Texas Red Filter Cube (excitation filter 532 nm–587 nm, barrier filter >590 nm). GFP or fluorescent maleimide stain was visualized using a C-FL HYQ FITC Filter Cube (excitation filter 460 nm–500 nm, barrier filter 515 nm–550 nm). Images were captured with a Photometrics Coolsnap HQ2 camera in black and white using NIS-Elements software and subsequently false-colored and superimposed using Fiji v.2.1.0. For microscopy of strains expressing GFP, cells were grown in LB broth at 37°C to an OD_600_ of 0.6–0.85. One milliliter of culture was pelleted, and the pellet was resuspended in 30 µL of 1× phosphate-buffered saline (PBS) containing 5 µg/mL FM 4-64 (Invitrogen #T13320) and incubated for 2 min in the dark at room temperature. Cells were washed with 1 mL 1× PBS and pelleted again. Cells were then resuspended in 30 µL 1× PBS. Cells were observed by spotting 4 µL of the resuspension on a glass microscope slide and immobilized with a poly-L-lysine-treated glass coverslip. Images were captured with NIS-Elements software.
For fluorescence microscopy of flagella, cells were grown in LB broth at 37°C to an OD_600_ of 0.6–0.85. One milliliter of culture was pelleted, and the pellet was resuspended in 50 µL of 1× PBS containing 5 µg/mL Alexa Fluor 488 C_5_ maleimide (Invitrogen; molecular probes A10254) and incubated for 3 min in the dark at room temperature. Cells were washed with 1 mL 1× PBS and pelleted again. The pellet was resuspended in 30 µL of 1× PBS containing 5 µg/mL FM 4-64 and incubated for 2 min in the dark at room temperature. Cells were washed with 1 mL 1× PBS and pelleted again. Cells were then resuspended in 30 µL 1× PBS. Cells were observed by spotting 4 µL of the resuspension on a glass microscope slide and immobilized with a poly-L-lysine-treated glass coverslip. Images were captured with NIS-Elements software.
Western blotting
B. subtilis strains were grown in LB to an OD_600_ of ~1 either from colony inoculation or back-dilution from overnight cultures. One milliliter (or (10 mL) was harvested by centrifugation, and pellets were resuspended to an OD_600_ of 10 (or OD_600_ of 100 if 10 mL samples) in lysis buffer (50 mM Tris pH 7.0, 100 µg/mL lysozyme, 10 µg/mL DNAse I, 100 µg/mL RNAse A, 1 mM phenylmethylsulfonyl fluoride, 10 mM MgCl_2_) and incubated at 37°C for 1 h. 6× sodium dodecyl sulfate (SDS) loading dye supplemented with β-mercaptoethanol (βME) was added to a 1× concentration. Samples were separated by 12 or 15% SDS-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were electroblotted onto nitrocellulose, and the membrane was blocked with 5% skim milk in Tris-buffered saline with Tween 20 and developed with varying dilutions of primary antibody (1:80,000 for α-Hag, 1:10,000 for α-SigD, 1:10,000 for α-DegU, 1:2,000 for α-SwrA, 1:40,000 for α-SigA) and a 1:10,000 dilution secondary antibody (horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G). Ten milliliter samples were used to blot for SigD and DegU, while 1 mL samples were used for the rest. Immunoblot was developed using the Immun-Star HRP developer kit (Bio-Rad).
β-Galactosidase assays
Three biological replicates of each strain were grown in 3 mL of LB broth at 37°C to an OD_600_ of 0.7–1.0. Two hundred microliters of culture was pelleted, and the pellet was resuspended in 200 µL of Z-buffer (16.1 g Na_2_HPO_4_ • 7H_2_O + 5.5 g NaH_2_PO_4_ • H_2_O + 0.75 g KCl + 0.246 g MgSO_4_ • 7H_2_O in 1 L H_2_O, pH = 7) with 0.27% βME. Four microliters of 10 mg/mL lysozyme was added, and samples were incubated at 30°C for 15 min to lyse. In a 96-well plate, lysates were diluted in Z-buffer to 200 µL depending on the strength of the promoter fused to lacZ (Phag = 1:25, Pfla/che = 1:10, PdegS = 1:2, PdegU = 1:1 [undiluted], PswrA = 1:2). Three wells had 200 µL of Z-buffer only and served as negative controls. Forty microliters of 4 mg/mL ortho-nitrophenyl-β-D-galactopyranoside in Z-buffer was added to each reaction. The plate was incubated at 30°C for 1 h, and the OD_420_ and OD_550_ of each well were taken every 2 min. The slope of each sample’s ODs over time was derived. The average slope of the three negative control wells was subtracted from the slope of each experimental well. The Miller Units were calculated using the following formula:
Sequencing of candidate genes
PCR product containing P_fla/che_ was amplified from chromosomal DNA from DS2231 suppressor sox16 using the primers 1921/3042. The Pfla/che PCR product was then sequenced using primers 1921 and 3042 individually. PCR product containing Pspx was amplified from DS2231 suppressors sox5, 10–11, and 13, and from DK6563 suppressor asx12 using the primers 6589/6590. The Pspx PCR product was then sequenced using primers 6589/6590. PCR product containing spx was amplified from DS2231 suppressors sox2–3, 7, 14, and 18–20, and from DK6563 suppressors asx4, 6, 16, and 27 using the primers 6589/6590. The spx PCR product was then sequenced using primers 6591/6590.
Whole-genome sequencing
One colony of each strain was grown overnight in 3 mL of LB broth at room temperature. Two milliliters of culture per strain was harvested, and the pellets were each resuspended in 180 µL enzymatic lysis buffer (20 mM Tris-HCl pH 8, 2 mM ethylenediaminetetraacetic acid, 1.2% Triton X-100). The samples were incubated at 37°C for 30 min. DNA was extracted using the DNeasy kit from Qiagen (#69504): 10 µL of 4 mg/mL RNAse A was added and mixed by pipetting. The samples were incubated at 37°C for 30 min. Twenty-five microliters of Proteinase K was added and mixed by vortexing for 30 s. The samples were incubated at 37°C for 10 min. Two hundred microliters of Buffer AL was added and mixed by vortexing for 30 s. The samples were incubated at 56°C for 30 min. Two hundred microliters of 100% ethanol was added and mixed by vortexing for 30 s. The entire sample was pipetted onto a DNeasy mini spin column in a 2 mL collection tube and centrifuged at 10,000 × g for 1 min. The collection tube was discarded and placed into a fresh one. Five hundred microliters of Buffer AW1 was added to the column, and the column was centrifuged at 10,000 × g for 1 min. The collection tube was discarded and placed into a fresh one. Five hundred microliters of Buffer AW2 was added to the column, and the column was centrifuged at maximum speed for 3 min. The flow-through was discarded, and the column was replaced and centrifuged again at maximum speed for 1 min to remove residual ethanol. The collection tube was discarded and placed into a fresh Eppendorf tube. One hundred microliters of Buffer AE was added to the column, and the column was incubated at room temperature for 2 min. The column was centrifuged at 10,000 × g for 1 min to elute DNA. The elution step was repeated to elute additional DNA, for a total of 200 µL. An additional RNAse A treatment was performed by adding 4 µL of 4 mg/mL RNAse A and incubating at room temperature for 10 min. Samples were submitted to the Center for Genomics and Bioinformatics at Indiana University for library preparation and data analysis.
RESULTS
ClpX promotes swarming motility by enhancing flagellar biosynthesis
The ClpX unfoldase subunit of the ClpP protease has been shown to be required for swimming motility through soft agar plates in B. subtilis laboratory strains (19). Consistent with a role in promoting flagellar motility, mutation of the clpX gene abolished swarming (Fig. 2A) (7, 44) and reduced swimming in the NCIB3610 ancestral strain background (Fig. 3), and both phenotypes were restored by complementation when clpX was expressed from its native promoter and inserted at an ectopic site in the chromosome ([Fig. 2A and 3](#F2 F3)). We conclude that ClpX is required for swarming and enhances swimming motility. We note that mutation of clpX in the ancestral strain did not completely abolish swimming through a soft agar matrix as previously reported, and we infer that the difference in phenotype may be due to genetic differences between strain backgrounds.
Mutation of LonA and Spx restores swarming in the absence of ClpX. Quantitative swarm expansion assays for the strains indicated in each panel. Genes in italics are mutated, genes in parentheses are complementation constructs expressed under the gene’s native promoter and inserted at an ectopic site in the chromosome, and a “+” indicates that the gene was induced with 1 mM IPTG. Each data point is the average of three technical replicates. (A) The clpX mutant phenotype was complemented by reintroducing clpX under its native promoter at an ectopic site. The following strains were used to generate the data in this panel: WT (DK1042), clpX (clpX) (DB22), clpX (DB10). (B) The clpX mutant phenotype was rescued when the fla/che operon was artificially overexpressed. The following strains were used to generate the data in this panel: fla/che+ (DK7584, 1 mM IPTG added), clpX fla/che+ (DK6647, 1 mM IPTG added), clpX (DK6647, 0 mM IPTG added). (C) The clpX mutant phenotype was partially rescued when the degU gene was artificially overexpressed. The following strains were used to generate the data in this panel: degU+ (DB644, 1 mM IPTG added), clpX degU+ (DB470, 1 mM IPTG added), clpX (DB470, 0 mM IPTG added). (D) The clpX mutant phenotype was partially rescued when the swrA gene was artificially overexpressed. The following strains were used to generate the data in this panel: swrA+ (DB548, 1 mM IPTG added), clpX swrA+ (DB3250, 1 mM IPTG added), clpX (DB3250, 0 mM IPTG added). (E) The clpX mutant phenotype was rescued when both the degU and swrA genes were artificially overexpressed. The following strains were used to generate the data in this panel: degU+swrA+ (DB3071, 1 mM IPTG added), clpX degU+swrA+ (DB2379, 1 mM IPTG added), clpX (DB2379, 0 mM IPTG added). (F) The clpX mutant phenotype was partially rescued when the lonA gene was also mutated. The following strains were used to generate the data in this panel: WT (DK1042), clpX lonA (DB23), clpX (DB10). (G) The clpX mutant phenotype was partially rescued when the spx gene was also mutated. The following strains were used to generate the data in this panel: WT (DK1042), clpX spx (DB17), clpX (DB10). (H) The clpX mutant phenotype was fully rescued when both the lonA and spx genes were also mutated. The following strains were used to generate the data in this panel: WT (DK1042), clpX lonA spx (DB53), clpX (DB10).
Mutation of LonA and Spx restores swimming in the absence of ClpX. Cells were centrally inoculated by toothpick on 0.3% agar swim plates, incubated for 12 h at 37°C, and filmed against a black background such that uncolonized agar appears black and bacterial growth appears white. Genes in italics are mutated, and genes in parentheses are complementation constructs expressed under the gene’s native promoter and inserted at an ectopic site in the chromosome. Each strain is additionally mutated for srfAA and epsH to abolish the contribution of sliding motility on the surface. The following strains were used to generate the data in this figure: WT (DK9597), clpX (DK9609), clpX (clpX) (DK9925), swrA (DB117), clpX swrA (DK9934), degU (DB1246), clpX degU (DB1304), clpX lonA (DB62), clpX spx (DK9618), clpX lonA spx (DB64).
One potentially relevant genetic locus that could explain the differential consequence of a clpX mutation in ancestral and laboratory strains is the gene swrA, which encodes the master flagellar activator component SwrA. Laboratory strains are naturally defective for SwrA, whereas ancestral strains are not (5, 10, 45). Mutation of swrA reduced swimming in the ancestral background (Fig. 3), and cells doubly mutated for both swrA and clpX recapitulated the more severe clpX mutant defect in laboratory strain swimming (Fig. 3) (19). SwrA cooperates with the response regulator DegU to activate flagellar gene expression, and like mutation of SwrA, mutation of degU alone reduced swimming and further exacerbated the clpX mutant phenotype when simultaneously mutated (Fig. 3). We conclude that ClpX enhances flagellar motility in B. subtilis, the extent of which depends on the genetic background.
One way in which a clpX mutant could be defective in swarming is if it failed to produce surfactin. Indeed, mutation of clpX has been shown to result in the inhibition of the promoter for the srf operon responsible for surfactin biosynthesis (46, 47). On swarm agar plates, however, a large zone of clear watery surface indicative of surfactin production was observed to surround the non-swarming colony (3, 48). It is unclear why cells mutated for ClpX appeared to produce abundant surfactin, but we note that previous work documenting reduced surfactin gene expression was performed in domesticated laboratory strains that lack surfactin production, and the presence of the ancestral allele of sfp alters surfactin gene expression (9). Whatever the case with regard to srf gene expression, we conclude that swarming is likely not limited by surfactin production in the absence of ClpX.
Another reason that the clpX mutant might be impaired for swarming motility is if it assembled fewer flagella on the cell surface (7, 49). To determine whether flagellar number was reduced in the absence of ClpX, a version of the flagellar filament protein Hag, which could be fluorescently labeled by the addition of a maleimide stain (Hag^T209C^), was introduced (50). Mutation of clpX appeared to dramatically reduce flagellar density per cell relative to wild type (Fig. 4A). Moreover, mutation of clpX decreased Hag protein accumulation in Western blot analysis, while there was no change in SigA protein levels, which were used as a loading control (Fig. 5). Mutation of clpX also reduced expression of a transcriptional reporter in which the hag promoter, Phag, was fused to the lacZ gene encoding β-galactosidase (Fig. 6A). For each assay, wild-type phenotypes were restored when clpX was ectopically complemented (Fig. 4A, Fig. 5, Fig. 6A). We conclude that the swarming defect in the absence of ClpX is correlated with reduced transcription from the Phag promoter which, in turn, reduces the filament protein pool and restricts flagellar filament assembly.
Cells mutated for ClpX have a reduced number of flagella per cell. Fluorescence microscopy observation of the cell membrane and flagella (A) or basal bodies (B) in a variety of genetic backgrounds. Genes in italics are mutated, and genes in parentheses are complementation constructs expressed under the gene’s native promoter and inserted at an ectopic site in the chromosome. (A) Fluorescent FM 4-64 membrane (false-colored red) and Alexa Fluor maleimide of HagT209C (false-colored green) staining in a variety of genetic backgrounds. Each strain is additionally mutated for the native copy of the hag gene and has the hagT209C allele expressed under the native promoter at an ectopic site in the chromosome. The following strains were used to generate the data in this panel: WT (DS6239), clpX (DK9981), clpX (clpX) (DK9985), clpX lonA (DB61), clpX spx (DK9984), clpX lonA spx (DB63). (B) Fluorescent FM 4-64 membrane staining (false-colored red) and fluorescent FliM-GFP translational fusion (false-colored green) in a variety of genetic backgrounds. Each strain is additionally mutated for the native copy of the fliM gene and has the fliM-GFP allele expressed under the native promoter at an ectopic site in the chromosome. The following strains were used to generate the data in this panel: WT (DS9446), clpX (DB1351), clpX (clpX) (DB1372), clpX lonA (DB1373), clpX spx (DB1371), clpX lonA spx (DB1391).
Mutation of LonA and Spx increases levels of key motility proteins in the absence of ClpX. Western blot analysis with the primary antibody indicated on the left and the genetic background indicated above. Genes in italics are mutated, and genes in parentheses are complementation constructs expressed under the gene’s native promoter and inserted at an ectopic site in the chromosome. Carets on the right indicate the location of the corresponding protein. SigA is used here as a loading control. We note a cross-reacting band below the band corresponding to SigD and also one below the band corresponding to SwrA. The following strains were used to generate the data in this figure: WT (DK1042), clpX (DB10), clpX (clpX) (DB22), clpX spx (DB17), clpX lonA (DB23), clpX lonA spx (DB53). The same samples were run in each Western blot shown, and each Western blot shown is from a different gel.
Mutation of ClpX reduces transcription from a variety of transcriptional reporters. β-Galactosidase assays measuring transcriptional activity from key motility promoters in a variety of genetic backgrounds. Genes in italics are mutated, and genes in parentheses are complementation constructs expressed under the gene’s native promoter and inserted at an ectopic site in the chromosome. Each bar shows the mean Miller Units of three biological replicates, with error bars indicating standard deviation. (A) Expression from the promoter of the hag gene (Phag-lacZ). The following strains were used to generate the data in this panel: WT (DK5457), clpX (DB20), clpX (clpX) (DB26), clpX lonA (DB28), clpX spx (DB24), clpX lonA spx (DB54). (B) Expression from the promoter of the fla/che operon (Pfla/che-lacZ). The following strains were used to generate the data in this panel: WT (DK2399), clpX (DB67), clpX (clpX) (DB68), clpX lonA (DB70), clpX spx (DB69), clpX lonA spx (DB71). (C) Expression from the promoter of the degSU operon (PdegS-lacZ). The following strains were used to generate the data in this panel: WT (DK9529), clpX (DB1527), clpX (clpX) (DB1529), clpX lonA (DB1530), clpX spx (DB1528), clpX lonA spx (DB1531). (D) Expression from the internal promoter directly upstream of the degU gene (PdegU-lacZ). The following strains were used to generate the data in this panel: WT (DB559), clpX (DB807), clpX (clpX) (DB811), clpX lonA (DB813), clpX spx (DB809), clpX lonA spx (DB848). (E) Expression from the promoter of the swrA gene (PswrA-lacZ). The following strains were used to generate the data in this panel: WT (DK6613), clpX (DB806), clpX (clpX) (DB810), clpX lonA (DB812), clpX spx (DB808), clpX lonA spx (DB847).
The Phag promoter is expressed by RNA polymerase and the alternative sigma factor SigD (51), and thus a reduction in Phag expression could be due to a reduction in SigD activity (Fig. 1). SigD activity is antagonized by direct binding with its cognate anti-sigma factor FlgM (52–54). Reduced SigD activity alone, however, was insufficient to explain the swarming defect of the clpX mutant, as simultaneous mutation of ClpX and FlgM did not restore swarming (Fig. S1A). SigD activity is also controlled by the level of SigD protein, and not only were SigD levels slightly reduced in the clpX mutant, the levels increased to wild type when clpX was ectopically complemented (Fig. 5). We conclude that although reduced SigD activity is not responsible for the lack of swarming in the clpX mutant, the reduction in SigD levels may indicate that the effect of ClpX is upstream of SigD expression.
One way in which ClpX could act upstream to modulate SigD levels is by increasing expression of the sigD gene. The gene encoding SigD, sigD, is the penultimate gene of the 27 kb-long fla/che operon that encodes many of the proteins required for the assembly of the flagellar basal body (Fig. 1) (55–58). To determine whether mutation of ClpX affects basal body formation, GFP was fused to the C-ring protein FliM, which forms puncta indicative of intact basal bodies (15, 59). Cells mutated for ClpX showed a reduction in the number of flagellar basal bodies relative to wild type, with a corresponding increase in the amount of cytoplasmic fluorescence, presumably due to excess unassembled FliM (Fig. 4B). Puncta number increased and cytoplasmic fluorescence decreased upon ectopic complementation of clpX (Fig. 4B). We conclude that both SigD levels and the number of flagellar basal bodies decrease in the absence of ClpX, both likely due to decreased expression of the fla/che operon.
Expression of the fla/che operon is controlled by the Pfla/che promoter recognized by RNA polymerase and the vegetative sigma factor SigA (Fig. 1) (60, 61). A reduction in transcription from Pfla/che could explain both the reduction in basal body number and SigD levels. Expression from a transcriptional reporter in which Pfla/che was fused to the lacZ gene was reduced twofold from wild-type levels in the absence of ClpX, a level of reduction previously shown to be sufficient to abolish swarming motility (6, 7), (Fig. 6B), and expression was restored by clpX ectopic complementation. To determine whether the inhibition of the fla/che operon was sufficient to abolish swarming motility, the native Pfla/che promoter was replaced with an artificial IPTG-inducible promoter (15). In wild-type cells, motility was IPTG-dependent, and induction with IPTG resulted in rapid swarming without the typically observed lag period (Fig. 2B). Induction of the fla/che operon in the absence of ClpX restored swarming, albeit with a lag period similar to that observed in wild-type cells (Fig. 2B). We conclude that overexpression of the fla/che operon is sufficient to restore swarming in the absence of ClpX. We infer that the inhibition of flagellar motility by ClpX occurs at or before the level of Pfla/che, consistent with previous results and models (19).
ClpX enhances swarming motility through the SwrA•DegU master activator
Pfla/che is activated by a heteromeric transcription factor complex made of the response regulator DegU and the accessory protein SwrA (8, 14, 15, 62) (Fig. 1), and Western blot analysis indicated that the levels of both DegU and SwrA were reduced in the absence of ClpX (Fig. 5). The gene degU, encoding DegU, is expressed from its own promoter but is also part of a two-gene operon preceded by the gene degS, encoding DegS, the DegU-cognate histidine kinase (Fig. 1) (63–66). Expression of a lacZ transcriptional reporter to PdegS marginally increased compared to wild type in the absence of ClpX (Fig. 6C), but expression decreased from reporters of PdegU (Fig. 6D) and PswrA (Fig. 6E), following the trends previously observed with other motility promoters. Transcription from both PdegU and PswrA could be restored by ectopic complementation of clpX (Fig. 6D, Fig. 6E). We conclude that the reduction of swarming motility in the absence of ClpX is correlated with a reduction in the transcription of genes encoding DegU and SwrA.
To determine if the repression of DegU and SwrA was sufficient to account for the swarming defect in the absence of ClpX, artificial overexpression constructs were introduced into wild type and ClpX mutant backgrounds. Overexpression of either degU or swrA abolished the swarming lag period in wild-type cells and increased swarming motility in the absence of ClpX (Fig. 2C, Fig. 2D). Simultaneous overexpression of both SwrA and DegU also enhanced swarming of the clpX mutant and did so to a higher level than overexpression of either SwrA or DegU alone (Fig. 2E). We conclude that the reduced expression of each component of the hybrid master regulator of flagellar biosynthesis contributes to the swarming defect of a clpX mutant.
Mutations that enhance fla/che expression suppress the absence of ClpX
To determine how swarming is inhibited in the absence of ClpX, spontaneous suppressors were obtained that restored swarming motility to a clpX mutant. To do so, a clpX mutant originally grows as a tight central colony on a swarm agar plate, but upon prolonged incubation, motile flares emerged and were clonally isolated. Twenty-three suppressor mutations were isolated from flares on independent plates, mapped by either candidate gene or whole-genome sequencing, and organized into three genetic classes (Table 2). Class I suppressors were in or near the Pfla/che promoter of the fla/che flagellar operon. One suppressor contained a mutation that changed the −10 box closer to consensus for SigA (class Ia), and another suppressor deleted an intrinsic terminator of codY, the gene immediately upstream of the fla/che operon (class Ib). Both mutations have been observed in other suppressor selection screens and have been shown to increase fla/che operon expression (6, 49, 67) (Fig. S2A and B). We conclude that both class I mutations increase fla/che transcription, thereby bypassing the need for ClpX. We further conclude that the mutations support the inference that ClpX acts at or above the level of the Pfla/che promoter.
Class II suppressors contained frameshift mutations in the gene lonA, encoding LonA, a protease responsible for the regulatory proteolysis of SwrA (Fig. S2C) (7). Mutation of lonA increased swimming motility in the absence of ClpX (Fig. 3) and partially restored the ability to swarm (Fig. 2F). The improved motility of the clpX lonA double mutant was correlated both with an increase in the number of flagellar basal bodies (Fig. 4B) and in the number of flagellar filaments on the cell surface (Fig. 4A). Moreover, the reductions in Hag and SigD protein levels (Fig. 5) as well as in hag and fla/che reporter expression were increased in the double mutant (Fig. 6A and B). The levels of swrA and degU expression by transcriptional reporter activity also increased in the clpX lonA double mutant (Fig. 6D, Fig. 6E), and while SwrA protein levels increased dramatically, DegU protein levels did not (Fig. 5). We conclude that the absence of LonA improves each of the motility phenotypes tested in cells lacking ClpX by increasing SwrA protein levels, which in turn increases fla/che operon transcription. Based on the class I and II mutations, we conclude that one way to enhance swarming motility in the absence of ClpX is by increasing expression of the fla/che operon.
Mutation of Spx suppresses the absence of ClpX by increasing SwrA and DegU levels
The final and largest class of suppressors was class III that disrupted the gene spx, encoding the global transcriptional regulator Spx. Three of the Class III suppressor mutants contained large deletions in the spx open reading frame (class IIIa) (Fig. S2D), and another two suppressors contained even larger deletions of multiple genes that included spx (class IIIb) (Fig. S2E). Four suppressors were point mutations in the −10 box of the spx promoter (Pspx) such that it was farther from consensus for SigA (class IIIc) (Fig. S2F), and one was a −10 box deletion (class IIId). Finally, seven suppressors introduced missense mutations in the spx coding region (class IIIe) (Fig. S2G through H), while an eighth inserted one full codon into the reading frame (class IIIf) (Fig. S2I). Spx is degraded by ClpXP with the aid of the adaptor protein YjbH and is often isolated as a suppressor of clpP and clpX mutant phenotypes (20, 68, 69). In sum, we infer that class III suppressor mutations likely decrease Spx levels and/or function.
To explore the mechanism of Spx-mediated repression of motility, we introduced a mutation in spx into our clpX mutant cells and assessed each of the phenotypes previously tested. Consistent with the spontaneous mutants, mutation of Spx increased swarming motility in the absence of ClpX (Fig. 2G), and also enhanced swimming (Fig. 3). Likewise, mutation in the spx enhanced flagellin gene expression (Fig. 6A), flagellin protein levels (Fig. 5), basal body number (Fig. 4B), and flagellar number per cell (Fig. 4A) when ClpX was mutated. While Hag and SigD protein levels appeared to be increased to wild-type levels in the clpX spx double mutant, the increase in the levels of both DegU and SwrA appeared to exceed that of the wild type (Fig. 5). Consistent with elevated DegU and SwrA, we observed a restoration in expression from Pfla/che in the clpX spx double mutant (Fig. 6B). We conclude that Spx accumulation inhibits the accumulation of SwrA and DegU, and the reduction in the flagellar master regulator components restricts the cells’ ability to produce flagella sufficient for swarming.
To determine whether Spx inhibited SwrA and DegU expression at the transcriptional level, reporter gene expression assays were conducted. Expression from the PdegU and PswrA reporters was increased in the clpX mutant background when spx was also mutated (Fig. 6D and E), but expression from the PdegS promoter was not (Fig. 6C). We conclude that in cells mutated for ClpX, Spx accumulates and most likely represses motility by inhibiting transcription from PdegU and PswrA, as artificial induction of both swrA and degU restored swarming to our clpX mutant close to wild-type levels (Fig. 2E). Thus, Spx inhibits motility at the highest level by restricting expression of the master activator components.
Our genetic analysis suggests that swarming motility can be restored to a clpX mutant by increasing expression of the fla/che operon, either directly by improving the promoter, or by increasing accumulation of SwrA and DegU. Mutation of LonA restores swarming by relieving proteolytic restriction of SwrA levels, and mutation of Spx restores swarming by relieving repression on both degU and swrA gene expression. Thus, each suppressor seems to restore components of the hybrid master regulator in different ways. Consistent with parallel mechanisms, cells mutated for ClpX, LonA, and Spx simultaneously increased swarming (Fig. 2H), Pfla/che, PdegU, and PswrA reporter gene expression (Fig. 6B, D, and E), and SwrA protein (Fig. 5) to levels greater than either clpX-containing double mutant alone. We note that while Spx has been shown to enhance expression of LonA (36, 70), we conclude that Spx and LonA act in parallel and by separate mechanisms to restrict flagellar biosynthesis at the level of the hybrid master activator of flagellar biosynthesis.
DISCUSSION
Flagellar gene expression is hierarchical and is organized in at least three regulatory tiers (71–74). In Bacillus subtilis, the first tier is the hybrid master activator SwrA•DegU, which in turn activates the second tier fla/che operon encoding the basal body, rod, and hook structural components (1). The third tier constitutes a regulon of genes under control of the alternative sigma factor SigD (6, 75), which becomes activated after basal body-rod-hook assembly is complete (76, 77), and contains genes required for the subsequent assembly of the flagellar filament (51, 78). Previous work indicated that when ClpX is mutated, Spx accumulates and inhibits swimming motility at the level of fla/che operon expression (tier II) (19, 70). Here, we show that swarming motility is also inhibited in the absence of ClpX, and genetic suppressor analysis indicated that Spx inhibits the master activator (tier I). Thus, our work reinforces previous observations that Spx inhibits expression of the fla/che operon but does so at one level higher than previously recognized by restricting production of both SwrA and DegU.
Spx is an unusual transcription factor in that it acts as a conditional subunit of RNA polymerase, binding between one of the alpha subunits and the vegetative sigma factor SigA (28, 79). Spx is thought to activate the expression of specific genes by stabilizing RNA polymerase interaction at promoters that contain a particular −44 element (25, 26, 36). Spx is thought to repress gene expression by a similar mechanism, whereby its presence in RNA polymerase interferes with transcriptional activators that require contact with the alpha subunit’s C-terminal domain (22, 30, 80). Here, we show that high levels of Spx inhibit transcription from the PswrA, PdegU, and Pfla/che promoters, and the effect can be bypassed either by artificial expression of the fla/che operon or both SwrA and DegU together. Consistent with sequence-independent inhibition, a previously published ChIP-seq analysis of Spx binding sites indicates that Spx does not bind to any of these promoters (36). Thus, our results seem consistent with a model whereby Spx inhibits by a DNA-independent anti-alpha mechanism, and it may antagonize the expression of all three motility promoters in parallel, as they are all activated by the transcription factor DegU (8, 62, 81, 82).
DegU is a highly pleiotropic DNA-binding response regulator, but the sequence to which it binds and the mechanism of transcriptional activation are poorly understood. Recent work indicated that DegU binds upstream of the Pfla/che promoter but does not activate it until SwrA interaction causes DegU to oligomerize, perhaps facilitating interaction with the alpha C-terminal domain of RNA polymerase (8, 14, 15, 62). ChIP-seq analysis also indicated that DegU bound near the PswrA and PdegU promoters in vivo, and DegU has been shown to be required for the activation of each (8, 14, 81, 83). As DegU activates all of the promoters that appear to be antagonized by Spx, we infer that at high levels, Spx inhibits swarming motility by blocking SwrA•DegU autoactivation. Consistent with Spx inhibiting both components of the master activator, mutation of Spx bypasses the need for ClpX and restores the levels of both proteins and robust swarming (Fig. 2G), whereas mutation of LonA partially compensates by increasing SwrA alone, and swarming restoration is weak (Fig. 2F). We infer that the primary role of Spx is to inhibit DegU-dependent activation, as high Spx levels inhibit swimming in laboratory strains that naturally lack SwrA (5, 10, 19, 84), and DegU activates other promoters related to motility (85).
Spx is normally kept at a low level in the cell by ClpXP and the proteolytic adaptor YjbH (68, 69), and proteolysis is thought to be relieved under stress conditions (22, 24, 86). Why flagellar motility would be inhibited under stress is unclear, and it is also unclear which stress conditions would lead to Spx-mediated inhibition. One possible stress could be flagellar assembly itself; as when cells are placed on a swarming-permissive surface, the density of flagella on the cell surface doubles and reaches nearly 30 flagella per cell (7). Enhanced transcription and translation of flagellar subunits could increase protein misfolding, proteotoxic stress known to increase Spx levels, and Spx in turn activates expression of protein-folding chaperones and other quality control systems (70, 87). Moreover, how flagella are assembled through the B. subtilis cell wall is unknown but could require peptidoglycan remodeling and/or insertion during peptidoglycan synthesis (1, 44, 59, 88), and recent work has indicated that cell wall stress also causes Spx to accumulate (89). In short, we posit that the production and insertion of a large number of trans-envelope machines may be stressful. Spx could be a means to restrict cellular damage from runaway flagellar biosynthesis, and our model suggests that Spx may dampen a DegU-mediated positive feedback loop.
Finally, our data provide two insights into the mechanism of Spx-mediated inhibition. First, simultaneous overexpression of both SwrA and DegU was sufficient to override high levels of Spx (Fig. 2E). Thus, although Spx interacts with RNA polymerase in the absence of DNA, there appears to be a competitive aspect to inhibition such that an excess of activator can either outcompete or displace Spx. We infer that however Spx inhibits transcriptional activation, it is likely not due to a non-competitive allosteric change in RNA polymerase. Second, the model that Spx acts like an anti-alpha factor is partly predicated on its inhibition of the ComA-activated PsrfAA promoter in domesticated laboratory strains that do not produce surfactin (9, 22, 23). However, we find that the inhibition of swarming motility by high levels of Spx in the ancestral strain is not due to a lack of surfactin production. Indeed, the clpX mutant produced levels of surfactin that were not only detectable to the naked eye but also sufficient to select for suppressors that restore swarming. Why Spx fails to restrict surfactin production under swarming conditions is unclear, but perhaps either the inhibitory activity of Spx is more specific than previously recognized or Spx impairs surfactin activation indirectly. Precisely how Spx antagonizes transcription remains unknown.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mukherjee S, Kearns DB. 2014. The structure and regulation of flagella in Bacillus subtilis. Annu Rev Genet 48:319–340. doi:10.1146/annurev-genet-120213-09240625251856 PMC 4869327 · doi ↗ · pubmed ↗
- 2Kearns DB. 2010. A field guide to bacterial swarming motility. Nat Rev Microbiol 8:634–644. doi:10.1038/nrmicro 240520694026 PMC 3135019 · doi ↗ · pubmed ↗
- 3Kearns DB, Losick R. 2003. Swarming motility in undomesticated Bacillus subtilis. Mol Microbiol 49:581–590. doi:10.1046/j.1365-2958.2003.03584.x 12864845 · doi ↗ · pubmed ↗
- 4Julkowska D, Obuchowski M, Holland IB, Séror SJ. 2005. Comparative analysis of the development of swarming communities of Bacillus subtilis 168 and a natural wild type: critical effects of surfactin and the composition of the medium. J Bacteriol 187:65–76. doi:10.1128/JB.187.1.65-76.200515601689 PMC 538812 · doi ↗ · pubmed ↗
- 5Calvio C, Celandroni F, Ghelardi E, Amati G, Salvetti S, Ceciliani F, Galizzi A, Senesi S. 2005. Swarming differentiation and swimming motility in Bacillus subtilis are controlled by swr A, a newly identified dicistronic operon. J Bacteriol 187:5356–5366. doi:10.1128/JB.187.15.5356-5366.200516030230 PMC 1196031 · doi ↗ · pubmed ↗
- 6Kearns DB, Losick R. 2005. Cell population heterogeneity during growth of Bacillus subtilis. Genes Dev 19:3083–3094. doi:10.1101/gad.137390516357223 PMC 1315410 · doi ↗ · pubmed ↗
- 7Mukherjee S, Bree AC, Liu J, Patrick JE, Chien P, Kearns DB. 2015. Adaptor-mediated Lon proteolysis restricts Bacillus subtilis hyperflagellation. Proc Natl Acad Sci USA 112:250–255. doi:10.1073/pnas.141741911225538299 PMC 4291670 · doi ↗ · pubmed ↗
- 8Mishra A, Hughes AC, Amon JD, Rudner DZ, Wang X, Kearns DB. 2024. Swr A-mediated multimerization of Deg U and an upstream activation sequence enhance flagellar gene expression in Bacillus subtilis. J Mol Biol 436:168419. doi:10.1016/j.jmb.2023.16841938141873 PMC 11462632 · doi ↗ · pubmed ↗