Wolbachia-mediated reproductive manipulation in rice planthoppers
Yue-Di Niu, Meng-Ke Wang, Zhi-Chao Yan, Xiao-Li Bing, Xiao-Yue Hong

TL;DR
This review explores how Wolbachia bacteria influence the reproduction of rice planthoppers, offering insights into pest control strategies.
Contribution
The paper highlights strain-specific Wolbachia effects and identifies host and bacterial factors involved in reproductive manipulation.
Findings
Wolbachia strains wLug, wStri, and wSfur have distinct effects on planthopper reproduction and cytoplasmic incompatibility.
The wStri genome contains three copies of CI-related genes, and host proteins CAL and NDUFA8 are linked to CI processes.
Wolbachia boosts planthopper reproduction by synthesizing B vitamins, upregulating vitellogenin, and promoting germ cell division.
Abstract
Rice planthoppers, including brown (Nilaparvata lugens), small brown (Laodelphax striatellus), and white-backed (Sogatella furcifera) planthoppers, are major agricultural pests in China and severely affect rice production and food security. The endosymbiotic bacterium Wolbachia is commonly found in these insects, where it regulates reproduction through mechanisms such as cytoplasmic incompatibility (CI) and increased fertility. In this review, we discuss the strain-specific effects of Wolbachia: wLug (in N. lugens, < 50% infection) increases fecundity without CI; wStri (in L. striatellus, 99% infection) induces complete CI and enhances reproduction; and wSfur (in S. furcifera, 90% infection) shows weak or no CI with minimal fecundity effects. Additionally, while wStri can induce CI in N. lugens, its intensity is reduced, suggesting that both the symbiont and the host influence CI…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
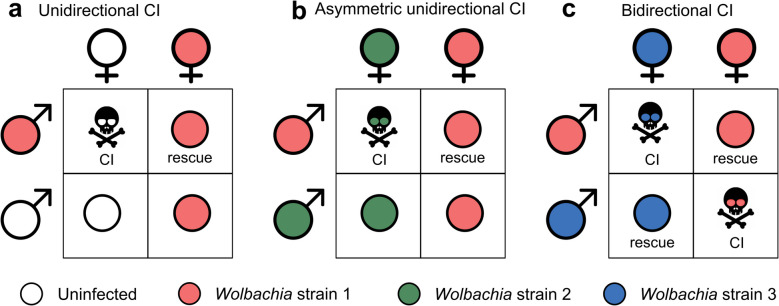 Figure 1
Figure 1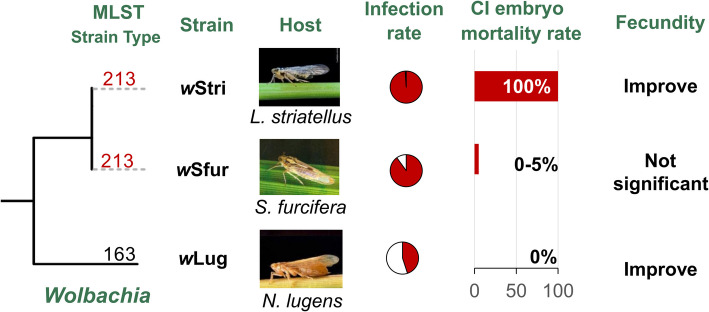 Figure 2
Figure 2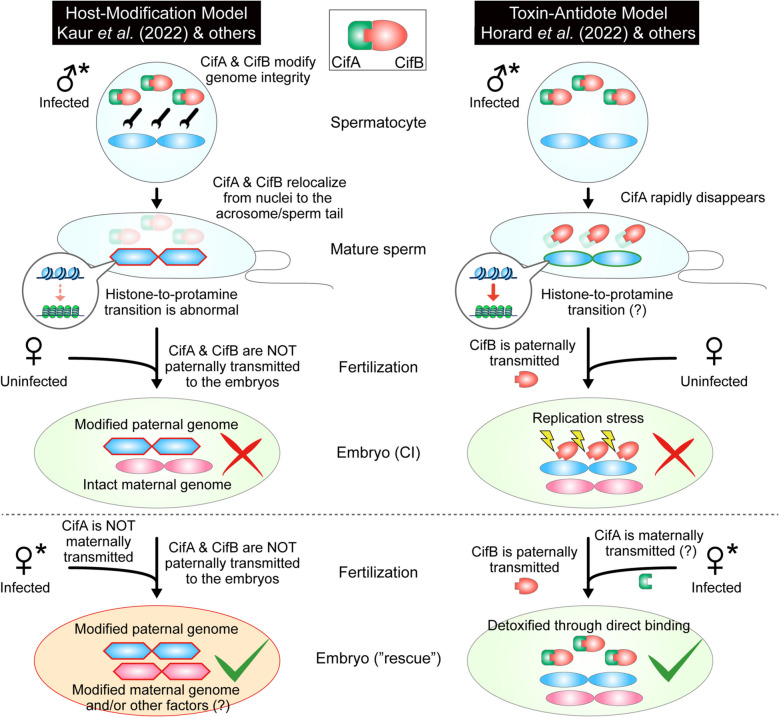 Figure 3
Figure 3- —http://dx.doi.org/10.13039/501100013142Key Research and Development Project of Hainan Province
- —Young Elite Scientists Sponsorship Program by Jiangsu Association for Science and Technology
- —http://dx.doi.org/10.13039/501100019001Center for Artificial Intelligence and Advanced Robotics, National Taiwan University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect symbiosis and bacterial influences
Introduction
Rice planthoppers include three main pests: the brown planthopper Nilaparvata lugens (Stål), the small brown planthopper Laodelphax striatellus (Fallén) and the white-backed planthopper Sogatella furcifera (Horváth). Rice planthoppers damage rice plants by directly piercing and sucking, penetrating the stems by laying eggs, and transmitting rice diseases [1]. Although individuals are small, rice planthoppers have a high reproductive capacity, transmit numerous viruses, migrate over long distances, and are highly resistant to pesticides [2, 3]. They are the most widespread and extensively important crop pests [4]. Among the three rice planthoppers, in addition to the brown planthopper, which feeds exclusively on rice, the small brown planthopper and the white-backed planthopper also damage other crops. In particular, maize and wheat virus diseases transmitted by the small brown planthopper have experienced increasing outbreaks and epidemics in recent years, causing severe losses in agricultural production.
Wolbachia is an intracellular symbiont that is widely distributed among insects and is capable of infecting reproductive tissues such as the testis and ovaries of the host. Wolbachia can be transmitted vertically to offspring through the maternal line [5, 6]. Wolbachia is known for regulating host reproduction, affecting host fecundity, and even regulating insect reproductive patterns and sex ratios. Wolbachia regulates host reproduction by killing infected males (male killing), causing genotypically male individuals to develop into females (feminization), producing females through parthenogenesis without mating or fertilization with a male (parthenogenesis), and inducing cytoplasmic incompatibility (CI) [7]. Wolbachia-induced CI refers to the phenomenon in which the offspring of fertilized eggs die during embryonic development after mating between a Wolbachia-infected male and an uninfected female (unidirectional cytoplasmic incompatibility) or a female infected with a different Wolbachia strain (bidirectional cytoplasmic incompatibility) (Fig. 1) [8]. These mechanisms can reduce or even kill males who do not directly help their transmission, increase the number of carrier females, and elevate the chance of bacterial transmission among host populations, which has important ecological significance.Fig. 1. Types of Wolbachia-induced cytoplasmic incompatibility [8]. a Unidirectional CI, b asymmetric unidirectional CI, c bidirectional CI
This review focuses on the interaction between rice planthoppers and Wolbachia and elucidates the role and mechanisms of Wolbachia in regulating rice planthopper reproduction, which may contribute to uncovering the mechanisms of rice planthopper disasters and exploring new technologies for integrated rice planthopper control via symbiotic bacteria.
Types of Wolbachia-mediated reproductive regulation in rice planthoppers
Rice planthoppers harbor a variety of symbiotic bacteria, among which the endosymbiont Wolbachia is highly abundant in their hosts [9]. In L. striatellus, Wolbachia is the most abundant in all wild populations [10–12]. The infection of Wolbachia in planthopper populations appears to play a significant role in shaping the structure of microbial communities [12, 13], thereby influencing the distribution of the host. It significantly affects reproductive capacity and induces CI [14, 15]. Studies have shown significant variations in the Wolbachia strains infecting these planthoppers, including differences in infection rates, effects on host fecundity, and the ability to induce CI among three rice planthopper species [16–19] (Fig. 2).Fig. 2. Reproductive effects of different Wolbachia strains on three planthopper species
The impact of Wolbachia on host reproductive capacity alters the base population size and is one of the factors influencing rice planthopper infestations. According to the CI principle, male insects carrying specific Wolbachia strains are released to mate with naturally uninfected females, causing fertilized eggs to die during embryonic development and effectively suppressing target pest populations [20]. This technique has been applied in mosquito control. In rice planthoppers, wStri from L. striatellus has been successfully transfected into N. lugens, resulting in cytoplasmic incompatibility in the new host and the inhibition of rice ragged stunt virus infection and transmission, with significant pest control potential [21, 22]. Given the considerable agricultural damage caused by these planthoppers and the close relationship between their Wolbachia strains, cross-infection and colonization of different Wolbachia strains across species are possible. This presents an ideal model system for investigating the mechanisms through which Wolbachia regulates host reproduction.
Bacterial factors associated with Wolbachia-induced cytoplasmic incompatibility
Wolbachia is transmitted mainly via maternal transmission. At the cytological level, although mature sperm do not contain Wolbachia, Wolbachia modifies or transforms sperm cells during the early stages of sperm maturation. This results in Wolbachia-infected (WI) males mating with Wolbachia-uninfected (WU) females. The resulting CI offspring, fertilized eggs, cannot effectively separate paternal chromosomes during cell division, forming a chromosome bridge and leading to embryonic death. However, Wolbachia within the eggs of WI females can "rescue" the paternal chromosomes, synchronizing the separation of maternal chromosomes and allowing normal embryonic development, a phenomenon known as “rescue” [23].
Because Wolbachia cannot be cultured freely in vitro, the key bacterial factors that induce CI have always been a challenge in studying its mechanisms. In recent years, with the development of sequencing technologies and advancements in research methods, the molecular mechanisms of CI have gradually emerged [8, 24]. Early studies revealed that the strength of the CI of Culex mosquitoes induced by Wolbachia was related to the genes encoding ankyrin (ANK) present in different strains [25]. In recent years, scientists have discovered that the proteins CifA (CI factor A, CidA or CinA) and CifB (CI factor B, CidB or CinB), which are encoded by two adjacent genes, may be key factors in the induction of host CI by Wolbachia. When WU male transgenic D. melanogaster expressing CifB and CifA mate with WU females, fertilized egg offspring exhibit the CI phenotype, and the embryos die. However, CifA and CifB can interact with each other. When WI male Drosophila infected with Wolbachia (wMel strain) mate with WU females expressing CifA, fertilized egg offspring can develop normally, resulting in a "rescue" phenomenon [26–28]. Mutations in the PD-(D/E)XK nuclease, deubiquitinase DUB and CifA peptidase domains of CifB affect the function of CI factors [27, 29–31], indicating that the integrity of a specific domain is important for the normal function of CI. The three-dimensional structures of CifA and CifB have been resolved, and the interaction between the two proteins is related to charge attraction at three interfacial regions [32, 33].
Although several studies have reported factors that induce CI in Wolbachia strains, Cif gene diversity is high, and evolutionary patterns are diverse [34]. The mechanism of CI is very complicated (Fig. 3) [35]. Currently, two models are used to explain the mechanism of CI, namely, the toxin-antidote model [36, 37] and the host modification model [38, 39].Fig. 3. Two proposed mechanistic models for Wolbachia-mediated CI [35]
The toxin–antidote model suggests that CifB is transferred to the embryo, where it is toxic. In the embryo, Wolbachia synthesizes CifA, which binds to CifB and neutralizes its toxic effects, thereby rescuing embryonic lethality. Both wPip Cids are localized in developing spermatids until the histone-to-protamine transition, after which only CidB remains in the mature sperm. Additionally, CidB induces apoptosis in Drosophila S2R + cells, an effect that can be rescued by co-expressing CidA. CidB likely closely associates with paternal DNA to inhibit DNA replication, with replication resuming upon CidA binding to CidB [37].
The host modification model proposes that Cif proteins modify specific targets in gametes: sperm modifications induce CI, whereas female modifications rescue embryos by reversing these alterations to ensure normal hatching. Since CifB-mediated modifications occur only in developing sperm, CI neither requires CifB sperm transport nor CifA-CifB physical interaction [38]. For example, in D. melanogaster infected with the Wolbachia wMel strain, CifA and CifB were detected in the sperm cell nucleus during spermatogenesis and sperm formation—this localization results in histone retention in elongating spermatids but protamine deficiency in mature sperm. In contrast, CifA was detected only in early-stage oocytes and was undetectable in mature oocytes or embryos, suggesting that CifA lacks the opportunity to bind CifB after fertilization for detoxification and is more likely to modify gamete DNA during gametogenesis. A similar Cif protein localization pattern has been confirmed in Aedes aegypti [39]. Functional studies further revealed that CifA, as a ribonuclease (RNase), degrades long non-coding RNAs (lncRNAs) involved in the histone-to-protamine transition during sperm formation. Moreover, both CifA and CifB act as deoxyribonucleases (DNases), exacerbating DNA damage in the late stages of sperm formation [40]. These molecular mechanistic findings collectively support the core hypotheses of the host modification model.
Most studies on the CI mechanism are still focused on mosquitoes and fruit flies, with some experimental results differing greatly between the two insects of the Diptera order [35, 41]. For example, CifA and CifB of wMel from D. melanogaster need to be expressed simultaneously to induce the death of CI embryos [26, 40]. However, the expression of a single CifB protein from wPip of Culex mosquitoes and* w*No of Drosophila was sufficient to induce CI [36, 42, 43], indicating that the functions of CI factors differ among different Wolbachia strains in different hosts.
The Wolbachia wStri strain induces complete cytoplasmic incompatibility (CI) in its native host, L. striatellus [14]. When N. lugens is transfected into a new host, N. lugens, wStri’s tissue distribution mirrors that of its native host, but its density is significantly higher than that of the wLug strain. Although wStri still induces CI in N. lugens, the intensity of CI is notably reduced [44]. These observations suggest that the strength of CI is influenced by both the symbiont and the host. Genomic analysis revealed that the wStri genome contains three copies of the cifA-cifB genes, which are likely crucial for CI induction. Phylogenetic analysis revealed that the CI factor groups expanded from 5–10 groups, with wStri factors belonging to newly identified groups VI and VIII [45, 46]. The functions of these CI factor candidate proteins remain unknown.
Host factors associated with Wolbachia-induced cytoplasmic incompatibility
In host insects, studies in D. melanogaster have revealed that karyopherin-a, the protamine histidine chaperone P32, and ubiquitin-conjugating enzymes can interact with CifB, influencing CI lethality [47, 48]. The male histone-regulated gene Hira, the acyl-CoA-binding protein Acbp2 and the mitochondrial membrane protein Mcad may be involved in CI caused by Wolbachia infection [49, 50], but whether these proteins directly interact with CifA and CifB remains unclear. The modes of action of CI in different insects may differ. Therefore, the study of the CI phenomenon of Wolbachia in other organisms is particularly important for clarifying the action pattern of CI.
With the advantages of proteomics and RNAi technologies, Huang et al. reported that the cytoplasmic aminopeptidase-like protein (CAL) of L. striatellus might be associated with Wolbachia-induced CI death [51]. RNA-seq analysis revealed that iLvE, a gene involved in branched-chain amino acid (BCAA) biosynthesis, was the most significantly downregulated gene in WI L. striatellus. RNAi-mediated knockdown of iLvE in WU male L. striatellus resulted in reduced fertility and lower embryo hatching rates. However, fertility was restored when these males were crossed with Wolbachia-infected females, suggesting that iLvE may also contribute to the reproductive dysfunction associated with CI [52]. Another gene potentially involved in Wolbachia-induced CI in L. striatellus is NDUFA8, which encodes a subunit of the NADH dehydrogenase [ubiquinone] 1 α subcomplex. In WI females, the mRNA expression of NDUFA8 was upregulated. RNAi-mediated suppression of NDUFA8 significantly increased early embryo mortality without affecting the number of eggs laid. Furthermore, treatment with dsNDUFA8 in WI females reproduced CI-like symptoms, suggesting that NDUFA8 plays a key role in the rescue phenotype associated with CI [53]. These findings indicate that the interaction mechanisms between CI factors and the host may differ across different species. However, whether these proteins directly interact with CifA or CifB remains unclear. Moreover, functional validation of candidate CI and fecundity-related genes in planthoppers via techniques such as CRISPR/Cas9 or transgenesis would provide the most critical evidence.
Mechanisms by which Wolbachia regulates host fecundity
Wolbachia infection can directly increase the fecundity of host insects. In Aedes albopictus, females infected with Wolbachia laid greater numbers of eggs [54]. Wolbachia is essential for host egg formation in Asobara tabida [55]. In D. melanogaster, Wolbachia manipulates the self-renewal and differentiation of reproductive stem cells to increase the fertility of the host [56]. Wolbachia can also provide nutrients to help the fruit fly D. innubila produce more female offspring under nutrient deficiency conditions[57]. The rice planthopper is a hemipteran insect that has undergone gradual metamorphosis and has significant biological differences from holometamorphic mosquitos, flies and wasps. The study of the function and mechanism of Wolbachia in other types of organisms is important for elucidating the interaction between Wolbachia and the host.
Wolbachia can also impact the fecundity of rice planthoppers. In N. lugens, the removal of Wolbachia results in a significant reduction in oviposition days and a decrease in the fecundity of female adults [58]. Similarly, wStri significantly promotes the fecundity of L. striatellus. A previous study utilizing comparative genomics and metabolomics technologies reported that both wStri and wLug possess the complete ability to synthesize members of the vitamin B family, including biotin and riboflavin. These nutrients play crucial roles in maintaining the fecundity of both L. striatellus and N. lugens [59]. Wolbachia-provided riboflavin and biotin have been demonstrated to increase the fitness of the bed bug Cimex lectularius [60]. Moreover, biotin operons are found in Wolbachia genomes scattered across four supergroups [59, 61, 62], supporting the notion that the biotin operons were possibly laterally transferred from another organism.
At the host level, the increase in egg laying in the planthopper caused by Wolbachia was related to the upregulation of the expression of vitellogenin Vg, the increase in germ cell mitosis, and the induction of apoptosis in ovarian trophoblasts [63–65]. Previous studies suggest that Wolbachia accelerates nurse cells in female L. striatellus during peak oviposition, thereby providing more ovarian nutrients and increasing egg production during this period [65]. Wolbachia also affects miRNA expression in L. striatellus, thereby altering the expression of genes related to fecundity [66]. Recently, we reported that Wolbachia infection caused the differential expression of mitochondrial-related genes in planthoppers, suggesting that energy metabolism may also be related to Wolbachia’s improvement in the fecundity of planthoppers (unpublished data), but the underlying mechanisms remain to be further studied.
In S. furcifera, Wolbachia may interact with another intracellular endosymbiont, Cardinium. Different endosymbiont-infected lines present distinct microbiomes and metabolite levels, which influence their fecundity. Notably, Cardinium and Wolbachia double-infected lines present lower fecundity than uninfected lines do [67]. These findings suggest that the combined effects of Wolbachia and other symbionts may have a more pronounced impact on the host's reproductive capacity than either symbiont alone. Future research could explore the specific mechanisms underlying these interactions and their potential for manipulating pest populations through biological control strategies.
Perspective
Wolbachia-mediated reproductive manipulation, particularly CI and fecundity enhancement, plays a pivotal role in shaping the population dynamics of rice planthoppers—among the most destructive pests in global rice production. The differential infection patterns, CI induction capacities, and reproductive benefits conferred by Wolbachia strains across N. lugens, L. striatellus, and S. furcifera highlight the complexity of symbiont–host interactions in hemipteran insects. Advances in comparative genomics, functional genetics, and proteomics have begun to unravel the molecular underpinnings of CI, identifying key effectors such as CifA and CifB and revealing potential host factors involved in modification and rescue pathways. Moreover, the role of Wolbachia in provisioning essential nutrients such as B vitamins and modulating host reproductive physiology offers new insights into mutualistic mechanisms that increase pest fitness. These findings not only deepen our understanding of insect–microbe coevolution but also open promising avenues for symbiont-based pest management strategies. Future research should focus on elucidating the host-specific determinants of CI strength, characterizing novel CI factor variants, and exploring the feasibility of leveraging Wolbachia for population suppression or replacement in rice planthopper control programs. Integrating Wolbachia biology into integrated pest management frameworks may offer sustainable, environmentally friendly solutions to mitigate the growing threat of planthopper outbreaks in rice agroecosystems. However, it is also essential to consider potential challenges in deploying Wolbachia-based control strategies in the field, such as the evolution of resistance or possible ecological side effects. However, the potential horizontal transmission of Wolbachia to non-target hosts must also be considered during risk assessments to avoid unintended consequences. In conclusion, while Wolbachia offers a promising avenue for controlling rice planthopper populations, the complexity of its interactions with both the host and the environment warrants careful consideration and further investigation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sinkins SP, Walker T, Lynd AR, Steven AR, Makepeace BL, J. Godfray HC, et al. Wolbachia variability and host effects on crossing type in Culex mosquitoes. Nature. 2005;436:257–60. 10.1038/nature 03629. https://www.nature.com/articles/nature 03629#supplementary-information.10.1038/nature 0362916015330 · doi ↗ · pubmed ↗
- 2Namias A, Martinez J, Boussou I, Terretaz K, Conner W, Justy F, et al. Recombination, truncation and horizontal transfer shape the diversity of cytoplasmic incompatibility patterns. bio Rxiv. 2025:2025.01.06.631454. 10.1101/2025.01.06.631454.10.1093/molbev/msaf 200PMC 1246156340796352 · doi ↗ · pubmed ↗
- 3Sun G, Zhang M, Chen H, Hochstrasser M, Cossart PF. The Cin B nuclease from w No Wolbachia is sufficient for induction of cytoplasmic incompatibility in Drosophila. m Bio. 2022;13(1):e 03177–21. 10.1128/mbio.03177-21.10.1128/mbio.03177-21PMC 878749035073749 · doi ↗ · pubmed ↗