Engineering strigolactone signaling: toward crops that resist parasites without sacrificing symbiosis
Zeming Huang, Fan Qi

Abstract
Click any figure to enlarge with its caption.
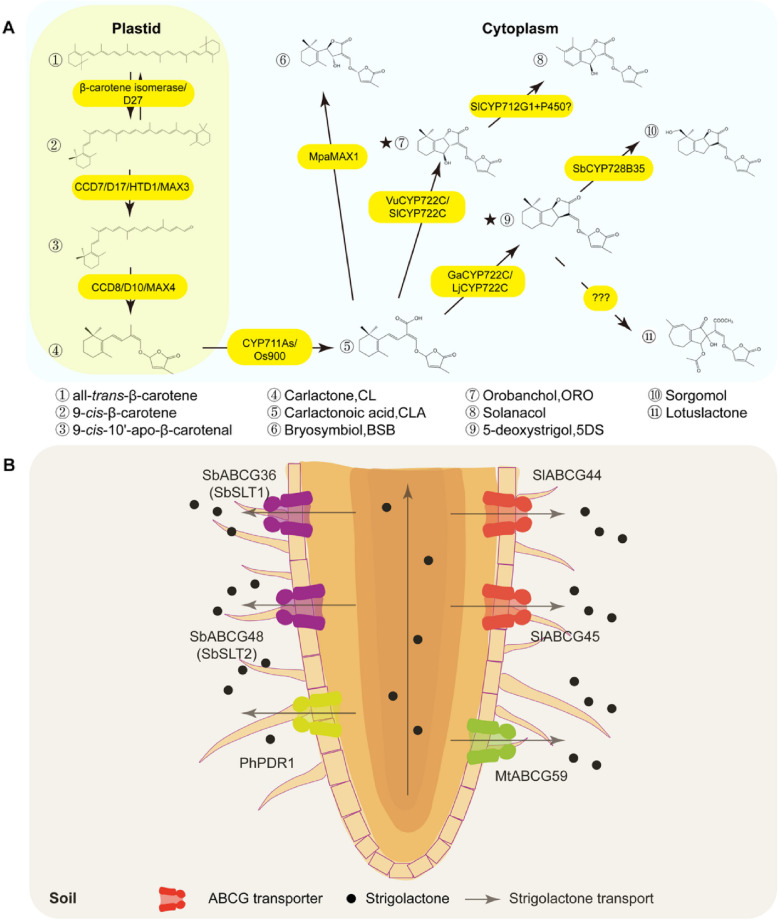 Figure 1
Figure 1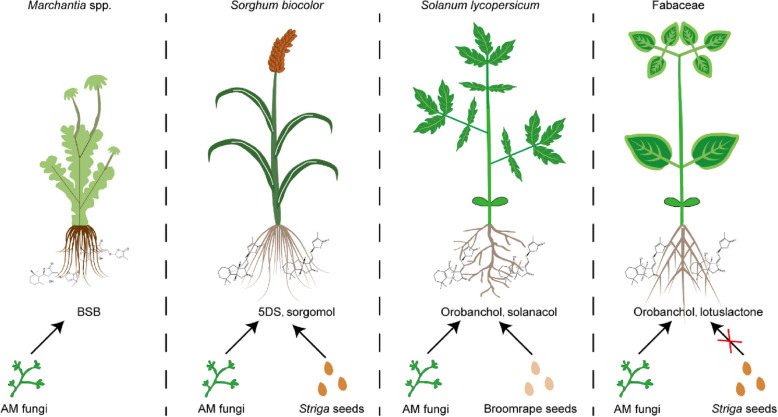 Figure 2
Figure 2- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
- —http://dx.doi.org/10.13039/501100002858China Postdoctoral Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Parasitism and Resistance · Plant and animal studies · Plant Molecular Biology Research
Strigolactones (SLs) constitute a class of carotenoid-derived phytohormones that orchestrate developmental plasticity and adaptive responses to environmental cues in plants [1]. The biosynthetic cascade of SLs is initiated by the stereoselective isomerization of all-trans-β-carotene to 9-cis-β-carotene, which is catalyzed by the isomerase DWARF27 (D27). This cis-configured intermediate undergoes sequential oxidative cleavage: first, carotenoid cleavage dioxygenase 7 (CCD7) catalyzes the stereo-specific cleavage of 9-cis-β-carotene to generate 9-cis-β-apo-10’-carotenal, which is subsequently remodeled by CCD8 to yield carlactone (CL), the central precursor of SLs [2]. The committed step in SL diversification involves cytochrome P450-mediated oxidation. Members of the cytochrome P450 family 711 subfamily A (CYP711A) oxidize CL to produce carlactonoic acid (CLA), a pivotal branch-point metabolite [2]. Divergent evolutionary trajectories within the P450 superfamily govern subsequent modifications. In Marchantia paleacea, the cytochrome P450 enzyme MORE AXILLARY GROWTH 1 (MAX1) catalyzes the conversion of CLA to bryosymbiol (BSB) [3], while in Vigna unguiculata and Solanum lycopersicum, enzymes from the CYP722C subfamily catalyze the conversion of CLA to the canonical SL orobanchol. The CYP712G1 enzyme then converts orobanchol to solanacol, the predominant SL in tomato root exudates [4]. Furthermore, some CYP722C-subfamily enzymes mediate the conversion of CLA to the strigol-type SL 5-deoxystrigol (5DS). In Sorghum bicolor, CYP728B35 subsequently converts 5DS to sorgomol [5]. Legumes may employ distinct enzymes to convert CLA into non-canonical SLs, including lotuslactone and medical compounds [6] (Fig. 1A).Fig. 1. Schematic diagram of strigolactones (SLs) synthetic and transport pathways. A Summary of SLs biosynthesis pathways among different plants. SLs molecules are numbered with serial identifiers, and enzymes involved in SLs biosynthesis are highlighted with a yellow background. Canonical SLs are marked with asterisks. B Summary of the molecular mechanisms of SLs transport in different plants and fields. Abbreviations: Os, Oryza sativa; Vu, Vigna unguiculata; Sl, Solanum lycopersicum; Sb, Sorghum bicolor; Lj, Lotus japonicus; Mt, Medicago truncatula;Mpa, Marchantia paleacea; Ga, Gossypium arboreum; Ph, Petunia hybrida. Enzyme ID: D27, C7AU21; CCD7、MAX3, Q7XJM2; CCD8、MAX4, Q8VY26; CYP711A2、Os900, M9R6D3; MpaMAX1, A0A8D4XJ04; VuCYP722C, Vigun03g264300; SlCYP722C, A0A3Q7F9H0; GaCYP722C, LC528626; LjCYP722C, A0A6F8PJQ1; SlCYP712G1, A0A3Q7IDV9; SbCYP728B35, Sb08g017540
The co-option of SL signaling by parasitic organisms is a byproduct of the synergistic co-evolution between plants and arbuscular mycorrhizal (AM) symbiosis. In the ancient bryophyte M. paleacea, secreted SLs, particularly BSB, function as phytohormones that regulate growth and development while also serving as rhizospheric signaling molecules that induce AM fungal symbiosis [3]. Recently, Wang’s group reported that M. paleacea ccd8a/8b mutants, which are deficient in SLs, presented significantly impaired AM colonization. This impairment could be partially rescued by exogenous application of GR24 (a synthetic SL analog) or root exudates from phosphorus-deficient plants, which contain endogenous SLs [7].
Divergent evolution in vascular plants has led to the structural diversification of SLs. Monocots, such as sorghum, predominantly secrete non-hydroxylated 5DS to optimize AM symbiosis under low-phosphorus conditions. However, this strategy inadvertently enhances susceptibility to Striga [8]. In contrast, plants within the Fabaceae family preferentially synthesize hydroxylated SLs, including orobanchol and lotuslactone [9]. This specialized profile of SLs diminishes the recognition of Striga while maintaining effective AM symbiosis, thereby representing an evolutionary adaptation that balances symbiotic benefits and parasitic defenses (Fig. 2).Fig. 2. Proposed models of strigolactone (SL) signaling mechanisms in Marchantia paleacea, Sorghum bicolor, Solanum lycopersicum and Fabaceae. In M. paleacea, root-excreted SLs predominantly consist of bryosymbols, which function as rhizosphere signaling molecules to promote arbuscular mycorrhizal (AM) fungal symbiosis [3]. S. bicolor primarily secretes 5DS under low-phosphorus conditions. While this SL enhances AM fungal colonization, it is paradoxically exploited by parasitic Striga seeds to stimulate germination [10]. S. lycopersicum primarily secretes orobanchol and solanacol under low-phosphorus conditions. While this SL enhances AM fungal colonization, it is paradoxically exploited by parasitic broomrape seeds to stimulate germination [11]. Fabaceae species predominantly produce orobanchol and noncanonical SLs through root exudation, and these compounds effectively promote AM fungal symbiosis while reducing susceptibility to parasitic plant exploitation [6]. Abbreviations: SL, strigolactone; BSB, bryosymbiol; 5DS, 5-deoxystrigol; AM, arbuscular mycorrhizal
This raises critical questions: do hydroxylation modifications in SLs determine their preferential recognition by AM fungi versus parasitic plants? How can SL transporter-targeted crop systems be engineered to spatially restrict SL secretion (e.g., root tip-specific expression) for precise suppression of Striga germination while preserving AM symbiosis in nutrient-deprived zones?
In addition to the biosynthesis of SLs, their secretion into the rhizosphere is pivotal for inducing AM symbiosis and the germination of parasitic plant seeds. In 2012, the ABC transporter G (ABCG) gene PLEIOTROPIC DRUG RESISTANCE 1 (PDR1) in Petunia hybrida was identified as the first SL transporter gene [12]. The functions of increasing numbers of SLs transporters, such as those in Medicago truncatula [13], S. bicolor [10] and S. lycopersicum [11], are being progressively elucidated (Fig. 1B).
In a breakthrough study, Shi et al. [10] identified two ABCG transporter genes in sorghum (S. bicolor), sorghum SL transporter 1 (SbSLT1) and SL transporter 2 (SbSLT2), which are responsible for exporting SLs from roots into the rhizosphere. These transporters were induced under low phosphorus conditions or GR24 treatment, as confirmed by transcriptomic and functional analyses. Using AlphaFold2, researchers have predicted the protein structures of SbSLT1 and SbSLT2 and validated F693 in SbSLT1 and F642 in SbSLT2 as key binding sites for SLs. CRISPR/Cas9-mediated knockout of these genes disrupted SLs efflux into the rhizosphere, leading to 67–94% suppression of Striga seed germination and parasitism in sorghum, thereby stabilizing crop yield. In field trials under Striga-free conditions, single (SbSLT1^ko^, SbSLT2^ko^) and double knockout (SbSLT1^ko^SbSLT2^ko^) mutants presented no growth defects compared with wild-type plants. However, in Striga-infested fields, the mutants presented fewer parasitic plants and significantly lower yield losses than did the wild type. Notably, the mutants presented greater aboveground biomass (fresh and dry weights) than did the wild type, which was attributed to prolonged leaf greenness and increased tillering. These results demonstrate that targeting SbSLT1/SbSLT2 effectively controls Striga parasitism in agricultural settings, suggesting a promising strategy to mitigate crop losses caused by parasitic weeds.
Furthermore, Ban et al. [11] identified the ABCG transporter genes SlABCG44 and SlABCG45 in S. lycopersicum, which are involved in the exudation and upward translocation of the SLs orobanchol and solanacol within tomato plants. Both pot experiments and field trials consistently demonstrated that single knockout mutants (Slabcg45^ko^ and Slabcg44^ko^) presented a significant reduction in parasitic infestation by Egyptian broomrape. Additionally, the yields of the two Slabcg45^ko^ mutants increased by more than 30% compared with those of the control group, underscoring the substantial potential application of SlABCG45 in the breeding of parasitic-resistant tomato varieties.
These studies reveal the central roles of SL transporters in rhizospheric SLs secretion, providing novel targets for precision editing to confer plant resistance against Striga (witchweed) and broomrape. By integrating AI-predicted evolutionarily conserved SLs transporter motifs, this strategy can be extended to major crops through homology-directed CRISPR editing, enabling tissue-specific SLs secretion control to balance parasite defense and symbiotic efficiency while minimizing off-target effects on root microbiome assembly.
The evolutionary arms race between parasitic plants and their hosts has driven co-adaptive innovations in SLs perception systems. Emerging evidence has demonstrated that noncanonical SLs derivatives in leguminous species are highly distinctive, enabling plants to maintain high efficacy in AM symbiosis while simultaneously evading detection by parasitic plant germination receptors. This structural divergence between canonical and noncanonical SLs variants likely constitutes an evolutionary countermeasure against parasitic infestation. Deciphering the molecular mechanisms underlying SLs biosynthesis pathway diversification and spatiotemporal transport regulation will not only resolve long-standing questions about functional trade-offs in plant chemical ecology but also establish a mechanistic framework for engineering SLs-mediated rhizosphere communication in agricultural ecosystems. Such insights could revolutionize crop improvement strategies by leveraging natural evolutionary solutions to optimize symbiotic partnerships while minimizing parasitic vulnerability.