Functional diversity of the Osiris gene family in the brown planthopper
Cui Zhang, Xinyi He, Ya Ma, Yaxin Liu, Xingxing Shen, Yanyuan Bao

TL;DR
This study explores the Osiris gene family in brown planthoppers, revealing their roles in development and reproduction.
Contribution
The study identifies and functionally characterizes Osiris genes in a hemimetabolous insect for the first time.
Findings
Knockdown of Osiris genes caused lethal phenotypes and developmental defects in brown planthoppers.
Osiris genes showed tissue-specific expression patterns in brown planthoppers.
RNAi experiments revealed roles in wing, ovary, and ovipositor development.
Abstract
Osiris is a large insect-specific gene family with multiple functions. Owing to highly conserved sequences and similar expression patterns across insect species, the Osiris gene family is believed to play specific roles in the insect kingdom. However, current studies on Osiris gene functions have focused primarily on several holometabolous insects, such as Drosophila melanogaster and Bombyx mori, with few systematic studies on hemimetabolous insects. In this study, we identified 20 Osiris genes by searching the genome and transcriptome databases of the brown planthopper Nilaparvata lugens. The Nilaparvata lugens Osiris genes presented similar development- and tissue-specific expression profiles. A functional investigation using RNAi in 4th instar nymphs and newly eclosed adult females revealed that the knockdown of Osiris genes resulted in multiple phenotypes, including a lethal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1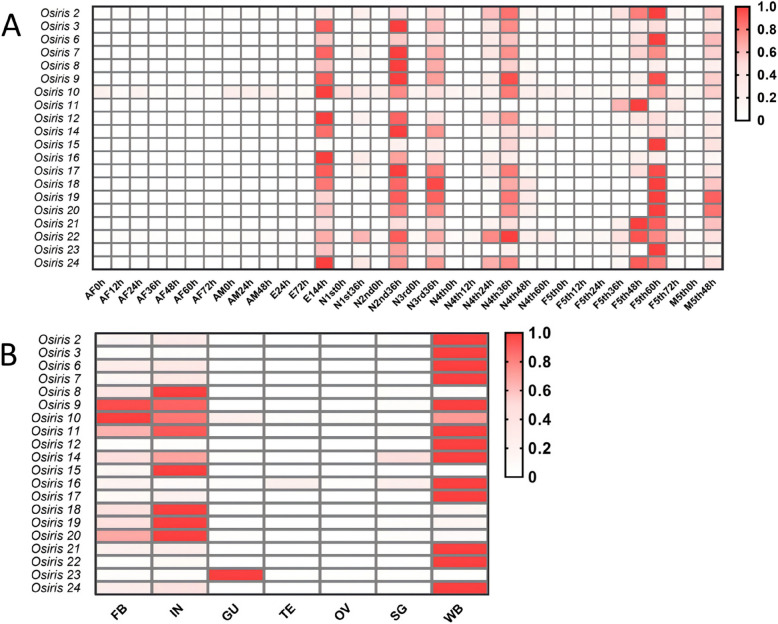 Figure 2
Figure 2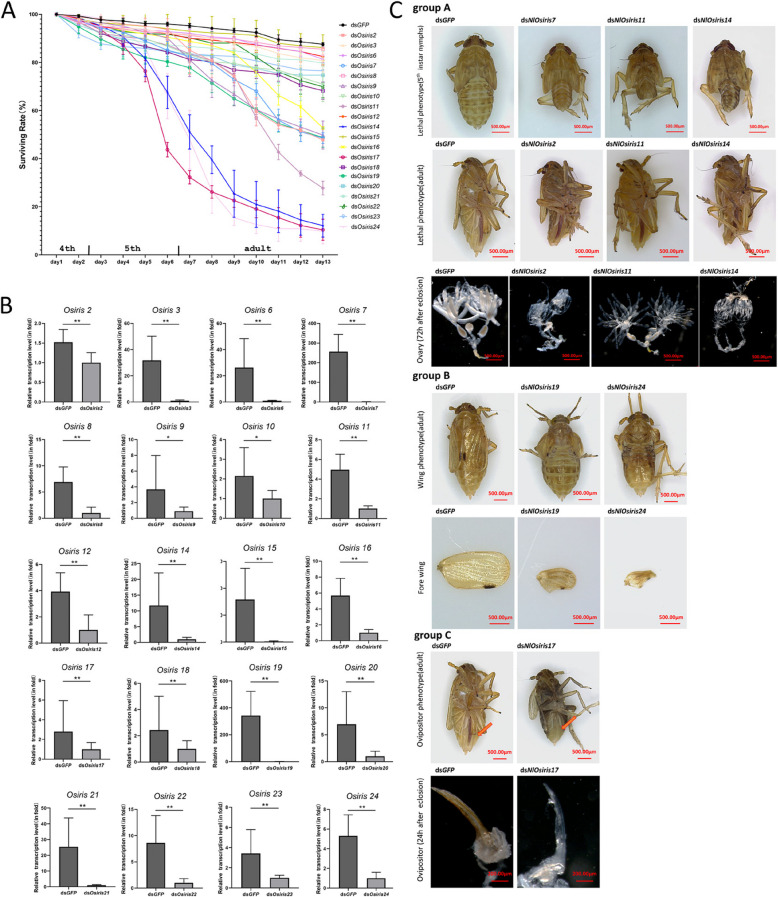 Figure 3
Figure 3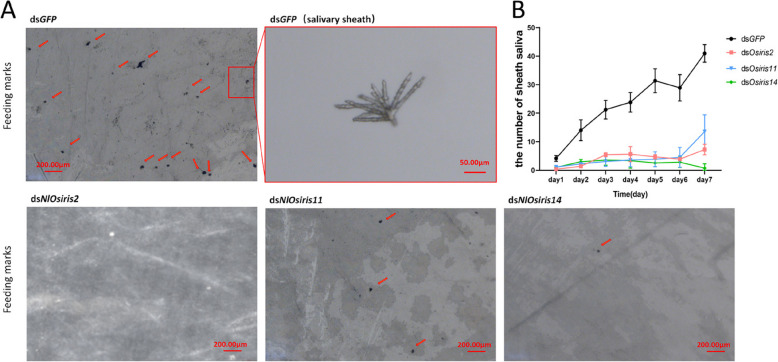 Figure 4
Figure 4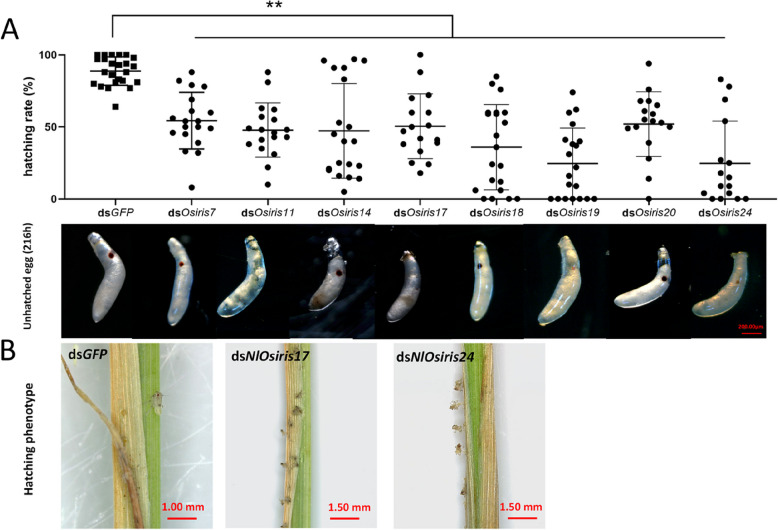 Figure 5
Figure 5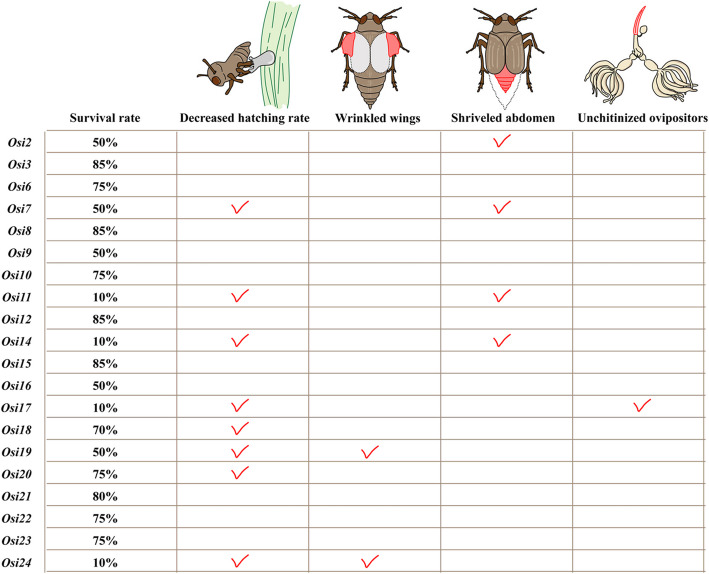 Figure 6
Figure 6- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
- —http://dx.doi.org/10.13039/501100012165Key Technologies Research and Development Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Photosynthetic Processes and Mechanisms · Plant Reproductive Biology
Introduction
The Osiris gene family was first described in Drosophila melanogaster [1]. Twenty-five Osiris genes have been identified in the Drosophila genome (FlyBase) [2]. Among these genes, 22 clustered within the dosage‐sensitive Triplo-lethal (Tpl) locus in the chromosomal region 83D-E. The other three genes (Osiris21, 22 and 23) are outside of this cluster and are located at the 32E, 99F and 87E sites in the genome [1]. The Osiris proteins are characterized by a DUF1676 domain (Pfam: PF07898) with unknown function [3]. In addition, they feature an N-terminal signal peptide, a pair of conserved cysteine residues, a transmembrane region and a C-terminal AQXLAY motif [4].
The Osiris genes are present only in insects, from the basal groups of mayflies and silverfish to highly evolved dipterans [4–6]. No obvious homologs have been reported in the genomes of non-insect Arthropoda or other invertebrates, including crustaceans, Myriapod, Chelicerata, and Entognatha (Collembola) [6–8]. A few studies, primarily in the fruit fly D. melanogaster and the silkworm B. mori, have addressed the specific functions of the Osiris genes [7–11]. In D. melanogaster, of the 25 Osiris genes, 16 were expressed in cuticle-secreting epidermal and sensory organ cells, and 4 were required for specific cuticle nanostructures [6]. The embryonic lethal Osiris6 and Osiris7 mutants presented strong defects in larval cuticle formation [7]. Osiris6, Osiris7 and Osiris8 are involved in resistance to octanoic acid (OA) [10, 12–14], a remarkable plant toxin to Drosophila species, with the exception of Drosophila sechellia, which is a dietary specialist on the host plant Morinda citrifolia and has evolved resistance to fruit OA [10]. Osiris8 is highly expressed in the tormogen support cells of the antenna trichoid sensilla and is required for pheromone detection [9]. Osiris9, Osiris15 and Osiris19 are highly expressed in the trachea and function redundantly to regulate tracheal tube maturation [8]. Osiris17 is required in the wing epithelium to produce expanded wings [6]. Osiris21 is involved in endolysosomal trafficking in photoreceptor neurons of the eye [15], suggesting that it plays a role in regulating the cellular signaling pathways involved in vesicular trafficking and protein sorting within this system. Osiris23 is essential for the formation of nanopores lining the olfactory sensillum [7]. In B. mori, 25 Osiris genes have been identified, and most of these genes are specifically expressed in the wings or epidermis [16]. In particular, the Osiris9a gene in the silkworm Osiris family is specifically expressed in the silk gland and contributes to the formation of silk fibers [17]. The understanding of the functions of Osiris, an insect-specific gene family, has been limited in the above two model insect species. Although a recent study reported that Osiris17 contributes to morphogenesis of the intestinal tract in the hemimetabolous insect Locusta migratoria [18], the physiological functions of this gene family in most insect species are still largely unknown. Although some genes with functions of both chitin-binding and peptidase activity are coexpressed with the Osiris family [11], the precise interaction mechanism remains elusive. To better understand the functional roles of the Osiris gene family, in this study, we aimed to study a Hemipteran insect, the brown planthopper (Nilaparvata lugens Stål), to conduct a comprehensive analysis of the expression and functions of the Osiris genes. Nilaparvata lugens is an important hemimetabolous rice pest that causes severe damage to rice by sucking rice phloem sap [19]. This insect species has emerged as the ideal model system for studying gene functions because the whole-genome sequence has been elucidated [20], and the susceptibility to RNA interference (RNAi) [21–26].
In this study, we identified 20 Osiris family genes by searching the N. lugens genome and transcriptome databases and analyzed their phylogenetic relationships, spatiotemporal expression patterns, and gene functions via RNA interference (RNAi). The knockdown of the Osiris genes resulted in various phenotypes, including lethality, wing and ovipositor deficiency, hatching and feeding failure. To our knowledge, this is the first report on the reproductive and feeding functions of Osiris genes. Our findings increase the understanding of the physiological functions of Osiris genes in development and reproduction. This gene family could be a potential target for pest management to develop biological control strategies involving genetic modification to reduce insecticide usage.
Materials and methods
Insects and plants
Brown planthopper populations were originally collected from a rice field located at the Huajiachi Campus of Zhejiang University, Hangzhou, China, in 2008. The insect strain was routinely reared on rice seedlings (Oryza sativa strain TN1) at a temperature of 26 ± 0.5 °C with 50 ± 5% relative humidity under a 16:8 h light:dark photoperiod as previously described [21].
Identification of the N. lugensOsiris gene family
The Osiris gene sequences were obtained from the N. lugens genome (GenBank accession number AOSB00000000 under BioProject PRJNA177647) and transcriptome (accession number SRX023419) databases via the BLASTX algorithm with a significance cutoff of E value < 1×10^−5^. The Osiris sequences were further verified and annotated via the National Center for Biotechnology Information (NCBI), Conserved Domains Database (CDD) (https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml) and SwissProt database (https://www.sib.swiss/swiss-prot). The open reading frames (ORFs) and amino acid sequences were predicted via Open Reading Frame Finder (https://www.ncbi.nlm.nih.gov/orffinder/). The signal peptide and characteristic domain were predicted via the Simple Modular Architecture Research Tool (SMART) (https://smart.embl.de/).
Phylogenetic tree construction
To understand the phylogenetic relationship, the Osiris sequences were obtained from 23 insect species in 8 orders, including Odonata: Ischnura elegans; Orthoptera: Schistocerca americana; Hemiptera: Laodelphax striatellus, N. lugens, Acyrthosiphon pisum, Rhopalosiphum maidis; Hymenoptera: Nasonia vitripennis, Vespula pensylvanica, Apis mellifera, Ooceraea biroi; Coleoptera: Agriotes lineatus, Tribolium castaneum, Diorhabda carinulata, Brassicogethes aeneus; Lepidoptera: Pararge aegeria, Pectinophora gossypiella, Trichoplusia ni, B. mori; Neuroptera: Chrysoperla carnea; Diptera: D. melanogaster, Anopheles merus, Aedes aegypti, Hermetia illucens. The Osiris sequences of these insect species were downloaded from the NCBI website (https://www.ncbi.nlm.nih.gov/), FlyBase Drosophila database (https://flybase.org/) and KAIKO (https://kaikobase.dna.affrc.go.jp/) by using D. melanogaster and B. mori Osiris sequences as queries to search the homologs of the other insect species via BLAST v2.12.0 (http://ftp.ncbi.nlm.nih.gov/blast/executables/blast+/2.12.0) at an E-value threshold of 1×10^−5^. The predicted Osiris genes were filtered on the basis of an identity threshold of ≥ 30%. InterProScan v5.66 (https://ftp.ebi.ac.uk/pub/software/unix/iprscan/5/5.66-98.0/interproscan-5.66-98.0-64-bit.tar.gz) [27] was used to determine the reliability of the homologous Osiris proteins across different insect species. Given the intrinsic homology among genes within the Osiris family, we employed IQ-TREE v2.3.3 (http://iqtree.cibiv.univie.ac.at/) to construct a maximum likelihood (ML) tree of the candidate Osiris protein sequence alignment under the Q. insect + F + R10 model [28, 29]. The reliability of the ML phylogeny was assessed via a bootstrap analysis of 1000 replications; bootstrap values N50% are shown on each node of the tree. The phylogenetic tree was visualized via iTOL v5 (https://itol.embl.de) [28].
Spatiotemporal expression patterns of NlOsiris genes
The spatiotemporal expression profiles of NlOsiris genes throughout the developmental stages and in various tissues were investigated by searching the N. lugens transcriptomic database (GenBank accession no. PRJNA714229) as described previously [30–32]. To verify the expression profiles, quantitative real-time PCR (qRT‒PCR) was conducted. Total RNA was extracted from each developmental stage and each tissue via a TRIzol Total RNA Isolation Kit (TaKaRa, Dalian) according to the manufacturer's instructions. The RNA concentrations were determined via a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific). For developmental stage-specific expression analysis, total RNA was extracted from the eggs in the rice leaf sheaths at 24, 72 and 144 h (n = 100) after laying; the whole bodies of the 1st-instar nymphs at 0 and 36 h after hatching (n = 100); the 2nd-instar nymphs at 0 and 36 h after molting (n = 80); the 3rd-instar nymphs at 0 and 36 h (n = 50); the 4th-instar nymphs at 0, 12, 24, 36, 48 and 60 h (n = 40); the 5th-instar female nymphs at 0, 12, 24, 36, 48, 60 and 72 h (n = 40); the 5th-instar male nymphs at 0 and 48 h (n = 40); the female adults at 0, 12, 24, 36, 48, 60 and 72 h (n = 25); and the male adults at 0, 24 and 48 h after eclosion (n = 25). For tissue-specific expression analysis, total RNA was extracted from the fat body (n = 100), integument (n = 80), wing bud (n = 80), gut (n = 100), and salivary gland (n = 200) of 5th-instar nymphs spanning early to late developmental stages and from the ovary (n = 50) and testis (n = 50) of female and male adults at 24–72 h post-eclosion. Prior to dissection, insects were anesthetized on ice and carefully dissected under a Leica S8AP0 stereomicroscope (Leica Microsystems GmbH, Wetzlar, Germany) with fine forceps. The isolated tissues were gently washed in phosphate-buffered saline (PBS) for RNA extraction. The total RNA of each sample served as a template for reverse transcription via a Hiscript® II QRT SuperMix for qPCR (+ gDNA wiper) Kit (Vazyme, Nanjing, China) to remove any contaminating genomic DNA. qRT‒PCR was performed on a CFX ConnectTM Real-Time System (Bio-Rad, Hercules, CA, USA) using ChamQ SYBR Color qPCR Master Mix (Vazyme, Nanjing, China) under the following reaction program: an initial denaturation step at 95 °C for 30 s, followed by 35 cycles at 95 °C for 5 s and 55 °C for 30 s. For each NlOsiris gene, specific primers for qRT‒PCR were designed via Primer Premier 6 (Premier, Biosoft, Canada) to amplify regions ranging from 150–250 bp (Table S1). The 18S ribosomal RNA gene (GenBank accession number JN662398.1) was used as an internal control. Relative expression levels were calculated via the 2^−ΔΔCt^ method [33]. Each sample was analyzed in triplicate. Developmental and tissue-specific expression heatmaps were generated via GraphPad Prism 8 (La Jolla, CA, USA).
RNA interference (RNAi)
RNAi was used to investigate the functions of the Osiris gene family in N. lugens. Each NlOsiris gene was amplified and cloned and inserted into the pMD-19 T vector (TaKaRa, Dalian, China). Double-stranded RNAs (dsRNAs) were synthesized with specific primers containing a T7 promoter in vitro through PCR-generated DNA templates via a T7 High Yield RNA Transcription Kit (Vazyme, Nanjing, China). The sequences of the NlOsiris genes used as templates for dsRNA synthesis ranged from 550–650 bp in length. dsGFP served as a negative control to assess nonspecific effects. The specific primers used for dsRNA synthesis are shown in Table S1.
For gene function analysis, the nymphs were anesthetized with carbon dioxide for 20 s, and then 10 µL of each dsRNA (5 ng/µL) was microinjected into the abdomen of 4th-instar nymphs (24–48 h after molting, n = 100) via a FemtoJet microinjection system (Eppendorf-Netheler-Hinz, Hamburg, Germany) [21]. For each treatment, three biological replicates were conducted. RNAi efficiency was assessed in 5th-instar nymphs at 48 h after molting. Phenotypes, including survival and any abnormally morphological phenotypes, were recorded daily. For reproduction studies, each newly emerged adult (0–2 h post-eclosion, n = 50) was anesthetized with carbon dioxide for 25 s and microinjected with 15 µL of each dsRNA (5 ng/µL). After injection, the insects were reared on fresh rice seedlings. Mating between one female and one male was performed 3 days after RNAi, and the females were removed from the rice seedlings on the 5th day after mating. The number of hatched nymphs was counted daily for 7 days. The seedlings were dissected to observe and calculate the unhatched fertilized and unfertilized eggs. RNAi efficiency in the eggs was assessed at 144 h post-laying within the rice seedlings via qRT‒PCR. Data analysis was performed via GraphPad Prism 8.0 (La Jolla, CA, USA). The data were analyzed via Student's t test, and the results are presented as the means ± standard deviations.
Feeding observation
The artificial D-97 diet was prepared according to methods described by Zu et al. [34]. The artificial diet was prepared under aseptic conditions, filtered through a 0.22 μm syringe filter (Millipore, MA, USA), and stored at -20℃. The 4th-instar nymphs were injected with dsOsiris RNAs and reared on fresh rice seedlings. On the 3rd day postinjection, the nymphs were transferred from the rice seedlings to glass tubes (length: 8 cm, diameter: 4.5 cm) and reared for 7 days. Both ends of a glass tube were sealed with two layers of Parafilm membranes (Pechiney Plastic Packaging Company, Chicago, IL, USA) with 40 μL of artificial diet in the space between the two layers of Parafilm. The glass tubes were placed at 26℃ ± 5℃, with a relative humidity of 50% ± 5% and a photoperiod of 16 h light and 8 h dark. Each glass tube contained 15 nymphs, with four biological replicates per treatment. The artificial diet was replaced every 24 h, and the number of surviving insects was calculated. The salivary sheaths were carefully separated from the inner layer of Parafilm via forceps under a stereomicroscope (Leica S8AP0, Wetzlar, Germany). The number of salivary sheaths was recorded. The data were analyzed via Student's t test, and the results are presented as the means ± standard deviations.
Results
Phylogenetic analysis of NlOsiris proteins
We identified 20 Osiris genes in N. lugens by searching genomic and transcriptomic databases (Table 1). The encoded proteins were validated through protein domain prediction (with a score < 0.001) (Table S2). We constructed a phylogenetic tree based on the complete amino acid sequences available in the GenBank database to understand the evolutionary relationships among NlOsiris and their homologs in other insect species. Phylogenetic analysis indicated that each NlOsiris protein is closely related to D. melanogaster and B. mori homologs (Fig. 1A). We follow the nomenclature strategy of D. melanogaster and B. mori Osiris genes to name the N. lugens homologs NlOsiris2-3, NlOsiris6-12 and NlOsiris14-24 on the basis of the the best match of the amino acid sequences with D. melanogaster and B. mori Osiris proteins. Although the E value of NlOsiris15 was > 1×10^−5^ according to the BLASTp algorithm, its evolutionary relationship suggests that it is a member of the Osiris family (Table S2). Table 1. Identification of Osiris genes in the N. lugens genomeNameGenomic locationSize (aa)OrientationDomainsBest matchE-valueosiris 2chr1[71781106–71785372]360-Duf1676D. melanogaster1.00E-33osiris 3chr1[16144454–16155056]289 + Duf1676B. mori2.00E-38osiris 6chr1[98218969–98225006]266-Duf1676D. melanogaster1.00E-25osiris 7chr1[21455532–21466129]310-Duf1676D. melanogaster5.00E-34osiris 8chr1[21416194–21423999]274-Duf1676D. melanogaster2.00E-29osiris 9chr1[21356327–21365789]300-Duf1676B. mori2.00E-27osiris 10chr1[78034913–78038815]286-Duf1676B. mori2.00E-14osiris 11chr1[77944874–77949318]292-Duf1676D. melanogaster1.00E-36osiris 12chr1[77897261–77910238]253-Duf1676D. melanogaster5.00E-20osiris 14chr1[77830153–77837895]283-Duf1676D. melanogaster6.00E-42osiris 15chr1[77812888–77815139]448-Duf1676D. melanogaster/osiris 16chr1[77763152–77782145]662-Duf1676B. mori1.00E-08osiris 17chr1[72092224–72117547]680 + Duf1676, DM4_12D. melanogaster7.00E-28osiris 18chr1[86484346–86489074]267-Duf1676B. mori3.00E-39osiris 19chr1[90906218–90909732]280-Duf1676B. mori2.00E-61osiris 20chr1[90888853–90893016]303-Duf1676D. melanogaster1.00E-26osiris 21chr3[14683004–14688778]228 + Duf1676B. mori2.00E-14osiris 22chrX[2469205–2491639]179-Duf1676B. mori4.00E-08osiris 23chr8[36186068–36194529]495-Duf1676D. melanogaster3.00E-06osiris 24chr1[90769914–90778891]478 + Duf1676, DM4_12D. melanogaster3.00E-54The Osiris sequences were obtained from the N. lugens genomic and transcriptomic databases. The Osiris genes were confirmed via the BLASTp algorithm with a cut-off E value of 1×10^−5^. The location, size and orientation indicate the locus, predicted amino acids (aa) and transcription orientation of the genes on the chromosome. Domains refer to the presence of the characteristic Duf1674 domain and/or the transmembrane domain DM4_12. D. melanogaster, Drosophila melanogaster; B. mori, Bombyx moriFig. 1. Bioinformatics analysis of the putative NlOsiris proteins*.* A Phylogenetic tree of insect Osiris proteins. A phylogenetic tree was constructed on the basis of the amino acid sequences of insect Osiris proteins via MEGA X (http://www.megasoftware.net/) with the maximum likelihood method with 1000 bootstrap replicates. Dm, D. melanogaster; Bm, B. mori; Nl, N. lugens. B Prediction of NlOsiris protein structure. The conserved domains of NlOsiris proteins were determined via SMART (http://smart.embl.de/), Pfam (http://pfam.xfam.org/) and NCBI (http://www.ncbi.nlm). The blue bars represent the number of amino acid residues, while the colored boxes indicate the characteristic domains. C Multiple sequence alignment of NlOsiris proteins. The conserved sequences, including a pair of conserved cysteine residues, the DUF1676 domain, the transmembrane region and the AQXLAY sequence, are highlighted in colored boxes
As the synteny of the Osiris genes was presented in the D. melanogaster and A. gambiae genomes, we examined the location of the Osiris genes in the N. lugens genome. Among the 20 NlOsiris genes, 17 were located on chromosome 1, whereas NlOsiris21, NlOsiris22, and NlOsiris23 were located on chromosomes 3, X, and 8, respectively. We still mapped the chromosomal locations of the Osiris genes across 23 insect species, and synteny was detected in the genomes of all 23 insect species (Figure S1). The predicted NlOsiris proteins had varying sizes, consisting of 179–680 amino acids (Table 1). All NlOsiris proteins contained a characteristic Duf1676 domain (Pfam: PF07898) (Fig. 1B). NlOsiris16 is special because it has two Duf1676 domains. The NlOsiris proteins had a predicted signal peptide at the N-terminus and a transmembrane region, except for NlOsiris9, which lacks a signal peptide sequence, and NlOsiris21, which lacks a transmembrane region. Additionally, NlOsiris17 and NlOsiris24 each contained a function-unknown Dm4_12 domain (Pfam: PF07841), a 115-aa motif with four highly conserved cysteine residues near the C-terminus. Multiple sequence alignments of NlOsiris proteins revealed conserved motifs, including a pair of conserved cysteine residues near the N-terminus, the Duf1676 domain, the transmembrane region and an AQXLAY sequence near the C-terminus (Fig. 1C).
Spatiotemporal expression analysis of NlOsiris genes
To understand the functions of the Osiris genes in N. lugens, we investigated their development- and tissue-specific expression patterns by searching the N. lugens transcriptome database and confirmed the development- and tissue-specific expression profiles of the NlOsiris genes via qRT‒PCR (Fig. 2A). The NlOsiris transcripts were almost undetectable or present at extremely low levels in female and male adults. For the eggs laid in the rice leaf sheaths, almost no transcripts were detected in the eggs at 24 and 72 h post-laying, but they were detected at high levels at 144 h, a time point before egg hatching. Notably, the NlOsiris genes presented periodic expression patterns throughout the nymphal stages. The transcripts were not detectable or at extremely low levels in newly hatched 1st–5th-instar nymphs at 0 h and increased to reach peak levels at the middle stage of each instar nymph, namely, at 36 h for the 1st–4th-instar nymphs and 48–60 h for the 5th-instar female and male nymphs. These results suggest that NlOsiris genes may function in egg hatching and the development of nymphs. Among the NlOsiris genes, NlOsiris11 presented a different expression pattern, whose transcripts were exclusively detected in 5th-instar female nymphs, with expression peaking at 36–48 h, implying a special function at the developmental stage.Fig. 2. Heatmap of the spatiotemporal expression profile of NlOsiris genes. A Developmental expression profiles of NlOsiris genes were obtained from the transcriptome data and visualized as a heatmap with GraphPad. The y-axis represents NlOsiris genes, and the x-axis denotes different developmental stages. AF, adult female; AM, adult male; E, egg; N, nymph; F5th, fifth-instar female nymph; M5th, fifth-instar male nymph. B Tissue-specific expression profiles of NlOsiris genes were obtained via qRT‒PCR and visualized as a heatmap with GraphPad. The y-axis represents NlOsiris genes, and the x-axis denotes the different tissues. FB, fat body; IN, integument; GU, gut; SG, salivary glands; WB, wing buds, which were dissected from fifth-instar nymphs. TE, testis; OV, ovary, dissected from male and female adults, respectively. Red indicates relatively high expression levels, and white represents relatively low expression levels
The NlOsiris genes presented tissue-specific expression patterns (Fig. 2B). The transcripts of most NlOsiris genes were detected at high levels in the wing bud, integument and/or fat body but were not detectable or at extremely low levels in the gut, testis, ovary and salivary gland. Despite the similar expression patterns, differences could be observed in these genes. NlOsiris8, NlOsiris15, NlOsiris18, NlOsiris19 and NlOsiris20 transcripts were detected at very high levels in the integument followed by the fat body but were not detected in the wing bud. The NlOsiris12 transcript was detected only in the wing bud, and the NlOsiris23 transcript was detected only in the gut. NlOsiris9, NlOsiris10 and NlOsiris11 transcripts were detected at similarly high levels in the wing bud, integument and fat body. The transcripts of the other NlOsiris genes, including NlOsiris2, NlOsiris3, NlOsiris6, NlOsiris7, NlOsiris14, NlOsiris16, NlOsiris17, NlOsiris21, NlOsiris22 and NlOsiris24, were detected at high levels in the wing bud but at low levels in the integument and fat body.
Effects of NlOsiris gene knockdown on development
To determine the functions of the Osiris gene family, we adopted an RNAi approach to knock down the transcript levels of each Osiris gene in 4th-instar N. lugens nymphs to observe the phenotypic changes. The knockdown of 9 genes, NlOsiris2, NlOsiris7, NlOsiris9, NlOsiris11, NlOsiris14, NlOsiris16, NlOsiris17, NlOsiris19 and NlOsiris24, led to a significant decrease in the survival rates of each target dsRNA-injected insect (Fig. 3A). Following RNAi, more than 80% of individuals in the dsNlOsiris14-, dsNlOsiris17-, and dsNlOsiris24-injected groups survived at 4 days post-injection (dpi), but these percentages decreased at 5 dpi (5th-instar nymphs) and declined dramatically to approximately l0% at 13 dpi (Fig. 3A). In the dsNlOsiris2-, dsNlOsiris7-, dsNlOsiris9-, dsNlOsiris16- and dsNlOsiris19-injected groups and the dsNlOsiris11-injected group, greater than 80% of individuals successfully molted into adults at 7 dpi; however, the survival rates decreased to 50% and 30% at 13 dpi, respectively. In contrast, knockdown of the other NlOsiris genes, including NlOsiris3, NlOsiris6, NlOsiris8, NlOsiris10, NlOsiris12, NlOsiris18, NlOsiris20-23, resulted in greater than 70% survival rates at 13 dpi, which were not significantly different, with 85% survival rates in dsGFP-injected individuals at 13 dpi. To confirm the RNAi effect, we conducted qRT‒PCR analysis to investigate the variation in the transcript level of each NlOsiris gene in 5th-instar nymphs at 48 h after molting. The results revealed that the transcript level of each NlOsiris gene was significantly lower in the dsNlOsiris-injected nymphs than in the dsGFP-injected controls (Fig. 3B).Fig. 3. Effects of NlOsiris gene knockdown on development. A Dynamic analysis of the survival rates of N. lugens. The 4th-instar nymphs were injected with dsNlOsiris (50 ng per insect) and observed for phenotypic variation at 24 h intervals for 13 days. dsGFP was injected as a negative control to determine the non-specific effects of dsRNA. Three independent biological replicates were conducted for each treatment (mean ± standard deviation; n = 80–100 nymphs). B Determination of RNAi efficiency. Total RNA was extracted from 5th-instar nymphs at 48 h after molting, and the transcript level variation of each NlOsiris was analyzed via qRT‒PCR, as described in Fig. 2. The results of triplicate experiments (n = 5) are shown with standard deviations. Significant differences were calculated via Student’s t test (P < 0.05 and **P < 0.01). C Abnormal phenotypes in N. lugens nymphs and adults upon NlOsiris gene knockdown. Group A: Abnormal body size and morphology were observed in the 5th-instar nymph (upper) and adult stages (lower panel). The ovaries were observed in female adults at 72 h after emergence. Group B: Abnormal wing phenotype. Deficient wing forms were observed in short-winged adults at 24 h after eclosion. Group C: Abnormal ovipositor phenotype. Abnormal ovipositors were observed in the adult females at 24 h after eclosion. The red arrowheads show the ovipositors in the abdomen integuments. For the RNAi experiments, each specific dsRNA (50 ng per insect) of the target genes was injected into 4th-instar nymphs, and the phenotypes were observed after RNAi. The dsGFP*-injected insects served as controls
The knockdown of NlOsiris genes generated different morphological defects in nymphs and/or adults (Fig. 3C), which can be divided into three major types: Group A: Abnormal body size: 20–25% of dsNlOsiris7-, dsNlOsiris11- or dsNlOsiris14-injected nymphs showed obviously shriveled abdomens and small body sizes at the 5th-instar nymphal stage. Only 50% of these nymphs could completely transition from 5th-instar nymphs to adults but had low survival rates at the adult stage. In addition, 50% of the dsNlOsiris2-injected nymphs and 80% of the dsNlOsiris11- or dsNlOsiris14-injected nymphs had flat abdomens at the adult stage, whereas more than 95% of the dsGFP-injected controls had obviously swollen abdomens and stretched intersegmental membranes in the tergums. Notably, these adults presented deficient ovaries without mature oocytes on the 3rd day post-eclosion, whereas dsGFP-injected adults presented normal ovaries with banana-shaped oocytes. Group B: Abnormal wing form: 85% of the dsNlOsiris19-injected nymphs and 90% of the dsNlOsiris24-injected nymphs successfully developed into adults, but the short-winged adults presented deficient wing forms. Their forewings were very small and irregularly shaped, whereas more than 95% of the dsGFP-injected controls had wings of normal size and regular shape. Group C: Abnormal ovipositor: Fewer than 50% of the dsNlOsiris17-injected nymphs developed into adults. However, 85% of the female adults presented abnormally transparent and soft ovipositors in the abdomen integuments, whereas more than 95% of the dsGFP-injected females presented brown and hard ovipositors in the abdomen integuments.
Effects of NlOsiris gene knockdown on feeding
We subsequently investigated whether flat abdomen was caused by feeding inhibition. The adults that were injected with dsNlOsiris2, dsNlOsiris11 or dsNlOsiris14 among the 4th-instar nymphs were fed an artificial diet, and their feeding behavior was observed by calculating the number of salivary sheaths in the parafilm membrane. As a result, dsGFP-injected adults generated more than 10 salivary sheaths/cm^2^ in the parafilm membrane on the 2nd day, which increased to 40 salivary sheaths/cm^2^ on the 7th day after feeding (Fig. 4A, B). In contrast, dsNlOsiris2-, dsNlOsiris11- or dsNlOsiris14-injected adults produced fewer than 10 salivary sheaths/cm^2^ in the parafilm membrane throughout 7 days of feeding (Fig. 4A, B), suggesting that NlOsiris knockdown inhibited feeding.Fig. 4. Effects of NlOsiris knockdown on feeding behavior. A Observation of salivary sheath production. The 5th-instar nymphs were injected with dsNlOsiris2, dsNlOsiris11 or dsNlOsiris14 (50 ng per insect) and were fed an artificial diet. The feeding behavior was observed, and the number of salivary sheaths on the Parafim/cm^2^ was calculated two days after injection. B Average number of salivary sheaths per insect. Each biological replicate consisted of 15 individuals, with a total of 4 biological replicates. The error bars indicate the standard deviations (n = 4)
Effects of NlOsiris gene knockdown on female reproduction
As most NlOsiris genes presented high transcript levels in eggs at 144 h after oviposition (Fig. 2A), we investigated the functions of NlOsiris genes in egg hatching by injecting each target dsRNA into newly emerged adults (0–2 h posteclosion). Knockdown of NlOsiris resulted in a significant decrease in hatching rates (Fig. 5A). Only 30–55% of the eggs laid from dsNlOsiris7-, dsNlOsiris11-, dsNlOsiris14-, dsNlOsiris17-, dsNlOsiris18-, dsNlOsiris19-, dsNlOsiris20- or dsNlOsiris24-injected female adults hatched to nymphs. In contrast, 90% of the eggs generated from dsGFP-injected female controls successfully hatched to nymphs. To understand the reason for hatching failure, we dissected the eggs laid in the rice leaf sheaths at 216 h, a time point just before hatching. More than 70% of the eggs from each target dsNlOsiris-injected female presented a typical banana shape with dark red and large eye spots. The presence of an eye spot in the egg was considered a viable fertilized egg, whereas those without an eye spot were considered unfertilized eggs [35]. Fewer than 30% of the eggs laid from dsNlOsiris11-, dsNlOsiris19- or dsNlOsiris24-injected females had no or only a light red and small eye spot, indicating unfertilized or abnormal eggs. Although these eggs were most likely to fail to hatch, our observations imply that the low hatching rates were probably not due to fertilization. Therefore, we observed egg–nymph transition in the rice leaf sheaths. We focused on dsNlOsiris17- and dsNlOsiris24-injected females to observe nymph hatching from the laid eggs. As a result, 60% of eggs were able to hatch into the 1st-instar nymphs; however, these insects stuck to the rice stems and could not leave, leading to hatching failure (Video S1). We confirmed the RNAi effects on the NlOsiris7, NlOsiris11, NlOsiris14, NlOsiris17, NlOsiris18, NlOsiris19, NlOsiris20 and NlOsiris24 genes in eggs 144 h after laying via qRT‒PCR. The transcript level of each NlOsiris gene was significantly lower in the dsNlOsiris-injected eggs than in the dsGFP-injected controls (Fig. S2).Fig. 5. Hatching rate following the injection of dsRNA targeting the Osiris gene. The dsRNA of the target gene was injected into female adults 0–2 h posteclosion. A Injection of dsOsiris7, dsOsiris11, dsOsiris14, dsOsiris17, dsOsiris18, dsOsiris19, dsOsiris20, or dsOsiris24 resulted in a significant decrease in the hatching rate of fertilized eggs. The error bars indicate the standard deviations (n = 3). A significant difference was detected via a Student’s t test (P < 0.05 and **P < 0.01). Reproductive phenotypes following the injection of dsRNA targeting the Osiris gene. The eggs were dissected from the rice leaf sheaths (216 h after laying). B After dsOsiris17* and dsOsiris24 were silenced, the 1st instar nymphs could not hatch normally
Discussion
With the completion of genomic and transcriptomic sequencing of the brown planthopper N. lugens, identification of the Osiris gene family via bioinformatics tools has become possible. Advanced technologies such as RNAi and gene editing have enabled researchers to explore the molecular functions of the Osiris family further. In this study, we identified and characterized 20 N. lugens Osiris genes by searching the N. lugens genomic and transcriptomic databases. The published literature indicates that insects have approximately 20 Osiris genes in their annotated genomes. Compared with the Diptera model insect D. melanogaster, N. lugens lacks the Osiris1, Osiris4, Osiris5 and Osiris13 genes, implying that these genes may play species-specific roles in D. melanogaster. Phylogenetic analysis revealed that each deduced N. lugens Osiris protein exhibited close evolutionary relationships with homologs from D. melanogaster and B. mori, suggesting a high degree of sequence conservation across diverse insect species. This conservation was evident not only in sequence homology but also in the synteny of the Osiris genes within their respective genomes. We analyzed the Osiris gene locus in the genomes of 23 insect species from eight orders. The results revealed that the synteny of the Osiris genes was present on one of the chromosomes of all the investigated insect genomes. The closer the evolutionary relationships are, the more similar the Osiris gene location and arrangement in the chromosome. Osiris gene clusters on one chromosome were most likely due to gene duplication during insect evolution.
Temporal and spatial expression profiles revealed that NlOsiris genes were almost undetectable in adult stages. However, pronounced expression peaks were observed during the mid-developmental stages of each instar nymph and in the eggs at 144 h post-laying within the rice leaf sheaths. These findings align with those of previous studies [11], which reported that Osiris genes are crucial for epidermal development during the larval and pupal stages. Notably, our data revealed that the timing of the peak expression of NlOsiris genes coincided with that of chitin binding-related genes (data not shown), suggesting that NlOsiris genes co-expressed with chitin metabolism-related genes and might be involved in the development of the epidermis during each nymphal stage. The tissue-specific expression profiles provided further evidence that most NlOsiris genes were specifically expressed in the integuments and wing buds of 5th-instar nymphs, which supported our speculations about the functions of Osiris in the cuticle. Several studies have established a link between Osiris gene expression and cuticle development. In D. melanogaster, 7 Osiris genes have been implicated in tracheal maturation, while chitin plays a crucial role in the construction of the tracheal system [8]. Silencing the locust LmOsiris17 gene resulted in abnormal development of the new cuticle [18]. Although our study did not provide direct evidence demonstrating the functional associations of NlOsiris genes with chitin metabolism, we observed deficient phenotypes of the cuticle morphology, which were caused by the knockdown of some NlOsiris genes (Fig. 6). (1) Knockdown of NlOsiris7, NlOsiris11 or NlOsiris14 resulted in significantly smaller body sizes with shriveled abdomens in 5th-instar nymphs, and knockdown of NlOsiris2 or NlOsiris11 or NlOsiris14 produced thin adults with visibly flattened abdomens. (2) Knockdown of NlOsiris19 or NlOsiris24 resulted in abnormally small and irregularly shaped forewings in adults. (3) Knockdown of NlOsiris17 resulted in abnormally transparent and soft ovipositors in the female abdomen integuments. These observations indicate that some Osiris genes have important functions in cuticle formation and structure. In addition, we found that the knockdown of NlOsiris2, NlOsiris11 or NlOsiris14 significantly affected feeding on the artificial diet. Few salivary sheaths formed at the feeding sites, which was consistent with the shriveled abdomen and decreased body weight of the NlOsiris2-, NlOsiris11- or NlOsiris14-injected insects. We hypothesize that expression silencing of these NlOsiris genes probably changed the mouthpart structure, thereby reducing feeding ability. Notably, the knockdown of NlOsiris2, NlOsiris11 or NlOsiris14 resulted in the generation of deficient ovaries in female adults that failed to lay eggs. These findings strongly suggest that the three Osiris genes have similarly important functions in feeding and female reproduction.Fig. 6. Abnormal phenotypes following the injection of dsRNA targeting each Osiris gene. The insects were treated as described in the Materials and Methods. Survival rate: survival rate on day 13
The transcripts of the NlOsiris genes were barely detectable at 24 and 72 h postlaying, corresponding to the early and middle development stages of the laid eggs. However, the transcript levels were significantly elevated in eggs at 144 h postlaying, a late developmental stage, implying that NlOsiris are involved in hatching. Our results revealed that the knockdown of 8 Osiris genes, NlOsiris7, NlOsiris11, NlOsiris14, NlOsiris17, NlOsiris18, NlOsiris19, NlOsiris20 or NlOsiris24, significantly decreased the hatching rates, indicating the functional importance of these genes for hatching. In N. lugens, the presence of an eye spot in the egg was considered a viable fertilized egg, whereas those without an eye spot were considered unfertilized eggs [35]. In our previous study, we reported that a large and dark red eye spot appeared in typical banana-shaped eggs approximately 120 h after laying and presented along with egg development until hatching. In this study, more than 70% of the eggs laid from the above dsNlOsiris-injected females presented an eye spot at 216 h, a time point just before hatching, implying that most of the laid eggs were fertilized eggs. On the basis of these observations, we suppose that the reason for hatching failure was not fertilization. The subsequent experiments confirmed our speculation. We found that the knockdown of NlOsiris17 or NlOsiris24 intriguingly generated phenotypes of hatching failure. Most of the laid eggs have transitioned to 1st-instar nymphs; however, these nymphs were not able to hatch successfully and died in rice leaf sheaths, strongly suggesting that NlOsiris genes are involved in the transition of egg–nymph.
The phylogenetic tree reflected, to some extent, the functional similarities and diversities of the Osiris family. For example, NlOsiris17 was closely related to NlOsiris24, and knockdown of its gene expression generated similar hatching failure phenotypes. Similarly, NlOsiris18 had a close evolutionary relationship with NlOsiris19, with gene knockdown leading to a significant reduction in hatching rates. Interestingly, despite divergent from each other, NlOsiris2, NlOsiris11, and NlOsiris14 presented similar RNAi phenotypes, suggesting that these genes may share common regulatory mechanisms or participate in overlapping pathways [36, 37]. In the future, it will be necessary to utilize comparative transcriptome sequencing to uncover the downstream pathways and molecular targets associated with Osiris gene expression, thereby providing insights into the detailed regulatory mechanisms involved.
Although the Osiris genes presented similar temporal‒spatial expression profiles, they presented functional diversity and specificity across insect species. For example, B. mori Osiris9a is specifically expressed in silk glands and is necessary for silk fiber formation [17, 38]. In contrast, knockdown of the homologous gene Osiris9 in N. lugens resulted in an apparently lethal phenotype. Drosophila melanogaster Osiris genes are highly expressed at 48 h in the pupal stage [11]. The knockdown of DmOsiris genes, such as Osiris7, in the larval stage resulted in larvae failing to complete metamorphosis and remaining at the prepupal stage. Nilaparvata lugens undergoes incomplete metamorphosis, in which the nymph is essentially similar to an adult, and there is no pupal stage. Knockdown of the homologous Osiris7 in N. lugens resulted in small body sizes in the nymphs and decreased hatching rates, suggesting that the functions of the homologous Osiris genes differ between holometabolous and hemimetabolous insect species.
Osiris genes densely clustered on one of the chromosomes across insect species. Synteny-generated genetic redundancy has been reported for Drosophila Osiris9, Osiris15, and Osiris19 in tracheal function [8]. In this study, the knockdown of nine NlOsiris genes, e.g., Osiris3, Osiris6, Osiris8, Osiris10, Osiris12, Osiris15, and Osiris21-23, did not cause notable changes at the individual or tissue level, such as changes in survival rates, hatching rates or cuticle morphology. However, this does not imply that they are functionally unimportant, as some NlOsiris genes may have redundant roles owing to their similar development-specific and tissue-specific expression patterns.
In conclusion, Osiris genes play crucial roles in wing, mouthpart, ovipositor and ovary development, as well as in egg hatching and feeding, ensuring survival and reproductive success. Our study provides insights into the physiological functions of Osiris and highlights their potential association with cuticle development. Further studies are needed to clarify the precise regulation and action mechanisms of the Osiris gene during insect development and reproductive processes, e.g., to elucidate the relationships between the Osiris genes and chitin metabolism in insect cuticles. Further studies should focus on determining the specific functional roles and interactions of Osiris gene family members via CRISPR/Cas9 gene editing in insects.
Supplementary Information
Supplementary Material 1.Supplementary Material 2.Supplementary Material 3.Supplementary Material 4.Supplementary Material 5.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Thurmond, J., et al., Fly Base 2.0: the next generation. Nucleic Acids Res, 2019. 47(D 1): D 759-D 765.10.1093/nar/gky 1003 PMC 632396030364959 · doi ↗ · pubmed ↗
- 2Finn, R.D., et al., The Pfam protein families database. Nucleic Acids Res, 2008. 36(Database issue): p. D 281–8.10.1093/nar/gkm 960PMC 223890718039703 · doi ↗ · pubmed ↗
- 3Shah, N., et al., Evolution of a large, conserved, and syntenic gene family in insects. G 3 (Bethesda), 2012. 2(2): 313–9.10.1534/g 3.111.001412 PMC 328433822384409 · doi ↗ · pubmed ↗
- 4Sun Z, et al. Osiris gene family defines the cuticle nanopatterns of Drosophila. Genetics. 2024;227(2):iyae 065.10.1093/genetics/iyae 065PMC 1115192938652268 · doi ↗ · pubmed ↗
- 5Mika, K., et al., Olfactory receptor–dependent receptor repression in Drosophila. Science Advances, 2021. 7(32): eabe 3745.10.1126/sciadv.abe 3745 PMC 834622034362730 · doi ↗ · pubmed ↗
- 6Suetsugu, Y., et al., Large scale full-length c DNA sequencing reveals a unique genomic landscape in a lepidopteran model insect, Bombyx mori. G 3 (Bethesda), 2013. 3(9): 1481–92.10.1534/g 3.113.006239 PMC 375590923821615 · doi ↗ · pubmed ↗
- 7Cheng, T., et al., Effects of Osiris 9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression. Int J Mol Sci. 2020;21(5):1888.10.3390/ijms 21051888 PMC 708479832164252 · doi ↗ · pubmed ↗
- 8Zu, H., et al., Jasmonate-mediated polyamine oxidase 6 drives herbivore-induced polyamine catabolism in rice. Plant J, 2024;120(5):2000–2013.10.1111/tpj.1709439432737 · doi ↗ · pubmed ↗