Spinning together agricultural and evo-devo research for Gynandropsis gynandra (spider plant)
Brandi Zenchyzen, Jocelyn C Hall

TL;DR
This paper reviews Gynandropsis gynandra, a leafy vegetable with agricultural and scientific value, and highlights the need for integrating research across disciplines to improve its cultivation and study.
Contribution
The paper bridges agricultural and evo-devo research on G. gynandra, proposing standardized approaches to enhance interdisciplinary collaboration.
Findings
G. gynandra is a valuable crop with potential for both agriculture and evolutionary developmental studies.
Molecular tools have advanced understanding of C4 photosynthesis and antiherbivore defenses in G. gynandra.
Distinctive floral features and phenotypic diversity offer insights for breeding and developmental research.
Abstract
Gynandropsis gynandra (Cleomaceae; formerly Cleome gynandra) is a leafy vegetable widely cultivated across Africa, uniquely positioned at the intersection of agricultural and evo-devo research. It is gaining recognition as an ‘opportunity crop’, valued locally for its nutritional and medicinal properties with ongoing agricultural research aimed at the development of improved cultivars and agronomic practices. Concurrently, its close evolutionary proximity to Arabidopsis thaliana, combined with its contrasting traits, positions G. gynandra as a model for studying C4 photosynthesis and floral development. Despite its relevance to both agricultural and evo-devo research, integration of findings between disciplines remains limited, hindered in part by inconsistent nomenclature and the lack of standardized morphological descriptors. To address this disconnect, this review synthesizes…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
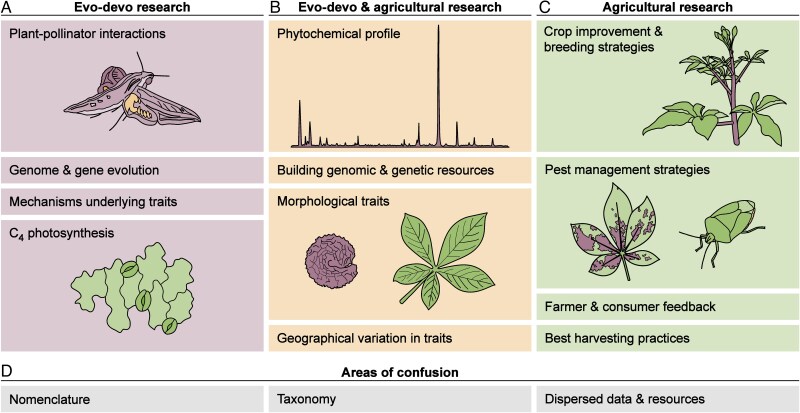 Fig. 1
Fig. 1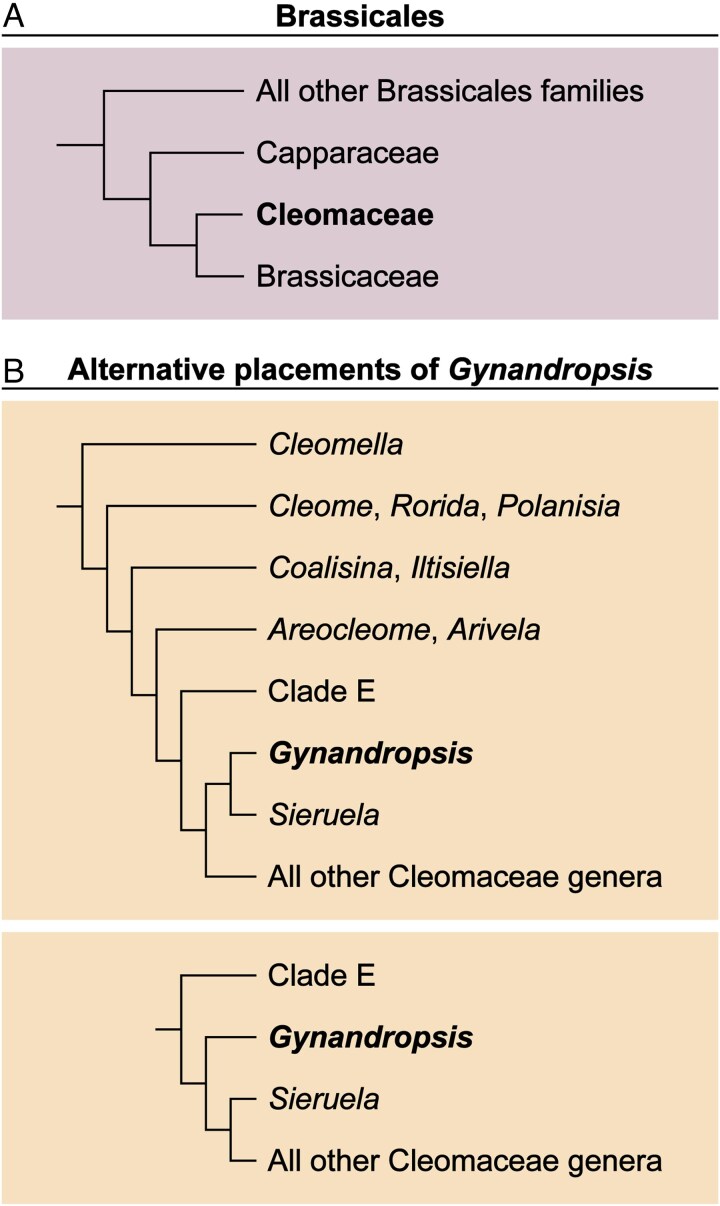 Fig. 2
Fig. 2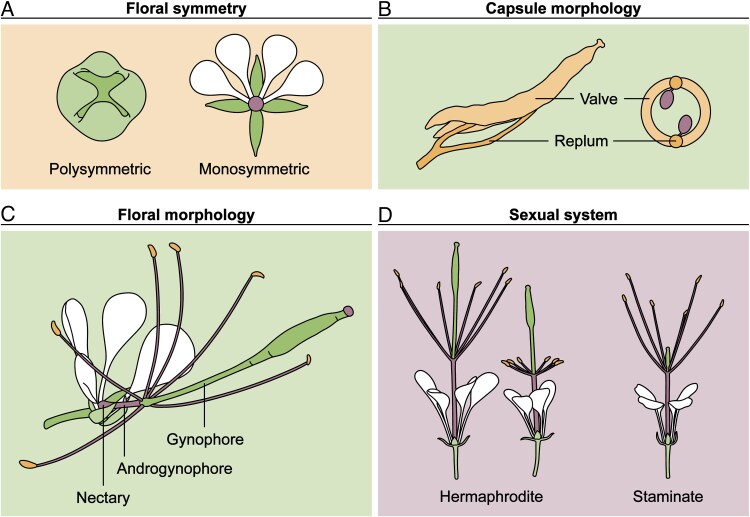 Fig. 3
Fig. 3| Chemicals analysed | Type of tissue | Number and origin of accessions | Publication |
|---|---|---|---|
| Amino acids, fatty acids, protein | Seed | 4 accessions, Zambia |
|
| Amino acids, minerals, fatty acids | Leaf | 1 accession, Ghana |
|
| Anthocyanins, phenolics, tannins | Leaf | 33 accessions, 8 African and 3 Asian countries |
|
| Ascorbic acid, carotenoids, chlorophylls, tocopherols | Leaf | 76 accessions, 4 Asian and 11 African countries |
|
| Ascorbic acid, carbohydrates, β-carotene, fatty acids, folate, minerals, protein, riboflavin, thiamin | Leaf | 1 accession, South Africa |
|
| Ascorbic acid, β-carotene, flavonoids, minerals, phenolics | Unspecified | 1 accession, Zimbabwe |
|
| Ascorbic acid, carotenoids, minerals, phytate, tocopherols | Leaf | 1 accession, Tanzania |
|
| Ascorbic acid, flavonoids, minerals, phenolics | Leaf | 8 accessions, Kenya and Zambia |
|
| β-Carotene, minerals, riboflavin | Leaf, stem | 1 accession, South Africa |
|
| Carotenoids, flavonoids, glucosinolates, phenolics | Leaf | 1 accession, Tanzania |
|
| Fatty acids | Seed | 1 accession, India |
|
| Fatty acids, folic acid | Unspecified | 1 accession, South Africa |
|
| Flavonoids | Aerial parts | 1 accession, Saudi Arabia |
|
| Flavonoids | Stem | 1 accession, Taiwan |
|
| Flavonoids, glucosinolates, minerals | Flower, fruit, leaf, stem | 30 accessions, 6 African countries |
|
| Flavonoids, phenolics | Flower, leaf and stem, seed | 8 accessions, Kenya |
|
| Flavonoids, phenolics | Leaf | 1 accession, India |
|
| Flavonoids, phenolics | Whole plant | 1 accession, Burkina Faso |
|
| Glucosinolates | Seed | 1 accession, Thailand |
|
| Glucosinolates, semi-polar metabolites, volatiles | Leaf | 48 accessions, 4 Asian and 10 African countries |
|
| Isothiocyanates | Seed | 1 accession, unspecified |
|
| Isothiocyanates | Seed | 1 accession, unspecified |
|
| Minerals | Leaf | 70 accessions, 4 Asian and 11 African countries |
|
| Minerals | Leaf | 1 accession, Kenya |
|
| Minerals | Leaf | 1 accession, Kenya |
|
| Minerals | Leaf | 1 accession, Kenya |
|
| Minerals, phenolics | Leaf | 17 accessions, 5 African countries |
|
| Phenolics | Leaf | 1 accession, Malaysia |
|
| Phenolics, condensed tannins | Leaf | 5 accessions, 3 African countries |
|
| Semi-polar metabolites, volatiles | Aerial parts | 1 accession, Kenya |
|
| Terpenoids | Whole plant | 1 accession, India |
|
| Volatiles | Inflorescence | 2 accessions, Malaysia and Malawi |
|
| Volatiles | Aerial parts | 1 accession, Kenya |
|
| Volatiles | Leaf | 5 accessions, Kenya and Tanzania |
|
| Volatiles | Leaf, whole plant | 5 accessions, Kenya and Tanzania |
|
| Type of data | Accession name and origin | Type of tissue | Publication | Data repository |
|---|---|---|---|---|
| Genome | Gyn, Malaysia | Leaf |
| BioProject ID (NCBI): PRJNA843598 |
| Genome, RNA-seq, transcriptome | Unspecified | Leaf (5 developmental stages, 2 temperature conditions), flower (2 developmental stages), capsule, stem, root |
| BioProject ID (NGDC): PRJCA017363, PRJCA017364 |
| RNA-seq, transcriptome | TOT8917, Malawi | Adaxial petals, abaxial petals, androgynophore (3 developmental stages), filaments, gynophore |
| BioProject ID (NCBI): PRJNA680567, PRJNA1030768 |
| RNA-seq, transcriptome | Taiwan | Leaf (developing) |
| BioProject ID (NCBI): PRJNA714768, PRJNA381122 |
| RNA-seq | Unspecified | Leaf (6 developmental stages), seedling (3 developmental stages), seed (3 developmental stages), sepal, petal, stamen, carpel, stem, root |
| BioProject ID (NCBI): PRJNA237449 |
| RNA-seq | Unspecified | Leaf (3 regions), leaf mesophyll cells, leaf bundle sheath cells |
| BioProject ID (NCBI): PRJNA243610 |
| RNA-seq | Unspecified | Leaf mesophyll cells, leaf guard cells |
| ArrayExpress (EBI): E-MTAB-3379 |
| miRNA-seq | Unspecified | Leaf (2 developmental stages) |
| BioProject ID (NCBI): PRJNA377356 |
| Targeted DNA sequence (Angiosperms353) | 857511.0 (PRE), South Africa | Leaf |
| BioProject ID (NCBI): PRJNA1080677 |
| RNA-seq, DNAseI-seq | Malaysia-01, Malaysia | Seedling (5 developmental stages/light treatments) |
| BioProject ID (NCBI): PRJNA640984 |
| DNaseI-seq | Unspecified | Leaf (3 developmental stages/light treatments) |
| BioProject ID (NCBI): PRJNA419285 |
| RNA-seq | Unspecified | Seedling (2 developmental stages/light treatments) |
| ArrayExpress (EBI): E-MTAB-4355 |
| Genome survey sequence, RNA-seq | Unspecified | Leaf |
| BioProject ID (NCBI): PRJNA542714 |
| Chloroplast genome sequence | DS20200516011 (NF), China | Leaf |
| BioProject ID (NCBI): PRJNA687638 |
| QTL | TOT7200, Malaysia × TOT8917, Malawi; TOT7199, Malaysia × TOT8918, Malawi | Leaf |
| GitHub: plycs5/GgQTL |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant biochemistry and biosynthesis · Photosynthetic Processes and Mechanisms · Plant Molecular Biology Research
INTRODUCTION
The study of Gynandropsis gynandra (L.) Briq. highlights the potential for cross-disciplinary insights through the integration of agricultural and evolutionary developmental biology (evo-devo) research. Yet, these fields remain largely disconnected for this flowering plant species. Gynandropsis gynandra (Cleomaceae), also referred to as spider plant or spider flower due to its elongated floral structures, is a nutrient-rich leafy vegetable and versatile medicinal plant native to Africa and Asia (Khuntia et al., 2022; Moyo and Aremu, 2022). As an essential crop for maintaining food security in resource-limited regions across Africa, G. gynandra is extensively studied for its agricultural-related properties such as nutrient content (Moyo et al., 2018; Sogbohossou et al., 2019; Chataika et al., 2021; Houdegbe et al., 2022a) and leaf yield (Houdegbe et al., 2018; Blalogoe et al., 2020; Chatara et al., 2023; Nzungize et al., 2024). Concurrently, G. gynandra is gaining recognition as a model organism for evo-devo research due to its close evolutionary relationship with Arabidopsis thaliana (Brassicaceae) and its contrasting traits including C_4_ photosynthesis (Huang et al., 2021; Zhao et al., 2023; Schreier et al., 2024) and atypical floral features (Zohoungbogbo et al., 2018; Zenchyzen et al., 2023b, 2023a). Despite significant advancements in each field, a disconnect persists. Agricultural reviews on G. gynandra often overlook potential insights from evo-devo studies, while evo-devo research is seldom linked to practical applications for crop improvement and commonly disregards data from agricultural literature. A complex taxonomic history compounds this division, with G. gynandra frequently referred to by its basionym Cleome gynandra L., outdatedly classified as a member of the family Capparaceae, or confused with other Cleomaceae species [e.g. Tarenaya houtteana (Schltdl.) Soares Neto & Roalson]. These inconsistencies make it challenging to determine whether journal articles are addressing the same species, further hindering cross-disciplinary research efforts (Fig. 1).
Focal areas of Gynandropsis gynandra research vary between disciplines. (A) Evo-devo studies centre on plant–pollinator interactions, genome and gene evolution, the mechanisms underlying traits, and C4 photosynthesis, while (C) agricultural studies emphasize crop improvement and breeding strategies, pest management strategies, farmer and consumer feedback, and best harvesting practices. (B) Shared resources, including phytochemical, genomic, genetic and morphological data, as well as information on geographical trait variation, offer valuable cross-disciplinary insights for both evo-devo and agricultural research. (D) However, integration is limited by inconsistent nomenclature, taxonomic confusion and scattered data and resources.
Integrated agricultural and evo-devo research offers a powerful framework to address pressing challenges in sustainable crop production while also advancing our understanding of flowering plant evolution. Agricultural research faces the urgent task of enhancing food security amid a changing climate and growing global population (Sogbohossou et al., 2018a; Henkhaus et al., 2020; Mashamaite et al., 2022). Advancements have focused predominantly on boosting the yield of a few major crop plants, resulting in just ten species dominating calorie production on cropland (Ray et al., 2019; Henkhaus et al., 2020). However, diversifying crops and domesticating underutilized species have proven effective in promoting ecosystem resilience and sustainability (Henkhaus et al., 2020; Knez et al., 2024). Meanwhile, evo-devo studies, which have long relied on model organisms, are expanding to include taxa with diverse traits (Schrager-Lavelle et al., 2017; Mabry et al., 2024; Sharma et al., 2024). Advances in genomic and genetic tools now enable detailed investigations into non-model species, enriching our understanding of plant evolution and diversification (Delaux et al., 2019). In the case of G. gynandra, morphological and phytochemical characterizations could provide critical context for evo-devo studies, while genetic discoveries have the potential to accelerate breeding and improvement programmes (Macel et al., 2010, Sogbohossou et al., 2018a; Jeiter and Smets, 2024). This review synthesizes findings from agricultural and evo-devo research on G. gynandra, providing an overview of its phylogenetic placement, geographical distribution, agricultural and medicinal uses, phytochemical profile, genomic and genetic resources, and morphological characteristics. While other reviews have focused primarily on its agronomically important traits, a comprehensive synthesis on the species’ morphology is lacking. By acknowledging key gaps and proposing future research directions, we aim to bridge agricultural and evo-devo perspectives, demonstrating their complementary potential to drive innovation and discovery in G. gynandra research.
TAXONOMIC HISTORY
Although G. gynandra was previously placed within the subfamily Cleomoidae of the family Capparaceae – a classification that persists in the G. gynandra literature – it is now a member of the family Cleomaceae following the division of Capparaceae s.l. into two distinct families, Capparaceae s.s. and Cleomaceae (Hall et al., 2002; Iltis et al., 2011; Angiosperm Phylogeny Group et al., 2016) (Fig. 2A). The separation of Capparaceae s.l. is well supported by molecular data, which reveal three clades, Capparaceae s.s., Cleomaceae, and Brassicaceae, with Cleomaceae more closely related to Brassicaceae than to Capparaceae s.s. (Hall et al., 2002; Iltis et al., 2011; Cardinal-McTeague et al., 2016; Edger et al., 2018 ; Hendriks et al., 2023). This recognition of Cleomaceae as a distinct family is further supported by morphological synapomorphies, including strongly incurved seeds, bracteate inflorescences, dry fruit with a replum (i.e. persistent placental tissue between the two valves of the capsule), and palmately compound leaves (Hall et al., 2002; Iltis et al., 2011).
The phylogenetic placement of Cleomaceae and Gynandropsis. (A) Capparaceae, Cleomaceae, and Brassicaceae are separate monophyletic families within the order Brassicales. Cleomaceae is sister to Brassicaceae. Cleomaceae plus Brassicaceae are sister to Capparaceae. (B) Phylogenetic studies place Gynandropsis either in a monophyletic clade with Sieruela, or sister to Sieruela plus the remaining Cleomaceae genera. The Brassicales and Cleomaceae phylogenies are based on Iltis et al. (2011) and Saunders et al. (2024), respectively. Clade E: Corynandra, Stylidocleome, Dipterygium, Puccionia, Thulinella, Gilgella, and Kersia.
Robust phylogenetic hypotheses for Cleomaceae have led to taxonomic revisions that align well-supported clades with generic classifications (Feodorova et al., 2010; Patchell et al., 2014; Roalson and Hall, 2017). Importantly, the large paraphyletic Cleome L. s.l. was segregated into multiple smaller genera (Feodorova et al., 2010; Patchell et al., 2014; Bayat et al., 2018). Supported by both molecular and morphological data, G. gynandra is the sole representative of its genus, which is more closely related to several other Cleomaceae genera (e.g. Sieruela Raf., Arivela Raf.) than it is to Cleome s.s. (Feodorova et al., 2010, Mabry et al., 2020; Saunders et al., 2024) (Fig. 2B). For simplicity, we refer to G. gynandra as Gynandropsis hereafter. The placement of Gynandropsis within the family remains unresolved. While some studies suggest that Gynandropsis is in a clade with Sieruela, others propose that it is sister to Sieruela plus several genera (Feodorova et al., 2010; Mabry et al., 2020; Saunders et al., 2024) (Fig. 2B).
Gynandropsis is a polymorphic species with a broad distribution, which has likely greatly contributed to its complex taxonomic history with more than 30 synonyms (Iltis, 1960; Roalson and Hall, 2017). Most frequently used synonyms include Cleome gynandra, Cleome pentaphylla L., and Gynandropsis pentaphylla L. (DC.) (Iltis, 1960). Gynandropsis is distinguished from other Cleomaceae genera by its three foliate bracts, white or pale coloured petals, and a prominent androgynophore and gynophore (i.e. stalk-like structures bearing the reproductive organs and pistil, respectively) (Iltis, 1960; Roalson and Hall, 2017). However, it is occasionally mistaken for other Cleomaceae taxa including the common ornamental T. houtteana (formerly T. hassleriana; Neto et al., 2022) and other species with elongated androgynophores, i.e. Podandrogyne Ducke and Cleoserrata speciosa (Raf.) Iltis. Notable morphological differences include T. houtteana’s stipular spines, bright pink to purple (rarely white) petals, and absence of an elongated androgynophore (Neto et al., 2022). Podandrogyne species feature unisexual flowers, arillate seeds, and often orange to red petals, while Cleoserrata speciosa is characterized by unifoliate bracts and pink to purple petals (seldom white) (Iltis, 1960; Iltis and Cochrane, 1989, 2007; Bayat et al., 2018). Additionally, Podandrogyne and Cleoserrata speciosa are native to the Americas rather than Africa and Asia (Iltis and Cochrane, 1989, 2007; Bayat et al., 2018; Neto et al., 2022).
GEOGRAPHICAL DISTRIBUTION
Gynandropsis is regarded as a ‘weedy’ species, having spread extensively from Africa and Asia alongside human movements (Iltis, 1960; Shilla et al., 2019). It is now widespread in tropical and subtropical regions across the world, including parts of North and South America, Europe, and Australia (Iltis, 1960; Bhattacharya et al., 2019; Shilla et al., 2019). Although Gynandropsis is considered native to both Africa and Asia, recent biogeographical analyses predict that a Gynandropsis and Sieruela clade originated in Africa and radiated outward (Saunders et al., 2024). Across its species range, Gynandropsis exhibits extensive morphological and phytochemical variation, which is correlated with geographical origin (Sogbohossou et al., 2019, 2020). Gynandropsis thrives in a variety of soil types and environmental conditions and is commonly found near human settlements and roadsides, possibly reflecting its early introduction routes (Iltis, 1960; Bhattacharya et al., 2019; Shilla et al., 2019). The success of Gynandropsis in hot, sunny, and dry environments is likely due in part to C_4_ photosynthesis, an innovation that has independently evolved multiple times within Cleomaceae (Rajendrudu and Rama Das, 1982; Feodorova et al., 2010; Hoang et al., 2023). In addition to its wild populations, Gynandropsis is cultivated in home gardens and croplands across Africa and Asia (Iltis, 1960; Rajendrudu and Rama Das, 1982; Shilla et al., 2019). As a result of its broad distribution and history of human use, Gynandropsis has numerous vernacular names, which vary across regions (see Iltis, 1960 ; Chweya and Mnzava, 1997).
AGRICULTURAL AND MEDICINAL USES
Gynandropsis is a versatile plant valued for both its agricultural and medicinal applications. Consumed primarily as a leafy vegetable, the leaves are prepared in various ways, including boiling as a potherb for use in sauces, stews, curries, and relish, pickling for a rice flavouring, and serving fresh in salads (Watt, 1890; Burkill, 1935, 1985; Chweya and Mnzava, 1997; Chataika et al., 2020). Tolerance for the bitterness of Gynandropsis leaves differs across regions (Flyman and Afolayan, 2006; Sogbohossou et al., 2018a). Methods used to reduce astringency include blanching and replacing the water prior to cooking, soaking in milk overnight, or mixing with other fruit or vegetables (Flyman and Afolayan, 2006; Onyango et al., 2013). Preservation practices such as blanching then sun-drying allow for year-round consumption, with dried leaves either reconstituted with water before cooking or ground and added to weaning food (Chweya and Mnzava, 1997; Flyman and Afolayan, 2006; Onyango et al., 2013). Gynandropsis seeds are crushed and made into mustard (Watt and Breyer-Brandwijk, 1932).
In addition to its culinary significance, Gynandropsis is occasionally used as livestock fodder (Burkill, 1985; Chataika et al., 2020) and has potential as a biological control by diminishing insect infestations and/or diverting insect pests from other cultivated plants such as Brassica crops (Brassicaceae), roses (Rosa hybrids; Rosaceae), and the common bean (Phaseolus vulgaris; Fabaceae) (Waiganjo et al., 2006; Nyalala and Grout, 2007; Kimbokota and Torto, 2013; Zedler et al., 2016). The attractiveness of Gynandropsis to insect pests also poses a significant challenge to its cultivation, necessitating the development of integrated pest management strategies (Francisco et al., 2024). Since many Gynandropsis farmers avoid chemical pesticides due to environmental and health risks, alternative strategies may include developing pest-resistant varieties and removing older crops, as insect infestations peak 8 weeks after sowing (Sogbohossou et al., 2018a; Francisco et al., 2024). Despite its susceptibility to infestations, the essential oil of Gynandropsis has tick repellent and acaricidal properties and has been proposed as a biological control against livestock ticks (Malonza et al., 1992; Lwande et al., 1999).
Beyond its agricultural importance, Gynandropsis is valued as a medicinal plant with all parts used in treating various ailments, as previously reviewed (see Khuntia et al., 2022; Moyo and Aremu, 2022) and summarized as follows. Leaf decoctions are traditionally used to treat malaria (Yetein et al., 2013), with experiments on mice supporting its efficacy (Igoli et al., 2016). Leaf poultices are used to alleviate pain, such as rheumatism discomfort and headaches (Burkill, 1935, 1985), as well as to treat abscesses and wounds (Shanmugam et al., 2012; Neamsuvan and Bunmee, 2016). Gynandropsis extracts possess anti-inflammatory, anti-nociceptive, and analgesic properties in rodents (Mule et al., 2008; Narendhirakannan et al., 2007 ; Ghogare et al., 2009; Bala et al., 2012), and anti-bacterial and anti-fungal activity (Borgio et al., 2008; Sridhar et al., 2014 ; Imanirampa and Alele, 2016 ; Kanimathi et al., 2019; Rotich and Mwafaida, 2024). Uses of Gynandropsis for pregnant women and postnatal mothers include chewing to induce labour and consuming to treat anaemia and stimulate milk production (Kamatenesi-Mugisha and Oryem-Origa, 2007; Dansi et al., 2008; Bosire, 2014). Clinical trials demonstrate that Gynandropsis consumption improves the haematological profile of lactating mothers (Bosire, 2014). The seed oil is used internally to expel parasitic worms (Watt, 1890; Burkill, 1985), and in vitro studies with Gynandropsis extracts confirm its anthelmintic properties (Ajaiyeoba et al., 2001; Fouche et al., 2016). Traditional ear-related treatments include dripping leaf sap into the ear to treat earaches and inserting the fruit into the ear canal to soften and extract wax (Watt, 1890; Watt and Breyer-Brandwijk, 1932; Burkill, 1935, 1985). It is occasionally noted that caution is needed when treating ailments with Gynandropsis as it may cause dermatitis (Watt, 1890; Watt and Breyer-Brandwijk, 1932; Mitchell, 1974; Burkill, 1985). Gynandropsis extracts also exhibit anti-cancer activity in mice (Bala et al., 2010). The numerous uses of Gynandropsis highlight the need for continued research to optimize its agronomic and pharmacological potential.
PHYTOCHEMICAL PROFILE
Plants produce a diverse array of chemicals that are essential for regulating plant growth and development, mediating plant–animal and plant–environment interactions, and serve as valuable nutritional and medicinal resources for human health (Hao et al., 2025). Metabolomics is a rapidly advancing field that has enabled comprehensive characterization of Gynandropsis phytochemicals through a variety of analytical methods (Sogbohossou et al., 2020; Hao et al., 2025) (Table 1). Phytochemical investigations on Gynandropsis have focused primarily on metabolites associated with nutrition, taste, and other health-promoting properties. Despite considerable variation in phytochemical concentrations across Gynandropsis accessions, the leaves are generally rich in minerals including calcium, copper, iron, magnesium, manganese, phosphorus, potassium, sodium, and zinc, which play vital roles in both plant physiology and human nutrition (Omondi et al., 2017; Moyo et al., 2018; Houdegbe et al., 2022a). Gynandropsis leaves are also high in vitamins, including provitamin A carotenoids (e.g. β-carotene), vitamin B (e.g. folate, riboflavin, thiamin), vitamin C (i.e. ascorbic acid), and vitamin E (e.g. tocopherols) (van Jaarsveld et al., 2014; Gowele et al., 2019; Sogbohossou et al., 2019). Although non-haem iron from plants is less bioavailable than haem iron found in meat, the high ascorbic acid content in Gynandropsis leaves may enhance iron absorption, making it a valuable dietary component for addressing iron deficiency (van Jaarsveld et al., 2014; Gowele et al., 2019; Houdegbe et al., 2022a).
Other non-nutritive chemical classes present in Gynandropsis leaves include flavonoids (e.g. anthocyanins), glucosinolates, and tannins, each containing compounds with health-promoting properties such as antioxidant and anti-inflammatory activity (Omondi et al., 2017; Moyo et al., 2018; Chataika et al., 2021). Dietary antioxidants are important for preventing the formation and activity of reactive oxygen and nitrogen species, which can cause damage to biomolecules, such as DNA, lipids, and proteins (Halliwell, 1996). Several studies have reported high antioxidant activity of Gynandropsis extracts (Meda et al., 2013; Moyo et al., 2018; Chandradevan et al., 2020a; Chataika et al., 2021; Thovhogi et al., 2021). Despite their therapeutic potential, glucosinolates and tannins contribute to the bitter taste of Gynandropsis leaves (Songsak and Lockwood, 2002; Kutsukutsa et al., 2014; Chataika et al., 2021). The levels of astringent compounds and undesirable elements (e.g. lead, cadmium) can be altered through selective breeding as well as cooking and cultivation methods (Hutchinson, 2011; Moyo et al., 2016; Omondi et al., 2017). For example, Gynandropsis can accumulate more glucosinolates when grown in the field compared to a glasshouse, and both the type and amount of fertilizer can affect leaf mineral content (Hutchinson, 2011; Omondi et al., 2017). Achieving a balance in the concentration of astringent compounds in Gynandropsis leaves is key to improving its palatability while maintaining its human health benefits and ability to deter herbivores.
Although pest management is essential for cultivating healthy Gynandropsis plants for both agricultural and research purposes, limited studies have identified the specific compounds involved in attracting or deterring herbivorous insects (Lwande et al., 1999; Nyalala et al., 2011, 2013; Kimbokota and Torto, 2013). Specialized plant metabolites can be unpalatable, antidigestive, or toxic to herbivores, and may be constitutively produced or induced in response to herbivory (Mithöfer and Boland, 2012). Trichomes not only serve as a physical barrier but also as storage and secretion sites for these metabolites (Mithöfer and Boland, 2012; Schuurink and Tissier, 2020). Broad chemical classes involved in plant defence include glucosinolates, terpenoids, and tannins (Barbehenn and Peter Constabel, 2011; Mithöfer and Boland, 2012). In Gynandropsis, glucocapparin is generally the dominant glucosinolate (Kjær and Thomsen, 1963; Songsak and Lockwood, 2002; Neugart et al., 2017; Sogbohossou et al., 2020), with the highest concentration in the flowers and seeds (Omondi et al., 2017). When plant tissues are damaged, glucosinolates are hydrolysed into volatile sulphur-containing compounds, such as isothiocyanates (Mithöfer and Boland, 2012). Isothiocyanates have been detected in Gynandropsis foliar volatiles (Nyalala et al., 2013; Sogbohossou et al., 2020) and contribute to its repellency against spider mites (Nyalala et al., 2013).
Floral phytochemicals not only play a role in reducing florivory, but also in pollinator attraction through the production of rewards, pigments, and volatiles (Parachnowitsch and Manson, 2015). Although Gynandropsis secretes small amounts of floral nectar, its high sugar concentration attracts nectar foraging ants, bees, butterflies, flies, and hawkmoths (Martins and Johnson, 2013; Raju and Rani, 2016; Zenchyzen et al., 2024). Like several other Cleomaceae species, Gynandropsis nectar fluoresces vibrant blue under UV radiation (Thorp et al., 1975; Zenchyzen et al., 2024). Additional research is needed to determine the chemical basis of this fluorescence and if it serves an ecological role, such as a visual signal for pollinators, or is simply a byproduct (Thorp et al., 1975; Zenchyzen et al., 2024). Floral volatile profiles vary significantly in Gynandropsis, with nitrogen-containing compounds as the major component in an African accession and benzenoids predominating in an Asian accession (Zenchyzen et al., 2025). Similar to isothiocyanates, the nitrogen-containing floral volatiles may be degradation products or biosynthetic precursors of glucosinolates (Halkier and Gershenzon, 2006; Mithöfer and Boland, 2012). The differences in chemical profiles align with the preferences of the effective pollinators in each region – hawkmoths in Africa (Monteiro, 1875; Werth, 1942; Oronje, 2011; Martins and Johnson, 2013) and bees and butterflies in Asia (Burkill, 1916; Chandra et al., 2013; Raju and Rani, 2016). Floral volatile profiling across a broader range of accessions is needed to assess whether these patterns are consistent across African and Asian populations.
Advances in metabolomics, such as high-throughput analytical techniques and reduced costs, allows for extensive characterization of phytochemicals across plant structures and between individual plants, and in response to environmental changes (Hao et al., 2025). As a result, metabolomics has become a useful tool for crop improvement, facilitating the analysis of intraspecific phytochemical variation to identify nutritional targets, select parental lines for breeding programmes, and monitor changes in the progeny (Fernie and Schauer, 2009; Sogbohossou et al., 2018a; Hao et al., 2025). These data can also inform strategies to retain anti-herbivory compounds during cultivar development or, if such compounds compromise palatability, guide the development of botanical pesticides (Mitchell et al., 2016; Lengai et al., 2020). Similarly, metabolomics can support drug development by enabling the identification of compounds from plant extracts for biological activity screening (Shyur and Yang, 2008). Given its wide range of medicinal uses, Gynandropsis is a strong candidate for phytomedicine research. Finally, integrating metabolomics with genomics and transcriptomics can uncover the regulatory mechanisms and biosynthetic pathways underlying phytochemical production, offering key loci to accelerate crop improvement and insights into the evolution and diversification of specialized metabolites (Hao et al., 2025).
GENOMIC AND GENETIC RESOURCES
Advancements in next-generation sequencing technologies have dramatically improved their accessibility, allowing for the research and development of non-model organisms, such as underutilized crops and taxa that possess characteristics absent in model organisms (Preston, 2021). Although work on well-established model organisms including A. thaliana, Antirrhinum majus (snapdragon; Plantaginaceae), Oryza sativa (rice; Poaceae), and Zea mays (corn; Poaceae) has significantly advanced our knowledge of plant genetics, focusing on few taxa limits our understanding of developmental networks and ability to establish a sustainable food supply as it provides a narrow representation of plant diversity (Damerval and Becker, 2017; Kamenya et al., 2021; Preston, 2021). Expanding genomic and genetic resources for non-model organisms is crucial for addressing fundamental questions in evo-devo (e.g. what are the molecular mechanisms underlying trait diversity?) and improving crops (Kamenya et al., 2021; Preston, 2021).
The order Brassicales is emerging as a model clade due to its extensive trait diversity, inclusion of A. thaliana along with several crops (e.g. rapeseed, capers, cruciferous vegetables, papaya, Gynandropsis), and rapidly expanding genomic and genetic resources (Mabry et al., 2024). Among these, Gynandropsis serves as a valuable comparative species for evo-devo research due to its contrasting traits relative to A. thaliana and other Cleomaceae species (Hoang et al., 2023; Zhao et al., 2023). In recent years, the germplasm collection of Gynandropsis has expanded significantly with the publication of two genome sequences (Hoang et al., 2023; Zhao et al., 2023) and numerous RNA sequences for a variety of plant structures, developmental stages, and experimental conditions (e.g. Zhao et al., 2023; Zenchyzen et al., 2025) (Table 2). These resources are bolstered by the close evolutionary relationship of Gynandropsis to A. thaliana, facilitating the transfer of knowledge and tools from this model species, as well as the genome sequences of Cleomaceae relatives T. houtteana and Cleome violacea L. (Cheng et al., 2013; Hoang et al., 2023; Zhao et al., 2023).
Both shared and independent polyploidy events have shaped the evolutionary history of Brassicaceae and Cleomaceae, including a whole-genome duplication (Gg–α) within Cleomaceae that is shared by Gynandropsis and T. houtteana (Mabry et al., 2020; Hoang et al., 2023). Genome and gene duplications may serve as drivers of evolutionary change, giving rise to novel traits through subfunctionalization and neofunctionalization, where duplicated genes either partition ancestral functions or acquire entirely new roles (Birchler and Yang, 2022). These events are associated with the origin of methionine-derived glucosinolates in the Cleomaceae, Brassicaceae, and Capparaceae clade – a chemical defence that diversified through an evolutionary arms race with butterflies (Edger et al., 2015; van den Bergh et al., 2016). Additionally, duplications have played a role in the independent evolution of C_4_ photosynthesis in at least three Cleomaceae species (Voznesenskaya et al., 2007; Koteyeva et al., 2011; van den Bergh et al., 2014; Huang et al., 2021; Hoang et al., 2023). While Gynandropsis has been extensively studied for its C_4_ photosynthesis mechanism (reviewed in Schlüter and Weber, 2020; Huang et al., 2023), the broader Brassicales order offers a comparative framework for understanding the evolution of different photosynthetic strategies (Marshall et al., 2007; Bayat et al., 2018; Mabry et al., 2024).
Though genomic and genetic resources are invaluable for identifying candidate genes, functional analyses are essential for confirming their roles. To facilitate such studies, a transformation system has been developed for Gynandropsis (Newell et al., 2010), and has since been used in investigations of the mechanisms underlying C_4_ photosynthesis (Huang et al., 2013; Aubry et al., 2014b; Williams et al., 2016; Reyna-Llorens et al., 2018). While stable transformation enables permanent gene edits that can be used for crop improvement, selective breeding remains a vital and cost-effective approach – especially for the often-underfunded advancement of ‘orphan crops’ (Kamenya et al., 2021; Venezia and Creasey Krainer, 2021). The development of effective breeding strategies benefits from knowledge on trait heritability and genetic variation (Kangai Munene et al., 2018; Sogbohossou et al., 2018a; Simpson et al., 2025). In Gynandropsis, several agronomically important traits, including days to flowering, plant height, and leaf biomass, have shown moderate to high broad-sense heritability and genetic advance, suggesting that selection based on phenotype is likely to produce measurable improvements in these traits over successive generations (Kiebre et al., 2017; Kangai Munene et al., 2018; Chatara et al., 2023; Mativavarira et al., 2024b; Simpson et al., 2025). Furthermore, quantitative trait loci (QTL) associated with key agronomic traits have been identified, providing the foundation for marker-assisted selection of parental lines (Simpson et al., 2025). Ultimately, integrating these genomic, genetic, and functional resources with accurate morphological descriptions and thorough chemical characterizations is essential for unravelling the mechanisms underlying trait evolution (Endress, 2002; Jeiter and Smets, 2024). This holistic approach could be utilized for crop improvement by informing the development of breeding strategies and genome editing protocols that optimize favourable traits (e.g. high leaf yield, vitamin content) (Sogbohossou et al., 2018a; Kamenya et al., 2021; Simpson et al., 2025).
MORPHOLOGICAL TRAITS
Morphological data continue to play a vital role in the molecular era of biology (Endress, 2002; Jeiter and Smets, 2024). Not only are detailed morphological studies essential for accurate species identification, but they can also provide critical insights into the form and function of plant structures (Endress, 2002; Jeiter and Smets, 2024). Although individual studies tend to focus on few features (e.g. only vegetative or only floral), data from both agricultural and evo-devo research, including several early in-depth anatomical studies, provide a cohesive picture of Gynandropsis morphology (e.g. Raghavan, 1939; Murty, 1953; Houdegbe et al., 2022b; Zenchyzen et al., 2025). Gynandropsis is an erect annual herb, reaching up to 2m in height, with variable branching (unbranched to profusely branched, bearing numerous inflorescences) (Iltis, 1960; Wu et al., 2018; Sogbohossou et al., 2019; Parma et al., 2023). The leaves are palmately compound with three to seven leaflets (typically 5-foliate), petiolate, estipulate, and alternately arranged (Iltis, 1960; Puri, 1971; Tucker and Vanderpool, 2010; Das et al., 2022). Both stems and petioles vary in vestiture (glabrous to glandular-pilose) and in colour (green to purple) (Iltis, 1960; Masuka et al., 2012; Wasonga et al., 2015; Wu et al., 2018; Sogbohossou et al., 2019). Leaflets are glabrous to pubescent and range in shape from oblanceolate to elliptic or rhombic, acuminate to acute (less often rounded) at the apex, and attenuate to cuneate at the base, with entire to serrulate or denticulate margins (Iltis, 1960; Puri, 1971; Tucker and Vanderpool, 2010; Wu et al., 2018; Sogbohossou et al., 2019; Das et al., 2022). The leaves have anatomical features associated with C_4_ photosynthesis including Kranz anatomy, characterized by a concentric arrangement of bundle sheath and mesophyll cells around vascular bundles (i.e. veins), large bundle sheath cells, few mesophyll cells between veins, and high vein density (Voznesenskaya et al., 2007; Koteyeva et al., 2011; Bayat et al., 2018). Along with the above-mentioned variation in plant form, there is considerable range in the size of vegetative and floral structures (Iltis, 1960; Wu et al., 2018; Sogbohossou et al., 2019; Houdegbe et al., 2022b).
The inflorescences are terminal racemes with numerous flowers and trifoliate bracts located at the base of each pedicel (Iltis, 1960; Zohoungbogbo et al., 2018). The flowers typically have four sepals (distinct, green, lanceolate to obovate), four petals (distinct, white or pale coloured varying from yellow to pink or purple, unguiculate with an obovate to suborbicular blade), a receptacular nectary, an elongated androgynophore and gynophore, six stamens (purple filaments), and a bicarpellate pistil (short style, capitate stigma) (Raghavan, 1939; Iltis, 1960; Tucker and Vanderpool, 2010; Das et al., 2022; Zenchyzen et al., 2023b). Flowers are monosymmetric (i.e. bilaterally symmetric/zygomorphic) at maturity due primarily to the upward curvature of the petals and stamens (Patchell et al., 2011) (Fig. 3A). However, during early organogenesis, the floral buds are polysymmetric (i.e. actinomorphic), aside from a slightly enlarged abaxial sepal (Patchell et al., 2011). Monosymmetry is established in the late stages of development when the stamen filaments are elongating and protruding adaxially out of the closed petals (Patchell et al., 2011; Zenchyzen et al., 2023a). The fertilized pistils of Gynandropsis flowers develop into dry, linear-cylindrical capsules (Iltis, 1960; Iltis et al., 2011), often referred to as pods (e.g. Zohoungbogbo et al., 2018; Wu et al., 2018) (Fig. 3B). The two valves of the capsule dehisce from the replum to disperse the seeds (Iltis et al., 2011). Unlike siliques, the characteristic fruit type of Brassicaceae, the capsules of Cleomaceae species, including Gynandropsis, lack a false septum (Hall et al., 2002; Iltis et al., 2011). Seeds are reniform to subglobose (i.e. strongly incurved with a narrow open cleft) with a rugulose to tuberculate and dark brown or black seed coat (Iltis, 1960; Iltis et al., 2011; Kwarteng et al., 2018; Blalogoe et al., 2020). While several studies provide detailed descriptions of the floral development, internal anatomy, and external morphology of Gynandropsis (Raghavan, 1938, 1939; Murty, 1953; Iltis, 1960; Patchell et al., 2011; Zohoungbogbo et al., 2018; Parma et al., 2023; Zenchyzen et al., 2023b, 2023a), we focus on floral traits associated with pollination and herbivory, as these are relevant to both agricultural and evo-devo research.
Key floral features and developmental traits of Gynandropsis gynandra. (A) The flower transitions from polysymmetric early in development to monosymmetric at anthesis, (B) has a pistil that matures into a capsule with valves that dehisce from the replum to release the seeds, (C) has notable structures including a receptacular nectary and an elongated androgynophore and gynophore, and (D) has an andromonoecious sexual system with staminate flowers and two types of hermaphroditic flowers, one with fully developed stamens and another with sessile stamens.
Floral structures of Gynandropsis that likely facilitate pollinator interactions include the petals, androgynophore, gynophore, and nectary (Fig. 3C). The androgynophore and gynophore presumably increase the likelihood of cross-pollination by optimally positioning the reproductive organs for pollinator contact and/or reduce the chance of self-pollination by spatially separating the stigma and anthers (Medan and Ponessa, 2003; Shakarishvili and Osishvili, 2013; Scorza and Dornelas, 2014; Rocha et al., 2015; Zenchyzen et al., 2023a). For instance, hawk moths with short proboscises feeding of the nectar near the base of the androgynophore contact the elevated reproductive organs while hovering over the flower (Oronje, 2011; Martins and Johnson, 2013). Although both stalk-like structures probably contribute to pollination, the androgynophore of Gynandropsis has been more thoroughly studied than the gynophore. The androgynophore is a relatively uniform cylindrical structure that rapidly elongates primarily through cell expansion during the late stages of floral development (Zenchyzen et al., 2023a). It contains a vascular cylinder, which diverges near the apex of the androgynophore into an outer ring of vascular bundles supplying the filaments and an inner ring that extends through the length of the gynophore (Raghavan, 1939; Murty, 1953; Zenchyzen et al., 2023a). This vascular pattern differs from the androgynophore of Cleoserrata speciosa, where the staminal bundles remain independent from the vascular cylinder throughout its entire length (Iltis, 1960). The receptacular nectary of Gynandropsis has an annular ring of nectary parenchyma (Murty, 1953; Zenchyzen et al., 2023b). Nectar is secreted via nectarostomata, often forming four droplets opposite the sepals (Raju and Rani, 2016; Zenchyzen et al., 2023b).
Micromorphology of the floral epidermis can influence pollinator interactions as well as provide defence against florivory and other environmental stressors (Riglet et al., 2021). In Gynandropsis, the epidermal cells of the petals, stamen filaments, and anthers are decorated with intricate striations (Zenchyzen et al., 2025). The cellular surfaces of the petal blades and anthers have disordered striations, while the petal claws and distal portion of the filaments (near the anthers) have linear (i.e. ordered) striations (Zenchyzen et al., 2025). Striations can generate structural colour through angle-dependent scattering of light, resulting in iridescence or blue halos (Moyroud et al., 2017). These optical effects may contribute to pollinator attraction by enhancing the flower’s visual signals (Moyroud et al., 2017; Riglet et al., 2021). In addition to striations, papillate epidermal cells are found on the petal claws, stigma, and sometimes the nectary of Gynandropsis flowers (Zenchyzen et al., 2025). Papillae may contribute to the visual signals and tactile cues of the flower by enhancing UV-absorption/-reflection patterns and improving grip for pollinators (Whitney et al., 2009; Alcorn et al., 2012; Schulte et al., 2019). Further, papillae can reduce the wettability of the floral structures, potentially mitigating adverse effects of surface wetness on plant–pollinator interactions, such as reduced light reflection and increased susceptibility to pathogen infection (Taneda et al., 2015). The epidermal surface of Gynandropsis flowers also includes glandular trichomes, which typically produce and store secondary metabolites (Schuurink and Tissier, 2020; Zenchyzen et al., 2025). In Gynandropsis, trichomes are located on the abaxial surface of the sepals and the valves of the pistil, presumably contributing to the defence of developing flowers and seeds against florivory (Zenchyzen et al., 2025).
The sexual system of Gynandropsis has been described as both polygamomonoecious and andromonecious, characterized by the presence of staminate, pistillate, and hermaphrodite flowers, or staminate and hermaphrodite flowers, occurring on the same plant, respectively (Zohoungbogbo et al., 2018; Parma et al., 2023) (Fig. 3D). The hermaphrodite flowers have a functional ovary and fertile stamens but vary in gynophore length, while the staminate flowers have a short or absent gynophore and a residual ovary (Raghavan, 1939; Murty, 1953; Raju and Rani, 2016; Zohoungbogbo et al., 2018; Parma et al., 2023). Flowers with sessile stamens have been described both as pistillate and hermaphroditic (Raju and Rani, 2016; Parma et al., 2023). However, Raju and Rani (2016) reported pollen production in flowers with sessile stamens, suggesting that andromonoecious is the most accurate term to describe the sexual system of Gynandropsis. In addition to the various reproductive morphs, several abnormal floral morphs have been documented (Raghavan, 1939; Murty, 1953). These include flowers with an extra sepal or petal, adnation of the petals to the androgynophore or the stamens to the gynophore and pistil, petaloid sepals, sepaloid or staminoid petals, an open ovary with exposed ovules, or a tri- or tetracarpellate pistil (Raghavan, 1939; Murty, 1953). Further research on the diverse and abundant floral morphs of Gynandropsis could be used to address questions regarding floral development and its underlying mechanisms (Meyerowitz et al., 1989), while understanding its sexual morphs is relevant for developing effective breeding programmes (Zohoungbogbo et al., 2018).
OUTSTANDING QUESTIONS AND FUTURE DIRECTIONS
Comprehensive sampling for comparative studies
Comparative studies have revealed geographical patterns in Gynandropsis phenology, C_4_ photosynthesis, morphology, phytochemistry, and pollinators, with variation between Asian, west African, and southern/east African accessions (Reeves et al., 2018; Wu et al., 2018; Sogbohossou et al., 2019, 2020; Chataika et al., 2021; Houdegbe et al., 2022a, 2022b; Zenchyzen et al., 2025). These patterns suggest that there may be genetically distinct, geographically separate subspecies of Gynandropsis, a hypothesis that could be supported with additional comparative studies using comprehensive geographical sampling. For example, genome sequences of west and southern/east African accessions would complement the existing Asian accession genome to provide insight into genomic differences and a more comprehensive picture of trait evolution and diversification. Integrated breeding programmes could utilize such knowledge while incorporating stakeholder input to define breeding objectives and develop improved cultivars (Sogbohossou et al., 2018a). Comparative studies play a crucial role in this process by identifying accessions with desirable traits for targeted breeding strategies (Chataika et al., 2021; Houdegbe et al., 2022a). Given the geographical differences, it is crucial for both agricultural and evo-devo research to specify the origin and accession of Gynandropsis plants used in studies to ensure the accurate interpretation and broader applicability of findings.
Alternative functional study approaches
Understanding the regulatory networks underlying traits is a fundamental component of evo-devo research and essential for developing efficient and effective breeding programmes (Sogbohossou et al., 2018a; Delaux et al., 2019; Kamenya et al., 2021). However, in Gynandropsis, transformation is the only established functional genomic tool for validating gene function. An alternative approach that circumvents the time-intensive development of stable transformants and allows for targeted suppression of candidate genes is virus-induced gene silencing (VIGS) (Purkayastha and Dasgupta, 2009). Cleome violacea has an established VIGS protocol, which has been used to study floral nectary initiation (Carey et al., 2021, 2023); however, its effectiveness in G. gynandra has yet to be evaluated. Expanding the functional genomic toolkit by testing and optimizing the Cleome violacea VIGS protocol for Gynandropsis would provide a valuable alternative for gene function studies. A key advantage to having an established transformation protocol is that it serves as a prerequisite for CRISPR/Cas, a powerful genome editing tool capable of generating stable and heritable mutations (Venezia and Creasey Krainer, 2021). Additionally, the availability of Gynandropsis genome sequences, along with the well-annotated genome of its close relative A. thaliana, facilitates CRISPR/Cas applications by enabling precise genomic target selection (Kamenya et al., 2021; Venezia and Creasey Krainer, 2021). The integration of CRISPR/Cas into Gynandropsis research would significantly accelerate the development of improved cultivars (Kamenya et al., 2021; Venezia and Creasey Krainer, 2021). To the best of our knowledge, Gynandropsis genome editing by CRISPR/Cas has not been documented in the scientific literature.
Building community across disciplines
Fostering collaboration across agricultural and evo-devo disciplines is key to accelerating research and maximizing the potential of Gynandropsis. Stakeholder surveys involving farmers and consumers (e.g. Sogbohossou et al., 2018b; Chataika et al., 2020; Francisco et al., 2024; Mativavarira et al., 2024a; Stoll et al., 2025) not only guide crop improvement strategies but also inform evo-devo research by identifying agriculturally relevant traits that might otherwise be overlooked. To ensure that findings are transferable, documenting the specific Gynandropsis accession studied and its seed source is key, especially given the species’ substantial phenotypic and genetic diversity. While examining a wide range of accessions is valuable for understanding intraspecific variation, focusing on accessions prioritized in breeding programmes or those with a reference genome can provide particularly meaningful insights. At the same time, consistent use of terminology and up-to-date taxonomy is critical for improving the discoverability of relevant literature. Although the term ‘orphan crop’ is commonly used to describe Gynandropsis and other underutilized species, it may not be the most appropriate due to its social connotations and inconsistent definitions (Lara et al., 2023). Alternatively, the term ‘opportunity crop’ offers a more positive and inclusive framing, highlighting the value and untapped potential of locally significant yet unimproved species (Simpson et al., 2025). Gynandropsis expands on this concept, not only offering opportunities for agricultural advancement but also serving as a focal species in evo-devo research – enhancing our understanding of complex processes such as C_4_ photosynthesis and floral development.
CONCLUSIONS
In this review, we have highlighted key areas of interest and ongoing research in agricultural and evo-devo contexts, including polyploidy, functional studies, C_4_ photosynthesis, floral morphology, sexual systems, and secondary metabolites. By integrating these shared interests with expanding genomic and genetic resources, morphological and phytochemical characterization, and documented within-species variation, Gynandropsis is well poised for synthetic investigations in crop improvement as well as plant development and evolution. To fully utilize this potential, the cohesive use of nomenclature will facilitate clearer communication, resource sharing, and data integration. Overall, improved collaboration across disciplines will accelerate the development of Gynandropsis as a model for both applied and fundamental plant science.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ajaiyeoba EO, Onocha PA, Olarenwaju OT. 2001. In vitro anthelmintic properties of Buchholzia coriacea and Gynandropsis gynandra extracts. Pharmaceutical Biology 39: 217–220. doi:10.1076/phbi.39.3.217.5936 · doi ↗
- 2Alcorn K, Whitney H, Glover B. 2012. Flower movement increases pollinator preference for flowers with better grip. Functional Ecology 26: 941–947. doi:10.1111/j.1365-2435.2012.02009.x · doi ↗
- 3Aparadh VT, Karadge BA. 2010. Fatty acid composition of seed oil from some Cleome species. Pharmacognosy Journal 2: 324–327. doi:10.1016/S 0975-3575(10)80103-8 · doi ↗
- 4Aubry S, Aresheva O, Reyna-Llorens I, Smith-Unna R, Hibberd JM, Genty B. 2016. A specific transcriptome signature for guard cells from the C 4 plant Gynandropsis gynandra. Plant Physiology 170: 1345–1357. doi:10.1104/pp.15.0120326818731 PMC 4775106 · doi ↗ · pubmed ↗
- 5Aubry S, Kelly S, Kümpers BMC, Smith-Unna RD, Hibberd JM. 2014 a. Deep evolutionary comparison of gene expression identifies parallel recruitment of trans-factors in two independent origins of C 4 photosynthesis. P Lo S Genetics 10: e 1004365. doi:10.1371/journal.pgen.100436524901697 PMC 4046924 · doi ↗ · pubmed ↗
- 6Aubry S, Kneřová J, Hibberd JM. 2014 b. Endoreduplication is not involved in bundle-sheath formation in the C 4 species Cleome gynandra. Journal of Experimental Botany 65: 3557–3566. doi:10.1093/jxb/ert 35024220652 PMC 4085951 · doi ↗ · pubmed ↗
- 7Bala A, Kar B, Haldar PK, Mazumder UK, Bera S. 2010. Evaluation of anticancer activity of Cleome gynandra on Ehrlich’s ascites carcinoma treated mice. Journal of Ethnopharmacology 129: 131–134. doi:10.1016/j.jep.2010.03.01020307641 · doi ↗ · pubmed ↗
- 8Bala A, Kar B, Karmakar I, Kumar RBS, Haldar PK. 2012. Antioxidant activity of cat’s whiskers flavonoid on some reactive oxygen and nitrogen species generating inflammatory cells is mediated by scavenging of free radicals. Chinese Journal of Natural Medicines 10: 321–327. doi:10.1016/S 1875-5364(12)60065-X · doi ↗