Combined effects of inflorescence developmental age and environmental conditions during microgametogenesis on pollen viability, germination and size in Musa acuminata ssp
Astrid Severyns, Rony Swennen, Steven B Janssens, Nico De Storme

TL;DR
The study shows how the age of banana flowers and environmental factors affect pollen quality, offering ways to improve breeding success in bananas.
Contribution
The study identifies optimal timing and environmental conditions for harvesting viable pollen in wild and cultivated bananas.
Findings
Wild bananas show reduced pollen germination with flowering duration and high light radiation.
Cultivated bananas benefit from higher temperatures during meiosis, improving pollen viability and germination.
Pollen quality can be optimized by adjusting harvest timing and environmental exposure in wild and cultivated bananas.
Abstract
Male fertility is often suboptimal in parthenocarpic banana cultivars, hindering their use as male parents in current banana breeding schemes. Next to genotype-specific traits, such as ploidy and structural heterozygosity, inflorescence developmental age and environmental factors can significantly influence male reproductive performance. In this study, pollen viability, germination capacity and diameter, were closely monitored throughout the entire male flowering phase of Musa acuminata wild diploids and diploid and triploid cultivars. Additionally, environmental parameters, including temperature, light radiation and humidity, were monitored up to 40 days before anthesis to determine their influence on microgametogenesis. Wild accessions showed a gradual reduction in pollen germination over the flowering period. High light radiation prior to pollen mitosis I and II reduced germination…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
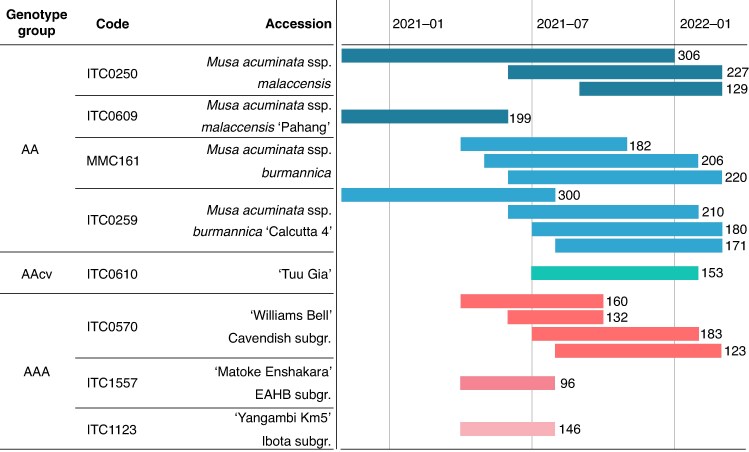 Fig. 1
Fig. 1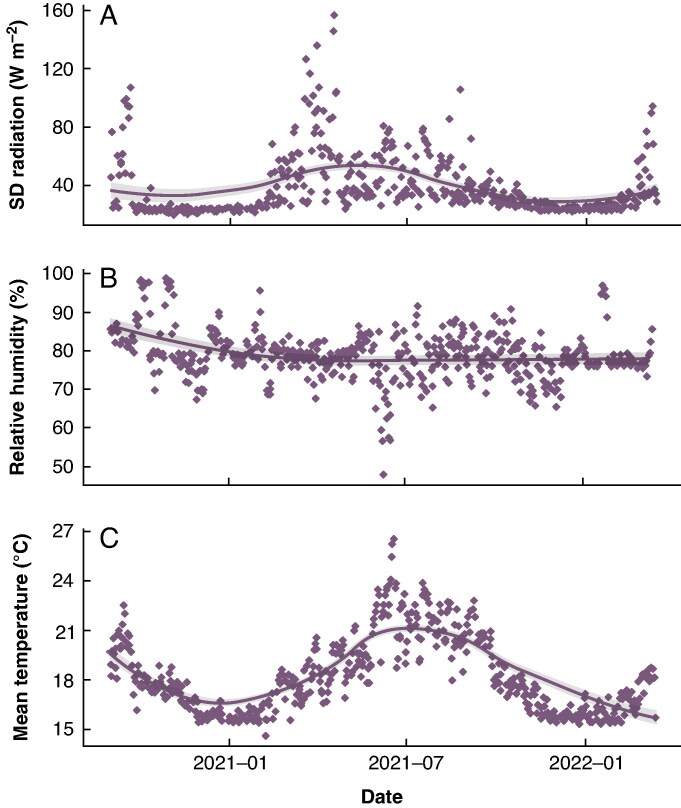 Fig. 2
Fig. 2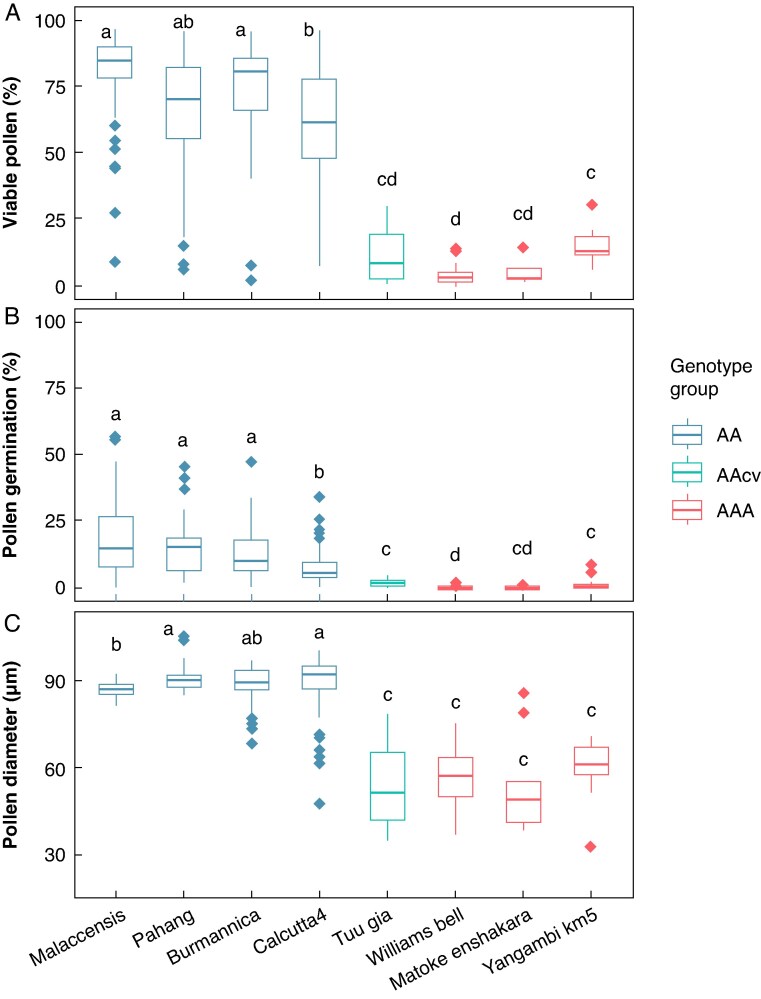 Fig. 3
Fig. 3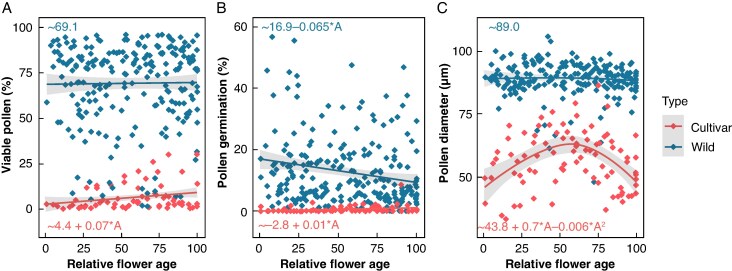 Fig. 4
Fig. 4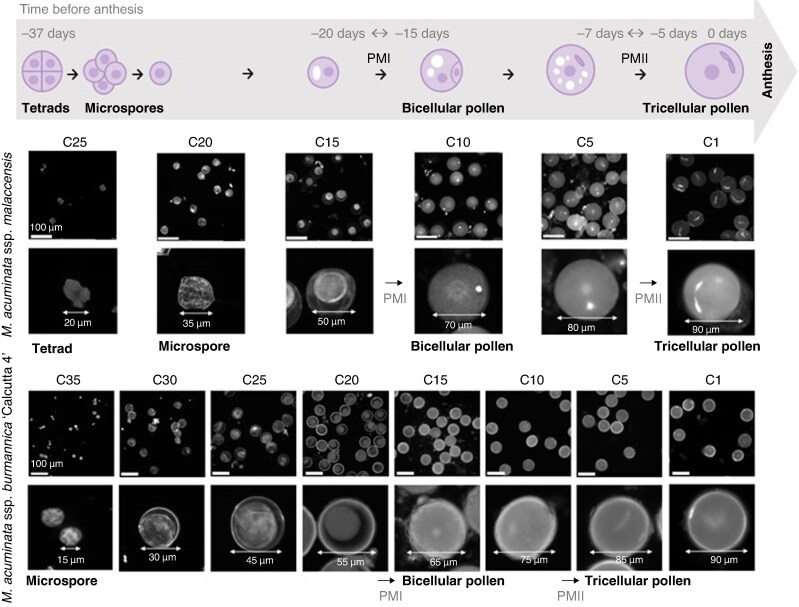 Fig. 5
Fig. 5| Variable | Timing | Set value |
|---|---|---|
| Heating | 06:00–22:00 h | 26 °C |
| 22:00–06:00 h | 16 °C | |
| Gradient, 1 °C per 15 min | ||
| Ventilation | When temperature = set + 0.5 °C | Open windows |
| Lighting | 06:00–22:00 h when natural light <450 W m−2 | 25 W m−2 additional |
| Relative humidity | Continuous | 80 % |
| Windows open | <80 % |
| Response | Term | Estimate | s.e. | d.f. |
| Pr(>| |
|---|---|---|---|---|---|---|
| Percentage viable pollen | Intercept | 40.056 | 10.522 | 65.645 | 3.807 | 0.000312 |
| s.d. rad −3 days | −0.326 | 0.069 | 221.601 | −4.714 | 4.29 × 10−6 | |
| Mean | 2.045 | 0.602 | 221.902 | 3.397 | 0.000807 | |
| s.d. rad −21 days | 0.320 | 0.089 | 221.178 | 3.570 | 0.000438 | |
| s.d. rad −37 days | −0.193 | 0.091 | 221.237 | −2.116 | 0.035462 | |
| Pollen germination percentage | Intercept | 24.919 | 11.771 | 215.540 | 2.117 | 0.035417 |
| Age | −0.082 | 0.020 | 227.756 | −4.070 | 6.50 × 10−5 | |
| Mean | 2.788 | 0.362 | 227.557 | 7.706 | 4.00 × 10−13 | |
| s.d. rad −10 days | −0.342 | 0.048 | 227.380 | −7.170 | 1.04 × 10−11 | |
| Mean RH −14 days | −0.287 | 0.076 | 227.183 | −3.755 | 0.000221 | |
| s.d. rad −24 days | −0.169 | 0.049 | 227.150 | −3.473 | 0.000616 | |
| Mean RH −37 days | −0.214 | 0.095 | 227.297 | −2.262 | 0.024620 |
| Response | Term | Estimate | s.e. | d.f. |
| Pr(>| |
|---|---|---|---|---|---|---|
| Percentage viable pollen | Intercept | −47.271 | 11.829 | 64.904 | −3.996 | 0.000167 |
| Age | 0.102 | 0.023 | 62.297 | 4.490 | 3.14 × 10−5 | |
| Mean T −37 days | 1.084 | 0.314 | 62.467 | 3.451 | 0.001006 | |
| Mean RH −37 days | 0.366 | 0.120 | 62.864 | 3.038 | 0.003468 | |
| Pollen germination percentage | Intercept | −1.919 | 0.968 | 56.537 | −1.983 | 0.05228 |
| Age | 0.010 | 0.004 | 83.628 | 2.448 | 0.01646 | |
| Mean | 0.117 | 0.043 | 83.429 | 2.715 | 0.00805 | |
| Pollen diameter | Intercept | −18.380 | 15.011 | 84.000 | −1.224 | 0.224220 |
| Age | 0.602 | 0.139 | 84.000 | 4.342 | 3.92 × 10−5 | |
| Age2 | −0.005 | 0.001 | 84.000 | −4.059 | 0.000110 | |
| Mean | 1.840 | 0.454 | 84.000 | 4.054 | 0.000112 | |
| Mean RH −27 days | 0.338 | 0.137 | 84.000 | 2.477 | 0.015247 | |
| Pollen diameter s.d. | Intercept | 28.434 | 11.283 | 15.492 | 2.520 | 0.02314 |
| s.d. rad −26 days | −0.269 | 0.080 | 83.907 | −3.337 | 0.00126 | |
| Mean | 1.044 | 0.477 | 83.443 | 2.188 | 0.03146 |
- —Bill and Melinda Gates Foundation10.13039/100000865
- —Accelerated Breeding of Better Bananas
- —Research Foundation Flanders (FWO)
- —Meise Botanic Garden
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBanana Cultivation and Research
INTRODUCTION
Bananas and plantains are among the most important crops in the world, together good for a production of almost 180 million tons in 2022, and being consumed by >400 million people daily (Brown et al., 2017; Crop Data: Banana, 2022). Any other crop of this importance has shifted towards hybrid breeding in the last few decades; however, for banana barely any improved varieties are currently in use. Banana breeding programmes have been active over the last 100 years but are hindered by low seed yield and viability (Vuylsteke et al., 1993; Batte et al., 2019). Despite the relevance of reproductive barriers in banana breeding, many factors in the pollination and subsequent process of seed formation remain insufficiently characterized. Suboptimal pollen and ovule viability related to genomic imbalances in parental lines, low receptivity of the female stigma, poor timing of pollination, lack of pollen tube growth and fertilization of ovules, and post-fertilization growth arrest in embryos all contribute to the failure of seed set in banana cultivars. However, the nature and relative share of these defects in the ultimate seed formation bottleneck remain to be determined (Uma and Arun, 2016; Amah et al., 2021).
Potential fertilization in flowering plants is a priori related to the capacity of pollen to germinate, thereby providing male sperm cells via the elongation of pollen tubes towards the embryo sac (Taylor and Hepler, 1997). Pollen germination is strongly influenced by external factors such as temperature, humidity and time of day, and by physiological factors, such as successful anther dehiscence, pollen quantity and quality, stigma receptivity, compatibility and enzymatic activity of stigmas and growing pollen tubes (Byers, 1995; Cheung, 1995; Sanzol et al., 2003; Boavida and McCormick, 2007; Distefano et al., 2012). Also, during early development of male gametophytes, factors such as temperature, drought stress and plant nutrition play an important role, because these conditions can lead to male sterility (Ito et al., 1970; Saini et al., 1984; Endo et al., 2009). Accordingly, previous research on pollen viability and germination capacity in banana has focused on separate interfering factors as conferred by differences in genotype, ploidy level, plant developmental age or environmental influences (Ortiz et al., 1998; Adeleke et al., 2004; Fortescue and Turner, 2004; Ssebuliba et al., 2008; Goigoux et al., 2013). An interpretation of the combined effect of these factors has not yet been performed.
Numerous studies in Musa have assessed pollen viability and fertility across genotypes, revealing noteworthy variations at inter- and intraspecific levels and at different ploidy levels within the Musa acuminata background (Adeleke et al., 2004; Fortescue and Turner, 2004; Soares et al., 2008; Mukasa and Rubaihayo, 1993; Bayo et al., 2024). According to Bayo et al. (2024) and Fortescue and Turner (2004), wild M. acuminata accessions (AA) show high pollen viability rates of ∼80 %, using tetrazolium chloride stain and Alexander’s stain, respectively, whereas both studies found reduced fertility in diploid cultivars (AAcv), with 40–60 % viable pollen. This variance is often attributed to the genomic make-up of parental lines and, in particular, to the interaction between contributions from multiple M. acuminata subspecies, resulting in structural heterozygosity (Shepherd, 1999; Goigoux et al., 2013; Šimoníková et al., 2020; Beránková et al., 2024). Triploid plantains (AAB) and East African Highland bananas (AAA), in contrast, typically show a high gametophytic sterility, with only 3–13 % pollen viability (Fortescue and Turner, 2004), which is attributed to their uneven number of chromosomes leading to irregularities in meiotic cell division and genomic imbalances in microgametogenesis (Shepherd, 1999).
Banana is protogynous, and its large conical inflorescence consists of enveloping bracts that contain clusters of ∼20 flowers. The inflorescence meristem continuously produces new flower primordia and is located in the centre of the cone. Towards the exterior of the cone, increasingly mature flowers are found, up to fully mature flowers in anthesis on the outside of the bud. Bracts open consecutively at an average rate of one to two per day, starting with 4–12 female flower clusters depending on the genotype, after which the meristem differentiates to male flower clusters, often with the formation of a few infertile or hermaphrodite flower clusters in between (Turner et al., 2024). Female flowers clusters remain attached to the stem to develop further into fruits in parthenocarpic genotypes and in wild genotypes if pollinated, whereas male flower clusters abscise 1–2 days after anthesis, leaving behind a scar. In general, the inflorescence continues to produce male flowers until fruit maturity and stem withering (Bakry et al., 2009). Overall, banana is outcrossing, except for some accessions with functional hermaphrodite flowers (Kallow et al., 2021). Outcrossing plants are known to show some degree of reproductive senescence (Malagon et al., 2019), and with Musa plants generating male flowers for several months, pollen performance might deteriorate throughout the flowering period. Krishnakumar et al. (1992) were first to note that in vitro pollen germination in Musa AA diploid cultivars is influenced by the age of the male bud, with weak germination of pollen obtained from the first male flowers, maximal pollen performance of flowers between clusters 20 and 30, and a gradual decline of germination capacity in the subsequent clusters. Likewise, a study by Ssebuliba et al. (2008) on triploid East African Highland bananas and the diploid cultivar ‘Pisang Lilin’ showed that flowers between clusters 30 and 40 exhibited the highest pollen viability according to acetocarmine stain. In accordance, Goigoux et al. (2013) reported a decline of pollen viability during male inflorescence development in ‘Mchare’ diploid cultivars via Alexander’s stain. In contrast, male bud age did not affect pollen viability in the wild diploid accession ‘Calcutta 4’, which showed high and uniform pollen viability along its complete rachis (Ssebuliba et al., 2008). Pollen viability in banana, as in many other plant species, is also significantly influenced by several environmental parameters. For a set of AA diploids, Ortiz et al. (1998) found that high solar radiation, high temperature, low relative humidity (RH) and low rainfall enhanced pollen viability (acetocarmine stain), whereas high rainfall and high minimum relative humidity reduced it. Uma and Arun (2016) confirmed that higher temperatures resulted in increased in vitro germination rates, although temperatures of >35 °C tended to decrease the germination percentage.
Next to pollen viability and germination, pollen size is another parameter relevant to overall pollen quality, which is also influenced by both intrinsic and external factors throughout microgametogenesis. Pollen smaller than the standard haploid pollen grain size indicates defects in pollen grain formation, resulting in small, infertile, aborted pollen (Ascari et al., 2020; Du et al., 2023). On the contrary, pollen larger than the haploid size might indicate the formation of pollen with a higher ploidy level, such as diploid (2n) pollen through meiotic restitution, a common phenomenon in plants (Brownfield and Köhler, 2011). When the number of chromosomes in a cell increases, its nucleus and cytoplasm will expand (Darlington, 1937; Karcz et al., 2000). Previous research has linked a rise in pollen diameter to an increase in pollen ploidy in bananas (Sathiamoorthy, 1994; Ortiz, 1997; Tenkouano et al., 1998). Findings by Ortiz et al. (1998) revealed that the production of 2n pollen in banana is positively associated with solar radiation, pan evaporation and temperature and negatively associated with rainfall and minimum RH.
Although various ad hoc studies have examined pollen viability, germination capacity and pollen size in banana, particularly in comparisons between different genotypes, no comprehensive study has yet explored how these parameters are impacted by the developmental age of the male flower as combined with environmental shifts occurring during male gametogenesis. In this study, we addressed this by monitoring the effect of the aforementioned factors and their putative interaction on pollen viability, germination capacity and size in a diverse set of banana genotypes, including wild diploids, a cultivated diploid and cultivated triploids.
MATERIALS AND METHODS
Biological material
Two types of plant material were collected, i.e. flowers in anthesis containing mature pollen that were used fresh for pollen viability, germination and size assessments immediately after harvest, and male flowers in different stages of microgametogenesis that were fixed for later nuclear staining.
Fresh mature flowers for pollen viability, germination and size assessments
From November 2020 to February 2022, fresh male flowers were sampled throughout male flowering from several banana inflorescences; i.e. per inflorescence, the new flower cluster in anthesis was harvested between 08:30 and 09:30 h every 2 weeks from the start until the end of the male flowering phase. During each sampling, the age of the inflorescence was determined by counting the number of male scars on the inflorescence rachis. This was done for two different accessions of both Musa acuminata ssp. malaccensis and Musa acuminata ssp. burmannica wild diploids (AA), ‘Tuu Gia’ as a diploid cultivar (AAcv), and triploid cultivar (AAA) accessions from three genetically different subgroups, namely ‘Williams Bell’, ‘Matoke Enshakara’ and ‘Yangambi Km5’ (Fig. 1). All plants were grown in full soil inside the same compartment of a greenhouse facility located in Heverlee, Belgium (50°N). Afterwards, the frequency of flower opening could be calculated by dividing the total number of days of the flowering period by the total number of scars on the rachis.
Different Musa accessions used in this study, including their genotypic group (AA, AAcv or AAA), reference code and full accession name. Each bar denotes a separate male inflorescence and the time interval during which it was monitored. The number at the right of each bar represents the total number of male nodes formed throughout the flowering period of that inflorescence.
Fixation of male flowers for assessment of microgametogenesis
To determine the time frame of microspore development in M. acuminata ssp., one M. acuminata ssp. malaccensis and one M. acuminata ssp. burmannica ‘Calcutta 4’ male flower bud were harvested in the KU Leuven greenhouse on 16 November 2020. The male flower buds were dissected by separating all enveloping flower clusters that it contained. Following the developmental gradient from the mature flower clusters at the exterior of the bud towards the flower clusters undergoing meiosis at the interior of the bud, the entire microgametogenesis of banana can be found. Every five flower clusters from the cluster in anthesis at the exterior of the bud, an entire cluster was fixed in Clarke’s fixative (75 % ethanol and 25 % acetic acid) to allow later cytological examinations. Each flower cluster contained ∼20 flowers. Following overnight fixation, the flower clusters were stored in 70 % ethanol at 4 °C. To allow additional observations of mature pollen nuclei at a different location and different time point, flower clusters in anthesis of M. acuminata ssp. malaccensis, M. acuminata ssp. burmannica ‘Calcutta 4’ and diploid AAcv cultivars ‘Pitu’, ‘Malaysian blood’ and ‘Uwati’ were harvested from plants in the field in Arusha, Tanzania (3°S), on 27 September 2021. The average temperature in Arusha in September 2021 varied around 20 °C, and the RH was ∼70 %. Solar radiation was not recorded. Following dissection of the anthers out of the flowers, they were fixed as described above.
Greenhouse conditions
Environmental conditions in the greenhouse were controlled by an automated system (Table 1). Additional lighting was provided by high-pressure sodium lamps (SON-T, Philips). Plants received nutrients (Supplementary Data Table S1) and water multiple times per day via an automated dripping system.
Environmental measurements
Every 5 min, light radiation (solar and artificial) was measured by a Quantum sensor (Sky instruments), and RH and temperature levels were recorded by a Box psychrometer (Priva). The daily s.d. in radiation intensity ranged from 20 to 160 W m^−2^, with a mean of 38 ± 21 W m^−2^ (Fig. 2A). Mean daily RH fluctuated around 80.0 ± 6.5 % but occasionally dropped to 50 % during summer months (Fig. 2B). Mean daily temperature was 18.2 ± 2.2 °C, although peaks of 35 °C were reached in summer and minimum values of 16 °C during winter (Fig. 2C).
Daily measurements of: (A) s.d. of light radiation intensity (in watts per metre squared); (B) mean relative humidity (as a percentage); and (C) mean ambient temperature (in degrees Celsius) in the greenhouse from November 2020 to February 2022. A smooth loess curve, with 95 % confidence interval in grey, was fitted to the general trend in the data.
The greenhouse climate in this study mimicked a humid subtropical climate that is found between 20° and 35° N and S latitude, with an average temperature of 27 °C during the warmest months and on average 12 °C during the coldest months, in which bananas can be managed successfully (Saúco Galán et al., 2012). In comparison to an annual average solar irradiance of 200–220 W m^−2^ in the subtropics (Loster, 2010), plants in the greenhouse received on average 220 W m^−2^ during summer; during winter, however, even with additional lighting, they received only 50 W m^−2^ on average. The latter is a major difference from the natural growth conditions of banana and should be taken into account when interpreting the results of this study.
Assessment on fresh mature pollen of pollen viability, germination and size
Every 2 weeks during flowering of the genotypes listed in Table 1, fresh male flower clusters in anthesis were harvested, after which pollen germination, viability and size were assessed immediately through in vitro pollen germination, fluorescein diacetate (FDA) staining and bright-field microscopy.
Pollen germination
The medium for in vitro assessment of pollen germination was prepared according to Brewbaker and Kwack (1963), with 0.01 % boric acid, 0.01 % potassium nitrate, 0.02 % magnesium sulphate, 0.03 % calcium nitrate, 10 % sucrose and 0.5 % plant agar, brought to a pH of 6.0. After autoclaving, 100 µL of the medium was spread over a microscopy slide and left to solidify for 5 min. Pollen of a freshly harvested flower was then picked up with a brush and distributed evenly over the medium patch. For each inflorescence, this was done for three different flowers per cluster. Next, the slides were placed inside a humid chamber and kept for 24 h at 25 °C in darkness (Nyine and Pillay, 2007; Soares et al., 2008). After overnight incubation, pollen germination was assessed quantitatively via bright-field microscopy (Olympus, Bx40) by counting the total number of pollen grains and the number of pollen tubes in four random 40× microscopy images per slide. Following Soares et al. (2008), protrusions shorter than the pollen grain diameter were not counted, under the assumption that truly elongating pollen tubes would have progressed beyond this distance after 24 h.
Pollen germination rate was calculated as:
Pollen size and viability measurements
Pollen grains were stained with FDA to assess their metabolic activity as a measure for pollen viability. For each inflorescence, pollen from three anthers from three different flowers within a cluster were taken with forceps, transferred to 1 mL of FDA staining solution and incubated for 5 min. The staining solution consisted of 900 µL of 10 % sucrose and 100 µL of 2 mg FDA per 1 mL acetone solution, with a final concentration of 0.2 mg FDA mL^−1^ (Heslop-Harrison and Heslop-Harrison, 1970). After centrifugation of the pollen solution at 30 g for 2 min, the supernatant was discarded. The pollen pellet was resuspended in 100 µL of 10 % sucrose solution and pipetted onto a microscopy slide. The pollen was imaged with a Leica DMi 8 microscope in bright-field mode to assess pollen size and in the Green Fluorescent Protein channel (450–490 nm) to measure FDA intensity. LAS X software was used to capture 18 images per microscopy slide automatically.
To identify pollen grains and perform size and FDA intensity measurements in a high-throughput manner, an image analysis pipeline was customized in CellProfiler software (version 4.2.4) (Lamprecht et al., 2007). This pipeline outputs .csv files with various size and fluorescent intensity parameters, of which the equivalent diameter and mean intensity were used in further analyses. To obtain robust pollen viability percentages based on the FDA staining assay, resulting .csv files were analysed using Rstudio software (R Studio Team, 2021). Pollen FDA intensity histograms typically showed two peaks: one at low fluorescence, representing dead pollen; and one at higher FDA intensity, indicating viable pollen. For each pollen sample, a kernel density function was used to fit a distribution on the FDA intensity histograms. Next, the R package ‘multimode’ was used to identify the exact position of the antinode between low and high FDA intensity; this value was used as a threshold to distinguish dead from viable pollen (Supplementary Data Fig. S1).
Pollen viability was then calculated as follows:
DAPI staining of fixed microspore and pollen nuclei
To determine the developmental state of microspores obtained from flower clusters at different stages throughout male bud development, their nuclear configuration was monitored using DAPI staining. Three anther pieces of length 1 mm were dissected out of three different flowers of each fixed cluster and were placed in 100 µL of DAPI staining solution, consisting of 1 mg DAPI, 1 mL EDTA and 20 µL Triton X-100 per 20 mL of 1× phosphate buffer (pH 7.4) (Park et al., 1998). Anther pieces were incubated in this staining solution for 1 h at 60 °C after which they were placed on a microscopy slide and squashed with a dentist’s hook. Finally, slides were imaged with a DMi8 microscope (Leica) using the LED405 channel (375–435 nm).
Statistical analysis
To compare the different pollen quality parameters, percentage viability, percentage germination and mean pollen diameter between genotypes, Kruskal–Wallis χ^2^ tests and post hoc Wilcoxon tests were performed in Rstudio (R Studio Team, 2021).
The effect of inflorescence age on the different pollen quality parameters was assessed with a linear mixed model using the R package lme4 (Bates et al., 2015; Pinheiro et al., 2023). The relative developmental age of each flower cluster was determined by dividing the number of scars that were present on the rachis when the cluster was harvested by the total number of scars at the end of male flowering, resulting in a scale ranging from 0 to 100. The model included genotype as a categorical random effect predictor and the relative developmental age (age) and its quadratic term (age^2^) as continuous fixed-effect predictors. Backward stepwise regression was performed to analyse whether age and age^2^ had significant effects. If not, the model was re-run without insignificant term(s), and coefficient estimates were obtained for the remaining parameters.
Environmental parameters up to 40 days prior to the pollen evaluation measurements were also added to the models as continuous fixed-effect predictors. Care was taken to avoid multi-collinearity between parameters, because they can inherently relate to each other. Prior to running the models, parameters showing least correlation were chosen, namely, daily standard deviation in radiation (SD rad; in watts per metre squared), daily mean temperature (mean T; in degrees Celsius) and daily mean RH (as a percentage), with correlations of 0.53 between mean T and SD rad, −0.22 between mean RH and SD rad, and −0.17 between mean T and mean RH. Additionally, during the model-selection step, parameters with variance inflation factor above three were removed. Again, linear mixed models were constructed via a combination of backward and forward selection using the lme4 package. Model quality was assessed by checking normality and homoscedasticity of the residuals and by conditional and marginal R^2^. Conditional R^2^ describes the proportion of variance explained by both fixed and random effects, whereas marginal R^2^ describes only the proportion of variance explained by fixed effects.
RESULTS
Genotypic differences in pollen viability, germination and pollen size
Large statistically significant differences were found between the percentages of viable pollen (χ^2^7 = 175.58, P < 2.2 × 10^−16^), percentages of pollen germination (χ^2^7 = 188.78, P < 2.2 × 10^−16^) and pollen diameters (χ^2^7 = 198.16, P < 2.2 × 10^−16^) of the genotypes included in this study. There was a clear separation between wild diploids and the diploid and triploid cultivars, with wild diploids showing significantly higher pollen viability, germination and pollen diameters than cultivars (Fig. 3). Therefore, the data were split into ‘wild’ and ‘cultivar’ for subsequent model construction to have two more homogeneous datasets that allow more accurate prediction within each of the groups.
Pollen quality parameters for the assessed varieties. Boxplots of percentage viable pollen according to FDA staining intensity (A), percentage germination (B) and pollen diameter of mature pollen (C). Genotypes showing significant differences in each of these parameters according to post hoc Wilcoxon tests are indicated by different letters.
Dynamics of pollen performance parameters over inflorescence development
For both wild accessions and cultivars, variation in the percentage of viable pollen, the percentage of pollen germination, and pollen diameter over the relative developmental age of the inflorescence was evaluated through the construction of linear mixed models. Inflorescence age did not significantly affect mean pollen viability in wild accessions, with 69.1 ± 22.6 % viable pollen throughout flowering (Fig. 4A). In cultivars, however, the developmental age significantly impacted pollen viability, with a small positive coefficient of 0.07 (s.e. = 0.025, P = 0.00734), equal to an increase of 0.07 % of viable pollen per unit of age (Fig. 4A). Accordingly, the mean percentage of viable pollen corresponded to 4.4 % at age 0 (intercept) and increased to 11.2 % by the end of male flowering. Pollen germination was impacted significantly by the developmental age of the inflorescence in both wild accessions and cultivars. In cultivars, pollen germination was overall close to zero but with a small positive effect of age (β = 0.01, s.e. = 0.004, P = 0.0296), whereas in wild accessions germination was generally much higher, starting at 16.9 %, but was affected negatively by age (β = −0.065, s.e. = 0.022, P = 0.003978), gradually decreasing towards a mean germination of 11.6 % at the end of male flowering (Fig. 4B). The mean pollen diameter of wild accessions was highly stable throughout inflorescence development, with a consistent value of 89.0 ± 6.6 µm (Fig. 4C), indicating a uniform haploid pollen population. For cultivars, the mean diameter of pollen grains was much lower and showed high variability over male flowering, as a consequence of the many small and aborted pollen grains that were formed, next to a variable fraction of pollen with a regular ‘haploid’ (90 µm) or even ‘diploid’ diameter (120 µm) (Supplementary Data Fig. S2). In general, the mean pollen diameter in cultivars was found to follow a parabolic curve over the male flowering phase, with a positive coefficient of 0.671 (s.e. = 0.151, P < 0.0001) for age and with a negative coefficient of −0.006 (s.e. = 0.001, P < 0.0001) for age^2^. The optimum of well-formed and potentially viable pollen grains in cultivars thus occurs mid-male flowering, more specifically around inflorescence developmental age 60 (Fig. 4C).
Temporal dynamics of pollen viability (A), germination rate (B) and pollen diameter (C) throughout male inflorescence development in Musa wild varieties and cultivars. The curves represent the linear mixed models fitted through the data points, with the grey area surrounding the curves indicating the 95 % confidence interval of predictions. Model formulas, with ‘A’ denoting the relative developmental age, and ‘A2’ its quadratic term, are shown in the graphs.
Time frame of male gametogenesis in Musa
Before adding the effect of environmental conditions during microgametogenesis to the models, we investigated where critical time points during male gametogenesis in Musa are situated relative to the moment of flower anthesis.
During the measuring period, M. a. ssp. malaccensis plants opened on average one flower cluster every 1.5 days, whereas for ‘Calcutta 4’, flower clusters opened daily (Supplementary Data Table S2). This indicates that ‘Calcutta 4’ inflorescences bear more flower clusters that are closer to each other in development in comparison to those in M. a. ssp. malaccensis. To confirm this, a M. a. ssp. malaccensis and ‘Calcutta 4’ male flower buds were dissected to isolate all developing flower clusters, starting from mature flowers at cluster one before anthesis to the end point of meiosis at clusters 25 and 35, respectively (Fig. 5). By correcting for one flower cluster opening every 1.5 days for M. a. ssp. malaccensis, the tetrad stage was calculated to take place 37 days prior to anthesis, closely coinciding with the early microspores found in ‘Calcutta 4’ at cluster 35 and thus 35 days prior to anthesis. With meiosis in banana during ∼2–3 days in total (Dodds and Simmonds, 1946), development from the pollen mother cell to mature pollen grain commences ∼40 days prior to anthesis. Nuclear staining of the developing microspores in the different flower clusters revealed that pollen mitosis I (PMI) occurs between flower clusters 10 and 15 in M. a. ssp. malaccensis and between clusters 15 and 20 in ‘Calcutta 4’, corresponding to a time frame between 15 and 20 days prior to anthesis (using the same correction factor for M. a. ssp. malaccensis). Pollen mitosis II (PMII) takes place between clusters 1 and 5 in M. a. ssp. malaccensis and between clusters 5 and 10 in ‘Calcutta 4’, implying that PMII takes place 5–7 days prior to anthesis.
Time line of the occurrence of distinct microspore stages in Musa accuminata ssp. malaccensis and M. accuminata ssp. burmannica ‘Calcutta 4’. Images show representative nuclear configurations of DAPI-stained microspores isolated from flower clusters in different developmental stages, where C1 represents one flower cluster before anthesis, C5 represents five flower clusters before anthesis etc., until the respective end point of meiosis marked by tetrads (C25 M. a. ssp. malaccensis) or early microspores (C35 ‘Calcutta 4’). Abbreviations: PMI, pollen mitosis I; PMII, pollen mitosis II.
To verify that PMII in M. a. ssp. malaccensis and ‘Calcutta 4’ consistently happens prior to flower anthesis, mature pollen from plants grown under different climatic circumstances in the field in Arusha, Tanzania, were subjected to nuclear staining, and although the more faint vegetative nucleus was not always visible, the germ unit was clearly composed of two separate vegetative nuclei (Supplementary Data Fig. S3A–H). Similar tricellular arrangements were visible in mature pollen of AAcv cultivars ‘Pitu’, ‘Malaysian blood’ and ‘Uwati’ (Supplementary Data Fig. S3I–T).
Overall, these data show that male gametogenesis starts as early as 40 days prior to anthesis. This is why, under the assumption of uniformity in the progression of microgametogenesis across wild M. acuminata ssp. and their derived AAcv and AAA cultivars, in the following analyses the environmental conditions 40 days prior to anthesis were included in the modelling of the three pollen quality parameters.
Impact of environmental conditions during gametogenesis on pollen viability, germination rate and pollen size
Taking advantage of the extensive data gathered on pollen size and pollen performance parameters from different Musa inflorescences that developed over a range of environmental conditions, putative effects of environmental conditions occurring during male gametogenesis (i.e. until 40 days before anthesis) on pollen viability, germination and size were determined via models. In this modelling, a distinction was made between wild M. acuminata accessions and cultivars.
The model predicting the percentage of viable pollen for wild M. acuminata accessions (Table 2) displayed a mediocre fit, with random and fixed effects combined accounting for 0.268 of variation (conditional R^2^) and with fixed effects accounting for 0.105 of variation (marginal R^2^). Although environmental conditions thus play a modest role here, radiation and temperature had significant impacts. More specifically, the radiation level during meiosis (−37 days) and 3 days before anthesis had a negative effect on the percentage of viable pollen. On the contrary, the light radiation level immediately before the first pollen mitotic division (−21 days) was found to have a positive effect on pollen viability. A higher temperature 10 days prior to anthesis also had a considerable positive effect on viability.
The model for pollen germination in wild accessions (Table 2) showed a better fit to the data, with random and fixed effects combined explaining nearly half of the observed variation (conditional R^2^ of 0.413). Fixed effects alone accounted for a marginal R^2^ of 0.243. Also here, several environmental factors occurring during microgametogenesis were found to affect pollen germination rate; in particular, radiation and mean RH had significant impacts. Germination was negatively influenced by high light radiation occurring 24 and 10 days before anthesis, corresponding to the developmental stages immediately preceding PMI and PMII, respectively. Additionally, RH at 37 days (during meiosis) and 14 days before anthesis (i.e. between PMI and PMII) negatively impacted the germination capacity of future pollen grains. Finally, temperature also influenced pollen germination rate, with especially higher temperatures occurring during PMII (−6 days) having a large positive effect.
Given that pollen diameters were highly uniform in wild accessions, no parameters were discovered that could explain variability in this trait, hence no model was constructed.
For the Musa cultivar group, the model explaining variation in pollen viability showed a relatively good fit when taking into account both random and fixed variables, with conditional R^2^ of 0.611. This, however, dropped to a marginal R^2^ of 0.184 when considering only fixed effects, indicating that a substantial part of the observed variation was attributed to the random effect of cultivar genotype. Still, a few environmental effects were uncovered, with pollen viability showing a positive relationship to the ambient temperature and RH occurring at 37 days before anthesis, corresponding to final phases of meiosis (Table 3).
Regarding pollen germination rate in cultivars, the overall fit of the model was low. A large part of the variation was again explained by differences between genotypes, whereas only a minor extent was determined by developmental age and environment (conditional R^2^ = 0.278, marginal R^2^ = 0.092). Only the temperature occurring 40 days before anthesis, i.e. corresponding to initiation of male meiosis, was positively related to the pollen germination rate (Table 3).
The significant relationship between mean pollen diameter and pollen viability observed in Musa cultivars (R = 0.47, P = 3.8 × 10^−17^; Supplementary Data Fig. S4) indicates that pollen size, besides acting as an indicator for gametophytic ploidy, is related to pollen functionality in these backgrounds. To this end, both mean pollen diameter and the associated s.d., as a proxy for the occurrence of larger sized pollen fractions, were modelled (Table 3). Owing to the quadratic term in the model for mean pollen diameter, no conditional R^2^ could be obtained; however, the marginal R^2^ was relatively high compared with the values of marginal R^2^ of other models for the cultivar group, with 0.338 of the variation explained by fixed effects. This includes strong positive effects by the mean temperature level 4 days before anthesis and by mean RH during early microspore development (−27 days). For the s.d. in pollen grain diameter, the observed variation is again largely explained by genotypic differences and only to a small extent by fixed effects (conditional R^2^ = 0.702 and marginal R^2^ = 0.052). However, some significant environmental effects were observed, with variability in pollen size, hence formation of larger pollen grains, being positively related to the ambient temperature during early meiosis (−39 days), whereas it was negatively linked to radiation in early microspore development (−26 days).
DISCUSSION
Developmental time frame of microgametogenesis in Musa
Our study provides, for the first time, an approximate time frame of microgametogenesis in M. acuminata. Cytological staging of developing male flowers revealed that initiation of meiosis occurs 40 days before anthesis and is completed ∼3 days later (i.e. tetrad stage), namely at 37 days before anthesis. Next, subsequent stages of microgametogenesis were also pinpointed, with PMI occurring between 20 and 15 days before anthesis and PMII taking place 7–5 days before anthesis. This is relatively long in comparison to the 30 days from anther primordium to anthesis in maize (Marchant and Walbot, 2022), but not abnormal, given that in apricot the development of meiocytes into mature pollen takes 30–45 days (Julian et al., 2014). Future screening of microgametogenesis stages in additional M. acuminata genotypes is needed to fine-tune the developmental time frame of male reproduction in this Musa spp. As an important observation in the present study, DAPI staining of mature M. acuminata pollen revealed that these are tricellular at the moment of anthesis, both in greenhouse conditions in Belgium and in the field in Tanzania, and not bicellular as was previously assumed (Brewbaker, 1967). Brewbaker (1967) stated that Musa pollen is bicellular, and attributed previous observations of tricellular pollen made by Schnarf (1939) to the use of old pollen. Our cytological data, however, indicate that PMII takes place ≥5 days before anthesis, hence that banana pollen grains are, by definition, trinucleate. This has major implications for the physical properties of the pollen with relevance to the conservation of banana pollen. In general, binucleate pollen undergoes maturation drying, a programmed dehydration process that occurs during pollen development and leads to desiccation tolerance. This allows binucleate pollen to remain viable over a long period of time, though these pollen grains exhibit a lower germination speed compared with trinucleate pollen (Brewbaker, 1967; Lora et al., 2009; Franchi et al., 2011). Trinucleate pollen maintains a higher water content and metabolic activity, because of which it germinates faster but survives for only a short period (Hoekstra and Bruinsma, 1975; Franchi et al., 2011; Williams and Brown, 2018). This aligns with the fact that bananas grow in humid tropical environments, where maturation drying is less necessary, and the observation that banana pollen loses its ability to germinate when dried (Youmbi et al., 2011). Future research should aim to implement cryopreservation protocols designed for short-lived, trinucleate pollen for conservation of banana pollen (Nagar et al., 2023).
From Supplementary Data Table S2 it can be deduced that the frequency of flower cluster opening is genotype dependent. The same was observed by Waniale et al. (2021) in a study on flower opening patterns in banana at two different sites in Uganda, with ‘Calcutta 4’ opening a bract every 0.7 days, ‘Mchare’ diploid cultivars and East African Highland Bananas (AAA) opening one bract per day, and Plantain cultivars (AAB) one every 1.5 days. The flower opening frequency might by influenced by environmental factors, because the ‘Calcutta 4’ in greenhouse conditions in this study opened flowers at an exact rate of one per day, whereas in Uganda they opened at a shorter interval of 0.7 days (Waniale et al., 2021). Although the difference in opening rate is minor, it might also be attributable to biological variation, given that Waniale et al. (2021) found no effect of ambient temperature or light intensity on bract opening rates in the genotypes in their study.
Dynamics of pollen viability and germination capacity throughout flowering
The male flowering period in Musa is marked by the almost daily production of flower clusters for several months up to 1 year. Our work demonstrates that wild Musa accessions showed a rather constant viable pollen percentage of ∼69.1 % and only a slight decrease in pollen germination during flowering. Likewise, Ssebuliba et al. (2008) saw a rather high and uniform pollen viability of 70–80 % according to acetocarmine staining, throughout the entire male inflorescence development in wild ‘Calcutta 4’. Despite substantial variation in our viability tests, the data obtained confirm the occurrence of a slight, yet significant, sexual degeneration in wild Musa accessions, which is typical for outcrossing plant species that have a long flowering period (Malagon et al., 2019). Notwithstanding this decline in pollen performance, enough potential to achieve successful pollination remains, even at the end of the male flowering phase.
In contrast, the percentage of viable pollen and the germination rate in Musa cultivars slightly increased throughout flowering, by 0.068 % and 0.009 % per unit of developmental age, respectively. This was also noted by Krishnakumar et al. (1992), who saw an initial increase in pollen germination in cultivars (from 20 % in the first male cluster to 30 % germination in cluster 15); unlike our results, they then observed a sharp decline in germination towards 6 % in cluster 50. Also Ssebuliba et al. (2008) saw that triploid East African Highland banana cultivars and the diploid ‘Pisang Lilin’ cultivar exhibit highest pollen viability in the mid clusters of the rachis. This initial increase in fertility is potentially attributed to the transition from female flowers (male infertile), via neutral flowers, towards male fertile flowers (Turner et al., 2024). The findings of Ssebuliba et al. (2008) and Krishnakumar et al. (1992) do coincide with our finding that most well-formed pollen grains of haploid and diploid sizes (>70 µm diameter) occur immediately after mid-flowering, at relative developmental age 60 of the male inflorescence in cultivars. The fact that we do not observe a downward trend in pollen viability and germination can be a consequence of the relatively short flowering period of most cultivars, where flowering is terminated before degeneration commences. Cultivars used in this study flowered for 4–7 months, whereas wild accessions flowered for 7–14 months (Fig. 1). Also in the study by Ssebuliba et al. (2008) cultivars produced ∼100 clusters, whereas wild diploid ‘Calcutta 4’ produced 200 clusters. We hypothesize that the earlier termination of male flowering in cultivars is related to the amount of energy that is reserved towards fruit development. In comparison to wild accessions, in cultivars a considerably larger amount of resources is diverted towards the development of the fruits, and this might negatively impact the sustenance of male flowering and overall pollen quality. Indeed, a study by Dzoyem et al. (2023) shows that following removal of female flowers soon after anthesis in cultivars, both the amount of pollen per anther and the pollen viability measured via Alexander’s stain increased significantly.
Environmental effects on pollen viability and germination
To date, studies on Musa pollen viability have taken into account the effects of environmental conditions only on the moment of anthesis (Ortiz et al., 1998; Uma and Arun, 2016). Pollen grains, however, are formed by a lengthy developmental process (Fig. 5) and are impacted by environmental conditions at several stages throughout pollen development (Ito et al., 1970; Saini et al., 1984).
Overall, in wild accessions, both pollen viability and germination are impacted by environmental parameters at several stages throughout microgametogenesis. Especially, the effect of light radiation was reoccurring. Light determines plant growth, morphology and many developmental changes (De Wit et al., 2016) and is thus, in many ways, necessary and beneficial to microgametogenesis (Quedado and Friend, 1978; Xie et al., 2014; Qin et al., 2022). However, too much light can cause accumulation of reactive oxygen species (ROS), which, in severe cases, can cause death and abortion of developing microspores (Hu et al., 2011). In line with this, pollen germination and viability were negatively linked to light radiation during early microspore development, i.e. before spores undergo PMI and are still dependent on the tapetal layer. Precocious induction of programmed cell death in the tapetal layer through ROS accumulation under strong light radiation could prevent crucial transfer of sugar and cell wall components from the tapetal layer to the developing spores, and thereby cause pollen lethality (Hu et al., 2011; Xie et al., 2014). Also at 10 days prior to anthesis, immediately before PMII, radiation showed a negative correlation with pollen germination, and again at 3 days before anthesis to pollen viability, indicating that ROS build-up in the microspore itself is also detrimental to pollen fertility (Muhlemann et al., 2018). Overall, our results show a general negative impact of high light intensity during microspore development on pollen quality in wild Musa accessions and thus contrast, in part, with findings by Ortiz et al. (1998), who reported that high solar radiation at the moment of anthesis is beneficial to pollen viability in Musa. It is likely that plants grown under a high year-round solar radiation, as in the study by Ortiz et al. (1998), have an increased antioxidant synthesis to counter the continuously high ROS levels, whereas ROS accumulation in the plants in our study might have had more severe effects from sudden increases in light intensity, affecting sensitive microspore stages (Szymańska et al., 2017; Liu et al., 2019). An increase in temperature immmediately before PMII was positively related to germination, and during PMII it was positively related to viability. A positive effect of temperature was expected, because M. acuminata is native to the humid (sub)tropics (De Langhe et al., 2009) and coincides with the positive relationship seen by Ortiz et al. (1998) between temperature at the moment of anthesis and pollen viability in wild bananas and cultivars in the field in Nigeria. For the same reason, a negative effect of mean RH during meiosis and between PMI and PMII on germination was unexpected, although also in the study by Ortiz et al. (1998) a high minimum humidity (during anthesis) decreased mature pollen viability. High relative air humidity (RH > 85 %) is known to deregulate the balance between plant CO_2_ uptake and water loss (Torre et al., 2003). Mean RH was on average 80 % during our study, but could occasionally reach ≤100 %, thereby possibly stressing the plants and impacting developing microspores. Given that the effects of environmental conditions on different stages of microgametogenesis on corresponding pollen performance observed in this study are inferred from correlations, future experimental work will have to determine whether the observed relationships can be confirmed via controlled exposure to certain environmental stresses. Especially, ROS accumulation during pollen development under high light intensities should be monitored in banana and related to the viability of the mature pollen.
In M. acuminata cultivars, we see a different response to environmental variables during pollen grain development compared with wild accessions, with less impact of radiation and more of temperature. Meiosis is clearly the most sensitive stage in terms of environmental effects on eventual pollen characteristics, whereas later stages during gametogenesis are not affected as much. Elevated temperatures during meiosis are correlated with increases in viable pollen percentage, germination percentage and s.d. in pollen size, indicative of increased formation of larger pollen grains. The association of larger pollen grains with higher temperatures aligns with previous studies that report enhanced production of unreduced (2n) pollen in banana cultivars upon exposure to high solar radiation and increased temperatures (Ortiz et al., 1998). This 2n pollen has an increased size owing to its increased DNA content. Although the occurrence of 2n pollen has not been demonstrated in the cultivars in our study, the observed correlation between temperature stress during meiosis and the eventual formation of enlarged pollen suggests instances of meiotic restitution and associated formation of 2n pollen. Given that meiotic restitution forms a natural route to restore the otherwise aneuploid karyotypic status of spores produced by triploids towards viable triploid gametes (Murthy and Tiwari, 1987), its induction in specific environmental conditions could explain the associated improvement in pollen viability. However, further in-depth analyses are needed to unravel the mechanistic link between enlarged pollen size and environmental (temperature) stress and whether and to what extent this contributes to improved pollen performance in Musa cultivars.
Conclusion
Male meiosis takes place in M. acuminata ssp. 40–37 days prior to anthesis. Pollen mitosis I and II take place between 20 and 15 days and between 7 and 5 days before anthesis, respectively. Musa acuminata pollen therefore appears to be tricellular at anthesis. Inflorescence developmental age and environmental conditions during male gametogenesis affect pollen performance simultaneously and should both be monitored to improve pollen quality. In wild accessions, pollen viability and germination gradually decrease throughout inflorescence development, whereas in cultivars the viability and germination improved slightly towards the end of flowering. Most well-developed pollen grains of haploid size or larger in cultivars were found slightly after the middle of the male inflorescence development. Pollen viability and germination in wild accessions were impacted negatively by increased light intensity during meiosis, before and during PMI, during PMII and several days before anthesis. Therefore, pollen quality in wild accessions might benefit when plants are sheltered from high light intensities. Pollen viability, germination and size were increased by elevated temperatures during meiosis in M. acuminata cultivars, implying that there might be potential for improvement of male fertility in cultivars by application of a heat treatment during meiosis.
Supplementary Material
mcaf268_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adeleke MTV, Pillay M, Okoli BE. 2004. Relationships between meiotic irregularities and fertility in diploid and triploid Musa L. Cytologia 69: 387–393. doi: 10.1508/cytologia.69.387 · doi ↗
- 2Amah D, Turner DW, Gibbs J, Waniale A, Gram G, Swennen R. 2021. Overcoming the fertility crisis in bananas (Musa spp.). In: Kema GHJ, Drenth A. eds. Achieving sustainable cultivation of bananas, Vol. 2. Cambridge: Burleigh Dodds, 457–470. doi: 10.19103/AS.2019.0054.13 · doi ↗
- 3Ascari L, Novara C, Dusio V, Oddi L, Siniscalco C. 2020. Quantitative methods in microscopy to assess pollen viability in different plant taxa. Plant Reproduction 33: 205–219. doi: 10.1007/s 00497-020-00398-633123804 PMC 7648740 · doi ↗ · pubmed ↗
- 4Bakry F, Carreel F, Jenny C, Horry J-P. 2009. Genetic improvement of banana. In: Jain SM, Priyadarshan PM. eds. Breeding plantation tree crops: tropical species. New York: Springer, 3–50. doi: 10.1007/978-0-387-71201-7_1 · doi ↗
- 5Bates D, Mächler M, Bolker BM, Walker SC. 2015. Fitting linear mixed-effects models using lme 4. Journal of Statistical Software 67, 1–48. doi: 10.18637/jss.v 067.i 01 · doi ↗
- 6Batte M, Swennen R, Uwimana B, et al 2019. Crossbreeding East African Highland Bananas: lessons learnt relevant to the botany of the crop after 21 years of genetic enhancement. Frontiers in Plant Science 10: 81. doi: 10.3389/fpls.2019.0008130804965 PMC 6370977 · doi ↗ · pubmed ↗
- 7Bayo SJ, Massawe V, Ndakidemi PA, et al 2024. Pollen amount and viability in Mchare and selected wild (AA) banana (Musa acuminata) genotypes: prospects for breeding. Hort Science: A Publication of the American Society for Horticultural Science 59: 632–638. doi: 10.21273/HORTSCI 17608-23 · doi ↗
- 8Beránková D, Čížková J, Majzlíková G, et al 2024. Striking variation in chromosome structure within Musa acuminata subspecies, diploid cultivars, and F 1 diploid hybrids. Frontiers in Plant Science 15: 1–14. doi: 10.3389/fpls.2024.1387055 PMC 1125541039027673 · doi ↗ · pubmed ↗