Assessing sexual dimorphism in the common vampire bat, Desmodus rotundus
Analorena Cifuentes-Rincon, Karen D. Sarmiento-Arias, Diego Soler-Tovar, Abelardo Rodríguez-Bolaños, Carlos Bravo-Garcia, Nicolas Reyes-Amaya, Laura Ávila-Vargas, Luis E. Escobar

TL;DR
This study finds that female common vampire bats are generally larger than males, with significant differences in forearm and tibia lengths.
Contribution
The study provides robust evidence of sexual dimorphism in D. rotundus using multivariate and univariate analyses with historical and contemporary data.
Findings
Females had significantly longer forearms than males (mean 61.8 mm vs. 58.5 mm) with non-overlapping confidence intervals.
Tibia length also differed significantly between sexes in wild-caught individuals, with females showing greater values.
Sex explained 25.7% of the variation in forearm length, with forearm dimorphism showing the strongest signal.
Abstract
Sexual dimorphism in bats is understudied, with conflicting evidence across species and geographic regions. For Desmodus rotundus, the common vampire bat, previous reports on morphological sex differences have been inconsistent. This study aimed to assess sexual dimorphism in D. rotundus using a combination of contemporary field measurements and historical museum specimens. We analyzed six morphometric traits, including body mass, head length, body length, tibia length, ear length, and forearm length. Data were collected from 46 wild-captured individuals from five locations across Colombia in South America. Additionally, forearm length was examined in an expanded dataset of 490 specimens, including additional 444 individuals from museum vouchers collected over the past century. Principal components analysis and hierarchical clustering of the six-trait dataset showed patterns of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
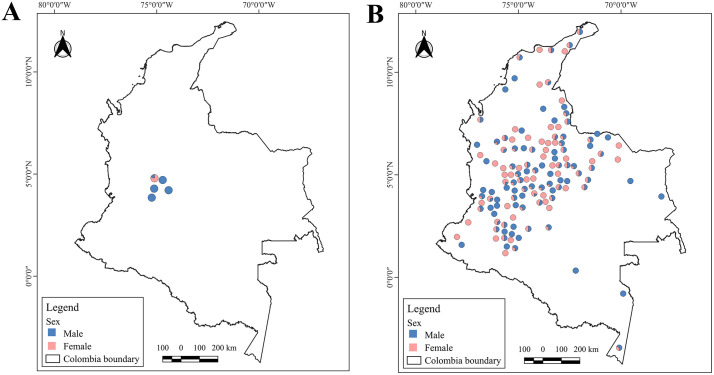 Fig 1
Fig 1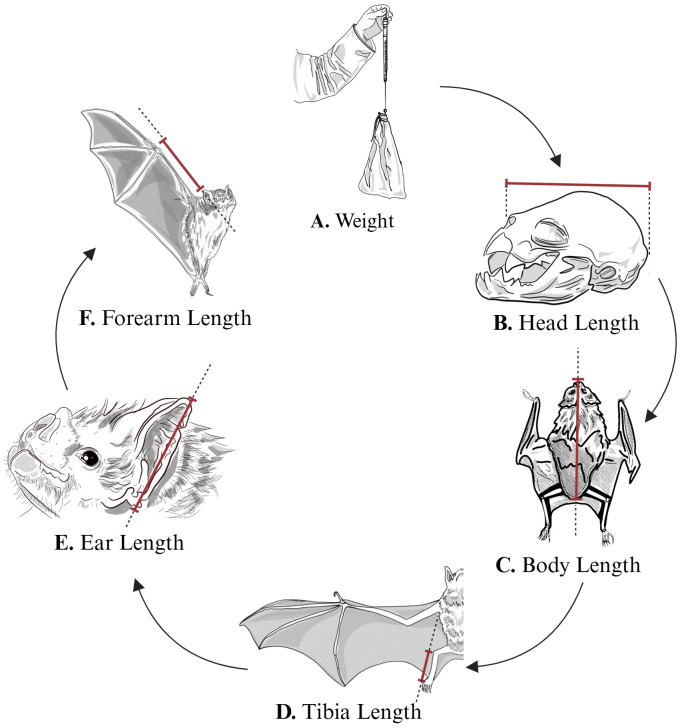 Fig 2
Fig 2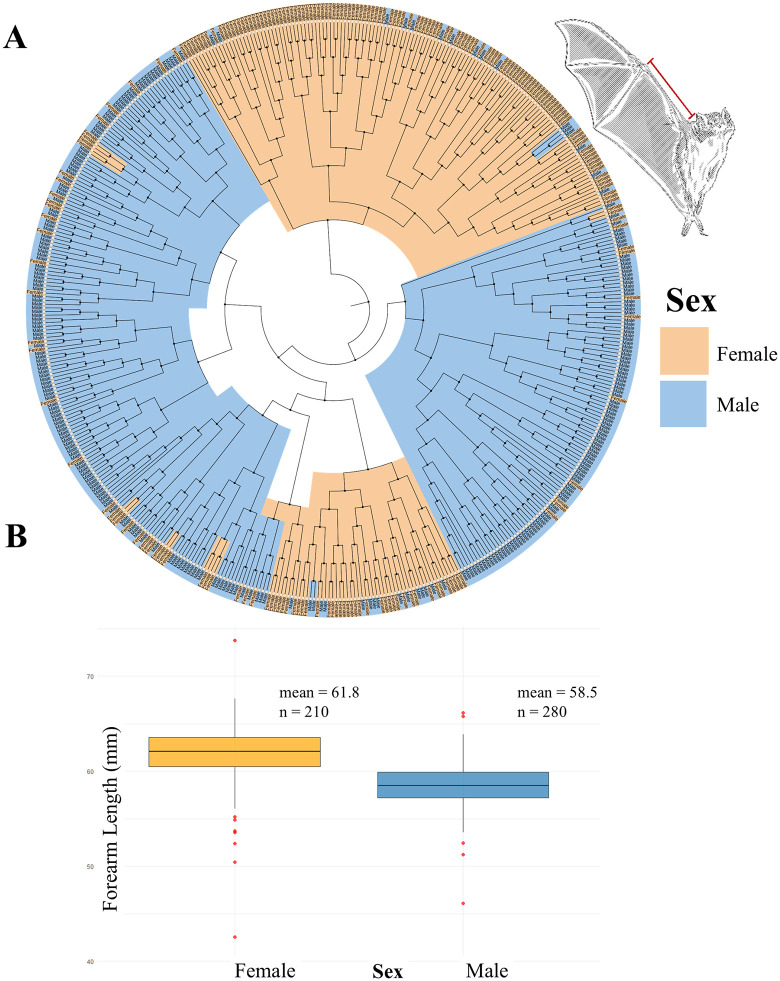 Fig 3
Fig 3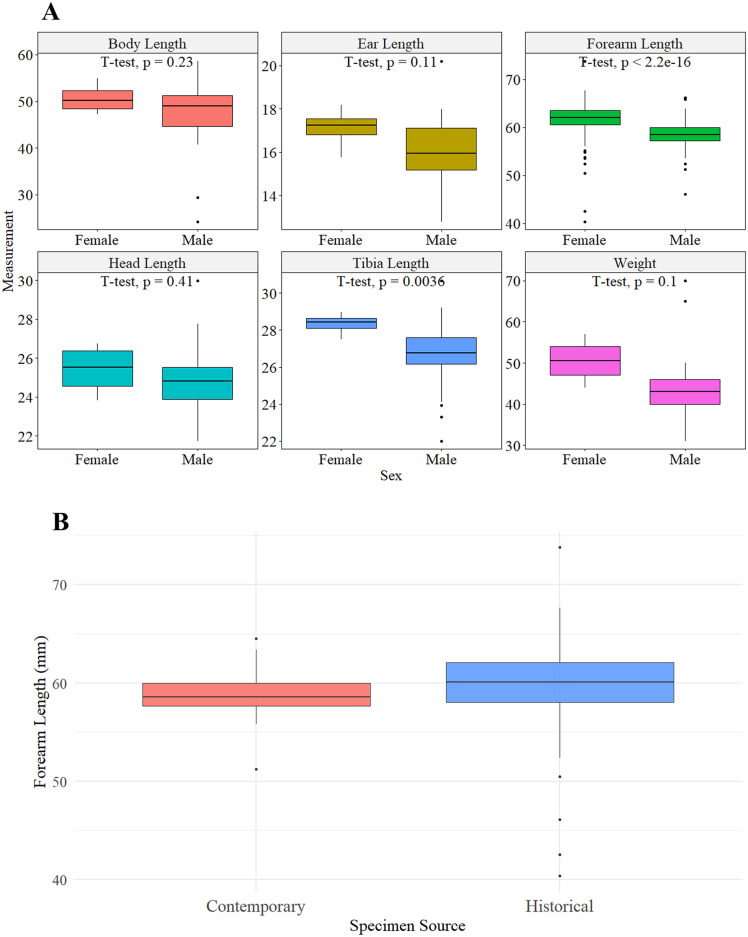 Fig 4
Fig 4- —http://dx.doi.org/10.13039/100000001National Science Foundation
- —http://dx.doi.org/10.13039/100000001National Science Foundation
- —http://dx.doi.org/10.13039/100014523Institute for Critical Technology and Applied Science
- —http://dx.doi.org/10.13039/100000060National Institute of Allergy and Infectious Diseases
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBat Biology and Ecology Studies · Amphibian and Reptile Biology · Rabies epidemiology and control
Introduction
Latin America is home to the only sanguivorous bat species in the world: Diphylla ecaudata (Spix, 1823), Diaemus youngi (Jentink, 1893), and Desmodus rotundus (E. Geoffroy, 1810) [1]. Among sanguivorous bats, D. rotundus is unique for being the only extant member of its genus and for specializing in feeding on mammals [2,3]. Its distribution spans from northern Mexico through Central America and much of South America [4–6], occupying a variety of habitats, from sea level to elevations over 3600 meters [7], including deserts, savannas, and tropical forests [8–10]. Its ability to thrive in disturbed environments, especially areas with livestock, underscores its ecological plasticity [3,11,12].
Desmodus rotundus is also a highly social species, living in colonies typically composed of a dominant male and several females [13]. Its reproductive biology is flexible, with breeding occurring year-round but peaking during rainy seasons, when an influx of juveniles can reshape colony dynamics [14–16]. The species’ obligate diet of blood has long intrigued researchers and generated public concern, particularly in rural areas of Latin America where D. rotundus can transmit rabies virus and other zoonotic pathogens to different species [6,15,16]. As a result, D. rotundus is not only ecologically significant but also a focal species in public health and wildlife disease management efforts.
Beyond its importance to public health, D. rotundus has drawn scientific interest due to its potential for use in biomedical research across Neotropics [6,15,17,18]. Recent studies have identified bioactive compounds in D. rotundus saliva that show promise in the treatment of cardiovascular diseases, such as stroke and hypertension [16,17,19–24]. These discoveries reflect the importance of studying D. rotundus from a public health perspective, as an important species for ecosystem services, and as a source of medical innovation [25–27].
In D. rotundus, sexual differences extend beyond morphology to include reproductive roles, social behavior, and feeding strategies. Females are often more involved in social cohesion activities such as grooming and regurgitative food sharing [28,29], while males invest more in maintaining dominance and reproductive access within colonies [11,14]. While behavioral, immunological, and physiological aspects of D. rotundus have received considerable attention, its morphological variation, including sexual dimorphism, remains poorly understood.
Current literature on sexual dimorphism in D. rotundus presents conflicting results. For instance, some studies report that females are larger than males, especially in traits linked to body size [30–33]. Other studies, however, suggest that males exhibit larger traits due to sexual selection and competitive behaviors [34]. Additional reports find no significant sexual differences, suggesting that environmental variables such as food availability may play a more dominant role [35]. These inconsistencies highlight a gap in our understanding of sexual dimorphism in D. rotundus and the need for a broader, more geographically inclusive analysis.
In this study, we explore sexual dimorphism in D. rotundus using a comprehensive morphological dataset that includes recently collected specimens (2022–2023) and historical museum records spanning over a century (1921–2023) from 11 biological collections. We focus on populations from Colombia, a country that offers a wide range of ecological and elevational conditions, to investigate whether consistent patterns of dimorphism emerge. Our aim is to contribute to a more nuanced understanding of D. rotundus’ morphological variability and its relevance to evolutionary biology, species management, and biomedical studies.
Materials and methods
We collected morphometric data from two distinct data sets (contemporary and historical) to evaluate sexual dimorphism in Desmodus rotundus. The contemporary dataset comprised 46 live-captured individuals from fieldwork at five field sites from 2022 to 2023 across Colombia (Fig 1A). The historical dataset included 444 adult D. rotundus from museum specimens. Combined, we obtained data from 490 adult D. rotundus, including the 46 live individuals with 444 museum specimens.
Geographical representation of the records of Desmodus rotundus in Colombia, represented by sex.(A) Contemporary dataset. Geographic distribution of 46 D. rotundus individuals from which six morphological traits were collected (i.e., weight, head length, body length, tibia length, ear length, and forearm length.) in the departments of Tolima and Cundinamarca. (B) Contemporary and historical dataset. Distribution of 490 D. rotundus museum specimens from which forearm measurements were collected for this study. Orange: female. Blue: male. Map was created using QGIS software/ Open source geospatial foundation project [36].
Ethics statement
Bat capture and sampling procedures were conducted in accordance with ethical guidelines for wildlife research and International Guiding Principles for Biomedical Research Involving Animals [37]. Protocols followed recommendations from the American Society of Mammalogists [38], and all methods were approved by the Virginia Tech Institutional Animal Care and Use Committee (IACUC-21–138) in the US, and Universidad de La Salle in Colombia under permit 1473 for the Collection of Specimens of Wild Species. Sampling to generate a contemporary dataset was carried out in compliance with national and international regulations for the ethical treatment of wild animals.
A veterinarian was present for all procedures to provide oversight and ensure appropriate handling, immobilization, anesthesia, euthanasia, and implementation of biosafety precautions to prevent human exposure to anesthetic agents [39].
Bat capture and sampling methods
Bats were collected using three capture methods (i.e., conventional mist nets, harp trap, and cone trap) and placed in disinfected cloth bags [40,41]. To minimize stress and potential harm during processing, bats were handled gently and restrained carefully by trained personnel. Handling time was minimized (approximately 2 minutes per individual).
We selected weight, head length, body length, tibia length, ear length, and forearm length as focal metrics to assess sexual dimorphism from the contemporary dataset (Fig 2). These measurements are standard in bat taxonomy and provide a robust source for exploring morphological variation [42–45]. Each metric was chosen based on its biological relevance, accuracy in differentiating sexes, and frequent use in bat studies [46].
Morphometric measurements of Desmodus rotundus.Six key morphological traits were measured in individual bats: (A) Weight, measured using a digital balance, recorded in grams (g). (B) Head length, from the tip of the nasal leaf to the posterior edge of the skull (in mm). (C) Body length, from the tip of the nose to the base of the tail. (D) Tibia length, measured from the proximal end of the tibia to the posterior base of the calcar (in mm). (E) Ear length, from the base of the ear to the tip (in mm). (F) Forearm length, from the wrist to the elbow when the wing is folded (in mm). Red lines indicate the specific landmarks used for each measurement to visually demonstrate the precise locations.
A spring balance was used to weigh individuals, and the breeding stage was assessed. Adults were classified based on their reproductive condition (females were classified as active or inactive, lactating or pregnant, and males as having inguinal or descended testicles) [7,47]. Morphometric data were collected using a Uline digital caliper (accuracy: 0.0005 mm), and only adult and semi-adult individuals were included. Sexually immature or pregnant specimens were excluded to avoid confounding effects. Specimens were confirmed to be the species D. rotundus using broadly accepted taxonomic keys [7,47,48].
Anesthesia and lethal sampling protocol
Specimens included in this study were obtained from individuals that had been lethally sampled as part of independent virological surveillance studies aimed at detecting rabies virus circulation in the study areas. Morphological measurements were taken prior to euthanasia, and no animals were euthanized specifically for the purpose of this morphological study.
For euthanasia, bats were gently placed inside a Ziploc-type plastic bag containing a cotton pad pre-soaked with 2 mL of USP-grade isoflurane, positioned in a corner to avoid direct contact with the animal. The bag was sealed immediately, and bats were monitored for signs of anesthesia (e.g., decreased responsiveness to stimuli, loss of righting reflex, and slowed respiration). Once deep anesthesia was confirmed (absence of reflexes), cardiac puncture was performed for exsanguination, followed by cervical dislocation to ensure death. Death was confirmed by the absence of heartbeat, respiration, and the presence of fixed, dilated pupils.
Organs were extracted for virological analyses, and voucher specimens were prepared and deposited at Colección de Mastozoología del Museo de La Salle – Bogotá, Colombia (MLS).
Historical specimen collection
In bats, forearm length which inludes the segment of the wing extending from the elbow to the wrist, is the most reliable and widely used measurement to assess size [49,50]. This metric is diagnostically valuable due to its relevance to aerodynamics, ecological adaptation, geographic variation, and sexual dimorphism [51]. For this study, we obtained additional forearm length measurements from 444 specimens resulting in a total sample size of n = 490 (210 females and 280 males), including 444 historical specimens and contemporary bats (Fig 1B).
For museum specimen collection, we visited 11 museums in Colombia, including Colección de Mastozoología del Museo de La Salle (MLS-mam) [52], Colección de Mamíferos, Museo de Historia Natural C.J. Marinkelle, Universidad de los Andes (ANDES-M) [53], Colección de Mamíferos “Alberto Cadena García” Instituto de Ciencias Naturales, Universidad Nacional de Colombia (ICN) [54], Colección de Mamíferos del Instituto Humboldt (IAvH-M) [55], Colección de Mamíferos, Museo de Ciencias Naturales de La Salle del Instituto Tecnológico Metropolitano (CSJ-m) [56], Colección de Mamíferos del Museo de Historia Natural de la Universidad de la Amazonia (UAM-M), Colección Mastozoológica, Museo de Historia Natural Unillanos (MHNU-M) [57], Museo de Historia Natural de la Universidad Industrial de Santander (MHN-UIS), Colección Zoológica de la Universidad del Tolima (CZUT), Colección de Mamíferos, Museo de Historia Natural Universidad de (Caldas -Uca) [58], and Museo de Historia Natural de la Universidad Distrital Francisco José de Caldas (MHNUD-M) [59]. We measured the forearm length of specimens of D. rotundus from museum collections between October 2022 and April 2023. All forearm length measurements were taken using a 0–150 mm (6″) electronic digital caliper with an accuracy of 0.01 mm. Forearm data were employed alone to estimate sexual dimorphism from these and wild individuals across Colombia (Fig 1B).
Data preprocessing and cleaning
All measurements were inspected for consistency, and individuals with missing or unclear sex assignment or forearm measurements were excluded. For the multivariate analysis, only the 46 live individuals with complete morphometric data were used. For the univariate analysis focused on forearm length, the complete sample of 490 individuals was analyzed.
Statistical analyses
Principal component analysis.
We performed a Principal Component Analysis (PCA) to explore multivariate patterns of sexual dimorphism using six external morphological variables collected from the 46 contemporary individuals. Measurements were scaled and centered prior to analysis to ensure comparability. PCA was conducted using the ‘prcomp’ function in R v4.4.0, and results were visualized with the ggplot2 and factoextra packages [60]. The first two principal components were plotted to evaluate clustering by sex.
Finally, the correlation between ear and forearm, forearm and weight, and body and weight measurements were measured to corroborate the influence of the variables.
Univariate analyses.
We conducted two-sample Welch’s t-tests comparing males and females to assess sexual dimorphism in forearm length across the larger sample (n = 490). Prior to the test, we assessed data for normality (via Q-Q plots) and homogeneity of variances.
A simple linear regression model was used to assess the effect of sex on forearm length. The model was fitted using the ‘lm’ function in R, with sex as a categorical predictor and forearm length as the response variable. The significance of the predictor variable (sex) was evaluated using the associated p-value of the regression coefficient, with a threshold of α = 0.05. Model fit was assessed using the coefficient of determination (R²), and the residual standard error was used as an estimate of the variability in forearm length not explained by sex.
Comparisons of the six morphometric traits between sexes were conducted using Welch’s t-tests [61]. To account for multiple testing, p-values were adjusted using the Benjamini-Hochberg false discovery rate (FDR) procedure [62], with Bonferroni correction applied for comparison. Differences between sexes were further tested using a MANOVA including all six morphometric traits simultaneously [63].
Descriptive statistics.
We calculated sample size (n), mean (m), standard deviation (SD), interquartile range (IQR), and 95% confidence intervals (CI) for forearm length by sex using the dplyr and ggplot2 packages in R [60]. These descriptive statistics provided context for interpreting the degree and consistency of sexual dimorphism.
Hierarchical clustering.
Hierarchical clustering was performed using the dendextend package, ‘hclust’ function in R to further evaluate morphometric differentiation between sexes [64], on the scaled morphometric data using Euclidean distances and Ward’s method (ward.D2). Pearson correlation coefficients were calculated to assess associations among traits and selected pairwise correlations were tested using ‘cor.test’.
Two separate dendrograms were generated from the hierarchical clustering. One dendogram was based on the six scaled morphological variables from the 46 contemporary individuals, offering a multivariate perspective on sex-based clustering. The second dendogram was created using the scaled forearm length data for all 490 individuals, enabling visualization of sexual dimorphism in this specific trait across a broader sample. In both cases, the resulting dendrograms were annotated by sex to assess clustering patterns visually.
The hierarchical clustering was converted into a phylogenetic tree through the ape package in R [65]. Finally, the hierarchical tree was edited in the Interactive Tree Of Life (ITOL V6) online program [66].
Comparative analysis between historical and contemporary specimens.
Morphological data were analyzed from two groups of individuals grouped as historical specimens from museum collections and contemporary individuals captured in the wild. The dataset included measurements of six morphological traits, including head length, body length, tibia length, ear length, body weight, and forearm length. Sex was recorded as a categorical variable (i.e., Male, Female).
Analyses were conducted in R (version 4.4.2), using the packages readr, dplyr, tidyr, ggplot2, and ggpubr. Initially, sex was converted to a factor for group comparisons. Descriptive statistics (mean values) were calculated separately for males and females.
Welch’s two-sample t-tests were performed for each morphological trait comparing male and female individuals, accounting for unequal variances to assess sexual dimorphism. The results were organized by p-values to identify significant differences.
Furthermore, we compared morphological differences between historical (museum) and contemporary (wild) specimens. Two-sample t-tests were applied within each sex group to compare forearm lengths between historical and contemporary individuals.Data visualization included boxplots of morphological traits by sex, created with ggplot2. Faceted boxplots displayed trait distributions stratified by sex, while separate boxplots compared forearm lengths between historical and contemporary specimens.
Some data has been provided via this manuscript and the associated Supporting Information files. Other data can be found via Figshare at: https://figshare.com/articles/dataset/Morphological_data_i_Desmodus_rotundus_i_csv/27858219?file=51814880.
Results
To assess sexual dimorphism in Desmodus rotundus, we performed two statistical analyses: one incorporating a series of morphological metrics and another focusing exclusively on forearm length. Welch’s t-tests indicated significant sexual dimorphism in forearm length (p = 6.8 × 10 ⁻ ⁵; FDR-adjusted p = 4.1 × 10 ⁻ ⁴; Bonferroni-adjusted p = 4.1 × 10 ⁻ ⁴) and tibia length (p = 0.003; FDR-adjusted p = 0.011; Bonferroni-adjusted p = 0.022). No other traits remained significant after correction for multiple testing.
A MANOVA on the six morphometric traits (body weight, head length, body length, tibia length, ear length, and forearm length) indicated a significant overall effect of sex (Pillai’s trace = 0.389, F(6,39)=4.14, p < 2.2 × 10 ⁻ ¹⁶). A few outliers (n = 12) were found in individuals from different localities. These outliers showed morphological differences from the bulk of samples and were retained in all analyses.
Forearm length measurements (n = 490) revealed that females were larger than males (t = −12.677, df = 366.8, p < 2 × 10 ⁻ ¹⁶). Females (n = 210) exhibited a significantly greater mean forearm length (61.79 mm, SD = 3.37) compared to males (n = 280; m = 58.50 mm, SD = 2.21). The 95% confidence interval for females ranged from 61.33 to 62.25 mm, while for males it ranged from 58.24 to 58.76 mm. Interquartile ranges were 3.05 mm for females and 2.69 mm for males, suggesting slightly greater variability in female measurements. Furthermore, hierarchical clustering based on forearm length show a separation between males and females (Fig 3A and 3B).
Comparison of forearm length in Desmodus rotundus by sex.(A) Distance tree by cluster analysis of D. rotundus according to forearm length from 490 historical and contemporary specimens. Orange: females. Blue: males. Note that the data tended to cluster in two groups based on sex. (B) Boxplot illustrating sexual dimorphism in forearm length. The median (central line) is higher in females than in males, indicating generally longer forearms in females. The 95% confidence interval for females spans 61.33–62.25 mm, and for males, 58.24–58.76 mm. The interquartile range (IQR) is slightly larger in females (3.05 mm) than in males (2.69 mm), suggesting greater variability within the female group. Outliers (red points) lie beyond the whiskers and represent individuals with forearm lengths notably outside the typical range for their sex. Mean forearm lengths were 61.8 mm for females and 58.5 mm for males.
We found a significant effect of sex on forearm length (t = −13.0, p < 2 × 10 ⁻ ¹⁶). The estimated mean forearm length for females was 61.8 mm while for males it was 58.5 mm, indicating that females have, on average, forearms 3.29 mm longer than males.
The model using sex as the only predictor explained 25.7% of the total variance in forearm length (R² = 0.26). The residual standard error was 2.77 mm, suggesting a moderate spread of individual values around the predicted means. These findings provide strong statistical support for sexual dimorphism in forearm length in this species, with females exhibiting significantly greater values than males.
Comparisons between historical and contemporary specimens were conducted using student’s t-tests. Of the six measurements evaluated, only two showed statistically significant differences between sexes, forearm in historical specimens and tibia length, in contemporary specimens (Fig 4A).
Morphometric comparisons by sex and origin in Desmodus rotundus.(A) Boxplots of six morphological traits (body length, ear length, forearm length, head length, tibia length, and body weight) comparing females and males. Each panel includes the result of a student’s t-test evaluating sexual dimorphism. Significant differences between sexes are observed in forearm and tibia length. (B) Comparison of forearm length between historical specimens and contemporary bats. Historical specimens tend to show a slightly broader range of variation. Forearm length was significantly greater in females than in males (t-test, p < 2 × 10 ⁻ ¹⁶), indicating a clear pattern of sexual dimorphism in this trait. Similarly, tibia length was greater in females, with a statistically significant difference (t-test, p = 0.0036). In contrast, no significant differences were observed between sexes in body length (p = 0.23), ear length (p = 0.11), head length (p = 0.41), or body weight (p = 0.1), although females tended to weigh more than males.
Distribution of morphological traits by sex reveal sexual dimorphism based on forearm length of historical and contemporary specimens (Fig 4A and 4B).
There was no difference between historical (museum) and contemporary (wild-caught) specimens for females (t = −0.93, df = 208, p = 0.355), and males (t = −0.01, df = 278, p = 0.992).
Discussion
Our study revealed female-biased sexual dimorphism in Desmodus rotundus in Colombia. Through both multivariate and univariate analyses, we found that D. rotundus females exhibit significantly larger forearm and tibia lengths compared to males. These findings align with prior reports of female-biased dimorphism in D. rotundus [30,31,67] and add to the growing body of literature on sex-based morphological variation in bats.
As an exploratory approach, we performed a principal components analysis and hierarchical clustering on a subset of 46 live-captured D. rotundus individuals with full morphometric data (see S1 Fig). Given the limited sample size and strong sex imbalance (only 4 females), results are not presented in the main text but may offer preliminary insights into multivariate morphological variation.
The second analysis, focused solely on forearm length in a larger sample of 490 individuals, reinforced these patterns with greater statistical power. Females had significantly longer forearms than males, with non-overlapping confidence intervals and consistent results across t-tests, linear regression, and clustering methods (Fig 3). The linear model explained over 25% of the variance in forearm length with sex as the sole predictor, a considerable proportion for a single biological variable, underscoring the strength of this dimorphic pattern in bats, as suggested by other authors [68–70].
In addition to multivariate methods, we performed univariate comparisons using student’s t-tests across six morphometric traits, including body length, ear length, forearm length, head length, tibia length, and weight, combining historic museum specimens and contemporary wild-caught animals (Fig 4). Significant sexual dimorphism was observed in traits forearm length and tibia length, with females being larger in both traits. No significant sex differences were found in other body metrics (i.e., body length, ear length, head length, and weight), although females tended to weigh more than males (Fig 4A). These findings further support a consistent pattern of dimorphism centered on locomotor traits rather than general body size, as observed by other authors [71,72].
Female-biased dimorphism in bats has been widely attributed to reproductive demands such as pregnancy, lactation, and pup rearing, which place substantial energetic and physiological burdens on females [73,74]. Larger body size in females may facilitate energy storage, thermoregulation, and load-carrying capacity during flight [73,74]. Our findings support this hypothesis and suggest that selective pressures, likely driven by ecological habits, are acting on D. rotundus, potentially leading to regional variation in forearm length, as has been observed in other Neotropical bat species [70–72]. This regional variation in forearm length should also be interpreted in the context of environmental and ecological variability. D. rotundus inhabits a wide range of environments—from tropical lowlands to montane regions—where factors such as prey availability, temperature, seasonal rainfall, and roosting conditions may impose distinct selective pressures that shape local morphology [75–77].
The presence of morphological outliers across several localities highlights the extent of individual variation within D. rotundus and likely reflects a combination of biological and ecological factors [78,79]. In total, we identified 12 outlier specimens—7 males and 5 females—distributed across 11 different localities. These individuals exhibited forearm lengths that fell beyond the expected range (based on the 1.5 × IQR rule) for their respective sex and locality groupings.
The most extreme outlier was a female from Agua de Dios (Cundinamarca) with a forearm length of 73.79 mm, more than 10 mm above the upper bound for females. This specimen, along with another unusually large female from Medina, Cundinamarca (67.65 mm), suggests the possibility of localized phenotypic divergence. These sites may exert selective pressures that promote larger body size, potentially in response to ecological strategy such as thermoregulatory demands or prey availability, as suggested by other authors [80–83]. Similarly, male outliers with notably small forearms, such as those from Chaparral (51.24 mm) and Venadillo Tolima (53.86 mm), and Tocaima, Cundinamarca (52.44 mm), also raise the possibility of ontogenetic or nutritional influences as evidenced in other bat species [84–86].
Given the wide ecological heterogeneity across Colombian landscapes, including transitional zones between lowland and montane habitats, there are unique opportunities to capture ecogeographic structure within populations [71,72,87,88]. The observed patterns of phenotypic, genetic, and environmental allometry differ markedly; notably, only environmental allometry reflects ontogenetic growth trends. This suggests that phenotypic allometry alone may not reliably predict how size and shape evolve over time [89]. For this reason, we chose to retain outliers in our analysis to preserve the inherent ecological variation present in natural populations. The geographic distribution and directionality of the observed relationships further underscore the importance of considering both environmental and ontogenetic factors when interpreting morphometric datasets, retaining the inherent ecological variation in natural populations [89–91].
The inclusion of historical museum specimens spanning over a century provided a valuable opportunity to assess long-term trends in morphological variation, a strategy widely used in evolutionary and ecological research [92–94]. While such collections offer valuable temporal breadth, they also present inherent limitations, including incomplete metadata, uncertain reproductive status, and unknown age of individuals [95]. Despite these constraints, the consistent pattern of female-biased sexual dimorphism across regions supports the interpretation of a biologically meaningful signal. This finding is in line with other studies demonstrating the utility of museum collections for detecting macroecological trends, even in the presence of data gaps [96,97].
Discrepancies in the literature regarding sexual dimorphism in D. rotundus may stem from regional variation, small or unbalanced sample sizes, and lack of control for confounding variables [98,99].
Beyond evolutionary implications, our findings may hold relevance for understanding behavior and disease ecology. Morphological differences between sexes can influence activity patterns, dispersal, and social structure [11, 100], all of which are relevant to the transmission dynamics of the rabies virus and other zoonoses in vampire bats [101].
Limitations
The reliance on measurements from the right forearm precluded assessment of bilateral asymmetry, an important indicator of developmental stability. For example, fluctuating asymmetry (FA), particularly in forelimbs, has been reported in bats [102]. Studies in Saccopteryx bilineata and Carollia perspicillata have been associated FA with reproductive success and fitness component [103,104]. Similarly, the use of historical specimens introduced potential biases, including uneven geographic representation, unknown age or reproductive condition and potential variation of size during preservation [105]. Lastly, additional traits—such as wingspan, digit length, wing loading, cranial metrics, and glands and non-glandular odour-producing structures —were not available due to the state of conservation of most of the specimens, and it could provide a more comprehensive view of functional dimorphism [106,107].
Outlier specimens that fall within the size range typical of the opposite sex may reflect individual variation due to factors such as age, health condition, or environmental stress [43,44]. In some cases, measurement errors or incomplete sexual dimorphism could also account for the overlap between groups [45,46]. Future studies using digital imaging and geometric morphometrics may offer improved precision and new insights into shape-based dimorphism [108].
Conclusion
Our morphometric analyses reveal modest but consistent female-biased sexual dimorphism in Desmodus rotundus, particularly in forearm and tibia length. This pattern likely reflects a combination of reproductive, ecological, and behavioral pressures acting on females. The inclusion of both historical and contemporary specimens strengthens the reliability of our findings, while also highlighting the value and limitations of museum-based research. Future studies incorporating genetic and ecological variables will be essential to fully understand the evolutionary drivers of sexual dimorphism in this species. As anthropogenic pressures continue to alter bat habitats and behaviors, such knowledge will be crucial for species conservation and for mitigating zoonotic disease risks.
Supporting information
S1 TableMorphometric measurements of Desmodus rotundus specimens.Morphometric data collected from Desmodus rotundus specimens, including forearm length and other standard body measurements.(CSV)
S2 TablePrincipal Component Analysis (PCA) of six morphometric traits in *Desmodus rotundus.*This table summarizes the results of a Principal Component Analysis conducted on six morphological traits. The upper section shows the standard deviation, proportion of variance, and cumulative proportion of variance explained by the first six principal components (PC1–PC6). The lower section lists the loadings of each morphological trait on the corresponding principal components, indicating the contribution of each variable to the respective PC.(CSV)
S1 FigPrincipal Component Analysis (PCA) of morphometric data.Principal Component Analysis (PCA) plot based on the morphometric measurements of Desmodus rotundus (n = 46).(TIF)
S1 TextR scripts used for statistical analyses and plots.This file contains the R code used for data processing, statistical analyses, and generation of figures presented in the manuscript.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Scheffer KC, Barros RF, Iamamoto K, Mori E, Asano KM, Lima JYO, et al. Desmodus rotundus - Biologia y comportamiento [Ph.D. Thesis]. Instituto Pasteur; 2022. doi: 10.37885/220308371 · doi ↗
- 2Sanchez-Gomez WS, Selem-Salas CI, Cordova-Aldana DI, Erales-Villamil JA. Common vampire bat (Desmodus rotundus) abundance and frequency of attacks to cattle in landscapes of Yucatan, Mexico. Trop Anim Health Prod. 2022;54(2):130. doi: 10.1007/s 11250-022-03122-w 35258761 · doi ↗ · pubmed ↗
- 3Brown N, Escobar LE. A review of the diet of the common vampire bat (Desmodus rotundus) in the context of anthropogenic change. Mamm Biol. 2023;:1–21. doi: 10.1007/s 42991-023-00358-3 37363038 PMC 10258787 · doi ↗ · pubmed ↗
- 4Wilson DE, Wilson DE, Mittermeier RA. Handbook of the mammals of the world: hoofed mammals. 2nd ed. Barcelona: Lynx Edicions; 2011.
- 5Iturra-Herrera L, Brito-Carrasco B, Daigre M, Arce P, Arriagada-Gajewski M. Ampliación del rango de distribución sur de Desmodus rotundus (É. Geoffroy Saint Hilaire, 1810), Islote de Pupuya. Región del Libertador Bernardo O´Higgins, Chile Central. Bol Mus Nac Hist Nat. 2020;68(2019):5–12. Available from: https://publicaciones.mnhn.gob.cl/668/w 3-article-95862.html
- 6Van de Vuurst P, Díaz MM, Rodríguez-San Pedro A, Allendes JL, Brown N, Gutiérrez JD, et al. A database of common vampire bat reports. Sci Data. 2022;9(1):57. doi: 10.1038/s 41597-022-01140-9 35173163 PMC 8850563 · doi ↗ · pubmed ↗
- 7Gardner AL. Mammals of South America, 1st ed: marsupials, xenarthrans, shrews, and bats. University of Chicago Press; 2019.
- 8Greenhall AM, Joermann G, Schmidt U, Seidel MR. Desmodus rotundus. Mamm Species. 1983;(202):1–6. doi: 10.2307/3503895 · doi ↗