Investigating the role of cytochrome bd oxidases in the antibacterial activity of madecassic acid and derivatives thereof
Samantha A. Henry, Geraud N. Sansom, Thao Thi Phuong Tran, Ryan A. Boughton, Guy Joiner, Calum M. Webster, H. Ireshika C. de Silva, Michelle D. Garrett, Christopher J. Serpell, Gary K. Robinson, Mark Shepherd

TL;DR
Madecassic acid and its derivatives show antibacterial activity by targeting bacterial-specific enzymes, offering a new approach to antibiotic development.
Contribution
The paper identifies cytochrome bd oxidases as a new antibacterial target for madecassic acid and its derivatives.
Findings
Madecassic acid and its derivatives inhibit cytochrome bd oxidases in bacteria.
Chemical modifications affect the biological activity and phenotypic effects of madecassic acid.
Abstract
Natural products are valuable starting points for drug discovery, although individual modes of action are often difficult to pin down. Ursanes such as madecassic acid have been shown to have antibacterial properties, but a variety of mechanisms have been proposed. In this paper, we report previously uninvestigated activity against cytochrome bd oxidases which are only found in prokaryotes and are therefore promising new targets, using madecassic acid and a set of synthetically modified derivatives. Our work shows that madecassic acid and its derivatives can block activity of these enzymes, while phenotypic effects in membrane and whole organism assays are more complex, consistent with modulation of multiple pathways depending upon molecular structure. This provides a new route to ursane-based antibacterial action while highlighting the importance of chemical modifications in fine-tuning…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
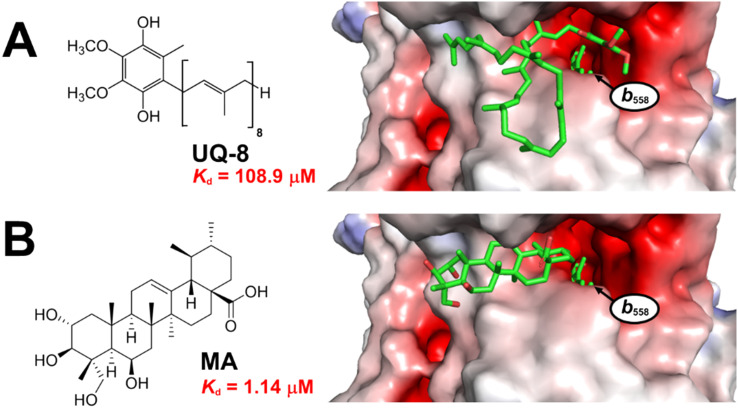 Fig. 1
Fig. 1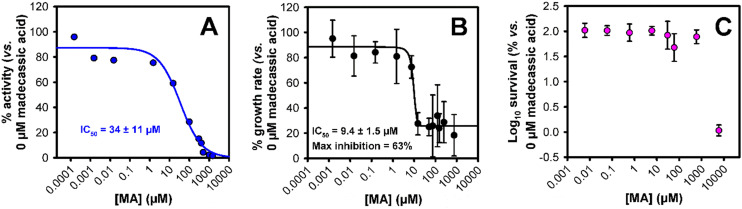 Fig. 2
Fig. 2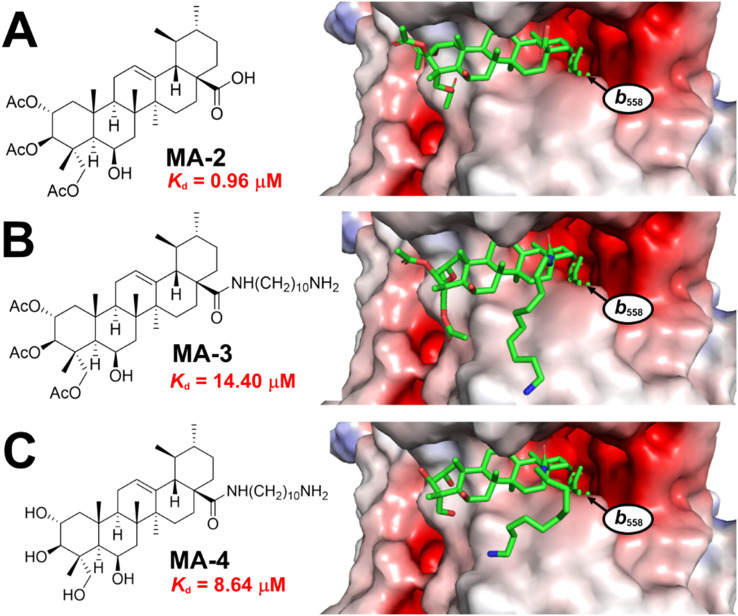 Fig. 3
Fig. 3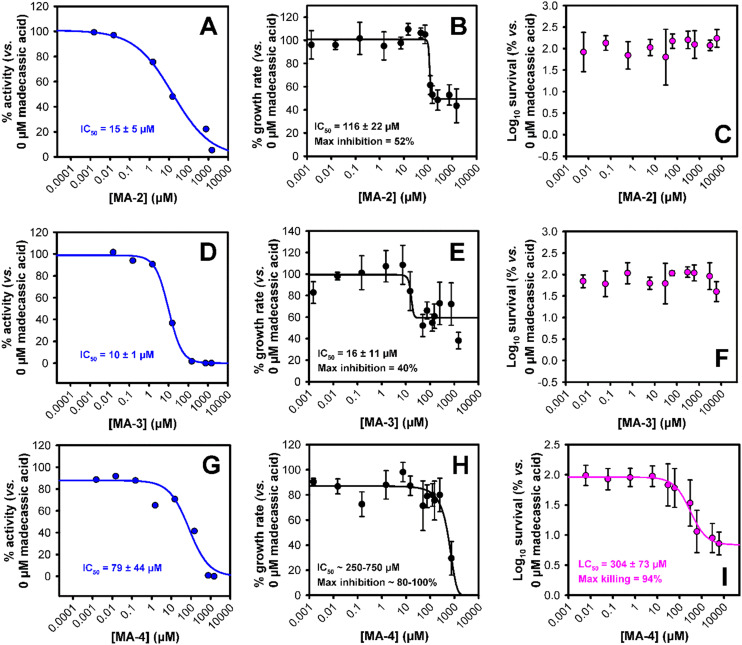 Fig. 4
Fig. 4- —Engineering and Physical Sciences Research Council10.13039/501100000266
- —University of Kent10.13039/501100001316
- —Vietnam Academy of Science and Technology10.13039/100012046
- —Biotechnology and Biological Sciences Research Council10.13039/501100000268
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Applications in Construction Materials · Biological Stains and Phytochemicals · Photodynamic Therapy Research Studies
Introduction
Ursanes are pentacyclic terpenoids which have been shown to have antibacterial properties.^1–4^ Madecassic acid, an ursane produced by the common Asian herb Centella asiatica, has been shown to have good activity against S. aureus and moderate activity against other bacterial species, with evidence consistent with a mechanism including increasing membrane and wall permeability, interaction with DNA, and inhibition of protein synthesis.^5^ This is similar to evidence for ursolic acid, a related compound also found to be active against Gram-positive bacteria.^6^ Unpurified extracts of C. asiatica have shown to be active against Gram-positive and -negative bacteria.^7^ These trends have been demonstrated across a range of modified ursanes,^8^ and the potential of asiatic acid to inhibit bacterial growth had led to its incorporation into gel materials to aid in vivo wound healing.^9^
Cytochrome bd oxidases are attractive drug targets for discovery of new antibacterials, since they are found only in prokaryotic organisms,^10^ and therefore permit specific killing of bacteria. The proteins are found on the inner membrane, where their role is to couple reduction of molecular oxygen with oxidation of quinols, resulting in a proton motive force used in the generation of ATP. Blocking cytochrome bd oxidase activity would therefore impede the capacity of bacteria to access energy required for growth.^11^Escherichia coli, a Gram-negative bacterium responsible for many urinary tract and bloodstream infections, has two bd oxidases: bd-I which is expressed in microaerobic environments, and bd-II which is predominant in anaerobic conditions.^12^
We have discovered effective steroidal ligands for bd-I which inhibit growth and provide lethality against bacteria, with the strongest measured affinities in E. coli.^13^ Since ursane-type structures resemble sterols, it is possible that binding of bacterial cytochrome oxidases is a key component in the antibacterial mechanism of action of the compound class. Since literature reports that a number of mechanisms are at play, it would also be useful to know if the balance of these processes can be modulated through structural refinement of the compounds. In this paper we test madecassic acid (MA) and three synthetic analogues for cytochrome oxidase inhibition through molecular docking, measurement of oxygen consumption in isolated membranes, and testing against live E. coli models, each engineered to have access to different oxidases.
Experimental procedures
Chemical synthesis
Madecassic acid (MA) was obtained by extraction of leaves grown in Quang Tho II Commune, Hue Province, VietNam, following our reported precedure.^14^ Deuterated solvents were purchased from Cambridge Isotope Laboratories Inc. Solvents and reagents were purchased from Thermo Fisher Scientific, Fluorochem, or Alfa Aesar.
A Bruker AVII 400 MHz spectrometer was used to record NMR spectra, and each spectrum was calibrated to the known chemical shift of the residual solvent peak of the deuterated solvent used. Chemical shifts were reported in part per million (ppm) and J coupling values were reported in Hz. Proton NMR spectra were obtained at 400 MHz and ^13^C spectra were obtained at 101 MHz. Spectral data was processed using MestReNova software. Electrospray mass spectrometry data was obtained using a Thermo MSQPlus instrument fitted with a Zorbax SB-C18 5 μm 3.0 × 150 mm column using H_2_O + 0.1% formic acid and MeOH + 0.1% formic acid or H_2_O + 0.1% TFA and MeCN + 0.1% TFA mobile phases. Data was analysed using Chromeleon™ Chromatography Data System (CDS) Software. A Bruker micrOTOF-Q LCMS system was used to obtain high-resolution mass spectrometry data, samples were dissolved in HPLC-grade methanol and injected using direct injection mode with a mobile phase system of 50 : 50 MeOH and H_2_O. Data was processed using Bruker Compass Hystar software.
MA-2
MA (201.6 mg, 0.40 mmol) was dissolved in 6 mL of dry pyridine under nitrogen and acetic anhydride (0.13 mL, 1.40 mmol) was injected into the reaction slowly. After 24 hours, 10 mL pyridine was added, and additional acetic anhydride (4 mL, 42.3 mmol) was added to the reaction. After 48 hours, TLC (after staining with vanillin) showed complete consumption of starting material. The reaction was transferred to a separating funnel, EtOAc (25 mL) was added, and the reaction organic layer was washed with 1 M HCl_(aq)_ (3 × 50 mL). The organic layer was dried over anhydrous MgSO_4_, filtered and the solvent removed under reduced pressure. Crude product was purified using gradient silica gel column chromatography (0–50%, EtOAc : CH_2_Cl_2_) giving MA-2 as a white needle crystalline solid (183 mg 72.6% yield). ^1^H NMR (400 MHz, CDCl_3_) δ 5.27 (d, J = 3.6 Hz, 1H), 5.22 (td, J = 11.3, 4.8 Hz, 2H), 5.00 (d, J = 10.3 Hz, 1H), 4.33 (d, J = 3.8 Hz, 1H), 3.93 (d, J = 11.9 Hz, 1H), 3.70 (d, J = 12.0 Hz, 1H), 2.19 (d, J = 11.2 Hz, 1H), 2.15–0.80 (m, 46H). ^13^C NMR (101 MHz, CDCl_3_) δ 183.9, 171.0, 170.6, 170.6, 137.3, 125.7, 75.0, 70.0, 68.0, 65.5, 52.5, 48.3, 48.0, 47.9, 45.9, 42.5, 40.8, 39.2, 38.9, 38.7, 37.4, 36.7, 30.7, 28.0, 24.2, 23.6, 23.4, 21.3, 21.2, 21.2, 21.0, 20.9, 18.8, 18.6, 17.1, 15.5, 14.3. HREI-MS: m/z calculated for C_36_H_54_O_9_Na [M + Na^+^] 653.3660; observed 653.3672.
MA-3
The tert-butyloxycarbonyl-protected version of MA-3 (MA-3Boc) was first synthesised. HATU (131.1 mg, 0.334 mmol) and MA-2 (175.5 mg, 0.278 mmol) were dissolved in 5 mL dry CH_2_Cl_2_ and DIPEA (0.145 mL, 0.835 mmol) was slowly added to the reaction and it was stirred for 1 hour. Tert-butyl 10-aminodecylcarbamate (152.8 mg, 0.556 mmol) was dissolved in dry CH_2_Cl_2_ (5 mL) and slowly added to the reaction and it was left to stir overnight. After 22 hours, the reaction was transferred to a separating funnel and diluted with CH_2_Cl_2_ (30 mL). It was then extracted with 1 M HCl_(aq)_ (3 × 50 mL) and brine (1 × 50 mL). Dried over anhydrous MgSO_4_, filtered and reduced under vacuum. The crude product was purified by isocratic column chromatography (25% : 75%, EtOAc : CHCl_3_) to yield MA-3Boc as a white solid (236 mg, 95.8%). ^1^H NMR (400 MHz, CDCl_3_) δ 5.85 (t, J = 5.4 Hz, 1H), 5.34 (t, J = 3.5 Hz, 1H), 5.22 (td, J = 11.0, 4.7 Hz, 1H), 5.01 (d, J = 10.3 Hz, 1H), 4.57 (s, 1H), 4.34 (t, J = 3.8 Hz, 1H), 3.94 (d, J = 11.9 Hz, 1H), 3.71 (d, J = 12.0 Hz, 1H), 3.23 (dq, J = 13.3, 6.8 Hz, 1H), 3.14–2.93 (m, 3H), 2.17–0.77 (m, 72H). ^13^C NMR (101 MHz, CDCl_3_) δ 177.9, 170.9, 170.5, 170.5, 139.4, 125.5, 79.1, 75.0, 70.1, 67.4, 65.4, 54.2, 48.1, 47.9, 47.8, 46.0, 43.1, 42.5, 40.6, 39.9, 39.6, 39.2, 38.8, 38.8, 37.4, 37.3, 31.0, 30.2, 29.5, 29.5, 29.3, 29.3, 29.3, 29.2, 28.5, 27.9, 27.2, 26.9, 24.9, 23.5, 23.3, 21.3, 21.2, 21.0, 20.9, 18.7, 18.3, 17.3, 15.4. HREI-MS: m/z calculated for C_51_H_84_N_2_O_10_Na [M + Na^+^] 907.6018; observed 907.6052.
MA-3Boc (131.2 mg, 0.148 mmol) was then dissolved in CH_2_Cl_2_ (3.7 mL), 4 M HCl in dioxane (3.7 mL, 14.8 mmol) was added to the solution at 0 °C. The reaction was stirred and left to warm to room temperature overnight. After 21 hours, solvent was removed under reduced pressure. The residue was washed into a separating funnel with a mixture of CH_2_Cl_2_ (50 mL) and sat. NaHCO_3(aq)_ (50 mL) and aqueous layer removed. It was then extracted with a further sat. NaHCO_3(aq)_ (2 × 50 mL) and brine (1 × 50 mL). Dried over anhydrous MgSO_4_, filtered and reduced under vacuum. Extracted product was clean by NMR and required no further purification yielding MA-3 (105.8 mg, 90.9%). ^1^H NMR (400 MHz, CDCl_3_) δ 5.92 (t, J = 5.4 Hz, 1H), 5.34 (d, J = 3.7 Hz, 1H), 5.21 (td, J = 10.9, 4.5 Hz, 1H), 5.00 (d, J = 10.3 Hz, 1H), 4.83 (s, 2H), 4.31 (d, J = 5.1 Hz, 1H), 3.94 (d, J = 11.7 Hz, 1H), 3.70 (dd, J = 12.5, 2.6 Hz, 1H), 3.28–2.94 (m, 3H), 2.85 (s, 2H), 2.25–0.73 (m, 62H). ^13^C NMR (101 MHz, CDCl_3_) δ 178.1, 171.1, 170.7, 170.6, 139.4, 125.5, 75.0, 70.2, 67.2, 65.4, 54.1, 48.1, 47.9, 47.8, 46.0, 43.8, 43.1, 42.5, 40.4, 39.9, 39.7, 39.2, 38.8, 37.4, 37.3, 31.0, 29.5, 29.4, 29.3, 29.3, 29.2, 29.1, 27.8, 27.1, 26.7, 24.9, 23.5, 23.3, 21.3, 21.3, 21.0, 20.9, 18.7, 18.3, 17.3, 15.4.
MA-4
MA-3 (52.0 mg, 0.0662 mmol) was dissolved in MeOH (5 mL). 4 M NaOH_(aq)_ solution (0.080 mL, 0.331 mmol) was injected into the reaction, and it was left to stir overnight. After 20 hours, reaction was transferred to a separating funnel, EtOAc (50 mL) was added. The organic layer was extracted with 1 M NaOH_(aq)_ (3 × 50 mL). Dried over anhydrous MgSO_4_, filtered and reduced under vacuum. Extracted product was pure and required no further purification, yielding MA-4 (43.6 mg, 99.9%). ^1^H NMR (400 MHz, methanol-d4) δ 5.39 (t, J = 3.6 Hz, 1H), 4.37 (t, J = 3.4 Hz, 1H), 3.74 (ddd, J = 11.3, 9.3, 4.3 Hz, 1H), 3.58 (d, J = 11.1 Hz, 1H), 3.43 (d, J = 11.1 Hz, 1H), 3.30 (d, J = 8.0 Hz, 1H), 3.21–3.01 (m, 2H), 2.75–2.66 (m, 2H), 2.20–0.79 (m, 54H). ^13^C NMR (101 MHz, methanol-d4) δ 180.1, 139.7, 127.1, 78.0, 69.6, 68.3, 65.7, 54.3, 50.2, 49.1, 48.9, 44.8, 43.9, 42.0, 41.2, 40.9, 40.8, 40.3, 40.1, 38.8, 38.5, 32.2, 32.0, 30.7, 30.7, 30.7, 30.6, 30.5, 30.3, 28.9, 28.3, 27.9, 25.2, 24.5, 24.1, 21.6, 19.3, 19.3, 17.7, 15.3. HREI-MS: m/z calculated for C_40_H_71_N_2_O_5_ [M + H^+^] 659.5357; observed 659.5392.
Computation and molecular docking
Ligand docking to an AlphaFold2 model of E. coli cytochrome bd-I was performed essentially as described in detail previously.^13^ Briefly, preparation of in silico ligand and protein files was performed using AutoDockTools and PyMOL.^15^ The equation Kd = exp(ΔG/(R × T)) with ΔG = binding energy (kcal mol^−1^), R = gas constant = (1.986 cal mol^−1^ K^−1^) and T = temperature (298 K) was used to estimate the dissociation constants.
Clog P values were calculated using the Chemical Properties tool in Signals ChemDraw v23.1.2.7.
Bacterial strains
EC958 is a multidrug-resistant E. coli O25: H4-ST131 isolate.^16,17^ Generation of the EC958 bd-I only strain (genotype ΔcyoA appCB::Cm) is described elsewhere.^13^
Growth assays
Starter cultures were grown in 10 mL LB (E. coli) in sterile 50 mL conical flasks at 180 rpm and 37 °C until stationary phase was reached and were used to inoculate 50 mL of fresh growth medium in 250 mL conical flasks. M9 minimal medium was used for E. coli (16 g L^−1^ Na_2_HPO_4_·2H_2_O, 3 g L^−1^ KH_2_PO_4_, 0.5 g L^−1^ NaCl, 1 g L^−1^ NH_4_Cl, 0.24 g L^−1^ MgSO_4_, 0.01 g L^−1^ CaCl_2_, 0.1% casamino acids and 2% glycerol). Drug stocks were prepared in DMSO so that their final concentrations were 40× higher than the working concentrations. Greiner F-bottom sterile 96-well plates were prepared by adding 100 μL of a 2× concentrated growth medium, 61.7 μL sterile milliQ H_2_O, 5 μL of the drug and 33.3 μL of cell culture (final OD_600_ of ∼0.1). Cells were grown in a FLUOstar Omega plate reader at double orbital pattern setting.
Oxygen consumption assays
Membrane preparation and oxygen consumption were performed as described previously.^13^ Briefly, cells were grown to exponential phase then harvested via centrifugation for 20 min at 4000 rpm and 4 °C. The cell pellet was resuspended in ice-cold sonication buffer (20 mM Tris/HCl at pH 7.4, 2 mM MgCl_2_, and 1 mM EGTA). The resuspended cells were sonicated (6 × 30s on ice at 15 μM) before centrifugation at 44 000 rpm for 1 h and 4 °C to isolate membranes. The membrane pellet was resuspended in 20 mM Tris/HCl (pH 7.4) at a final concentration of 100 mg mL^−1^ and stored at −20 °C. For oxygen measurements a Rank Brothers oxygen electrode with a 4 mL closed chamber was used at 37 °C, which contained 50 mM HEPES pH 7.4, 0.5 mg mL^−1^ membranes (based on wet membranes) and DMSO-solubilised drug (added from 40× final concentration). A final concentration of 8 mM succinate (pH 7.4) was added (from 160 mM stock) to initiate the reaction with a single run lasting 15–20 min.
Viability assays
Viability assays were performed essentially as described previously.^13^ Briefly, E. coli was grown overnight in 10 mL LB medium and was used to inoculate M9 medium. 20 μL of each drug was added to the wells of row A of a 96 well plate followed by 180 μL of cells (OD_600_ of 0.1). Cells were exposed to drug for 3 hours at 37 °C. Following drug exposure, serial dilutions were performed in 1 × phosphate buffered saline (pH 7.4) before being spotted onto LB agar plates overnight to determine changes in cell survival. Six repeats were performed for each concentration of drug which included two biological repeats and three technical repeats of each.
Results
Both MA and ubiquinol-8 (UQ-8) the natural substrate of cytochrome bd-I were docked to the quinol site of the E. coli AlphaFold2 cytochrome bd-I structural model.^13^ The binding affinity values predicted by AutoDock Vina (Fig. 1) indicate that MA is substantially more tightly bound by cytochrome bd-I (1.14 μM) than UQ-8 (108.9 μM). The carboxylic acid of MA was found to reside in the hydrophobic cleft (red, Fig. 1) close to where the aromatic unit of UQ-8 sits, while the pentacyclic hydrocarbon occupies a similar space to that of the oligoisoprene tail of UQ-8. The polar units on MA point out of the protein cavity towards the aqueous environment.
Docking of ubiquinol-8 (A) and MA (B) to the quinol-binding site of an AlphaFold 2 model for E. coli cytochrome bd-I. Haem b558 is a cytochrome bd-I cofactor that accepts electrons from the ubiquinol substrate, and is labelled on the figure. Predicted affinities from Autodock Vina are shown in red.
These predictions suggest that MA would be able to competitively inhibit cytochrome bd-I. To test this prediction, we performed oxygen consumption assays to measure oxidoreductase activity using isolated E. coli EC958 cytochrome bd-I only membranes (Fig. 2A),^13^ and MA obtained by extraction of C. asiatica,^14^ resulting in an IC_50_ of 34 ± 11 μM. This activity is consistent with the docking results in that MA would be expected to outcompete UQ-8. On this basis, we advanced to assessing the ability of MA to inhibit the growth of cytochrome bd-I only E. coli cells (Fig. 2B), giving an IC_50_ of 9.4 ± 1.5 μM, with a maximum inhibition of 63%. We further conducted a viability assay (Fig. 2C) which showed that MA did not kill bd-I only E. coli cells. The observed lack of lethality may relate to permeability of the cell membrane and/or wall to MA, but is consistent with other tests on Gram-negative bacteria.^5^
Impact of madecassic acid upon cytochrome bd-I activity, E. coli growth, and E. coli survival. (A) Dose inhibition curve of madecassic acid against oxygen consumption activity of E. coli EC958 cytochrome bd-I-only membranes. (B) Growth assay of MA against E. coli cytochrome bd-I only cells. (C) Survival assay of MA against E. coli EC958 cytochrome bd-I only cells. Dose response data were fitted to three- or four-parameter logistic equations using nonlinear regression (Sigmaplot) to generate IC50 values with standard error. Error bars represent standard deviations for at least four technical repeats, including two biological repeats.
To investigate the contribution of cytochrome bd-I to MA-mediated respiratory inhibition, growth experiments were also performed on the wild type EC958 strain, which encodes an additional respiratory oxidase cytochrome bo′ that is expressed under the aerobic conditions tested (Fig. S1A). These data revealed an IC_50_ of ∼10 μM and maximal inhibition of 48%, which is consistent with the wild type responding to respiratory inhibition in a similar way to the bd-I only strain. In addition, the wild type strain was not susceptible to MA-mediated killing at concentrations below 1 mM (Fig. S1B), confirming that MA is bacteriostatic but not bactericidal towards both strains.
We then decided to explore variations upon the MA skeleton which might alter access to proteins on the bacterial inner membrane. For this reason, we examined modified versions. These were based on transformations prominent in the literature for such compounds,^18,19^ enabling easy development in future, and operated on opposite ends of the molecule to provide initial independent evaluation of modification sites. Acetylating the alcohols at positions 2, 3, and 23 gave MA-2, lowering polarity from Clog P of MA at 3.97 to 6.64. Additionally, conversion of the carboxylic acid into an amide using 1,10-diaminodecane to add a hydrophobic chain and introduce a cationic group (likely to interact with bacterial lipids) gave MA-3 (Clog P = 8.63). We also produced the 1,10-diaminodecane amide without acetylation (MA-4, Clog P = 5.96) to create a double-mutant cycle. These compounds have been previously studied for anticancer activity, and were synthesised in the same manner, starting from MA.^18^ As before, these were docked into the quinol binding site of cytochrome bd-I (Fig. 3). Compared with the predicted Kd of 1.14 μM for MA, acetylation resulted in a slightly stronger affinity (Kd = 0.96 μM) for MA-2. However, when the long chain amine was also added to give MA-3, the affinity was substantially weakened (Kd = 14.4 μM). The chain appears to add too much bulk, and impedes the binding geometry. Removal of some of the molecule's steric bulk in the unacetylated MA-4 resulted in a moderate improvement in Kd (8.64 μM). Importantly, all of these are modelled to bind more strongly than UQ-8, and therefore we could expect inhibitory activity.
Madecassic acid derivatives docked to the quinol binding site of an AlphaFold 2 model for E. coli cytochrome bd-I. Haem b558 is a cytochrome bd-I cofactor that accepts electrons from the natural substrate ubiquinol (labelled as b558). (A) Docking of MA-2. (B) Docking of MA-3. (C) Docking of MA-4.
Oxygen consumption assays were then performed with bd-I-only membranes (Fig. 4a). The tighter predicted binding of MA-2 compared with MA correlated with elevated inhibition (IC_50_ = 15 μM, Fig. 4A), as did the slightly weaker binding of MA-4 correspond to a lowered inhibition (IC_50_ = 79 μM, Fig. 4G). However, MA-3 was more active than would be expected from the predicted binding (IC_50_ = 10 μM, Fig. 4D). This is potentially due to the more lipophilic nature of that compound which may result in it being drawn into the membranes. The same compound performed well in the growth inhibition of bd-I only E. coli cells (IC_50_ = 16 μM, Fig. 4E), although was not as potent as MA (IC_50_ = 9.4 μM, Fig. 2B). MA-2 was much less active (IC_50_ = 116 μM, Fig. 4B) despite having good activity against membranes, while MA-4′s weak performance against membranes was echoed in its limited (IC_50_ > 250 μM, Fig. 4H) growth inhibition. In light of these results, it was therefore a surprise when MA-4 revealed itself as the only compound tested which had any capacity (albeit weak) to kill E. coli bd-I only cells (Fig. 4I), with an LC_50_ of 304 μM.
Impact of madecassic acid derivatives upon E. coli growth, cytochrome bd-I activity, and E. coli survival. (A, D and G) Oxygen consumption activities of E. coli EC958 cytochrome bd-I-only membranes exposed to madecassic acid derivatives. (B, E and H) Growth assays E. coli cytochrome bd-I only cells exposed to madecassic acid derivatives. (C, F and I) Survival assays for E. coli EC958 cytochrome bd-I only cells exposed to madecassic acid derivatives. Where possible, data were fitted to three- or four-parameter logistic equations using nonlinear regression (Sigmaplot) to generate IC50 values (or LC50 for killing data) with standard error. Error bars represent standard deviations for at least four technical repeats, including two biological repeats.
Discussion
These results show that inhibitory activity of bacterial cytochromes is involved in the antibacterial activity of MA, with binding to the ubiquinol site of cytochrome bd-I being of particular interest, to which MA and all our derivatives are modelled to bind more strongly than the natural substrate. In line with these results, MA itself shows inhibition of enzymatic activity and bacterial growth, although without any observed killing of E. coli – this is consistent with other studies which found that Gram-positive bacteria were more vulnerable.^5^
Having applied chemical modifications to MA, we were able to improve enzymatic inhibition activity in membrane assays for two out of three variants (MA-2 and MA-3). Unmodified MA proved best for growth inhibition, although MA-3, the most highly modified version was close behind, despite being expected to bind the protein the least strongly. However, it was only MA-4, ranked third in predicting binding and last in enzymatic effects, which was found to kill E. coli. It must be said therefore that the links between predicted binding, and the different levels of biological activity (protein activity in isolated membranes, growth inhibition, cell killing) are mixed. This supports previous work by other groups which have identified a variety of mechanisms through which MA has antibacterial effects including membrane disruption, inhibition of protein synthesis, and topoisomerase inhibition.^6^ In particular, we note that our modifications make the compound significantly more hydrophobic as reflected in a shift of calculated partition coefficient of four log_10_ units between MA and MA-3, as well as swapping an anionic (at biological pH) carboxylate for a cationic ammonium; both of these changes are likely to increase interactions with lipids. The proteins of interest reside in the inner membrane of E. coli, and their natural substrate is primarily found in the lipid bilayer. This increased lipidic interaction could therefore both increase the local concentration of MA derivatives, or conversely result in competitive sequestration within the lipid bilayer, away from the protein. Convolution of these effects with the other mechanisms at play have led to the overall result that MA-4 has the greatest bactericidal activity. The myriad ways in which natural products such as MA can interact with biological macromolecules leads to complications which are hard to avoid – in our case, the possibility that MA might interact with succinate dehydrogenase^5^ was a potential limitation for the isolated membrane components of this study, since the oxygen consumption assay relies upon this complex to maintain the supply of ubiquinol for cytochrome bd-mediated oxygen consumption. It was therefore necessary to perform additional control experiments to exclude MA-mediated SDH inhibition and further demonstrate the inhibition of cytochrome bd-I activity where electron delivery is independent of SDH. To introduce an additional level of rigour to these investigations, an alternative bd-I only strain was engineered where an oxidase null mutant strain was complemented with a plasmid encoding the cytochrome bd operon (SI). Membranes from this E. coli EcoM4 pSU2718G-cydABX-his_6_ strain were prepared and SDH activity measurements confirmed that 1 mM MA did not inhibit SDH in isolated membranes (Fig. S2). To verify that MA directly targets cytochrome bd-I directly, oxygen consumption experiments were undertaken using the same membranes and duroquinol as the electron donor (delivers electrons directly to bd-I). These data (Fig. S3) confirmed that 1 mM MA completely abolishes bd-I activity, further supporting the direct binding of MA to bd-I in EC958 membranes.
Plants have evolved a certain set of natural products which have a particular role for that plant. Because they interact with the biological machinery in one organism, there is a good chance that they will do something in other organisms, and this has historically led to many successful medicines.^20^ However, there is no reason to expect natural products to behave exactly as we would wish, and off-target effects are common, since the compounds were selected for a different role. Chemical modification can make a big difference,^21^ and we have shown here that modifications can alter the behaviour of MA in bacterial models, and this provides a route to focus the activity of a natural product into a particular mechanism. Through strategic chemical modification, it should therefore be possible to both better understand the different mechanisms of triterpenoid antibacterial activity, and create molecules which maximise their potency against specific targets. Cytochrome bd oxidases are particularly attractive in this regard since they are only found in prokaryotic organisms. Given the versatile modification chemistry open to madecassic acid,^18,19,22,23^ further work to maximise its activity against these proteins would be well justified.
Conclusion
Madecassic acid and three derivatives thereof have been modelled to, and shown to have, a high affinity for cytochrome bd-I oxidase, and experimental observations of enzymatic activity, bacterial growth, and bactericidal assays are consistent with this modelling. Alterations of the chemical structure have been shown to influence the biological activity, although the relationships between the different assay outcomes are non-linear. These findings show that inhibition of cytochrome bd oxidases is a further component of the antibacterial activity of madecassic acid, in addition to mechanisms already investigated by others. Further strategic chemical modification could result in new antimicrobial compounds based upon madecassic acid or other similar plant-derived compounds, which are specifically targeted towards the unique aspects of bacterial biochemistry.
Author contributions
All authors contributed to writing the manuscript. TTPT extracted and purified the MA from C. asiatica. GNS and HICDS performed organic transformations. SAH was involved in planning and performing the microbiological experiments and data analysis. RAB developed the duroquinol assay, CMW engineered the complementation vector, GJ performed the SDH assays and duroquinol experiments, GKR contributed to the planning of the microbiological experiments and data analysis. CJS and MDG obtained funding and supervised the chemistry. MS also obtained funding and was responsible for all microbiological experimental design and data analysis.
Conflicts of interest
There are no conflicts of interest.
Supplementary Material
MD-OLF-D5MD01116G-s001
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wojnicz D. Tichaczek-Goska D. Korzekwa K. Kicia M. Hendrich A. Anti-Enterococcal Activities of pentacyclic Triterpenes Adv. Clin. Exp. Med.201726348349010.17219/acem/6224528791824 · doi ↗ · pubmed ↗
- 2Sycz Z. Tichaczek-Goska D. Jezierska-Domaradzka A. Wojnicz D. Are Uropathogenic Bacteria Living in Multispecies Biofilm Susceptible to Active Plant Ingredient—Asiatic Acid?Biomolecules 20211112175410.3390/biom 1112175434944398 PMC 8698853 · doi ↗ · pubmed ↗
- 3Liu W.-H. Liu T.-C. Mong M.-C. Antibacterial Effects and Action Modes of Asiatic Acid Biomedicine 2015531610.7603/s 40681-015-0016-726280399 PMC 10723755 · doi ↗ · pubmed ↗
- 4Wojnicz D. Tichaczek-Goska D. Kicia M. pentacyclic Triterpenes Combined with Ciprofloxacin Help to Eradicate the Biofilm Formed in Vitro by Escherichia coli Indian J. Med. Res.2015141334335310.4103/0971-5916.15663125963496 PMC 4442333 · doi ↗ · pubmed ↗
- 5Wei C. Cui P. Liu X. Antibacterial Activity and Mechanism of Madecassic Acid against Staphylococcus aureus Molecules 2023284189510.3390/molecules 2804189536838882 PMC 9967526 · doi ↗ · pubmed ↗
- 6Liu G. Qin P. Cheng X. Wu L. Zhao W. Gao W. Evaluation of the Mechanistic Basis for the Antibacterial Activity of Ursolic Acid against Staphylococcus aureus Front. Microbiol.20241510.3389/fmicb.2024.1389242 PMC 1114014738827151 · doi ↗ · pubmed ↗
- 7Chonsut P. Romyasamit C. Konyanee A. Niyomtham N. Goodla L. Mordmuang A. Potential Activities of Centella Asiatica Leaf Extract against Pathogenic Bacteria-Associated Biofilms and Its Anti-Inflammatory Effects Adv. Pharmacol. Pharm. Sci.202420241595907710.1155/2024/595907739296309 PMC 11410404 · doi ↗ · pubmed ↗
- 8Xie Z. Dai X. Wang L. Novel 23-Hydroxybetulinic Acid Derivatives: Semi-Synthesis and Antibacterial Activity Russ. J. Gen. Chem.202494102770278810.1134/S 1070363224100219 · doi ↗