The Tonoplast Topology Index—a new metric for describing vacuole organization
Helena Kočová, George Alexandru Caldarescu, Radek Bezvoda, Fatima Cvrčková

TL;DR
The paper introduces a new metric called the Tonoplast Topology Index to better analyze plant vacuole organization, offering advantages over existing methods.
Contribution
The novel Tonoplast Topology Index (TTI) provides a more efficient and statistically robust way to quantify vacuole organization.
Findings
TTI outperforms the Vacuolar Morphology Index (VMI) in capturing vacuole compartment organization and statistical normality.
TTI successfully detected subtle changes in vacuole organization in Arabidopsis roots based on growth conditions.
The TTI protocol is semi-automated, user-friendly, and suitable for high-throughput analysis.
Abstract
The plant vacuole arises by orchestrated interplay of membrane trafficking, cytoskeletal rearrangements and a variety of signaling pathways. In the root, the characteristic large central vacuole develops by endomembrane reorganization occurring mainly in the transition zone. The vacuole’s bounding membrane—the tonoplast—can be visualized in vivo using fluorescent protein markers, allowing for quantitative analysis of confocal microscopy images. Tonoplast organization can thus serve as a sensitive indicator of changes to any of the processes involved in vacuole biogenesis. The Vacuolar Morphology Index (VMI) is widely accepted as a quantitative measure of vacuole structure. However, this metric has two drawbacks—it only reflects the size of the largest vacuolar compartment (missing therefore possible differences in the organization of smaller compartments), and its determination is labor…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
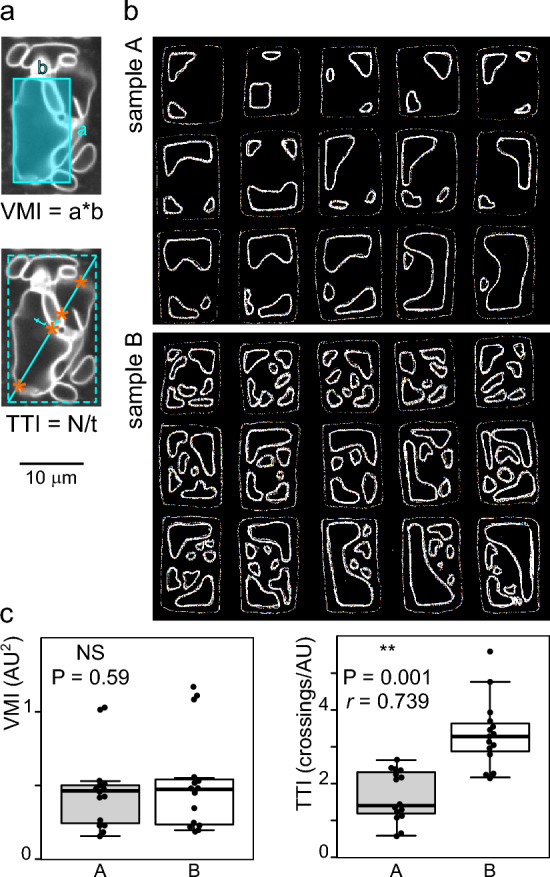 Figure 1
Figure 1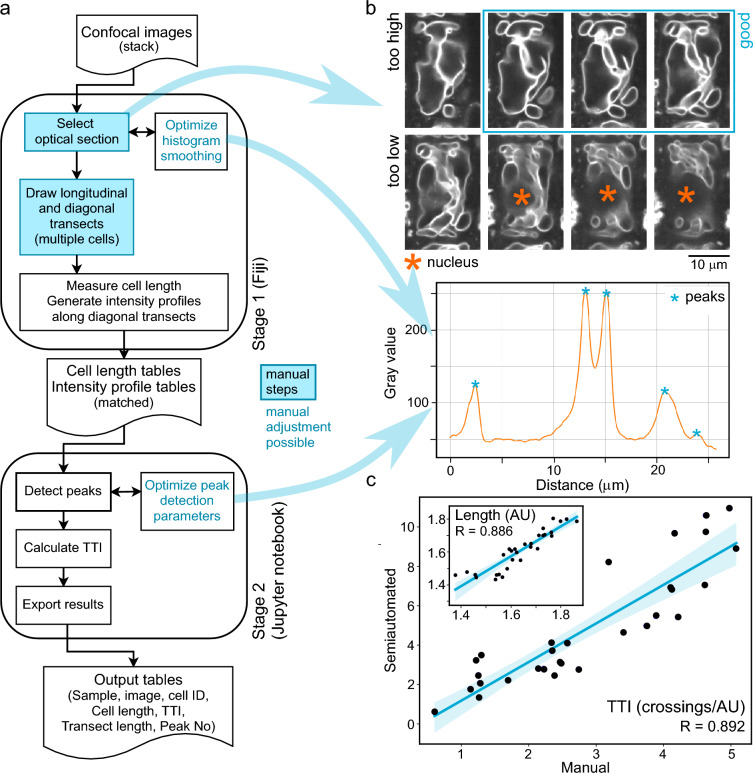 Figure 2
Figure 2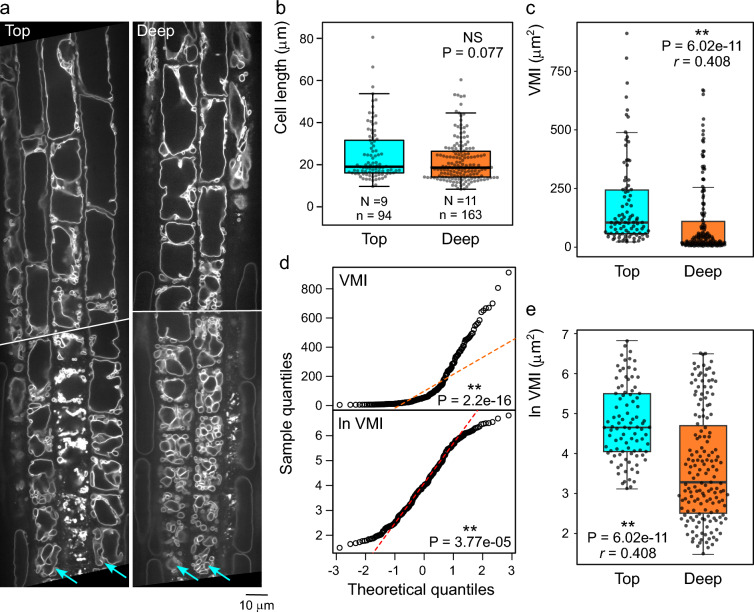 Figure 3
Figure 3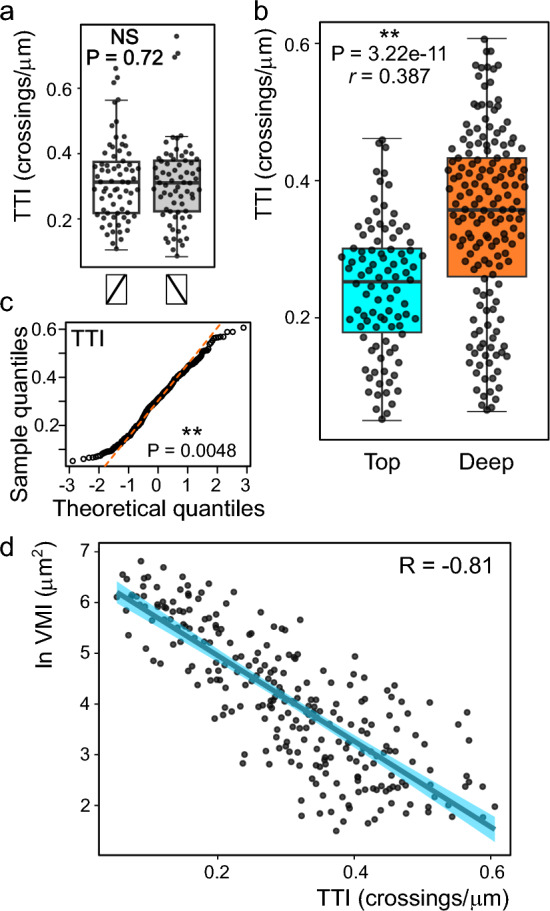 Figure 4
Figure 4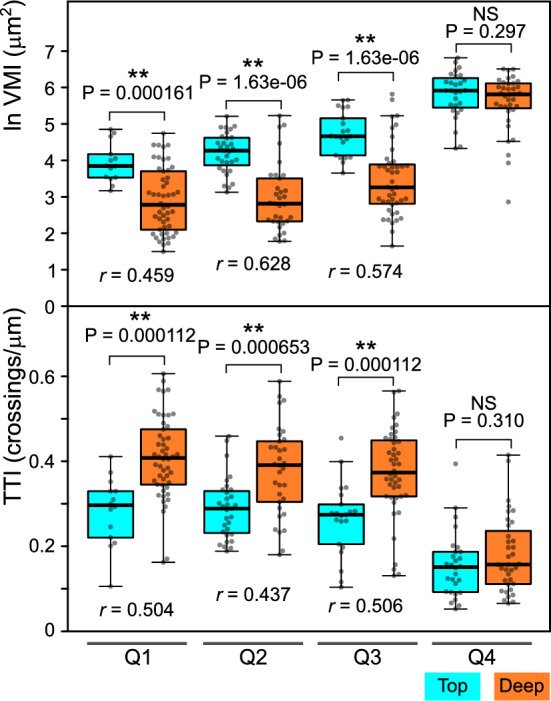 Figure 5
Figure 5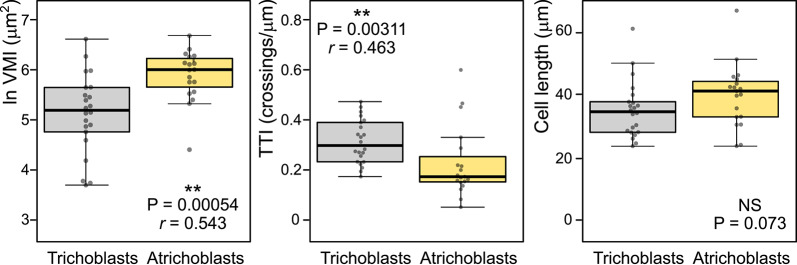 Figure 6
Figure 6- —https://doi.org/10.13039/501100001824Grantová Agentura České Republiky
- —https://doi.org/10.13039/501100001823Ministerstvo Školství, Mládeže a Tělovýchovy
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCellular transport and secretion · ATP Synthase and ATPases Research · Bacterial Genetics and Biotechnology
Background
Plant vacuoles are the largest intracellular membrane compartments, participating in various processes including storage or degradation of cellular components, ion homeostasis or cell expansion (see, e.g., [1]). The vacuoles, distinguishable by the presence of specific tonoplast markers on their bounding membrane, attain various shapes, ranging from a highly fragmented collection of smaller compartments or a tubular network in meristematic cells [2, 3] to one big central vacuole with a few invaginations or occasional cytoplasmic strands, which can take up to 90% of the cell volume in expanding or differentiating cells (reviewed in [4]).
In typical angiosperm roots (such as those of Arabidopsis thaliana), establishment of the large central vacuole takes place in the transition zone and is a prerequisite for rapid turgor-driven cell expansion. The central vacuole arises by fusion and/or expansion of precursor compartments in a consolidation process that involves trafficking of membranes originating from the endoplasmic reticulum, depends on the integrity of the actin cytoskeleton and is intimately linked to auxin signalling (e.g. [2, 3, 5–7]). Any changes to this process may cause variability in tonoplast organization in otherwise similarly-sized cells.
Vacuole shape is affected by various conditions, such as cell type [8], pH [9], sucrose availability and concentration in the media [10], as well as by stiffness of root surroundings, with differences reported between roots penetrating inside media compared to those growing on the agar surface [9]. Various mutations or pharmacological treatments can modulate vacuole organization as well (summarized in [11]). Mutations disabling the critically important molecular machinery responsible for central vacuole development, for example subunits of the HOPS complex [12], are lethal at the embryo stage.
Since the tonoplast can be easily visualized by confocal microscopy using fluorescent dyes or tonoplast-resident proteins fused to various fluorescent proteins (e.g. [3, 13, 14]), quantitative description of tonoplast morphology in differentiating root tissues, whose cells exhibit a relatively regular shape, may serve as a good indicator of changes in the process of vacuole biogenesis. A “vacuolar morphology index” (VMI), defined as the product of the length and width of the largest vacuole compartment section in an optical section of a cell (see Fig. 1a), has been introduced as a quantitative metric for describing the state of vacuole development [6]. This metric has been widely adopted in subsequent studies to document effects of auxin, other signaling molecules, mutations or environmental factors on vacuole organization (e.g., [7, 9, 15–18]).Fig. 1. Comparison of the vacuolar morphology index (VMI) and tonoplast topology index (TTI) metrics. a A representative optical section of a transition/elongation zone rhizodermis cell expressing GFP-tagged vacuolar H + pyrophosphatase with schematic drawing of the principle of VMI and TTI determination. N is the number of membrane crossings along a transect (marked by asterisks). Dashed rectangle approximates the cell outline. b Hand-drawn simulated images of two populations of cell optical sections with similar VMI but different vacuole organization. c Result of manual VMI and TTI determination from simulated data shown in (b), where only TTI can distinguish between the two populations. (t-test P values shown, with strength of effect r value provided for the significant difference; AU—arbitrary units).
However, while the VMI captures well the overall progress of central vacuole enlargement, it does not provide any information about the organization of vacuolar compartments outside the largest (nascent) central vacuole. We are proposing an alternative metric of vacuole organization, which overcomes this drawback of the VMI, specifically focusing on the overall extent of vacuole fragmentation and complexity of vacuole shape. This method has been tested on simulated data and on a benchmark set of images taken from Arabidopsis roots expressing GFP-tagged vacuolar H + pyrophosphatase [14] that were grown on the surface or inside agar medium, reproducing a previously reported observation [9]. Together with comparison of TTI and VMI results on these datasets, we are presenting a user-friendly, platform-independent, freeware-based protocol for semi-automated, relatively high throughput TTI determination.
Methods
Plants and culture conditions
An Arabidopsis thaliana line expressing plant vacuolar H + pyrophosphatase 1 fused with monomeric GFP (VHP1:mGFP), obtained by crossing an original Col-0 background line [14] to the Col-8 background, was used in all experiments.
Seedlings were grown in vitro on vertically positioned Petri dishes containing half-strength Murashige and Skoog (MS) medium with 1.6% (w/v) agar and 1% (w/v) sucrose, pH adjusted to 5.7, at 22 °C with 16 h light/8 h dark cycle as described previously [19] for 5 days prior to imaging. Roots grown on the media surface were directly mounted into a microscopy chamber, while roots growing inside agar media were carefully pulled out from agar before mounting.
Imaging
Confocal Z-stack images were acquired using a vertical spinning disc confocal microscopy system consisting of a Zeiss Axio Observer 7 microscope integrated with a Yokogawa CSU-W1-T2 spinning disk unit featuring 50 µm pinholes and a VSHOM1000 excitation homogenizer (Visitron Systems), along with VisiView software (Visitron System, v4.40.14) and alpha Plan-Apochromat 100 × /1.46 Oil immersion objective. A 488 nm laser was employed for GFP (excitation 488 nm, emission 500–550 nm). Images from at least 8 roots were collected for each studied condition. Typically, two minimally overlapping Z-stacks with 15 to 20 optical sections 1 µm apart through the thickness of the rhizodermis were taken per root, covering the range from the meristem to the mid-elongation zone.
Image processing and image analysis
Routine image processing, VMI determination and the first stage of the TTI determination procedure (including its development) has been performed on a personal computer running the Fiji distribution [20] of ImageJ 1.54p under a 64-bit version of Windows 10 or Windows 11 (Enterprise/Pro or Home edition). Prior to analysis, raw confocal image stacks were processed by algorithmic contrast enhancement to ensure good visibility of the tonoplast. For long-time archiving, the image stacks were subsequently converted to 8 bit, rotated to position the root vertically, cropped, and converted to the *.ome.tiff format (see Additional File 1). All image quantification procedures have been successfully performed both on raw or contrast-enhanced images and on files that underwent processing for archiving with practically identical results obtained for a random image sample. Quantitative analyses were performed on atrichoblasts of the rhizodermis, whose vacuole organization differs from that of the trichoblasts [21], unless stated otherwise.
Typically, 10–20 cells per root were subjected to quantitative image analysis. Consistent results were obtained upon independent experiment replication including a different plant genotype, as well as upon repeated measurements of a subset of images. Data from a representative replica are shown.
VMI was determined by multiplying manually measured values of the length and width of the largest vacuole compartment found across the optical sections (preferably in, but not limited to, the perinuclear region, i.e. above the nucleus and below the cortical cytoplasm layer largely devoid of vacuoles [6]), obtained using built-in Fiji functions. For any given root, all atrichoblasts with suitably positioned optical sections (Additional File 1) were analyzed (in some cases using more than one microscope field or optical section but always ensuring that a non-redundant set of cells is measured).
Data processing and statistics
For numeric data processing during TTI protocol development and TTI determination, a Windows PC running Jupyter Notebook as a part of the Anaconda 2.5.3 package has been used together with scripts written in the Python language version 3.11.7. Raw TTI data processing was performed using Scipy library [22] v. 1.11.4.
The Matplotlib library [23] v. 3.8.0 was used to generate graphs, while data tables were produced using the pandas library v. 2.1.4 [24, 25]. Simple data processing was performed in MS Excel. In some cases, R version 4.4.2 [26] was used for plotting, statistical analysis, and additional data processing, employing the ggplot2 and dplyr libraries from the Tidyverse package [27]. Some plots were produced also using the R-based BoxPlotR tool [28] or Matplotlib 3.6.3 [23] employing Seaborn 0.13.2 [29].
The Shapiro–Wilk test (α = 0.05; [30]) was applied to assess data distribution. Data that did not conform to the normal distribution were analyzed using the Mann–Whitney test [31], while Student’s t test [32] was used for normally distributed data. For statistically significant differences (i.e., P < 0.05), effect size r was determined as described in [33]; r of 0.5 or greater was considered a strong effect, 0.3 < r < 0.5 as medium strong effect. Asterisks were used in figures to denote significant statistical differences with P < 0.01 (**) or 0.01 ≤ P < 0.05 (*).
Results
Rationale for developing the TTI metric
The VMI metric relies solely on measuring the size of the largest vacuolar compartment crossed by a single representative optical section of a cell (Fig. 1a; see “Methods” for details of optical section selection). However, cells with large vacuoles of similar size and shape may differ, possibly dramatically, in the presence, number and organization of additional smaller vacuolar compartments crossed by the optical section, as illustrated by a hand-drawn simulated dataset comprising two populations with similar large vacuolar compartments but dissimilar organization of smaller ones (Fig. 1b). In spite of a readily visible difference between these populations, there is no significant difference between their average VMI values (Fig. 1c).
We propose a new metric, the Tonoplast Topology Index (TTI), that should capture not only the progress of the large vacuole development reflected in the VMI, but also the overall complexity of vacuole shape, which should be reflected in the number of tonoplast intersections with a defined linear transect across a cell´s optical section. While the transect could be positioned in various ways, we chose to use the cell diagonal. The number of transect-tonoplast crossings, normalized to the transect length to compensate for cell size variation, can then serve as a metric of vacuole shape complexity. We thus defined the TTI as the number of tonoplast intersections divided by the diagonal transect length (Fig. 1a). Higher TTI indicates a more complex vacuome organization, regardless of the underlying phenomena (which could include, e.g., vacuole fragmentation, reduced tubular network expansion, delayed coalescence or even increased number of transvacuolar strands) or their causes (i.e. changes in membrane trafficking or cytoskeletal organization). As expected, TTI determination on the simulated dataset using visual peak counting and transect length measurements in Fiji revealed a significant difference between the two model populations (Fig. 1c).
A semi-automated pipeline for TTI determination
While the TTI can be determined manually, partial automation of the procedure can substantially speed up the process while enhancing reproducibility and decreasing the probability of human-introduced errors. We thus developed a two-stage protocol for determining the TTI (Fig. 2a), where the first stage involves manual transect definition with simultaneous cell length measurement, and subsequently employs Fiji´s built-in Plot profile function to measure and export intensity profiles of cell transects. The second stage is performed in the Jupyter notebook environment and analyses the exported profile data to identify peaks corresponding to membrane crossings, counts the peaks, divides the peak count by transect length and outputs a table containing matched cell length and TTI values for individual cells. We are providing the necessary scripts and detailed, beginner-friendly instructions for performing the procedure as Additional File 2.Fig. 2. Semi-automated TTI determination procedure. a A flowchart of the TTI determination pipeline. b Examples of optical section selection (top, the middle one of three sections labelled as good is at the optimal position), and acceptable histogram smoothing and peak detection (bottom). Both examples correspond to the cell and diagonal transect shown in Fig. 1a. c Comparison of results of manual and semi-automated TTI determination on the simulated dataset from Fig. 1b (with results of cell size measurements shown as an inset). The same transect direction was used for both approaches.
Both stages involve steps that either require manual intervention or include an opportunity to perform manual parameter adjustments. First of them is the selection of an appropriate image from the confocal stack during the first (Fiji) stage. Measurement of comparable focal planes is crucial, as the tonoplast structure varies greatly across the cell’s Z dimension. Similar to VMI determination, the measurements should be performed as close to the cell “equator” as possible but outside the tonoplast-free region around the nucleus. Typically, a suitable optical section can be located a few micrometers above the nucleus, which appears as a roundish area devoid of tonoplast membranes, within the top half of the cell’s Z dimension but below its top third (Fig. 2b; detailed visual instructions for consistent focal plane selection are provided in Additional File 1).
Second manual step is the selection of cells to be analyzed and positioning of longitudinal and diagonal transects of each cell for determining cell length and TTI, respectively. The diagonal transects should be oriented uniformly to avoid observer-induced bias, and non-redundant sets of cells have to be measured when analyzing several planes of focus or image fields.
Third, the intensity profile generation step involves image denoising by repeated rounds of smoothing. The number of smoothing steps can be adjusted depending on the quality of microscopy images, with the preset value optimized for our benchmark data set (see below) but with a possibility of modification (Fig. 2b; see also detailed manual in Additional File 2 and examples in Additional File 1).
Lastly, the second (Jupyter notebook) stage of the procedure involves application of several filters employing adjustable parameters (Table 1) to identify genuine maxima while avoiding detection of false positive peaks (local maxima close to the background; Fig. 2b). The default parameter values are optimized for our benchmark images but adjustment may be required for other data. It is advisable to optimize the stage 1 denoising, as well as the stage 2 filtering, on a subset of source images prior to analyzing large datasets. Some examples of effects of parameter modification are provided in Additional File 1; the readers are also encouraged to experiment with the provided benchmark data or their own images. At suitable parameter settings, most cells should yield only correct peak detection, with misdetection affecting, on average, less than one membrane per transect and no more than 5% of membranes overall (although occasional false negatives or false positives can never be completely avoided).Table 1. Overview of adjustable parameters of stage 2ParameterDescriptionBackgroundFilters out all peaks below the set value (i.e., background noise). If increasing this value, all analyzed plots should have similar intensity ranges to avoid loss of peaks in low intensity plots.Med_ratioMultiplier of the median intensity values from each separate profile, used to filter out maxima lower than medianmed_ratio value. Useful to compensate for varying intensity ranges among different profiles, not affected by changing bit depth of the source images. Typically around 1 or slightly lower.Min_ratioMultiplier of the minimum intensity values from each separate profile, used to filter out maxima lower than minimummin_ratio value. Useful to compensate for varying intensity among different profiles, not affected by changing bit depth of the source images. Typically higher than 1.DistanceMinimum distance between two maxima to be considered as separate membrane crossings (scale-independent, takes next/previous value in plot as 1). Value can be adjusted to avoid artificial peak clusters coming from a single membrane.ProminenceMinimum height of a peak compared to the closest local minima. Adjust to avoid loss of minor peaks next to major peaks.SigmaGaussian smooth. Can be used for further denoising in addition to stage 1 smoothing to eliminate artifactual minor peaks.
To test the performance of our pipeline, we used it to analyze our simulated dataset (Fig. 1b) and examined the correlation between results of manual and semi-automated TTI determination for individual cells. Although the semi-automated method tends to overestimate the TTI due to detecting weak satellite lines on many of the hand-drawn vacuole contours (which is unlikely to occur on real-life microscopy data), we found an overall strong correlation (close to very strong sensu [34]) between results of manual and semi-automated analysis for both the TTI and cell length (Fig. 2c).
A benchmark data set for TTI procedure validation
In order to validate the TTI determination procedure, we decided to test it by reproducing the previously reported difference in vacuolar organization in atrichoblasts of roots grown on agar media surface compared to those penetrating the agar media, where cells from surface-grown roots were found to have higher VMI than those from roots embedded in the medium [9]. Confocal images obtained from both types of roots are further referred to as the “top” (surface-grown) and “deep” (agar-embedded) population. Our plants were expressing the specific tonoplast marker VHP1:mGFP, and were found to be free of visually obvious marker oligomerization artifacts known as “bulbs”, in agreement with the original description of the marker [14]. Following the published report [9], we focused on the atrichoblasts, which were readily distinguishable from trichoblasts in the late meristematic zone by their less fragmented vacuolar compartments and overall weaker tonoplast signal (Fig. 3a).Fig. 3. Characterization of the benchmark data set using the VMI metrics. a Examples of optical sections of transition and elongation zone rhizodermis of a root grown on the agar surface (labelled “top”) or embedded inside the agar medium (labelled “deep”). Rows of atrichoblasts are marked by arrows. b Atrichoblast length distribution for a set of transition and elongation zone cells from top and deep agar-grown roots (N denotes the number of roots, n the number of cells). c Comparison of atrichoblast VMI distribution for the cell set from (b). d QQ plots of pooled VMI values from top and deep agar showing that the log-transformed VMI value distribution for the dataset from (c) deviates from normal distribution less than that of raw data. Shapiro–Wilk P-values are shown. e Comparison of log-transformed atrichoblast VMI distribution for the same data as in (c). Mann–Whitney P-values are shown in (b), (c) and (e), accompanied by strength of effect r values for significant differences.
The atrichoblasts of both populations included in our analyses exhibited a similar asymmetric cell length distribution; although the average cell length was somewhat smaller in the “deep” group, the difference was not statistically significant (Fig. 3b). The distribution of VMI values of both populations was even more noticeably asymmetric (Fig. 3c), and significantly deviating from the normal distribution. A quantile–quantile (QQ) plot suggested a possible exponential distribution. Indeed, logarithmic transformation brought the VMI value distribution somewhat closer to normal, with its shape possibly corresponding to the Cauchy distribution (Fig. 3d). Therefore, the Mann–Whitney test had to be used to assess the statistical significance of observed VMI differences. For both original and log-transformed data (Fig. 3c, e), we found significantly higher VMI values in the “top” population compared to the “deep” one.
We thus successfully confirmed that our image data set behaves in agreement with the previous report that cells from roots penetrating into agar media possess more fragmented vacuoles compared to those from surface-grown ones [9], and proceeded to use the same images (also deposited in the EMBL-EBI BioImage Archive repository, accession number S-BIAD2226) as a benchmark set to test the performance of our TTI procedure.
Comparison of the TTI and VMI metrics on the benchmark data set
We have first used the semi-automated TTI procedure on a subset of 70 cells from the “top” population of the benchmark data set to examine the effect of the diagonal transect direction on the analysis outcome. There was no difference between average TTI values due to transect direction (Fig. 4a), indicating that, if a sufficiently large set of cells is analyzed, the transect direction does not affect the analysis outcome. Although vacuole organization in individual cells is typically asymmetrical (see Fig. 3a), we did not detect any evidence of cell chirality on the supracellular scale.Fig. 4. Testing the TTI method on the benchmark data set. a Comparison of TTI values obtained on a subset of the benchmark data set using the indicated directions of the diagonal transect (Mann–Whitney P value shown). b Comparison of atrichoblast TTI distribution among roots grown on top of the agar medium or deep inside (data from the same set of cells as in Fig. 3, Mann–Whitney P value and strength of effect r value shown). c QQ plots of pooled TTI values from top and deep agar indicating near-normal distribution. Shapiro–Wilk P-value shown. d Pearson correlation of TTI and log-transformed VMI values for individual cells from the benchmark data set, indicating strong negative correlation between the two metrics.
Analysis of the complete benchmark image set using the TTI pipeline revealed significantly higher TTI values (and thus more complex tonoplast topology, as already suggested by VMI determination results) in cells from agar-embedded roots compared to those grown on the media surface (Fig. 4b). TTI value distribution was near-normal but not normal, possibly corresponding to the Couchy distribution (Fig. 4c), justifying the use of the Mann–Whitney test for statistical evaluation.
Both methods thus yielded comparable results, documenting higher vacuole shape complexity, associated with smaller size of individual vacuolar compartment sections, in the “deep” population compared to the “top” one. Furthermore, comparison of TTI and log-transformed VMI values for the same cells revealed a strong negative correlation between the results of both methods (Fig. 4d), showing that they produce mutually consistent results.
To further examine the effects of root growth conditions on vacuolar organization by both methods, we divided the benchmark cell population into subgroups according to cell length. Splitting the population into quartiles based on cell length (with quartile border values determined from pooled “top” and “deep” populations) allowed us to compare cells not only of similar size, but also of approximately similar developmental age (Fig. 5). Independent from the measurement method, significant differences with moderate to strong effect sizes were confined to the first three quartiles (Q1-Q3), corresponding to smaller cells located in the late meristematic and transition zones, while large (Q4) cells did not exhibit statistically significant differences. Thus, the effects of root growth conditions are only detected in small cells but diminish or disappear later due to developmental progression and the expansion of the central vacuole. Importantly, both the conventional VMI method and the newly proposed TTI metric are comparably sensitive with respect to detection of vacuolar organization differences present in our benchmark image data set.Fig. 5. Comparison of TTI and VMI metrics for size-matched cells. Top: Log-transformed VMI value distribution in atrichoblast populations from top and deep agar-grown roots (the same dataset as in Fig. 3 and Fig. 4). Bottom: TTI value distribution for the same set of cells. For both plots, the cell population was split into subsets corresponding to quartiles of pooled cell length distribution. Mann–Whitney P-values, separately calculated for the indicated pairs and subjected to Benjamini–Hochberg correction for multiplicity, are shown, together with strength of effect r values for pairs showing significant differences.
To further examine the performance of the TTI metric, we evaluated the difference in tonoplast organization between comparably sized mid-elongation zone trichoblasts and atrichoblasts of surface-grown roots from our benchmark images set. Even when evaluating substantially fewer cells than in the previous examples because of a conscious effort to select cells of similar length, both TTI and VMI exhibited comparable ability to detect increased vacuole shape complexity in the trichoblasts, i.e., smaller vacuolar compartments manifesting as lower VMI and higher TTI (Fig. 6).Fig. 6. Comparison of vacuolar organization in trichoblasts and atrichoblasts. Left: Log-transformed VMI value distribution in trichoblast and atrichoblast populations from mid-elongation zone rhizodermis of top agar-grown roots. Center: TTI value distribution for the same set of cells. Right: Cell length distribution of the same set of cells. Mann–Whitney P-values, are shown, together with strength of effect r values for pairs showing significant differences.
Discussion
Study of vacuole organization often relies on qualitative descriptions or 3D modelling, typically restricted to a specific cell type such as stomata guard cells [35], tobacco BY-2 cells [36], or rhizodermis cells (see Background; reviewed also in [11]). While qualitative description may be sufficient if morphological differences are clearly visually distinguishable, quantitative approaches are necessary to address more subtle phenotypes, especially in cells with complex vacuolar structure.
While quantitative metrics of tonoplast morphology evaluation are already being used (see Background and [37]), their application is hampered by technical complexity and computational demands of high data volume image analysis. The Vacuolar Morphology Index (VMI, [6]) is a popular and accessible option especially for following vacuole development in root tissues, as it does not require proprietary software or advanced computational skills. However, it may not capture all aspects of tonoplast organization, as it focuses solely on the size of the largest vacuole compartment detected in a single optical section of a given cell.
Several alternatives to VMI have been introduced. Examples include vacuole volume, surface to volume ratio, or vacuolar occupancy, i.e. the vacuole to cell volume ratio [7], determined in the open-source MorphoGraphX 3D segmentation and quantification environment [38] or in the commercial Imaris software [39, 40]. Despite increasing availability of relevant software in microscopy facilities, transferring these tools (and related skills) to diverse experimental environments may be challenging. A “vacuole shape index” defined as the ratio between the width and length of the largest vacuole compartment (therefore sharing the detection limits of the VMI) has been recently used alongside VMI (renamed to “vacuole area”), and occupancy [10], and vacuole compactness has been quantified by measuring the tonoplast to plasmalemma distance at cell corners [15]. Overall, there is so far an obvious lack of standardized workflows for quantitative description of vacuole organization, and development of a new, simple metric that can be scaled up for large datasets may be beneficial.
Here we present the Tonoplast Topology Index (TTI), a new metric that provides a complementary quantitative perspective on vacuolar architecture Our protocol is amenable to upscaling, somewhat reduces the requirement for subjective observer´s decisions (such as the choice of the largest vacuolar compartment) that could be introducing artifacts, and may represent an alternative or valuable addition to currently available tools. While TTI shares an important limitation of the VMI, namely reliance on a single optical section and therefore inability to assess vacuole shape, fragmentation or connectedness, it should be able to detect additional aspects of vacuole organization besides those captured by the VMI, since it also reflects the complexity of smaller vacuolar compartments. Although we could not yet demonstrate this feature on real-life data, we believe that it may help to capture subtle changes in vacuole organization not detectable using the VMI metric. For example, it may be interesting to use TTI for re-assessment of previously reported qualitative effects of brassinazole treatment, which in some situations appears to noticeably increase complexity of small vacuolar compartments without much apparent change in larger ones [41].
While establishing a benchmark data set for testing our method, we successfully reproduced the previously reported difference in vacuolar organization in the rhizodermis of roots grown on the surface of agar media compared to those embedded inside, which may be reflecting the plant´s response to varying media stiffness [9]. However, characterization of our benchmark data set using the conventional VMI metric alerted us to the notably asymmetric distribution of VMI values in real-life rhizodermis cell populations that is, in our view, only rarely reflected in reported vacuole organization analyses. Multiple studies [6, 7, 15, 17, 42] report statistical analyses performed using Student’s t-test on VMI data that are unlikely to meet assumptions of normality and equal variance, and in some cases even display clear asymmetry or variance differences that are not accounted for [15, 17]. Non-parametric statistical methods that do not assume normal data distribution, although clearly more appropriate, were used only in some cases [9, 16, 43]. Although the importance of choosing a non-parametric test for data deviating from normal distribution may be reduced for very large samples, typically consisting of hundreds of individual values [44, 45], most studies employ a relatively small number of individual plants per sample and cells per plant; we could find only a single report involving 100 or more cells per sample [15].
In contrast to VMI, our new TTI method generates data exhibiting a distribution that is generally symmetric and relatively close to normal. At the same time, the semi-automated procedure enables large scale analyses, and collection of several hundred data points per sample, though laborious, is possible (if enough good quality confocal images can be obtained). Thus, while use of non-parametric methods for statistical evaluation is still recommended for non-normally distributed data, with a sufficiently large sample [45], parametric tests could be employed. For an example, ANOVA can be used if multiple treatments, genotypes or combinations thereof are compared, substantially simplifying data analysis.
Besides of successfully employing TTI to detect vacuole architecture differences between rhizodermis atrichoblasts of roots grown on the surface and inside agar medium [9], we demonstrated that it is able to detect also the known difference in vacuome organization between trichoblasts and atrichoblasts [21] with sensitivity similar to that of VMI. In addition, while the present study was under review, we reported application of the TTI method alongside VMI to document altered vacuole organization in plants with mutationally or pharmacologically altered formin function, as well as the effects of actin disruption on vacuole architecture [46]. Both methods produced qualitatively similar results in this case as well, although VMI appeared to be somewhat more sensitive.
We believe that our new TTI metric can contribute towards a more transparent and reproducible framework for quantitative analysis of tonoplast organization. Moreover, while originally intended for measurements of vacuole shape complexity, the method might be suitable for adaptation for other applications involving quantification of geometrically complex biological structures (the endoplasmic reticulum might be one such example).
Supplementary Information
Supplementary Material 1. A visual guide to interactive steps in TTI determination (*.pdf file). Supplementary Material 2. Archive file in *.zip format containing scripts and a detailed manual for performing TTI determination.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1R core team. R: a language and environment for statistical computing. https://www.R-project.org. Accessed on 31 July 2025.