In Silico Investigation Reveals IL-6 as a Key Target of Asiatic Acid in Osteoporosis: Insights from Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation
Wanatsanan Chulrik, Aman Tedasen, Nateelak Kooltheat, Rungruedee Kimseng, Thitinat Duangchan

TL;DR
This study uses computer modeling to show that asiatic acid may help treat osteoporosis by targeting the IL-6 protein, which plays a key role in inflammation and bone health.
Contribution
The study identifies IL-6 as a novel molecular target of asiatic acid in osteoporosis using an integrative in silico approach.
Findings
Network pharmacology identified 135 overlapping targets between asiatic acid and osteoporosis, with IL-6 as a key hub.
Molecular docking and dynamics simulations confirmed strong and stable binding of asiatic acid to IL-6.
The PPAR signaling pathway is suggested as a potential mechanism for the anti-osteoporotic effects of asiatic acid.
Abstract
Background/Objectives: Osteoporosis is a multifactorial skeletal disorder in which chronic inflammation, dysregulated cytokine signaling, and metabolic imbalance contribute to excessive bone resorption and impaired bone formation. Asiatic acid has demonstrated bone-protective effects, but its molecular mechanisms in osteoporosis remain incompletely understood. This study aimed to investigate the anti-osteoporotic mechanisms of asiatic acid using an integrative in silico strategy. Methods: Network pharmacology analysis was performed to identify osteoporosis-related molecular targets of asiatic acid. Molecular docking was used to predict the binding modes and affinities between asiatic acid and its target proteins. Molecular dynamics simulation was used to assess the structural stability and interaction persistence of the asiatic acid–protein complex. Results: Network pharmacology…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
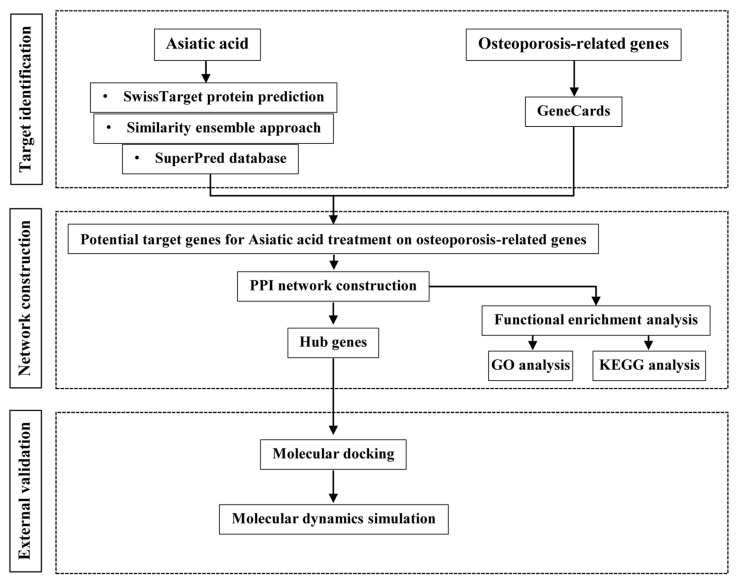 Figure 1
Figure 1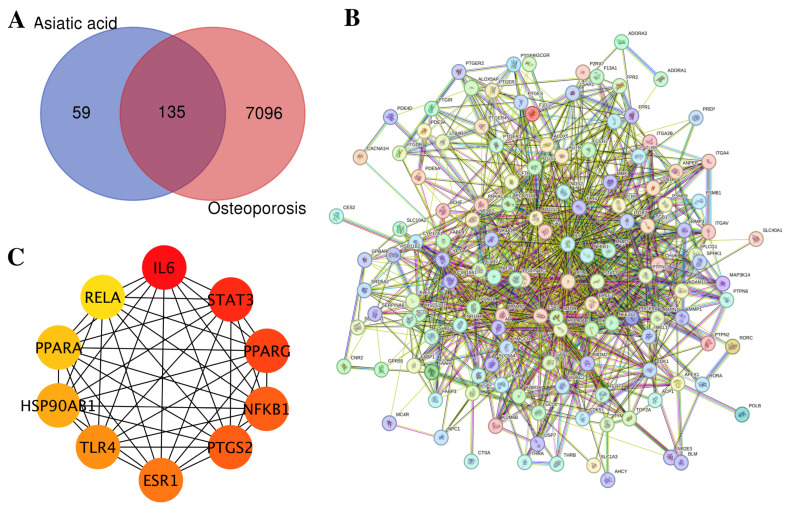 Figure 2
Figure 2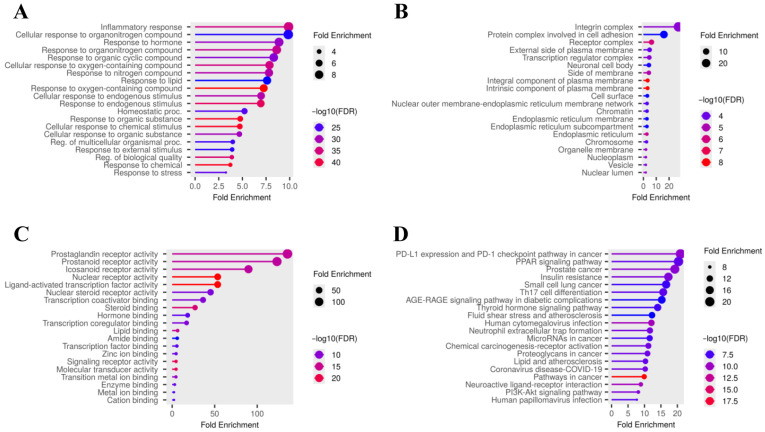 Figure 3
Figure 3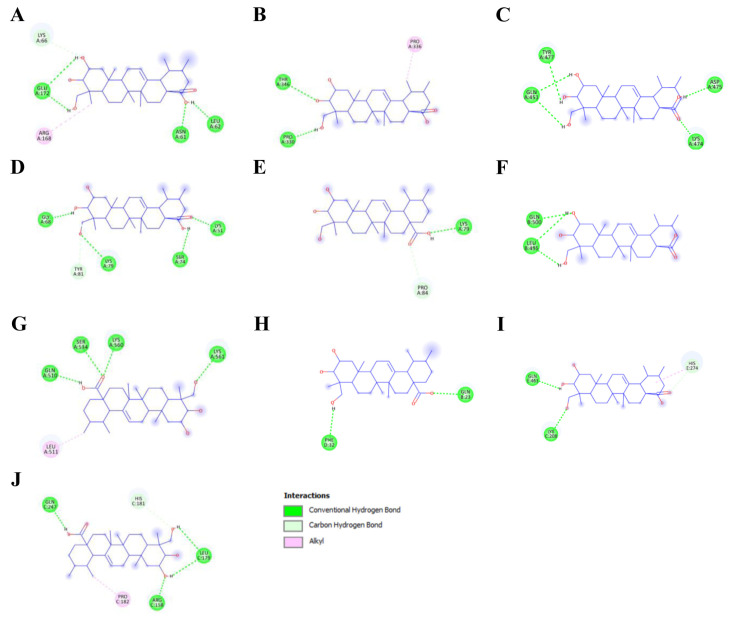 Figure 4
Figure 4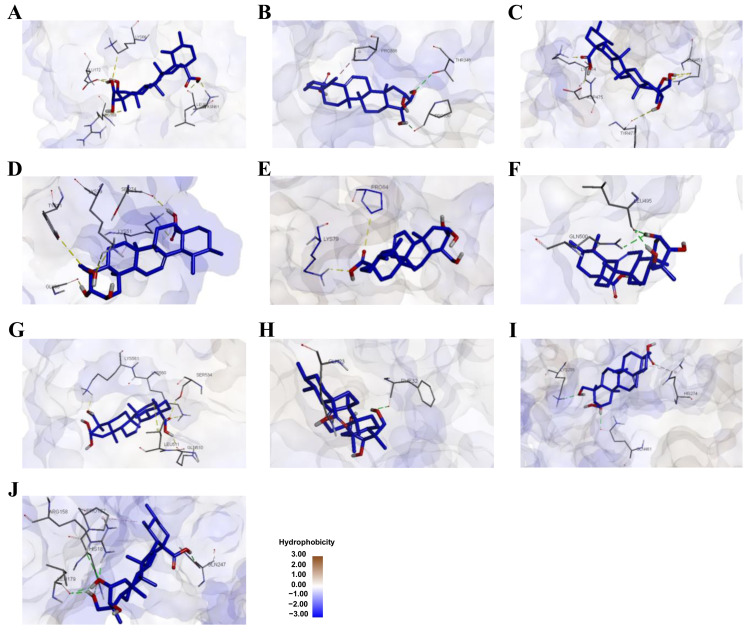 Figure 5
Figure 5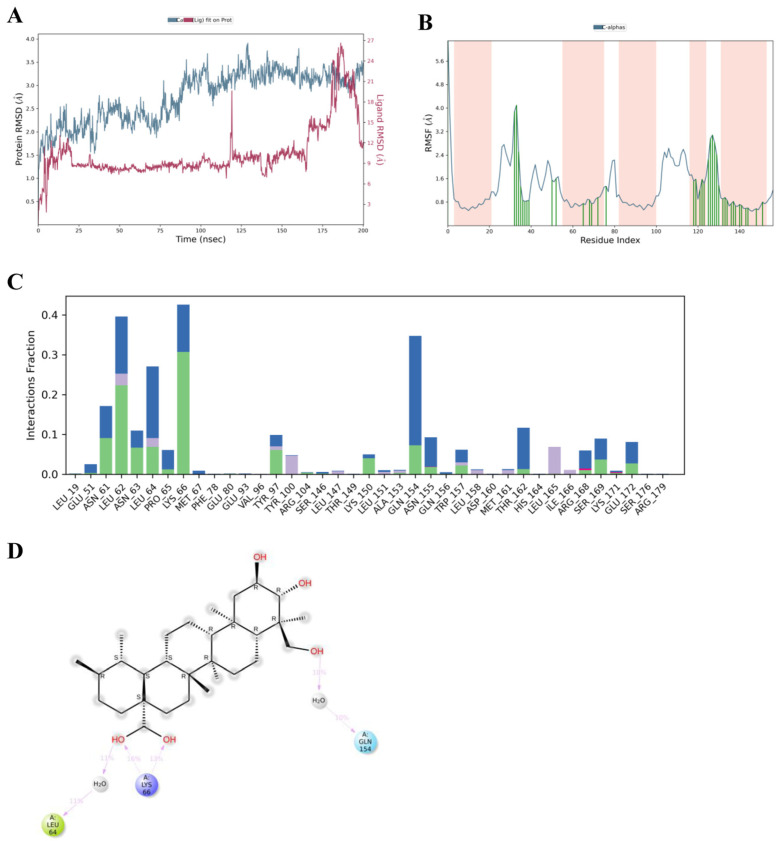 Figure 6
Figure 6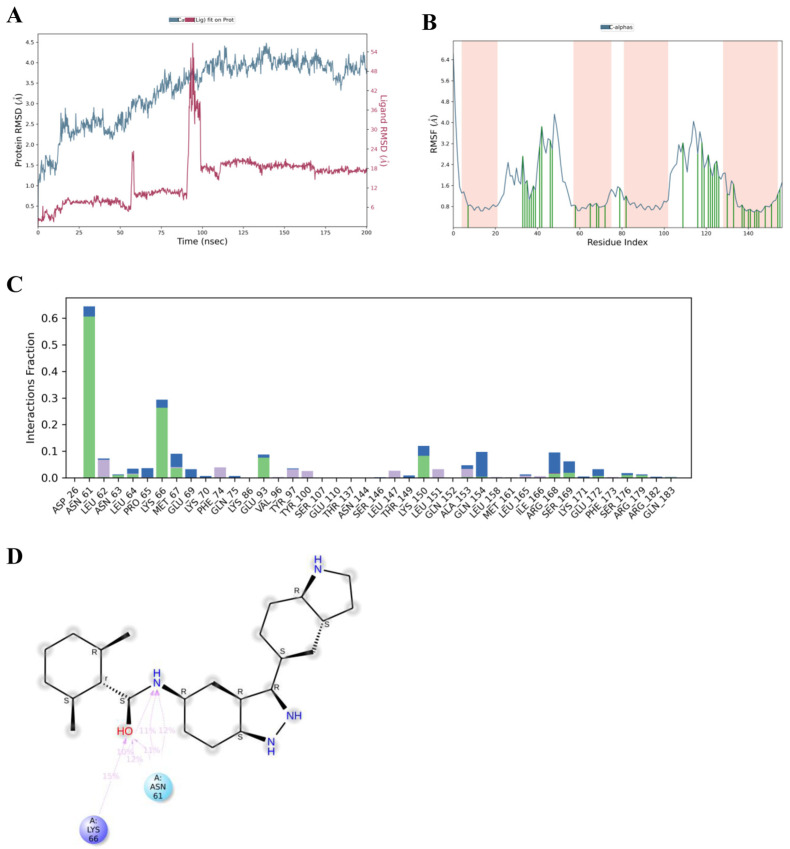 Figure 7
Figure 7- —Walailak University under the New Researcher Development Scheme
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMedicinal Plants and Neuroprotection · Bone Metabolism and Diseases · NF-κB Signaling Pathways
1. Introduction
Osteoporosis is a prevalent metabolic bone disorder characterized by decreased bone mass and deterioration of bone microarchitecture, leading to increased fracture risk and substantial morbidity and mortality worldwide. With population aging, osteoporosis has emerged as a major public health concern, particularly among postmenopausal women and older adults [1,2]. Although estrogen deficiency is a primary driver of postmenopausal osteoporosis, accumulating evidence indicates that osteoporosis is a multifactorial disease involving chronic low-grade inflammation, dysregulated cytokine signaling, and altered metabolic regulation of bone remodeling [3,4,5,6]. Inflammatory cytokines play a pivotal role in bone homeostasis by modulating the balance between osteoblast-mediated bone formation and osteoclast-driven bone resorption. Among these, interleukin-6 (IL-6) is a key mediator linking inflammation to bone loss, promoting osteoclastogenesis through activation of the gp130/Janus kinase (JAK)/signal transducer and activator of transcription 3 (STAT3) pathway and upregulation of receptor activator of nuclear factor-κB ligand (RANKL) [7,8]. In parallel, nuclear factor-κB (NF-κB) signaling is essential for osteoclast differentiation downstream of RANKL–RANK interaction and is further amplified under inflammatory and estrogen-deficient conditions [9]. Metabolic regulators such as peroxisome proliferator-activated receptor γ (PPARγ) also contribute to osteoporosis by shifting mesenchymal stem cell differentiation toward adipogenesis at the expense of osteoblastogenesis, thereby impairing bone formation [10]. These pathways are highly interconnected, underscoring the complex, multi-target nature of osteoporosis pathophysiology.
Current pharmacological treatments for osteoporosis, including bisphosphonates, selective estrogen receptor modulators, and monoclonal antibodies, effectively reduce fracture risk but are associated with limitations such as adverse effects, high cost, and reduced long-term adherence [11,12]. Consequently, there is growing interest in identifying safer therapeutic agents capable of modulating multiple disease-relevant pathways simultaneously. Natural compounds with pleiotropic biological activities have attracted particular attention as potential adjuncts or alternatives for osteoporosis management [13]. Asiatic acid, a pentacyclic triterpenoid derived from Centella asiatica, has been reported to exert anti-inflammatory, antioxidant, and metabolic regulatory effects in various disease models [14,15,16]. Emerging experimental evidence suggests that asiatic acid can attenuate osteoclastogenesis and protect against bone loss, primarily through suppression of NF-κB signaling and modulation of adipogenic differentiation [17,18]. However, these studies have largely focused on individual pathways or downstream effects, and a comprehensive understanding of asiatic acid’s molecular targets and system-level mechanisms in osteoporosis remains lacking. In particular, whether asiatic acid directly interacts with upstream inflammatory mediators such as IL-6 and STAT3, and how these interactions integrate with known NF-κB and PPAR signaling networks, has not been systematically explored.
Network pharmacology, combined with molecular docking and molecular dynamics simulation, offers a powerful systems-level approach to elucidate the multi-target and multi-pathway mechanisms of bioactive compounds in complex diseases [19,20,21]. By integrating target prediction, pathway enrichment, structural interaction analysis, and dynamic stability assessment, this framework enables mechanistic hypothesis generation beyond single-target paradigms.
Accordingly, the aim of the present study was to elucidate the molecular mechanisms underlying the anti-osteoporotic potential of asiatic acid using an integrative in silico strategy. Specifically, network pharmacology, molecular docking, and molecular dynamics simulation were employed to predict the putative targets of asiatic acid, evaluate its binding affinities with osteoporosis-related proteins, and assess the stability of the asiatic acid–protein complex. Through these integrative computational approaches, this study seeks to provide mechanistic insight into the potential of asiatic acid as a natural therapeutic candidate for the prevention and management of osteoporosis.
2. Materials and Methods
2.1. Network Pharmacology Analysis
2.1.1. Prediction of Asiatic Acid and Osteoporosis Targets
Network pharmacology analysis was conducted to identify potential molecular targets associated with asiatic acid and osteoporosis. The identification of asiatic acid targets (species: Homo sapiens) was carried out using three databases, namely SwissTargetPrediction (https://www.swisstargetprediction.ch; accessed on 5 March 2025), the Similarity ensemble approach (SEA) (https://sea.bkslab.org; accessed on 5 March 2025), and SuperPred (https://bio.tools/superpred; accessed on 5 March 2025). Osteoporosis-related targets were retrieved from the GeneCards database (https://www.genecards.org; accessed on 5 March 2025). The overlapping targets between asiatic acid and osteoporosis were visualized using a Venn diagram online tool (https://bioinformatics.psb.ugent.be/webtools/Venn/; accessed on 5 March 2025).
2.1.2. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis
GO and KEGG pathway enrichment analyses were performed using ShinyGO version 0.82 (http://bioinformatics.sdstate.edu/go/; accessed on 5 March 2025). Enrichment results were evaluated based on fold enrichment values and −log10 (false discovery rate, FDR) values, with p < 0.05 considered statistically significant. Cytoscape version 3.10.3 (www.cytoscape.org; accessed on 5 March 2025) was used to visualize the networks between asiatic acid and its anti-osteoporotic targets. The cytoHubba plugin (https://apps.cytoscape.org/apps/cytohubba; accessed on 5 March 2025) was employed to identify hub targets based on degree centrality.
2.1.3. Construction of Protein–Protein Interaction (PPI) Network
PPI network plays a pivotal role in a variety of cellular biological processes, which are crucial for understanding molecular mechanisms, cellular processes, and pathways, thereby facilitating drug discovery [22]. We therefore constructed a PPI network using the STRING database (https://string-db.org; accessed on 5 March 2025) with the species set to Homo sapiens and a medium-confidence score (>0.4) to balance interaction reliability and network completeness in the context of osteoporosis. The resulting network enabled the identification of highly connected proteins, which were considered hub targets based on their degree values.
2.2. Molecular Docking
2.2.1. Protein Preparation
Molecular docking analysis was performed using AutoDockTools (ADT) version 4.2 [23]. Three-dimensional (3D) crystal structures of 10 target proteins (receptors) were obtained from the Research Collaboratory for Structural Bioinformatics (RCSB) Protein Data Bank (PDB) (https://www.rcsb.org, accessed on 5 March 2025) with a resolution of less than 3.5 Å, in PDB format. Protein structures were prepared by removing ligands/inhibitors and water molecules using the Visual Molecular Dynamics (VMD) software version 2.0.0. Polar hydrogen atoms were subsequently added to the proteins using ADT software version 4.2. The prepared protein structures were saved in PDBQT format files (PDB, Partial Charge (Q), and Atom Type (T)).
2.2.2. Ligand Preparation
The 3D structures of asiatic acid and native ligands were retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov, accessed on 5 March 2025). Ligand structures were converted to PDB format using the online SMILES Translator and Structure File Generator (https://cactus.nci.nih.gov/translate/, accessed on 5 March 2025). Polar hydrogen atoms were added to the ligands and further prepared in PDBQT format files using ADT.
2.2.3. Docking Protocol
Semi-flexible molecular docking was conducted, in which the receptor proteins were treated as rigid while the ligand molecules remained flexible. Grid maps were generated using ADT. Partial atomic charges of the proteins and ligands were assigned using the Gasteiger–Marsili method [24]. A blind docking strategy was employed; therefore, a cubic grid box was defined to encompass the entire receptor structure for each target protein, without assuming a predefined active pocket. The grid center was positioned at the geometric center of each protein, and the grid dimensions were individually adjusted to fully cover the receptor surface, allowing exploration of all potential ligand-binding regions. Docking simulations were performed using the Lamarckian genetic algorithm with 50 independent runs and a population size of 200 to identify energetically favorable binding sites for asiatic acid. The docked conformation with the lowest binding energy (ΔG_docking_; kcal/mol) and the corresponding inhibitory constant (K_i_) in the most populated cluster was selected for each protein. Docked conformations with the lowest binding energies and interaction residues were visualized using BIOVIA Discovery Studio Visualizer (Dassault Systèmes BIOVIA Corp., San Diego, CA, USA).
2.3. Molecular Dynamics Simulation
Molecular dynamics (MD) simulation, a powerful computational technique, simulates the physical movements of atoms and molecules over time and provides insights into the dynamic behavior and properties of biomolecular systems under various conditions [25]. MD simulation was conducted to further investigate the stability, interaction patterns, and binding dynamics of asiatic acid and MD2-TLR4-IN-1 (a native IL-6 inhibitor) with the IL-6 protein. Discovery Studio’s protein preparation technique was used to prepare the IL-6 protein structure, including the protonation state of any ionizable side chain. The AMBER ff14SB and generalized AMBER force field were used for protein and ligand, respectively. For the simulations, the system was built using the TIP3P solvation model and centered within an orthorhombic cubic box measuring 10 Å × 10 Å × 10 Å, filled with single-point charge water molecules. The system was neutralized by adding random counterions (sodium and calcium ions), and an isosmotic state was maintained by adding 0.15 M NaCl. The molecular dynamics simulation was performed under an NPT (constant number of particles, constant pressure, and constant temperature) ensemble at 310 K and 1.01 bar, with a 200-nanosecond simulation duration. Finally, root mean square deviation (RMSD) and root mean square fluctuation (RMSF) were calculated to gain insight into the stability and identify flexible regions in the IL-6–asiatic acid and IL-6–MD2-TLR4-IN-1 complexes throughout the simulation. A detailed flowchart depicting the study process is presented in Figure 1.
3. Results
3.1. Network Pharmacology Analysis of Asiatic Acid Targets Related to Osteoporosis
A network pharmacology analysis was conducted to investigate the potential molecular mechanisms underlying the anti-osteoporotic effects of asiatic acid. A total of 194 predicted gene targets of asiatic acid were identified using SwissTargetPrediction, SEA, and SuperPred. In parallel, 7231 osteoporosis-related targets were retrieved from the GeneCards database. Intersection analysis revealed 135 overlapping targets, suggesting that asiatic acid may exert multi-target therapeutic effects (Figure 2A). The top 10 of 135 overlapping targets is presented in Table 1, whereas the complete list of all 135 overlapping asiatic acid–osteoporosis targets is provided in Table S1. The STRING database revealed a PPI network comprising 135 nodes and 1019 edges, with an average node degree of 15.1 (Figure 2B). Subsequent degree centrality analysis using the cytoHubba plugin in Cytoscape identified the top 10 hub genes from the PPI network based on their topological importance. The hub genes were ranked in descending order according to their cytoHubba scores as follows: IL-6, STAT3, PPARG, NFKB1, PTGS2, ESR1, TLR4, HSP90AB1, PPARA, and RELA, indicating their relative centrality and potential regulatory significance within the network (Figure 2C). In the hub gene network, node color reflects cytoHubba ranking (red = highest, yellow = lowest) (Figure 2C).
To explore the biological functions of asiatic acid in relation to osteoporosis-related targets, a comprehensive GO enrichment analysis was performed on the 135 candidate targets using ShinyGO. The analysis yielded 1789 significant GO terms in total, encompassing 1000 biological process terms, 228 cellular component terms, and 561 molecular function terms. The top 20 GO biological process enrichment terms were primarily associated with inflammatory response, response to hormone, response to oxygen-containing compound, response to organic cyclic compound, and homeostatic process (Figure 3A and Table 2). Cellular component analysis revealed enrichment in the integrin complex, receptor complex, protein complex involved in cell adhesion, and external side of the plasma membrane (Figure 3B and Table 2). Molecular function analysis identified nuclear steroid receptor activity, nuclear receptor activity, ligand-activated transcription factor activity, and hormone binding (Figure 3C and Table 2). Moreover, KEGG pathway analysis was performed to elucidate the relevance of asiatic acid to osteoporosis-related pathways. Among the 184 identified pathways, the top 20 pathways are presented in Figure 3D and Table 2, including the PI3K–Akt signaling pathway, PPAR signaling pathway, Th17 cell differentiation, and the thyroid hormone signaling pathway. These findings highlight the pleiotropic potential of asiatic acid in regulating several osteoporosis-associated pathways through multiple targets.
3.2. Molecular Docking Analysis of Asiatic Acid with Osteoporosis-Related Proteins
Based on the network pharmacology analysis, 10 key osteoporosis-related proteins encoded by predicted target genes, including IL-6, STAT3, PPARG, NFKB1, PTGS2, ESR1, TLR4, HSP90AB1, PPARA, and RELA, were selected for molecular docking analysis. Molecular docking was performed using ADT software to predict the binding interactions between asiatic acid and the selected targets. The results showed that asiatic acid exhibited favorable binding affinities toward all selected proteins, with the binding energies in the most populated cluster lower than −5.00 kcal/mol, indicating strong binding activity. Among these targets, asiatic acid showed the strongest binding affinity toward IL-6 (−8.64 kcal/mol), followed by PPARγ (−7.92 kcal/mol), NF-κB p105 (−7.70 kcal/mol), cyclooxygenase-2 (COX-2) (−6.86 kcal/mol), PPARα (−6.41 kcal/mol), STAT3 (−6.17 kcal/mol), NF-κB p65 (−6.15 kcal/mol), estrogen receptor α (ESRα) (−5.98 kcal/mol), toll-like receptor 4 (TLR4) (−5.87 kcal/mol), and heat shock protein 90 β (HSP90-β) (−5.57 kcal/mol). Notably, asiatic acid exhibited comparable or lower binding energies than the corresponding native ligands for several key targets, particularly STAT3, PPARγ, NF-κB p105, ESRα, TLR4, and NF-κB p65. Detailed binding poses and key amino acid interactions of asiatic acid with the osteoporosis-related proteins are illustrated in Figure 4 and Figure 5, with quantitative docking parameters summarized in Table 3.
For IL-6, asiatic acid occupied a binding pocket similar to that of the native ligand and interacted with several conserved residues, including GLU172, LYS66, and ARG168 (Table 4 and Table S1). Specifically, asiatic acid formed hydrogen bonds with ASN61, GLU172, LEU62, and LYS66, along with a hydrophobic interaction involving ARG168. In addition, asiatic acid formed stable interactions with STAT3, including hydrogen bonds with THR346 and PRO330, along with hydrophobic interactions involving PRO336 (Figure 4 and Figure 5 and Table 4). Native ligand interaction details are provided in Figures S1 and S2 and Table S1. Collectively, these findings support the role of asiatic acid in targeting key proteins associated with osteoporosis, particularly within the STAT3, NF-κB, and PPAR signaling pathways.
3.3. Molecular Dynamics Simulations of Asiatic Acid and IL-6
The molecular docking analysis demonstrated a strong binding interaction between asiatic acid and IL-6. To further evaluate the stability of this interaction and validate the docking predictions under dynamic conditions, molecular dynamics simulations were performed. The structural stability and dynamic behavior of the asiatic acid-IL-6 complex were assessed using RMSD, RMSF, and ligand–residue interaction analyses. During the simulation, the RMSD of the IL-6 backbone increased rapidly during the initial equilibration phase (0–20 ns) and subsequently stabilized, fluctuating between 2.5 and 3.5 Å throughout the trajectory. This behavior indicates moderate conformational flexibility while maintaining overall structural stability. The RMSD of asiatic acid (Lig fit Prot) in complex with IL-6 remained relatively stable during most of the simulation, with average values of approximately 2–4 Å, suggesting that asiatic acid maintained a stable binding conformation within the IL-6 binding site. Transient RMSD increases were observed at approximately 120 ns and again during the late simulation phase (170–190 ns), suggesting short-lived conformational rearrangements of asiatic acid within the binding pocket rather than complete dissociation (Figure 6A). Overall, the RMSD profiles indicate a sustained association of asiatic acid with IL-6 over the 200 ns trajectory.
RMSF analysis revealed that most IL-6 residues exhibited fluctuations below 1.5 Å, indicating a generally rigid protein structure that may support stable ligand binding (Figure 6B). However, increased flexibility was observed in specific loop regions, particularly around residues 35–45 and 120–130. Ligand–residue interaction fraction analysis demonstrated that asiatic acid formed persistent contacts with several key IL-6 residues throughout the simulation. Notably, residues GLN154, ASN61, LEU62, LEU64, PRO65, and LYS66 exhibited the highest interaction fractions, indicating their major contribution to ligand stabilization within the binding pocket. Additional interactions were observed with ASN63, ASN155, TRP157, THR162, SER169, LEU165, GLU51, GLU172, LYS150, TYR97, TYR100, PRO65, and ARG168, suggesting an extended interaction network involving hydrophobic contacts, hydrogen bonds, and water bridges (Figure 6C). A 2D interaction map from the molecular dynamics simulation further confirmed that asiatic acid formed stable hydrogen bonds with LYS66, highlighting its anchoring role within the IL-6 AB-loop region. Moreover, water-mediated contacts with LEU64 and GLN154 were observed, suggesting that dynamic hydration also contributes to ligand stabilization (Figure 6D). Collectively, these results validate the docking predictions and confirm that asiatic acid binds stably to IL-6 through a combination of hydrogen bonding and hydrophobic interactions, supporting its potential role as an IL-6-modulating anti-inflammatory agent.
To provide a comparative assessment with asiatic acid, molecular dynamics simulations were also performed for MD2-TLR4-IN-1, a native IL-6 inhibitor, in complex with IL-6. Over the 200 ns trajectory, the protein RMSD (Cα atoms) remained stable within 2.5–4.0 Å, indicating moderate conformational flexibility and overall structural stability of IL-6. The ligand RMSD (Lig fit Prot) initially stabilized at approximately 0.5–1.0 Å, suggesting strong and consistent binding. Although transient spikes were observed near 50 ns and 95–100 ns, these deviations were followed by rapid re-stabilization, indicating temporary reorientation within the binding pocket rather than unbinding (Figure 7A).
The RMSF profile for the MD2-TLR4-IN-1-IL-6 complex revealed a pattern similar to that observed with asiatic acid, with most residues displaying low fluctuation amplitudes (<2 Å), suggesting good complex stability. Slightly elevated mobility was again noted in the loop regions around residues 35–50 and 115–130, consistent with flexible surface-exposed motifs involved in ligand accommodation (Figure 7B). Interaction fraction analysis revealed that ASN61 showed the highest interaction fraction among all residues, followed by LYS66, LYS150, GLN154, GLU93, ARG168, MET67, LEU62, SER169, ALA153, PHE74, PRO65, LEU64, GLU69, TYR97, TYR100, LEU147, LEU151, and GLU172, emphasizing their role as anchoring residues in IL-6 binding (Figure 7C). The 2D protein–ligand contact map of MD2-TLR4-IN-1 further confirmed the presence of stable and recurrent hydrogen bonding with ASN61 and LYS66, two residues that were also critically involved in asiatic acid binding (Figure 7D). These interactions showed consistent occupancy over the trajectory, with multiple polar contacts anchoring the ligand. In contrast to asiatic acid, MD2-TLR4-IN-1 exhibited slightly fewer water-mediated interactions, suggesting a more rigid and direct engagement with key IL-6 residues. Nonetheless, the shared contact residues between both ligands, especially ASN61 and LYS66, highlight the pharmacophoric significance of the AB-loop region in IL-6 inhibition.
4. Discussion
Network pharmacology has emerged as a pivotal role in systems biology, providing a powerful framework for discovering novel drug targets and elucidating disease mechanisms at the level of interconnected biological targets and pathways, particularly in multifactorial disorders such as osteoporosis [26,27]. Accumulating experimental evidence indicates that asiatic acid influences bone remodeling by attenuating osteoclast activity and modulating inflammatory and metabolic signaling, particularly through NF-κB- and PPARγ-associated mechanisms, leading to reduced bone loss in experimental osteoporosis models [18,28,29]. However, osteoporosis pathogenesis is driven by coordinated interactions among multiple signaling pathways rather than by isolated molecular events. Consistent with this concept, GO and KEGG enrichment analyses in the present study highlighted the concurrent involvement of inflammatory response-related processes, IL-6/STAT3 signaling, and PPAR signaling pathways, together with enrichment of Th17 cell differentiation and AGE–RAGE/PI3K–Akt-related pathways, suggesting functional crosstalk among inflammatory and metabolic networks that collectively regulate bone resorption and formation [7,9,10,30,31,32,33]. While these findings support the anti-osteoporotic potential of asiatic acid, they do not fully explain how these effects are coordinated at the network level or whether upstream regulatory nodes are involved. Network pharmacology therefore offers an appropriate strategy to examine the system-wide actions of asiatic acid and to identify key molecular hubs underlying its effects in osteoporosis.
Several key targets were identified through network pharmacology, including IL-6, STAT3, PPARG, NFKB1, PTGS2, ESR1, TLR4, HSP90AB1, PPARA, and RELA. These genes encode IL-6, STAT3, PPARy, NF-κB p105, COX-2, ESRα, TLR4, HSP90-β, PPARα, and NF-κB p65, respectively, which are critically involved in inflammation, hormone signaling, and bone remodeling. Among the identified targets, IL-6/STAT3 signaling axis plays a central role in skeletal biology. IL-6, especially via trans-signaling with soluble IL-6 receptor, activates gp130/JAK/STAT3, leading to increased RANKL in stromal/osteoblast cells and subsequent activation of nuclear factor of activated T-cells, cytoplasmic 1 (NFATc1) and osteoclast-specific genes in precursor cells. This axis is further amplified under estrogen-deficient conditions, linking it directly to postmenopausal bone loss [28,29,33]. In contrast, STAT3 signaling in osteoblasts and osteocytes supports load-induced bone formation, highlighting the importance of cellular context in determining its net skeletal effect [34]. Clinically, the relevance of this pathway is underscored by evidence that IL-6 blockade with tocilizumab lowers bone resorption and preserves bone mineral density in inflammatory conditions such as rheumatoid arthritis [35].
Additionally, PPARγ was identified as a key metabolic regulator associated with osteoporosis. Activation of PPARγ promotes adipogenic differentiation of bone marrow mesenchymal stem cells at the expense of osteoblastogenesis, increasing marrow fat and reducing bone formation [36]. Moreover, PPARγ facilitates osteoclastogenesis through PGC-1β-dependent activation of the NFATc1 pathway, boosting bone resorption. These molecular effects align with the clinical observations of increased fracture risk in patients treated with thiazolidinedione PPARγ agonists [37]. The NF-κB pathway also emerged as a critical signaling hub in osteoporosis. Activation of the p65/p50 heterodimer (where p50 is derived from the NFKB1 precursor p105) downstream of RANKL-RANK signaling induces NFATc1 and osteoclast-specific genes, driving osteoclast differentiation [9,38]. Inflammatory cytokines and estrogen deficiency further enhance NF-κB activity, while its activation in osteoblast-lineage cells suppresses bone formation, collectively contributing to net bone loss [39,40]. Consistent with these mechanisms, previous experimental studies have demonstrated that asiatic acid protects against osteoporosis mainly by inhibiting osteoclastogenesis through suppression of NF-κB signaling. In RANKL-stimulated osteoclast precursors, asiatic acid reduced TRAP^+^ osteoclast formation, bone resorption, and nuclear translocation of NF-κB p65, thereby downregulating NFATc1 and osteoclast-specific genes; consistent protection against bone loss was observed in OVX mouse models [18,28]. Asiatic acid also upregulates Smad7, further antagonizing TGF-β and NF-κB signaling to attenuate OVX-induced bone resorption [28]. In addition to antiresorptive effects, asiatic acid suppresses PPARγ expression and adipogenic markers in bone marrow mesenchymal stromal cells, shifting differentiation toward osteoblastogenesis [17]. Collectively, these findings establish NF-κB p65 and PPARγ as experimentally validated hub targets of asiatic acid in osteoporosis, whereas direct modulation of IL-6 and STAT3 has remained insufficiently characterized. Given its central hub position in the network analysis, its convergence across enriched inflammatory pathways, and its well-established upstream role in inflammation-driven osteoclastogenesis, IL-6 was prioritized for further molecular dynamics simulation to evaluate the stability and persistence of its interaction with asiatic acid under dynamic conditions.
Molecular docking analysis is widely employed in biological research and drug discovery to predict the binding affinity of ligands to receptor proteins [41]. In the present study, molecular docking analysis revealed that asiatic acid exhibited stronger binding affinities toward several osteoporosis-related proteins, including STAT3, PPARγ, NF-κB p105, COX-2, ESRα, TLR4, and NF-κB p65, compared to their native ligands. In contrast, for IL-6, the native ligand exhibited a slightly more negative predicted binding energy than asiatic acid, indicating a higher binding affinity. Nevertheless, asiatic acid was predicted to bind IL-6 through hydrogen bonding with ASN61, GLU172, LEU62, and LYS66, along with hydrophobic interaction involving ARG168. Notably, GLU172, LYS66, and ARG168 have been reported as key residues targeted by MD2-TLR4-IN-1, a potential IL-6 inhibitor that mitigates inflammation-driven bone loss in osteoporosis patients [42]. Furthermore, asiatic acid was predicted to interact with STAT3 through hydrophobic interaction with PRO336, a residue similarly targeted by natural STAT3 inhibitors [43]. Importantly, as NF-κB p65 plays a crucial in osteoclast differentiation [44], asiatic acid was predicted to bind directly to the DNA-binding subdomain of p65 through hydrogen bonds with ARG158 and HIS181, as well as a hydrophobic interaction with PRO182. Consistently, a curcuminoid derived from Curcuma longa has been shown to preferentially bind these same amino acid residues of NF-κB p65 in model of osteoclastogenesis [45]. These interactions suggest that binding of asiatic acid within the DNA-binding domain may interfere with the association of the p65-p50 heterodimer with nuclear DNA [45,46]. Nevertheless, molecular docking provides theoretical predictions of binding interactions, and experimental confirmation, such as biophysical assays, crystallographic studies, or site-directed mutagenesis, is needed to validate the precise interactions of asiatic acid with IL-6, STAT3, and NF-κB proteins.
Molecular dynamics simulation is a powerful computational technique widely used to evaluate the temporal stability, conformational flexibility, and interaction persistence of ligand–protein complexes in an explicit solvent environment [21,47]. To complement molecular docking results and provide a more realistic insight into the interaction between asiatic acid and IL-6 under near-physiological conditions, a 200 ns MD simulation was conducted. Although molecular docking revealed that asiatic acid exhibited a strong binding affinity for IL-6 (−8.64 kcal/mol), molecular dynamics simulation further demonstrated the stability and durability of this interaction over time. Notably, asiatic acid was predicted to bind with amino acid residues that overlap with those targeted by a known IL-6 inhibitor [42], reinforcing its potential as an IL-6 antagonist. Throughout the simulation, the asiatic acid-IL-6 complex maintained a stable RMSD trajectory, remaining below 3 Å for most of the simulation period and exhibiting only minor fluctuations between 170–200 ns. These observations indicate that the ligand maintained a consistent and well-anchored binding conformation throughout the simulation period. The analysis also revealed that asiatic acid formed persistent interactions with ASN61, LEU62, and LYS66 located in the AB-loop, and ARG168 in helix D of the IL-6 structure [48,49]. These structural motifs are known to constitute Site I, the primary binding interface between IL-6 and its receptor IL-6Rα [49,50]. The AB-loop and helix D are essential for IL-6 receptor recognition and downstream signaling [48,51]. Thus, asiatic acid’s engagement with these critical residues suggests that it may competitively interfere with IL-6–IL-6Rα complex formation, blocking receptor-mediated signal transduction and offering a plausible anti-inflammatory mechanism [52]. Consistent with this interpretation, similar residue hotspots have emerged in other IL-6 inhibitor studies, as shown in molecular docking of dexamethasone, which showed binding to ASN60/ASN61, while thymoquinone, a natural anti-inflammatory compound, was shown to target ARG168, overlapping directly with the asiatic acid interaction site [52]. To further assess the relevance of asiatic acid’s interaction profile, MD simulation was also performed for MD2-TLR4-IN-1, a known IL-6 inhibitor. Interestingly, both ligands interacted with common anchoring residues, ASN61, LEU62, LYS66, and ARG168, but asiatic acid displayed slightly greater flexibility while maintaining persistent binding. This suggests that asiatic acid may mimic the native ligand’s pharmacophore while offering additional adaptability within the binding pocket.
Collectively, these findings indicate a common pharmacophoric pattern among small-molecule IL-6 inhibitors, wherein effective compounds anchor into the receptor-binding groove, stabilize local protein conformations, and reduce flexibility in the AB-loop and helix D regions through hydrogen bonding and hydrophobic interactions. This study, integrating network pharmacology, molecular docking, and molecular dynamics simulation, provides compelling mechanistic insight into asiatic acid’s potential as an IL-6 modulator. The identification of IL-6 as a potential upstream target of asiatic acid has important translational implications. Given that IL-6 blockade reduces bone resorption in inflammatory diseases, asiatic acid may represent a natural, multi-target adjunct candidate for inflammation-driven osteoporosis, particularly in postmenopausal or chronic inflammatory settings [53]. Unlike conventional antiresorptive agents that primarily target osteoclast activity, asiatic acid appears to modulate upstream cytokine signaling, metabolic regulation, and transcriptional networks simultaneously, which may theoretically reduce compensatory pathway activation.
These findings provide a strong foundation for future drug development targeting inflammation-driven bone loss. In recent years, integrative in silico approaches have been increasingly recognized as valuable tools in natural product-based drug discovery, particularly for elucidating multi-target mechanisms and prioritizing candidates for experimental validation [54,55,56]. In this context, the present study employed an integrative in silico framework to investigate the potential anti-osteoporotic effects of asiatic acid. Nevertheless, such computational analyses are inherently hypothesis-generating in nature and require experimental validation to confirm their biological relevance. Therefore, the IL-6-specific anti-osteoporotic effects of asiatic acid remain to be validated in both in vitro and in vivo models. In vitro validation represents a critical next step, particularly using osteoblast and osteoclast cell models to directly assess the effects of asiatic acid on IL-6–mediated signaling pathways, including STAT3 and NF-κB, as well as on functional endpoints related to bone resorption and formation. Despite the promising in silico results, it is important to acknowledge that favorable binding affinity does not necessarily translate to in vivo efficacy. Asiatic acid, as a pentacyclic triterpenoid, may exhibit limited oral bioavailability and tissue distribution [53]. Accordingly, optimization strategies, including formulation enhancement or structural modification, may be required to achieve therapeutically relevant concentrations in bone tissue. In vivo studies employing ovariectomized or inflammation-induced osteoporosis models will be essential to confirm efficacy and safety. Additionally, biophysical approaches such as surface plasmon resonance or isothermal titration calorimetry could directly verify IL-6 binding, while transcriptomic or phosphoproteomic analyses may further clarify pathway-level effects [57,58].
5. Conclusions
This study highlights the potential anti-osteoporotic role of asiatic acid through an integrated in silico approach combining network pharmacology, molecular docking, and molecular dynamics simulation. Network pharmacology identified key osteoporosis-related targets involved in inflammatory signaling, hormone regulation, and bone remodeling, with key hubs including IL-6, STAT3, NF-κB, and PPARγ. Molecular docking revealed favorable binding affinities between asiatic acid and these targets, particularly IL-6. Molecular dynamics simulation further confirmed the structural stability of the asiatic acid-IL-6 complex, demonstrating persistent protein–ligand interactions throughout the simulation period. Collectively, these findings suggest that asiatic acid may exert osteoprotective effects via multi-target regulation of inflammation-driven bone loss, extending its known downstream actions on NF-κB and PPARγ signaling to potential upstream regulation of IL-6-mediated pathways. By providing system-level and structural evidence supporting IL-6 as a plausible molecular target of asiatic acid, this study advances the mechanistic understanding of its anti-osteoporotic potential. Although experimental validation is warranted, the present work establishes a robust mechanistic framework for future in vitro, in vivo, and translational studies and supports further exploration of asiatic acid as a natural multi-target therapeutic candidate for the prevention and management of osteoporosis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Rachner T.D. Khosla S. Hofbauer L.C. Osteoporosis: Now and the future Lancet 20113771276128710.1016/S 0140-6736(10)62349-521450337 PMC 3555696 · doi ↗ · pubmed ↗
- 2Compston J.E. Mc Clung M.R. Leslie W.D. Osteoporosis Lancet 201939336437610.1016/S 0140-6736(18)32112-330696576 · doi ↗ · pubmed ↗
- 3Hsu S.-H. Chen L.-R. Chen K.-H. Primary Osteoporosis induced by androgen and estrogen deficiency: The molecular and cellular perspective on pathophysiological mechanisms and treatments Int. J. Mol. Sci.2024251213910.3390/ijms 25221213939596206 PMC 11593909 · doi ↗ · pubmed ↗
- 4Ginaldi L. Di Benedetto M.C. De Martinis M. Osteoporosis, inflammation and ageing Immun. Ageing 200521410.1186/1742-4933-2-1416271143 PMC 1308846 · doi ↗ · pubmed ↗
- 5Zhu X. Du L. Zhang L. Ding L. Xu W. Lin X. The critical role of toll-like receptor 4 in bone remodeling of osteoporosis: From inflammation recognition to immunity Front. Immunol.202415133308610.3389/fimmu.2024.133308638504994 PMC 10948547 · doi ↗ · pubmed ↗
- 6Castellani C. De Martino E. Scapato P. Osteoporosis: Focus on bone remodeling and disease types Bio Chem 202553110.3390/biochem 5030031 · doi ↗
- 7Xu J. Yu L. Liu F. Wan L. Deng Z. The effect of cytokines on osteoblasts and osteoclasts in bone remodeling in osteoporosis: A review Front. Immunol.202314122212910.3389/fimmu.2023.122212937475866 PMC 10355373 · doi ↗ · pubmed ↗
- 8Wu Q. Zhou X. Huang D. Ji Y. Kang F. IL-6 enhances osteocyte-mediated osteoclastogenesis by promoting JAK 2 and RANKL activity in vitro Cell. Physiol. Biochem.2017411360136910.1159/00046545528278513 · doi ↗ · pubmed ↗