Imaging Cell Competition in Ex-Vivo Drosophila Adult Brains
Andrés Gutiérrez-García, Mariana Marques-Reis, Eduardo Moreno

TL;DR
This study introduces a new method for long-term imaging of adult fruit fly brains to observe cell competition after injury.
Contribution
A novel ex vivo protocol for extended live imaging of Drosophila adult brains without complex culture conditions.
Findings
The ex vivo protocol allows stable imaging of adult Drosophila brains for up to 32 hours.
Cell competition in optic lobes peaks at 48 hours after traumatic brain injury.
The method successfully visualizes Azot expression in loser cells during elimination.
Abstract
Live imaging has been instrumental in understanding cellular dynamics in Drosophila tissues, but technical limitations have prevented the long-term visualization of cell competition in adult brains. Here, we describe a simple ex vivo protocol that enables extended live imaging of adult Drosophila brains for up to 32 h. The method relies on non-supplemented Schneider’s Drosophila medium and hydrophobic interactions to maintain brain stability during imaging, eliminating the need for complex culture conditions or embedding procedures. We validate this approach by studying cell competition in the optic lobes following traumatic brain injury, where cell competition is expected to occur with a peak at 48 h after damage. We demonstrate the value of this method by visualizing the expression of the fitness checkpoint Azot in a loser cell and its subsequent elimination. This protocol offers a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
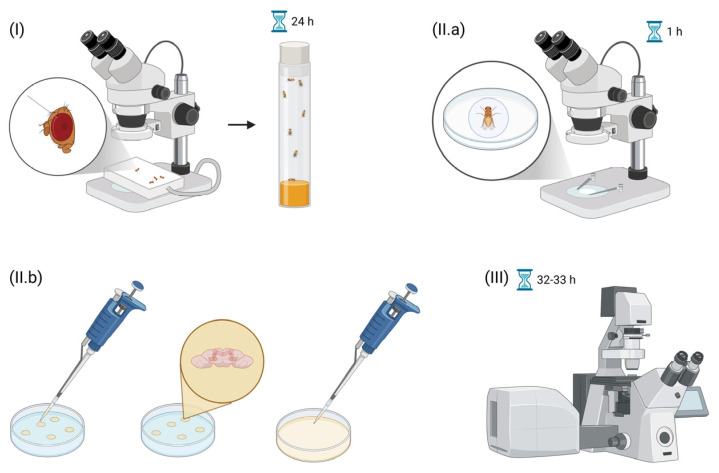 Figure 1
Figure 1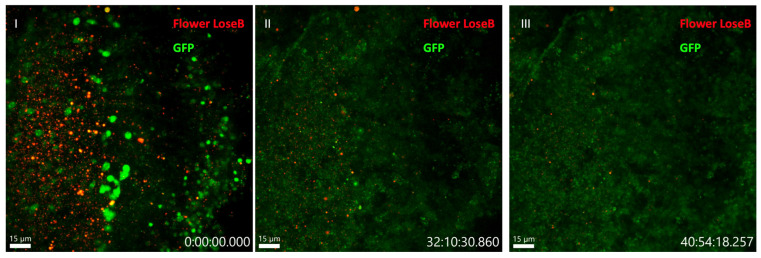 Figure 2
Figure 2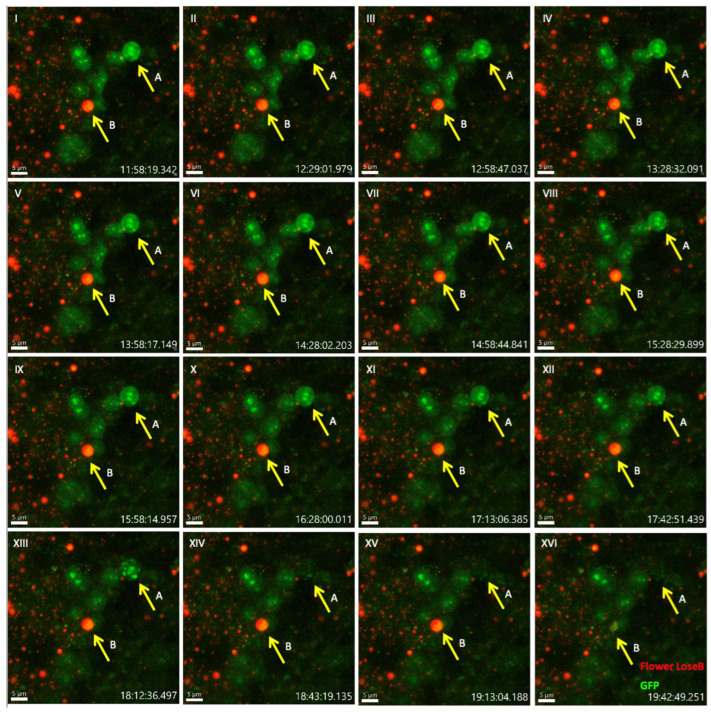 Figure 3
Figure 3 Figure 4
Figure 4- —FCT—Fundação para a Ciência e a Tecnologia—PhD studentship
- —Portuguese national funds
- —European Research Council
- —CONGENTO
- —FCT (Portugal)
- —Lisboa2020
- —Portuguese Platform of BioImaging
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHippo pathway signaling and YAP/TAZ · Neurobiology and Insect Physiology Research · Developmental Biology and Gene Regulation
1. Introduction
The Darwinian principle of ‘survival of the fittest’ extends beyond macroscopic organisms to the cellular level through a phenomenon known as Cell Competition. This process, first described in 1975 in Drosophila wing imaginal disks, demonstrated that wild-type cells could outcompete slow-growing cells carrying mutations in minute genes [1]. Since then, the concept has evolved to encompass the elimination of viable but suboptimal cells when they encounter cells with superior fitness within the same tissue compartment. This fundamental biological process is conserved from Drosophila to mammals, including humans [2,3]. Cell competition can be triggered by various factors, including competition for survival factors [4,5], mechanical forces [6,7], and differential expression of fitness markers [8,9,10,11].
Among these fitness markers, the Flower protein family plays a crucial role in cellular fitness communication. Conserved across Drosophila, mice, and humans, the Flower protein in Drosophila exists in three isoforms: Flower ubi, present in cells with higher fitness status (‘winners’), and Flower Lose A and Flower Lose B, present in cells with lower fitness status (‘losers’) [8,12,13]. Notably, cell elimination occurs only in heterogeneous populations where cells of different fitness levels interact [8,14]. In such contexts, loser cells activate the fitness checkpoint protein Azot, which triggers the pro-apoptotic gene hid, ultimately leading to programmed cell death [15].
While previous studies have demonstrated the co-expression of Flower Lose B and Azot in fixed tissue samples, particularly in the adult brain, the temporal dynamics of this process remain poorly understood [16]. Specifically, the time course from initial Flower Lose B expression to Azot activation and subsequent cell elimination requires further investigation. Ex vivo live imaging studies of cell competition in Drosophila have been primarily confined to larval imaginal disks [17,18], while other ex vivo imaging approaches have been established for larval and pupal brains in different contexts, typically for periods less than 18 h [19,20]. However, competitive events require longer observation windows, as the comparison of fitness status, the decision-making process for cell elimination, and the execution of cell death are multi-step processes. Current in vivo techniques using imaging windows allow visualization of the midgut and certain regions of the adult brain, but these methods are optimized for light-sheet microscopy or two-photon microscopy rather than confocal imaging [21,22,23,24]. For adult brains, confocal microscopy can offer superior spatial resolution and more straightforward multicolor imaging workflows than traditional light-sheet microscopy, while avoiding the complexity of two-photon systems [25]. Recent efforts have been made to study ex vivo Drosophila adult brains with confocal microscopy, but they still require complex pumping systems that might be unnecessary if no pharmacological manipulation is needed [26].
To address these limitations, we have developed a simplified ex vivo imaging approach that enables the long-term observation (32 h) of adult Drosophila brains. Our system utilizes a non-supplemented Schneider’s Drosophila Medium and leverages hydrophobic interactions between the brain tissue and the imaging chamber to maintain specimen stability. To induce competitive events, we performed traumatic brain injury (TBI) in the right optic lobe, as this injury triggers an initial wave of apoptosis due to mechanical damage, followed by a second wave of cell elimination peaking at 48 h post-injury that is driven by flower-dependent cell competition [27,28]. This methodology will not only facilitate the study of cell competition dynamics but also provide a versatile platform for extended ex vivo imaging of the adult Drosophila brain.
2. Experimental Design
In this protocol, we describe the experimental setup and procedures for live imaging of ex vivo adult Drosophila melanogaster brains following TBI, enabling the observation of cell competition events through fitness fingerprint comparisons (Figure 1). Although developed for studies of cell competition and tissue remodeling after TBI, the sample preparation and imaging procedures described here can be broadly applied to any ex vivo live imaging study of adult Drosophila brains.
2.1. Reagents
Schneider’s Drosophila Medium (Biowest, Nuaillé, France; Cat. No.: L0207-500).
2.2. Materials
2.PIPETMAN P1000, 100–1000 µL, Metal Ejector (Gilson, Middleton, WI, USA; Cat. No.: F144059M).3.PIPETMAN P200, 20–200 µL, Metal Ejector (Gilson, Middleton, WI, USA; Cat. No.: F144058M).4.Pipette Tips SureOne™ Micropoint, Universal Fit, Non-Filtered 200 µL, 1000 µL (Thermo Scientific, Waltham, MA, USA; Cat. No.: 02-707-410; 02-707-407).5.Minutien Pins (Fine Science Tools, Heidelberg, Germany, Cat. No.: 26002-10).6.Petrolatum (Unilever, NJ, USA; Cat. No.: 305212352009).7.Sterilin™ 30 to 140 mm Petri Dishes (Thermo Scientific, Waltham, MA, USA; Cat. No.: PF55).8.Fine Forceps (Fine Science Tools, Heidelberg, Germany, Cat. No.: 11403-11).9.Nunc™ Glass Bottom Dishes (Thermo Scientific, Waltham, MA, USA; Cat. No.: 150682).
2.3. Drosophila Melanogaster Lines
10.flower{KO; KI-flowerLoseB::mCherry} [8].11.azot{KO; KI-LexA::p65} [16].12.26xLexAop-CD8::GFP (Bloomington Drosophila Stock Center, Bloomington, IN, USA; Cat. No.: 32207).
2.4. Equipment
Flystuff^®^ Drosophila Anesthesia Systems (Genesee Scientific, El Cajon, CA, USA; Cat. No.: 59-121C).Zeiss Stemi 305 Stereo zoom microscope (ZEISS Microscopy, Oberkochen, Germany, Cat. No.: 435063-9020-100).Zeiss LSM 880 Airyscan Inverted Confocal Microscope with stage incubator (ZEISS Microscopy, Oberkochen, Germany).Objective Plan-Apochromat 40x/1.4 Oil DIC M27 (ZEISS Microscopy, Oberkochen, Germany, Cat. No.: 420762-9900-000).
3. Procedure
3.1. Drosophila Husbandry and Preparation for Experiments (Duration: Minimum of 13 Days)
CRITICAL STEP: Start by preparing the fly stocks needed for the experiment. In our experimental conditions, we first developed the stocks w ; azot{KO; KI-LexA::p65}/CyO, 26xLexAop-CD8::GFP/Tm6B and w; ; flower{KO; KI-flowerLoseB::mCherry}/Tm6B from the lines mentioned in Section 2.3.
Cross 3 males of the genotype w; azot{KO; KI-LexA::p65}/CyO, 26xLexAop-CD8::GFP/Tm6B with 10 female virgins of the genotype w; ; flower{KO; KI-flowerLoseB::mCherry}/Tm6B. Keep the cross in wide plastic vials with Vienna standard media [29] in a chamber with a controlled temperature of 25 °C and humidity of 70%.Discard the adults after 6 days.OPTIONAL STEP: These adults can be put in a new vial to establish a new cross in case it is needed for another experiment.The F1 generation should hatch 10 days after the cross. Collect the recently hatched flies into a new vial. CRITICAL STEP: Wait 2 days to perform the TBI on these flies.Use a minutien pin to stab the young adult flies (2 days old) in the right optic lobe, through the cuticle on the dorsal part of the head (as described in [27]), next to the second pair of bristles that surround the retina. CRITICAL STEP: Wait 24 h before starting the dissections.
3.2. Drosophila Adult Brain Dissection (Duration: 1 h)
We followed the principles stated in [30] with small modifications.
CRITICAL STEP: Before the dissections, place the Schneider’s Drosophila Medium in a 4 °C chamber.
Prepare 15 mL of chilled Schneider’s Drosophila Medium and place it on ice.Coat a Petri dish with a thin layer of Petrolatum.Anesthetize the flies with CO_2_ and, with the forceps, place them belly up in the Petri dish with the wings embedded in *Petrolatum.*Fill the Petri dish with chilled Schneider’s Drosophila Medium.With the forceps, detach the proboscis and grab from below the cavity with each forcep, break the head cuticle, and expose the brain. CRITICAL STEP: At this point, the brain is still attached to the rest of the body.Clean the brain from the surrounding trachea without damaging its structure.Repeat steps 1–6 until the desired number of brains is obtained (in our case, we dissected 5 brains).In a Nunc™ Glass Bottom Dish, place 40 μL droplets of chilled Schneider’s Drosophila Medium.Use the forceps to detach the brain from the body, perform final cleaning if necessary, and transfer each brain to a droplet in the Nunc™ Glass Bottom Dish.Place each brain at the bottom of each droplet, facing down. CRITICAL STEP: The brain–glass adhesion relies upon hydrophobic contact with the glass. The hydrophobic contact is very fragile, so it is advisable to dissect more than one brain.Gently fill the Nunc™ Glass Bottom Dish with 5 mL of chilled Schneider’s Drosophila Medium.
Troubleshooting:
The droplets provide mechanical stability to the brain, reducing susceptibility to turbulence and preventing detachment. We observed that embedding the brains in droplets before filling the Nunc™ Glass Bottom Dish with 5 mL of chilled Schneider’s Drosophila Medium visibly decreased the likelihood of tissue detachment compared to adding the brains after the dish had been filled.
3.3. Imaging (Duration: 32 to 33 h)
CRITICAL STEP: Set the stage-top incubator to 25 °C
- Follow the instructions for imaging according to your laboratory’s system. In our case, we used the Zeiss LSM 880 Airyscan confocal microscope, and the images were taken with the objective Plan-Apochromat 40x/1.4 Oil DIC M27 and a frame size of 2048 × 2048. The “GFP” and “Flower LoseB” channels refer to the specific ranges of wavelengths chosen for redirection to the detector. For the “GFP” channel, we selected the range from 499 nm to 548 nm, and for the “Flower LoseB” channel, we selected the range from 565 nm to 644 nm. The depth of the Z-stack was 40 μm, the step between frames was 1 μm, and each frame of the video corresponded to 15 min for a total duration of 32 h from the beginning of the imaging session.
-
Troubleshooting:
-
1.We imaged one brain per session to minimize the Nunc™ Glass Bottom Dish movement and prevent tissue detachment. To ensure the robustness of the protocol, we repeated the imaging session three independent times.
4. Expected Results and Discussion
The ex vivo system successfully maintained brain tissue viability for 32 h, as evidenced by the persistent fluorescent signal throughout the imaging period (Figure 2). From 32 to 40 h, we ceased to see competitive events. To analyze cell competition dynamics, we focused on a temporal window between 36 and 42 h post-injury (corresponding to 12 to 20 h since the beginning of the imaging session), when most competitive interactions are expected to occur [28] (Figure 3, Supplementary Video S1).
To visualize these competitive events, we employed a dual-labeling system: Flower LoseB was tagged with mCherry, while Azot expression was monitored using the bipartite LexA-LexAop system driving GFP expression [16]. Our time-lapse imaging revealed two cellular behaviors. In the initial frame (Figure 3I), we identified two cells of interest: cell A expressing only Azot (GFP-positive) and cell B co-expressing both Flower LoseB (mCherry-positive) and Azot (GFP-positive). Between frames (XIII) and (XIV), cell A exhibited a morphological disruption and disappearance, and between frames (XV) and (XVI), one can observe the morphological disruption and disappearance of cell B. These disappearances suggest apoptotic events, as Azot precedes hid activation and consequent cell apoptosis [15], aligned with the dynamics of Flower-dependent cell competition [15,16]. However, definitive confirmation of apoptosis would require additional cell death markers not included in our current setup.
This ex vivo imaging approach complements existing in vitro competition assays by enabling the direct visualization of competitive cell behaviors in intact brain tissue [13]. The straightforward nature of our protocol, requiring only standard laboratory equipment and commercially available Drosophila culture medium, makes it readily adaptable for any Drosophila research laboratory interested in studying cellular dynamics in the adult brain.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Morata G. Ripoll P. Minutes: Mutants of Drosophila Autonomously Affecting Cell Division Rate Dev. Biol.19754221122110.1016/0012-1606(75)90330-91116643 · doi ↗ · pubmed ↗
- 2Morata G. Cell Competition: A Historical Perspective Dev. Biol.2021476334010.1016/j.ydbio.2021.02.01233775694 · doi ↗ · pubmed ↗
- 3Cong B. Cagan R.L. Cell Competition and Cancer from Drosophila to Mammals Oncogenesis 202413110.1038/s 41389-023-00505-y 38172609 PMC 10764339 · doi ↗ · pubmed ↗
- 4Moreno E. Basler K. Morata G. Cells Compete for Decapentaplegic Survival Factor to Prevent Apoptosis in Drosophila Wing Development Nature 200241675575910.1038/416755 a 11961558 · doi ↗ · pubmed ↗
- 5Martins V.C. Busch K. Juraeva D. Blum C. Ludwig C. Rasche V. Lasitschka F. Mastitsky S.E. Brors B. Hielscher T. Cell Competition Is a Tumour Suppressor Mechanism in the Thymus Nature 201450946547010.1038/nature 1331724828041 · doi ↗ · pubmed ↗
- 6Brás-Pereira C. Moreno E. Mechanical Cell Competition Curr. Opin. Cell Biol.201851152110.1016/j.ceb.2017.10.00329153702 · doi ↗ · pubmed ↗
- 7Wagstaff L. Goschorska M. Kozyrska K. Duclos G. Kucinski I. Chessel A. Hampton-O’Neil L. Bradshaw C.R. Allen G.E. Rawlins E.L. Mechanical Cell Competition Kills Cells via Induction of Lethal P 53 Levels Nat. Commun.201671137310.1038/ncomms 1137327109213 PMC 4848481 · doi ↗ · pubmed ↗
- 8Rhiner C. López-Gay J.M. Soldini D. Casas-Tinto S. Martín F.A. Lombardía L. Moreno E. Flower Forms an Extracellular Code That Reveals the Fitness of a Cell to Its Neighbors in Drosophila Dev. Cell 20101898599810.1016/j.devcel.2010.05.01020627080 · doi ↗ · pubmed ↗