Improved Sphincter Muscle Regeneration by Myoblasts from M. extensor carpi radialis in a Large Animal Model of Urinary Incontinence
Niklas Harland, Lukas Schwarz, Meltem Avci-Adali, Andrea Buzanich-Ladinig, Lina M. Serna-Higuita, Arnulf Stenzl, Wilhelm K. Aicher

TL;DR
This study shows that myoblasts from a specific muscle improve sphincter function in a large animal model of urinary incontinence.
Contribution
Myoblasts from M. extensor carpi radialis outperformed those from M. semitendinosus in restoring sphincter function.
Findings
Myoblasts from M. extensor carpi radialis showed high expression of myogenic markers and formed myotubes in vitro.
These myoblasts restored 92% of sphincter function compared to baseline levels in the study model.
The experimental group showed significant recovery compared to mock controls (p < 0.045).
Abstract
Purpose: Stress urinary incontinence (SUI) is a significant medical challenge affecting substantial parts of modern societies. Several studies suggested that cell therapy may alleviate the symptoms. However, in many cases, the overall efficacy was not satisfactory for the patient’s needs. Moreover, in our recent preclinical studies, myoblasts isolated from M. semitendinosus failed to restore significant urethral sphincter function. We, therefore, investigated in our large animal SUI model whether myoblasts from other muscles yielded better sphincter recovery. Methods: Urethral sphincter deficiency was induced surgically in six female littermates and confirmed by measuring the urethral wall pressure. Three days after induction of sphincter deficiency in gilts, homologous myoblasts were injected into the sphincter complex. The urethral wall pressure and urine status were monitored weekly…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
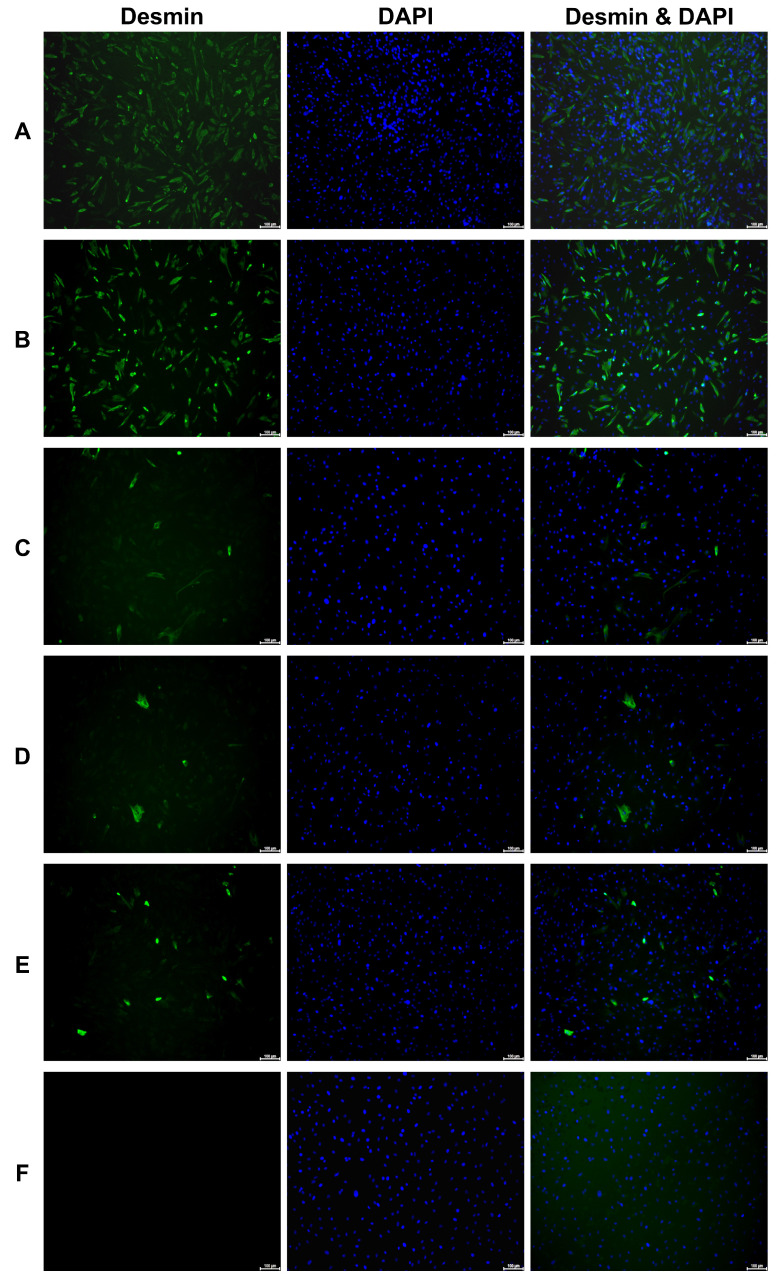 Figure 1
Figure 1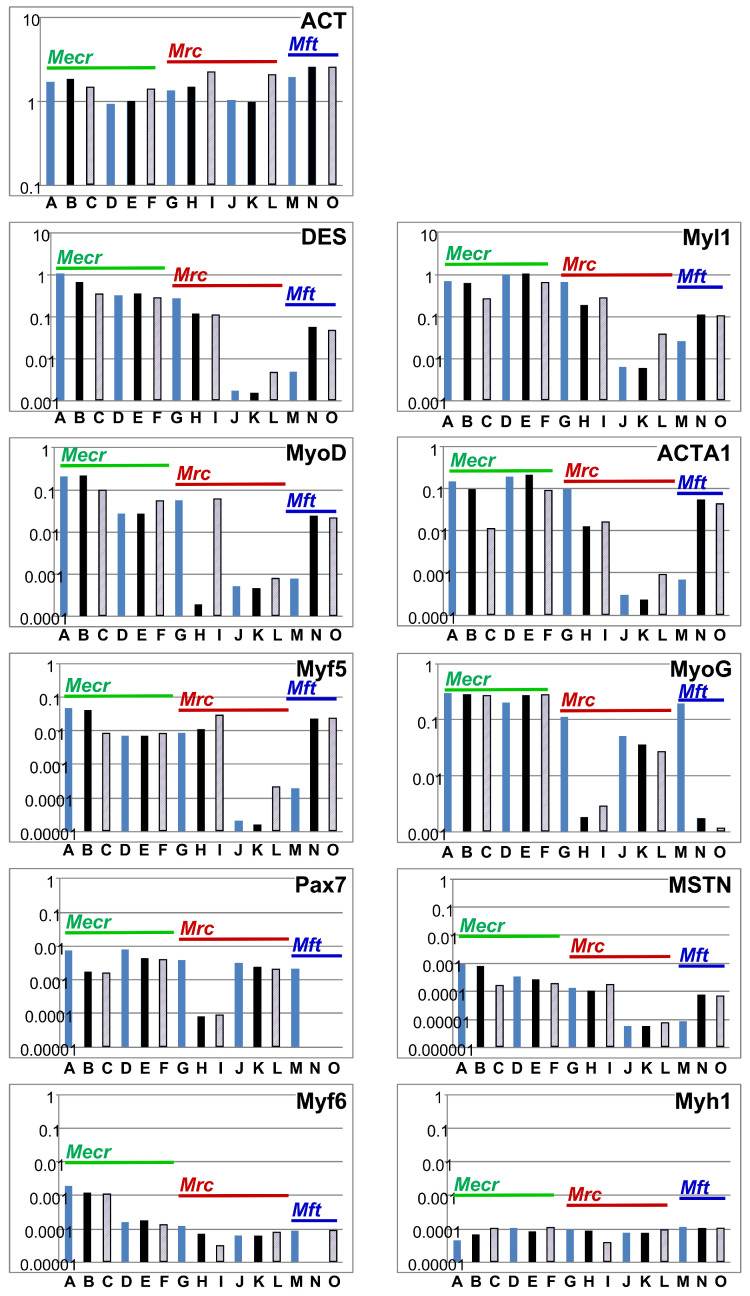 Figure 2
Figure 2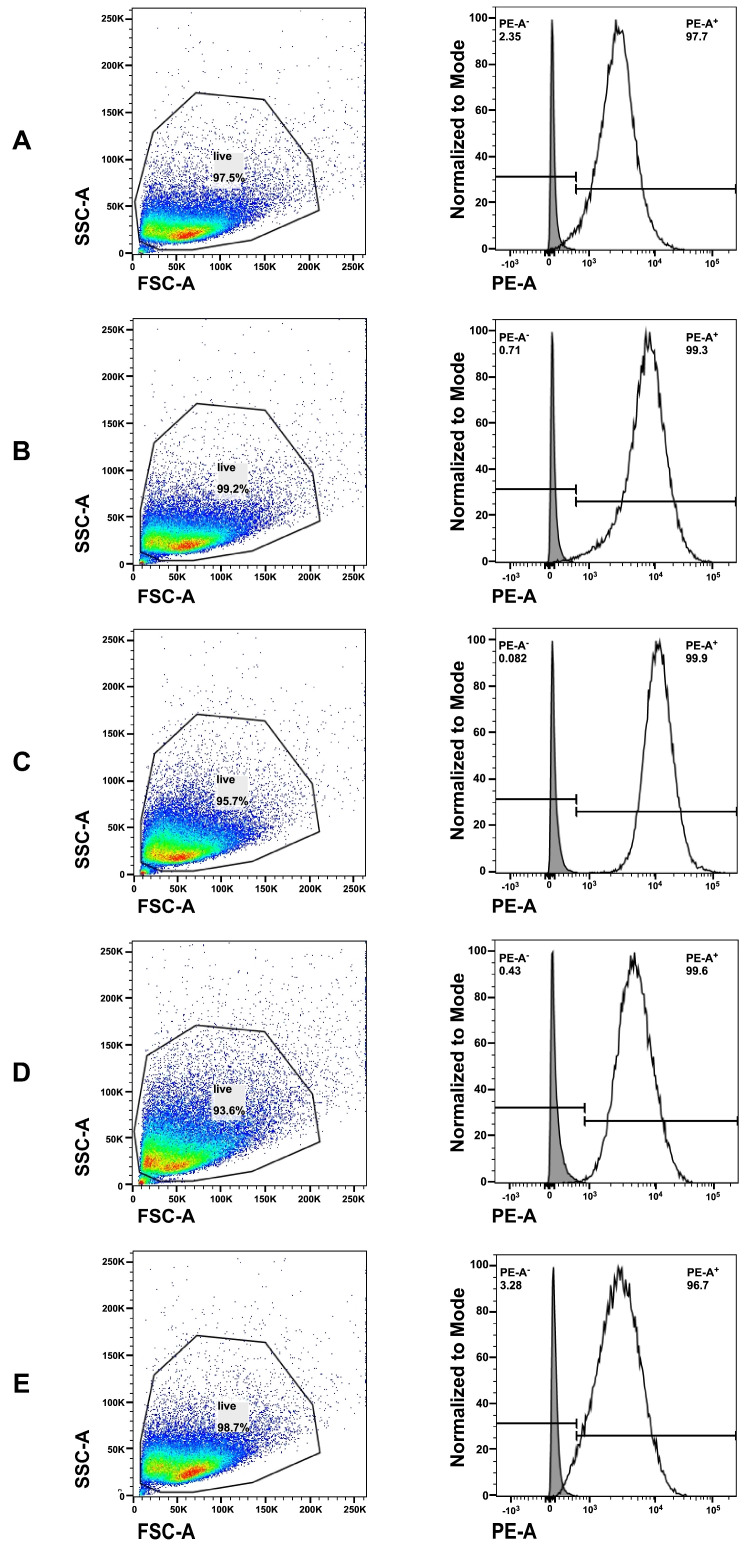 Figure 3
Figure 3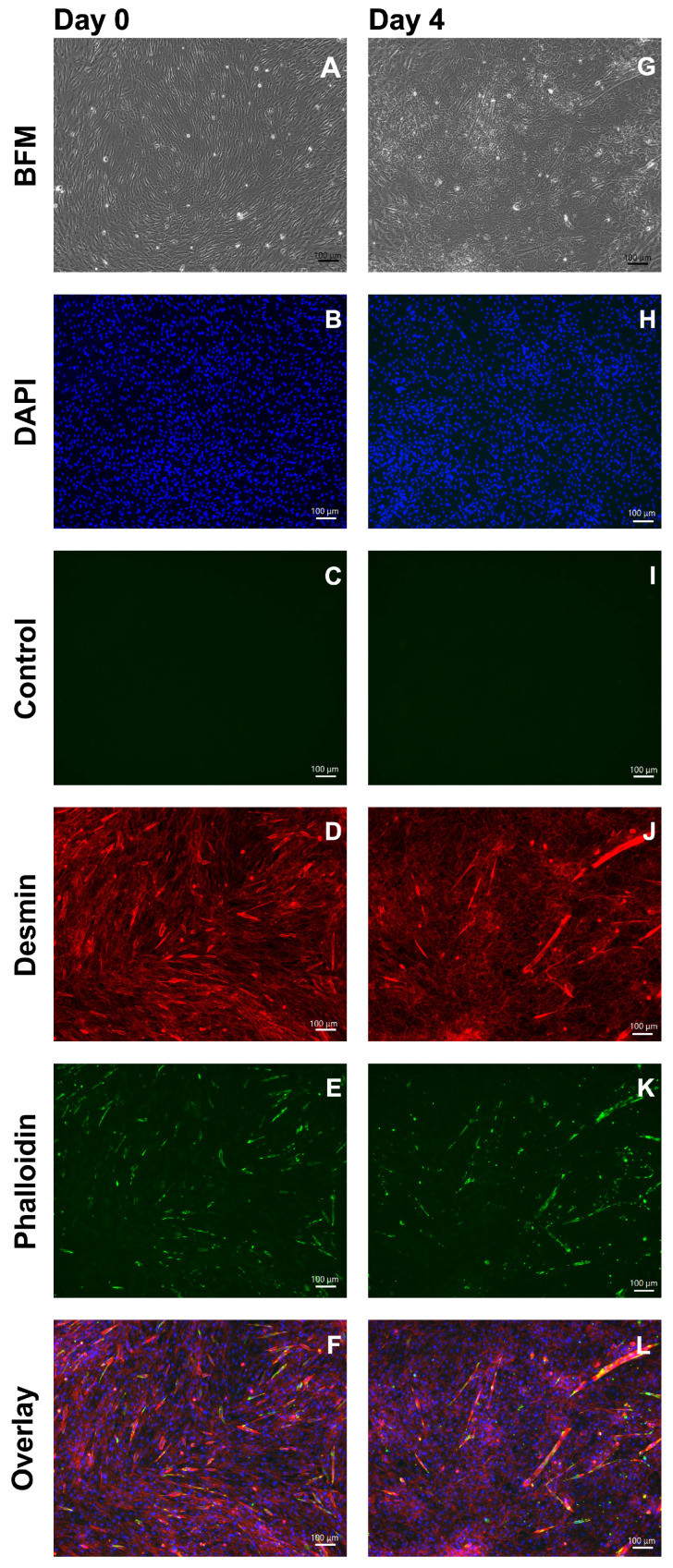 Figure 4
Figure 4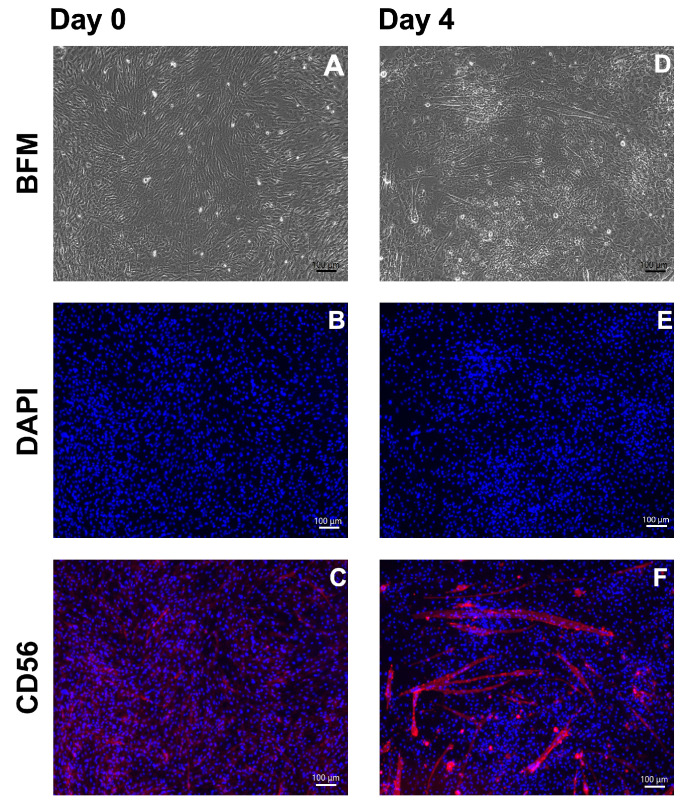 Figure 5
Figure 5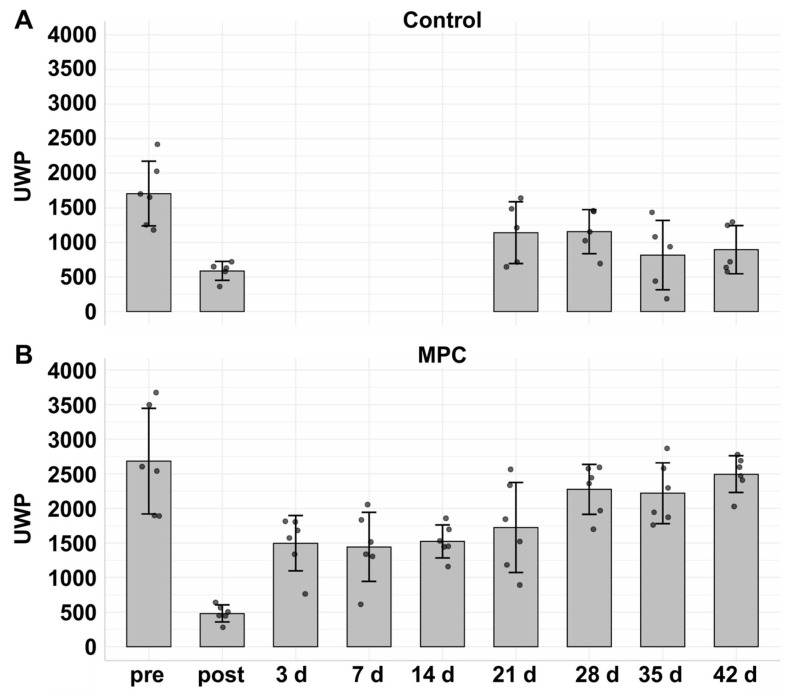 Figure 6
Figure 6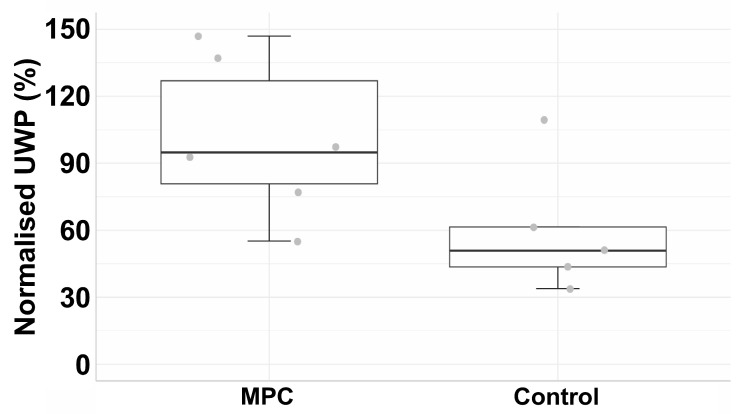 Figure 7
Figure 7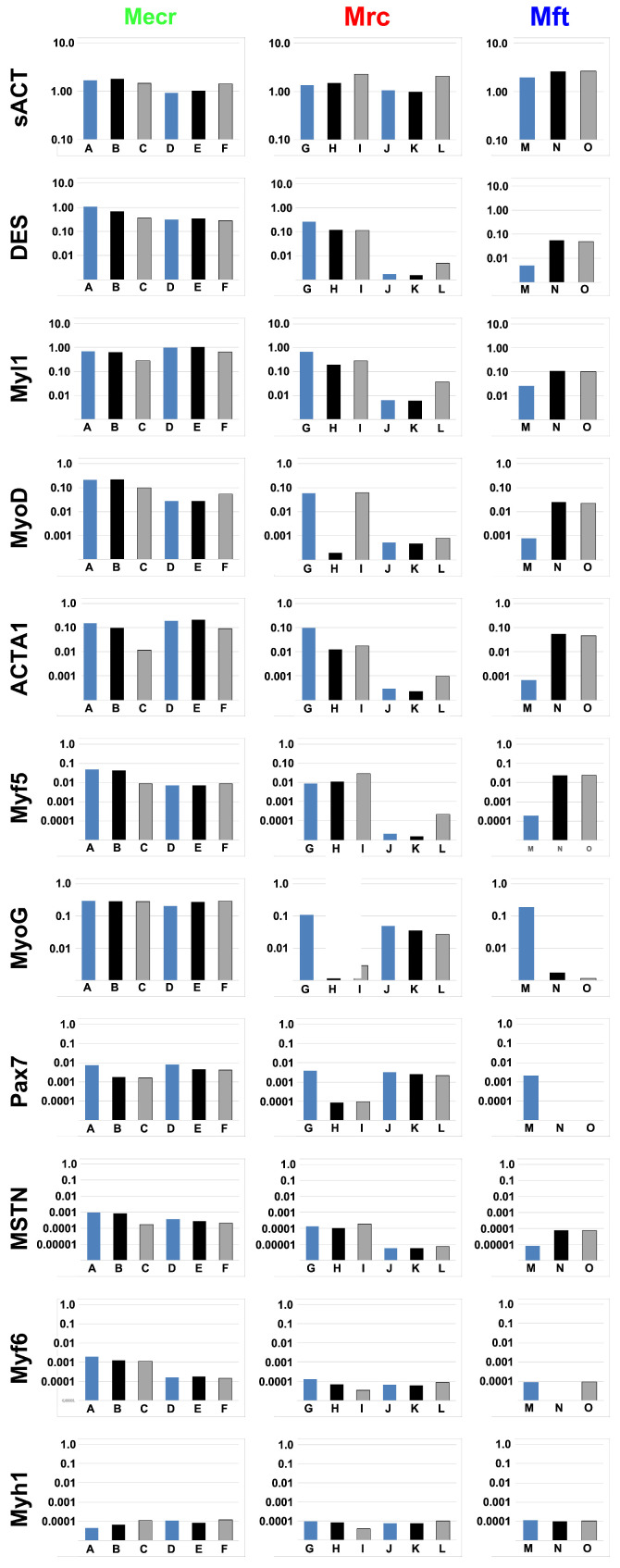 Figure 8
Figure 8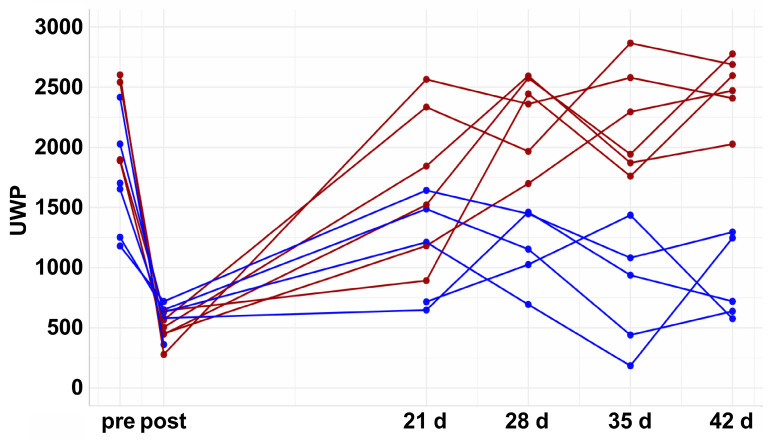 Figure 9
Figure 9- —VetMed University in Vienna
- —University of Tuebingen
- —Adolf-Leuze-Stiftung
- —Förderverein Urologie e.V.
- —Open Access Publication Fund of the University of Tübingen
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPelvic floor disorders treatments · Tissue Engineering and Regenerative Medicine · Urinary Bladder and Prostate Research
1. Introduction
Stress urinary incontinence (SUI) is an increasing challenge for many modern societies as its incidence rises among the elderly in ageing populations [1,2,3,4,5]. Medical records and patient surveys suggest that about 37% of elderly women suffer from any form of urinary incontinence (UI) [2,6], while the prevalence in men can reach up to 30% [5]. Incontinence is linked with various secondary conditions, including social isolation, recurrent urinary tract infections, and dermatitis [7,8,9]. Individuals experiencing mild UI may manage the condition by adjusting fluid intake and output. Physical exercise and medication can help alleviate the symptoms [7,10]. In severe cases of SUI, surgical interventions may be necessary to achieve satisfactory results [7].
SUI was linked to a loss of functional muscle tissue, urethral scar formation, and impaired innervation [7,11]. This has led to the idea that improving sphincter muscle function could be achieved by locally applying active components that promote tissue regeneration, including adipose tissue-derived mesenchymal stromal cells (ADSCs) and myogenic progenitor cells (MPCs) [12]. Several preclinical studies have indicated that cell therapies might improve incontinence [13]. Similarly, some clinical studies have reported success [14,15,16,17,18]. Others, however, failed to show convincing results [19]. The double-blind ADMC-USR study reported placebo responses [20]. Nevertheless, cell-based SUI therapies demonstrated high cost-effectiveness [21]. However, the current evidence and situation remain unsatisfactory for the field.
The partly contradictory reports may be due to technical differences between the studies. One challenge is the application of active components in the region of interest [22,23,24]. The quality of injected cells may also influence the outcomes of SUI therapies. In our recent study, ADSCs demonstrated a significant recovery in urethral wall pressure in a porcine large animal model of SUI cell therapy [25]. In contrast, MPCs derived from M. semitendinosus did not produce a full and significant recovery [25]. Furthermore, in clinical studies of SUI cell therapy, myoblasts from various muscles were used, likely following different protocols [11,14,17,18,26,27], and most of these studies did not characterise the myoblasts in detail. In this context, it gains importance that different porcine muscles contain quite distinct numbers of myogenic progenitor cells [28]. Thus, we hypothesised that the number and quality of the MPCs applied for sphincter regeneration might play a role in the regeneration of the deficient urinary closure apparatus in our model. Therefore, we expanded our research to investigate, in an exploratory study, whether MPCs isolated from other striated muscles might offer a better outcome in SUI cell therapy within this model.
2. Materials and Methods
2.1. Production of Myogenic Progenitor Cells
A 5-day-old male landrace littermate of the gilts to be employed in the SUI cell therapy study was anaesthetised (10 mg/kg body weight (BW) ketamine, 0.5 mg/kg BW azaperone i.v.) and sacrificed. The death of the boar was confirmed via auscultation of absent heartbeat noises and testing the corneal reflex. Based on the amounts of satellite cells and MPCs in different muscles [28], tissue samples of M. extensor carpi radialis (Mecr), M. rhomboideus cervicis (Mrc), and M. fibularis tertius (Mft) were isolated aseptically. The MPCs were prepared as described [28,29]. The tissue samples were minced and subjected to proteolytic digestion (HBSS, 0.2% collagenase 1, 0.01% DNase 1, 0.025% trypsin). The cells were enriched by a cell strainer, centrifuged (800× g, 10 min. 4 °C), purified by a Percoll gradient (15,000× g, 10 min; Sigma-Aldrich, Taufkirchen, Germany), washed, and expanded (=bath 1 MPCs). Cells not released by proteolysis from the minced tissue were collected and expanded in a separate lot (=batch 2 MPCs).
The cells were expanded in expansion medium (E-medium: F10 medium, 15% FBS, 5 ng/mL bFGF, antibiotics, all from Sigma-Aldrich) [28,29]. The cell proliferation rate was monitored, and the MPCs were characterised. Differentiation and myotube formation were induced by differentiation medium (DMEM, low glucose, 2% horse serum, antibiotics) and documented by brightfield microscopy and immunofluorescence.
2.2. Analysis of Transcript Expression
The mRNA expression of myogenic marker genes was enumerated by RT-PCR [29,30]. The RNA was isolated (Aurum Total RNA mini kit; BioRad, Dreieich, Germany), yield and purity were determined by UV spectroscopy (Eon BioTec Instruments, Hillborough, NC, USA), and 1 μg was transcribed reversely (iScript cDNA synthesis kit; BioRad). For amplification of target genes and the housekeeping reference GAPDH (Table 1), a hot-start protocol followed by PCR amplification (39 cycles; iQ SYBR Green Supermix chemistry (BioRad)) and melting point analysis was performed (CFX95 Real-Time System; BioRad). The expression of the myogenic marker genes was normalised to the expression of GAPDH by the 2^(−∆∆CT)^-method [31,32] as described by the manufacturer (CFX Maestro software V2.3, BioRad).
2.3. Immunocytochemistry of Myoblasts
The expression of myogenic markers was explored by immunocytochemistry (ICC) [33]. The MPCs were washed with PBS, fixed (4% paraformaldehyde), washed, permeabilised (0.1% Triton X-100 in PBS (=T-PBS), and washed again with PBS. The cells were pre-incubated with blocking solution (5% dry milk powder). Desmin was detected by a rabbit antibody (# 15200, 1:200, abcam, Cambridge, UK) and a detection antibody (FITC- or PE-labelled F(ab’)2-donkey-anti-rabbit IgG, #711-096-152, 1:200, Jackson-Dianova, Hamburg, Germany). Unbound antibodies were aspirated, and the samples were washed with PBS-T. F-actin was detected by phalloidinstaining (ReadyProbes, Invitrogen, Carlsbad, CA, USA) and CD56 was detected by PE-labelled anti-CD56 antibody (clone MEM-188, 1:200, BioLegend, San Diego, CA, USA). The cell nuclei were counterstained by DAPI (Sigma-Aldrich). Samples omitting the primary reagents served as controls. The marker expression was observed by fluorescence microscopy (BZ-X800, Keyence, Mechelen, Belgium), evaluated by proprietary software (BZ-H1ase, Keyence). To compare the staining intensity of desmin on the different MPCs, the desmin- and DAPI-stained micrographs were each converted to an 8-bit grey-scale image (Atkinson mode; ImageJ V1.54g, NIH), and the positive signals were enumerated to compute a desmin staining intensity index, normalised to DAPI.
2.4. Flow Cytometry of Myoblasts
The expression CD56 was quantified by flow cytometry (FC) [34,35,36]. The MPCs were detached, washed, and counted, and 5 × 10^5^ MPCs were resuspended in FC sample buffer and incubated with PE-labelled anti-CD56 antibody (NCAM, # 318305, 1:10, BioLegend). The cells were washed, resuspended in 300 µL FC sample buffer, and analysed by FC (FACSCelesta; BD Biosciences, Heidelberg, Germany). Cells without antibody staining and FC compensation particles (BD Bioscience) were used to set the gates. The size and granularity of the MPCs were determined by the forward (FSC-A) and sideward (SSC-A) scatters, respectively. Debris and dead cells were excluded, and the expression of CD56 was enumerated. The data were processed using proprietary software (FACSDiva V9.0, FlowJo10.7.1; BD Biosciences).
2.5. Pig Husbandry and Providing a Large Animal Model of Urinary Incontinence
In our recent studies, cohort sizes of five pigs were computed to yield significant results in the pig SUI cell therapy model [25,37]: In this study, the sample size estimation was preformed using a two-group repeated measures design with an intra-subject correlation of 0.5 between repeated measurements. A sample size of five animals per group was required to achieve 80% to detect a mean difference of 80 mm H_2_O by a urodynamics device (see below) [38] at a two-side significance level of α = 0.05, assuming a correlation of 0.5a. To account for potential attrition due to withdrawal or loss of animals during the study, one additional animal was included in each group, and six gilts were enrolled in each group of the study. General health indicators, intake of water and food, and behaviour were monitored daily. On the day of the first surgery (=day 0), pigs were pre-medicated (10 mg/kg BW ketamine, 0.5 mg/kg BW azaperone i.m.), anaesthetised (4 mg/kg Propofol i.v.) and intubated for isoflurane anaesthesia. The urine status was determined (Combur 10M, Roche, Basle, Switzerland). The urethra was examined by cystoscope (Obturator 27026 U, 17 Ch, 27005BA Hopkins II 0° optics, KarlStorz, Tuttlingen, Germany). The urethral wall pressure (UWP) was recorded by a urodynamics device (T-DOC 7Fr dual sensor, Nexam Pro WPU-L4; Laborie, Portsmouth, NH, USA; ≙UWP day 0 “pre”). Then, the gilts were put in deep anaesthesia (isoflurane 1 Vol% under controlled respiration). The sphincter deficiency was induced in two steps [25] by electro-dissection of the urethra (i.e., electrocautery, Vio300D, monopolar cut, effect 4, power maximum = 100 W; ERBE Elektrotechnik, Tübingen, Germany) and balloon dilatation as described [22,25,39]. Immediately after its induction, sufficient sphincter deficiency was confirmed by urodynamics as described above (≙UWP day 0 “post”). Randomisation or blinding were not applied.
2.6. Cell Therapy of Incontinent Gilts and Follow-Up
Sufficient sphincter deficiency was confirmed on day three in sedated gilts of the experimental cohort by urodynamics (≙UWP day 3) before cell injections, and the urine status was recorded. The MPCs (2.4 × 10^6^/mL in E-medium) of the male littermate were injected by Williams needle laterally (as it were 3 and 9 o’clock) in two aliquots of 250 µL each via transurethral route by cystoscope under visual observation [22,25]. During follow-up, the health status was monitored daily. From day seven onward, the urine status was recorded (Combur 10M, Roche, Basle, Switzerland), and sphincter deficiency and its recovery were monitored weekly for six weeks by urodynamics as described above. In the control cohort, SUI was induced and monitored as in the experimental cohort, but neither solvent nor MPCs were injected on day three of follow-up.
2.7. Statistics
The generated data were exported to Microsoft Excel for processing and subsequently transferred to GraphPad Prism (V.106.1) for statistical analysis. Continuous variables were summarised as means with standard deviations (SDs) or as medians with interquartile ranges (IQRs), depending on their distribution. Data normality was assessed using kurtosis, skewness, Q–Q plots, histograms, and the Shapiro–Wilk test.
The primary endpoint of the study was the urinary weight pressure (UWP) measured repeatedly in each animal over the six-week follow-up period. To evaluate longitudinal changes in recovery and differences between groups (control vs. therapy) over time, a linear mixed-effects model was fitted. The model included group, time (days), and their interaction as fixed effects. A random intercept for each animal was included to account for within-animal correlation due to repeated measurements. Models were fitted using restricted maximum likelihood (REML). Degrees of freedom were estimated using the Satterthwaite method. In case of significant group-by-time interaction, post hoc comparisons were performed by splitting the data by group. Tukey correction was applied for multiple testing, and statistical significance was set at p < 0.05.
To analyse the secondary outcomes of the Merc’s cell therapy, comparisons between two independent groups were performed using the independent samples t-test for continuous variables. Levene’s test was used to verify homogeneity of variance, and in cases where this assumption was not met, Welch’s t-test was applied. A 95% confidence interval and a p-value < 0.05 were considered statistically significant. Given the limited sample size, missing values were addressed using a single imputation approach to minimise data loss, preserve statistical power, and reduce potential bias associated with case-wise deletion.
3. Results
3.1. Production and Quality Management of Porcine Myogenic Progenitor Cells
Myoblasts were isolated from striated muscle tissue containing elevated numbers of MPCs compared to the numbers of MPCs found in M. semitendinosus and used previously [28]. Accordingly, the MPCs were prepared from M. extensor carpi radialis (Mecr), M. rhomboideus cervicis (Mrc), and M. fibularis tertius (Mft); expanded; and characterised. Cells isolated from Mecr expressed desmin at the highest levels and on most cells (Figure 1A,B) when compared to cells from the Mrc or Mft muscle, respectively (Figure 1C–F). Only minor differences in desmin expression between Mecr batch one (Figure 1A) versus Mecr batch two cells (Figure 1B) were observed. Comparably, major differences in desmin expression were not noted in batch one versus batch two of Mrc MPCs (Figure 1C,D). In contrast, the low desmin expression in batch one Mft MPCs was even undercut in batch two cells (Figure 1E,F). The data provided evidence for a higher yield of desmin-positive MPCs isolated from Mecr when compared to Mrc and Mft in pig muscle tissues [28]. The normalised desmin staining intensity index of Merc MPCs was 4.25, while Mrc MPCs (0.78) and Mft MPCs (0.95) scored below 1. This confirmed elevated desmin expression in Merc MPCs compared to the MPCs from the other muscles.
The expression of myogenic marker transcripts was investigated by RT-qPCR (Figure 2). In batches one and two of the Mecr MPCs, prominent transcript levels encoding desmin (DES) and myosin light chain-1 (Myl1) were detected. In Mrc MPCs batch two and in Mft MPCs, expression of DES and Myl1 were lower compared to Mecr MPCs (Figure 2). Transcripts encoding myogenic differentiation factor (MyoD), skeletal muscle actin alpha 1 (ACTA1), myogenic factor 5 (Myf5), myogenin (MyoG), as well as the early marker of MPCs paired box 7 (Pax7) were somewhat lower in all MPCs (Figure 2). Again, transcript levels of these factors were partially very low in batch two Mrc MPCs and in Mft MPCs (Figure 2). In addition, expression of myostatin (MSTN), myogenic factor 6 (Myf6), and myosin heavy chain-1 (Myh1) were explored and, overall, very low levels of transcripts were uncovered (Figure 2). The expression of Myf6, was elevated only in batch one MPCs from Mecr. The transcript analyses of the individual MPC batches in the second passage of in vitro culture can be depicted in the online supplement (Figure A1).
Furthermore, the expression of myogenic marker transcripts was investigated as a function of cell passage in 2°P MPCs (Figure 2 A, D, G, J, M; blue bars) versus 3°P MPCs before terminal differentiation (Figure 2 B, E, H, K, N; black bars) and after terminal differentiation (Figure 2 C, F, I, L, O, grey bars). Overall, batches one and two of Mecr MPCs presented promising and stable mRNA expression patterns throughout expansion and in vitro differentiation. Batch one Mrc MPCs expressed some markers, including DES, Myl1, ACTA1, and Myf5 in 2°P and 3°P cells at levels comparable to MPCs from Mecr, but MyoD, MyoG, and PAX-7 were very low in 3°P Mrc MPCs (Figure 2). Batch two of Mrc MPCs were excluded from cell therapy as Myl-1, MyoD, ACTA1, and Myf5 were found at low levels as well (Figure 2). In Mft MPCs, satellite cell marker Pax7 and myogenic progenitor cell marker MyoG were not detected in 3°P cells (Figure 2). Therefore, Mft MPCs were excluded from cell therapy for this reason as well. Myf6, a key factor associated with terminal differentiation of MPCs, was expressed at the highest levels in batch one Mecr MPCs. All other batches expressed this factor at considerably lower levels (Figure 2). The transcript levels of the individual MPCs in passage 3 before and after differentiation are disclosed online in Figure A1.
CD56 is an adhesion molecule, and it is also considered a myogenic marker [34]. Thus, the CD56 expression was enumerated on the different MPC batches by flow cytometry (FC). More than 95% of the MPCs expressed CD56 with high mean fluorescence intensities. The narrow staining profiles of the histograms indicated uniform CD56-positive populations (Figure 3). This finding aligns with our recent studies [25,29].
Terminal differentiation of MPCs and generation of myotubes are key to facilitating their integration in striated muscles during muscle tissue regeneration [40,41]. The terminal differentiation and generation of multinuclear myotubes were explored in pooled Mecr MPCs. The changes in cell shape and the generation of elongated multinuclear myofibres were recorded by microscopy and ICC (Figure 4 and Figure 5). Staining with anti-desmin antibodies and phalloidin (Figure 4) or with anti-CD56 antibodies (Figure 5) presented elongated myotubes after differentiation of the MPCs. Based on these analyses, Mecr MPCs were selected as candidate cells for the preclinical SUI cell therapy study.
3.2. Induction of Sphincter Deficiency in a Pig Model of Incontinence and Cell Therapy
Sphincter deficiency was induced in the six gilts of the control cohort as described [22,25,39]. A robust sphincter deficiency was corroborated by urodynamics immediately after SUI induction in both groups (control cohort: 38.82% and experimental cohort: 19.3%) when compared to the percentage of UWP before SUI induction (=100%; Figure 6). Four weeks after sphincter injury, the spontaneous recovery reached 91.6% in the experimental cohort and 79.5% in the controls. To evaluate longitudinal changes in UWP recovery and differences between the therapy and control groups over time, a linear mixed model was performed. The model shows a significant group by time interaction, indicating that recovery trajectories differed between the groups over the four-week follow-up period (p value_interaction_ < 0.001). Therefore, a subgroup analysis was performed by stratifying the data according to group.
In the control group, an increase in the UWP after 21 days was recorded compared to the UWP immediately after SUI induction (“post”; Figure 6A). However, after 35 and 42 days of follow-up, the mean UWP dropped to 55.4% and 59.9%, respectively (Figure 6A). Longitudinal changes in UWP within the control group were assessed by using a linear mixed-effects model. No significant change in UWP was observed over time (β = 6.33, 95% CI −5.43 to 18.1, p = 0.304), suggesting that UWP does not improve over the follow-up period. The UWP timelines are presented online in Figure A2. These results are in line with our recent studies [25]. Due to a technical failure of the hardware for measuring urodynamics, UWPs could not be measured seven and fourteen days after SUI induction. Regrettably, a repetition of this control experiment was impossible. The animal welfare license did not allow for an additional six control animals, as our recent studies had already shown that the applied procedure yielded significant sphincter deficiency in landrace pigs (see [25,29]). We thus refer to these publications.
In the experimental cohort, sphincter deficiency was induced in six gilts as described above. Immediately after SUI induction, sphincter deficiency was confirmed by urodynamics, and the UWP was reduced significantly to a mean level of 19.3% compared to levels before SUI induction (=100%) (Figure 6B). After urodynamics on day three, 1.2 × 10^6^ MPCs were injected by a Williams needle in the zone of electro-dissection. The UWP of MPC-treated pigs remained at levels between 56.7% and 64.9% for three weeks. However, a robust rise in mean UWP from 64.9% to 91.6% and 86.3% was noted on days 28 and 35, respectively (Figure 6B). The linear mixed model in the therapy group shows a significant increment in the UWP over time (β = 52.1, 95% CI: 40.4 to 63.7, p < 0.001; Figure 6B). Injection of MPCs facilitated an almost full and functional recovery to 92% compared to “post” (17%) (Figure 6B). After six weeks of follow-up, the UWP of the experimental cohort was significant compared to the mock controls (β = 1.32, 95% CI: 0.03 to 2.56, p < 0.045; Figure 7). The UWP timelines of the individual animals are presented in the supplement Figure A2. We conclude that, in contrast to our recent study employing MPCs from M. semitendinosus [25], MPCs from M. extensor carpi radialis facilitated a significant (p < 0.001) and almost full recovery (92%) of the urethral sphincter complex in the porcine large animal model of SUI (Figure 6 and Figure 7).
4. Discussion
The efficacy of cell therapy for urethral sphincter deficiency has been investigated in porcine SUI models. Yet, therapies using cells isolated from M. gracilis or M. semitendinosus were not convincing [25,42]. In contrast, injection of ADSCs yielded significant and full recovery of the sphincter complex in the same model [25]. Hence, the porcine large animal model of SUI seems suitable for investigating the efficacy of SUI therapies with different cells and distinct surgical technologies. In this context, we hypothesised that the MPCs applied in the two studies mentioned above [25,42] might have contributed to the outcome of these studies. Significant differences in the composition of muscle tissue and the blend of muscle precursor cells in distinct muscles are well known [43]. This includes differences in satellite cell and MPC densities, as well as differences in species, sex, and age of the donor [28,44,45,46,47,48]. Such differences may even depend on the applied cell culture protocols [28,29,49,50]. An analysis of Pax7^pos^ in muscular stem cells provided evidence that porcine M. semitendinosus contained fewer Pax7^pos^ muscle stem cells compared to, e.g., M. psoas major or Mecr [28]. The considerably low content of satellite cells and MPCs in M. semitendinosus could explain the insufficient regenerative potential in sphincter repair observed in our recent study [25]. Analogously, one would assume that MPCs from Mecr may have facilitated a superior performance and contributed to the significant and almost complete regeneration of the sphincter complex reported in this exploratory study.
Moreover, the composition of the sphincter muscle may influence the outcome of an MPC therapy of SUI: While in the human sphincter complex type-I fibre muscles predominate [51], in the porcine urethra type-II fibres prevail [52]. But one must not overlook that in the quadruped pig, striated muscles are predominantly localised in the distal part of the urethra [53,54], while the urodynamic peak is found in young female gilts more proximally [55]. In this zone, smooth muscle but not striated muscle builds the urethral closure complex in pigs [54]. Thus, the regeneration of the deficient sphincter by injection of MPCs most likely does not require integration and differentiation of the cells into the muscle but may be facilitated by the release of cytokines and growth factors. This, however, remains to be investigated in our future studies.
The published clinical SUI studies employed cells isolated from quite different muscles, including, for instance, cells from M. quadriceps femoris [56], M. vastus lateralis [20], M. deltoideus [57,58], M. biceps brachii [26,27], and M. soleus [17], or did not specifically mention the cell source. The rationale behind the choice of cell sources employed was often not disclosed, and the results reported in the clinical studies spanned from almost complete success to predominant failure. The different outcomes reported from the clinical SUI cell therapy studies might be associated, in part, with the different MPC populations employed. Moreover, the cells applied were not characterised in depth in all studies. This brings about additional complexity when comparing the outcomes of such studies. Despite all limitations, our results could suggest that successful SUI treatment of patients with muscle-derived cells might be more successful when using proliferation- and differentiation-competent MPCs. However, based on the data obtained in our porcine SUI animal model, we, at least, are wary of defining criteria for cellular or molecular quality measures of MPCs produced for SUI cell therapies on human patients. A meta-analysis of the results of previous SUI cell therapy studies combined with histological and/or cell biological analyses of the muscle tissue and MCPs used in the individual studies could provide some initial information on which myogenic markers might be relevant for therapeutic success in human SUI patients.
We selected the donor muscles for MPC production based on their content of progenitor cells [28], as MPCs can be expanded in vitro, while differentiated myotubes, and even more so myofibres, cannot be expanded in cultures under normal conditions. The differentiation of MPCs to myotubes was studied in vitro to investigate the phenotype of the Merc MPCs at this level. This does not imply that terminal differentiation of the injected MPCs and formation of myofibres in situ is key for sphincter regeneration in this model. Therefore, analyses of type-I or type-II myofibres in tissue samples ex vivo were not included after follow-up. Moreover, our recent studies provided evidence that injected cells cannot be tracked for such a long period of time [23,59]. Hence, our study falls short in providing robust evidence on the cellular processes of sphincter regeneration. Our proof-of-principle study suggests that, at least in the pig model of SUI, chances for better outcomes of sphincter regeneration are associated with MPCs isolated from a muscle enriched for satellite cells and/or myogenic progenitor cells.
In the current experiments, we did not plan a randomised controlled trial with large cohorts and multiple tissue donors per cohort. In contrast, this study focused on identifying more efficient MPCs for sphincter muscle regeneration. To this end, MPCs from different muscles were compared to the cells derived from M. semitendinosus used in our previous studies [22,25,38]. Pigs with induced sphincter deficiency without sham injections of interfering substances served as mock controls. The protocol for induction of sphincter deficiency, injection of MPCs, or urodynamics during follow-up was not altered in this study compared to our previous ones [25]. Nonetheless, future studies should be designed on a conformational level with larger cohorts and include additional controls such as a sham-injection of biologically inactive components. To avoid bias, such studies should be blinded with respect to verum versus placebo treatment and possibly even include independent teams or centres for cell production, animal surgery, and follow-up.
However, each preclinical animal model has specific disadvantages, and translating our results into clinical studies requires additional studies. In rabbits, for instance, regeneration of the urethral sphincter has been reported following the application of cells isolated from the M. soleus [60]. However, this regeneration was not superior to cell-free injection of the solvent, which contained plasma [60]. Moreover, the phenotype of the cells applied in this rabbit study was not investigated. One may conclude that the solvent complemented by plasma brought about sphincter regeneration in this set-up, not the cells. Thus, the efficacy of defined MPCs from M. soleus in sphincter regeneration remains to be investigated in animal models. By contrast, the application of MPCs from M. soleus in combination with electrophysiological stimulation seemed promising in women suffering from SUI [17]. Moreover, in most preclinical SUI animal studies, young animals were utilised. We noticed differences in the UWP profiles when young versus old as well as virgin versus multipara Göttingen minipigs were explored [55]. By contrast, most SUI patients are typically in an advanced stage of life. This necessitates a further restriction for the translation of our results in clinics. In addition, the individual breed of the pigs employed in such studies is critical when establishing an SUI model [37]. Reasons for these differences and the clinical relevance remain to be investigated.
Another factor contributing to the regenerative efficacy is the cell expansion protocol applied [28,29,49,50]. This includes the variables observed in the distinct cell sets or batches obtained in parallel preparations from the same muscle and donor animal reported here. For instance, batch one MPCs from Merc expressed higher levels of the early MPC marker MyoD than batch two MPCs from Merc. This is probably associated with the slightly different method utilised for the isolation of the cells. Likewise, different expression patterns between batches one and two were detected in MPCs from Mrc and Mft as well. An almost complete loss of the MPC-marker MyoG was reported in undifferentiated Mft MPCs over time [61], which is corroborated by our results from batch one MPCs from Mrc and Mft, but not from Merc. Changes in the expression of β-actin (ACT) as a function of cell culture passage or differentiation were not expected, as this protein is expressed in basically all nucleated cells. The consistently high expression levels of ACT demonstrated a comparable efficiency of RNA extraction, cDNA synthesis, and PCR amplification throughout the study.
The situation of the muscle to be targeted by MPC therapy may also contribute to the clinical success. Thus, in preclinical animal studies of SUI cell therapies, the injury of the urethra and the timeline until cell application will influence the outcome [41]. In addition, cell dosages investigated in animal studies may not translate directly to human therapies [14]. Moreover, the exact positioning of the injected cells contributes to the therapy’s outcome [24]. This, again, differs in humans in comparison, e.g., to the porcine SUI model due to the anatomical and physiological characteristics in these quadruped animals [22,53,62].
In our previous studies, a transient elevation in the UWP was recorded in the controls as well as in the verum cohort peaking approximately two weeks after SUI induction [24,25]. This data is missing for the control cohort in our study. Spontaneous muscle regeneration after mechanical injury will take about two weeks for measurable functional recovery [63]. Thus, this transient elevation in the UWP was associated with the spontaneous processes of sphincter repair after mechanical injury. But this was not the focus of the study. The focus of this study was to explore whether Merc MPCs facilitated better sphincter regeneration compared to the cells utilised previously [25,29].
5. Conclusions
Porcine myogenic progenitor cells isolated from M. extensor carpi radialis expressed high and stable levels of muscle stem cell markers Pax7 and Myf5, as well as myogenic progenitor markers MyoD and MyoG. Such cells facilitated a full and significant recovery from urethral sphincter deficiency in our large animal model of stress urinary incontinence. This study complements our recent publication, which provided evidence that ADSCs but not MPCs isolated from M. semitendinosus were capable of doing so. Thus, both types of cells, ADSCs as well as MPCs from Mecr, are promising candidates for further pre-clinical and possibly even clinical studies of cell therapy of incontinence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pizzol D. Demurtas J. Celotto S. Maggi S. Smith L. Angiolelli G. Trott M. Yang L. Veronese N. Urinary incontinence and quality of life: A systematic review and meta-analysis Aging Clin. Exp. Res.202133253510.1007/s 40520-020-01712-y 32964401 PMC 7897623 · doi ↗ · pubmed ↗
- 2Batmani S. Jalali R. Mohammadi M. Bokaee S. Correction: Prevalence and factors related to urinary incontinence in older adults women worldwide: A comprehensive systematic review and meta-analysis of observational studies BMC Geriatr.20222245410.1186/s 12877-022-03111-635614400 PMC 9134701 · doi ↗ · pubmed ↗
- 3Shang X. Fu Y. Jin X. Wang C. Wang P. Guo P. Wang Y. Yan S. Association of overweight, obesity and risk of urinary incontinence in middle-aged and older women: A meta epidemiology study Front. Endocrinol.202314122055110.3389/fendo.2023.1220551 PMC 1059834537886637 · doi ↗ · pubmed ↗
- 4Olagundoye O. Odusanya B. Kung J.Y. Gibson W. Wagg A. A scoping review of risk factors for urinary incontinence in older men BMC Geriatr.20232353410.1186/s 12877-023-04249-737660036 PMC 10474661 · doi ↗ · pubmed ↗
- 5Gacci M. Sakalis V.I. Karavitakis M. Cornu J.-N. Gratzke C. Herrmann T.R.W. Kyriazis I. Malde S. Mamoulakis C. Rieken M. European Association of Urology Guidelines on Male Urinary Incontinence Eur. Urol.20228238739810.1016/j.eururo.2022.05.01235697561 · doi ↗ · pubmed ↗
- 6Milsom I. Gyhagen M. The prevalence of urinary incontinence Climacteric 20192221722210.1080/13697137.2018.154326330572737 · doi ↗ · pubmed ↗
- 7Harland N. Walz S. Eberli D. Schmid F.A. Aicher W.K. Stenzl A. Amend B. Stress Urinary Incontinence: An Unsolved Clinical Challenge Biomedicines 202311248610.3390/biomedicines 1109248637760927 PMC 10525672 · doi ↗ · pubmed ↗
- 8Wilson L. Brown J.S. Shin G.P. Luc K.-O. Subak L.L. Annual direct cost of urinary incontinence Obstet. Gynecol.2001983984061153011910.1016/s 0029-7844(01)01464-8 · doi ↗ · pubmed ↗