The effects of postbiotics and glycyrrhetinic acid on immune response and inflammation-related genes during H. pylori eradication therapy
Erhan Tek, Nizami Duran, Elif Yaprak Colak, Gulay Gulbol Duran, Tuncer Kutlu, Hamdullah Suphi Bayraktar, Sibel Dagli, Mehmet Demir

TL;DR
This study shows that postbiotics and glycyrrhetinic acid can reduce inflammation and H. pylori bacterial load in gastric cells and rat models.
Contribution
The study introduces a novel combination of postbiotics and glycyrrhetinic acid as potential adjuncts for H. pylori eradication therapy.
Findings
Postbiotics from S. thermophilus and L. casei reduced NF-κB and TNF-α expression by up to 10-fold in AGS cells.
The quaternary treatment group achieved complete H. pylori eradication with zero detectable bacterial load in rat models.
Combination treatments showed significant reductions in proinflammatory cytokines like IL-1β, IL-6, and IL-8.
Abstract
This study aims to test the effectiveness of postbiotics from Streptococcus thermophilus and Lactobacillus casei, along with Glycyrrhetinic acid, against H. pylori in both AGS cells (a human gastric adenocarcinoma cell line) and an experimental rat gastritis model. The effectiveness of the compounds (postbiotics and G. acid) against H. pylori was evaluated by measuring the expression levels of various genes, including NF-κB, IL-1β, IL-6, IL-8, IL-10, TNF-α, COX-2, FOX-M1, and IL-33 in AGS cells using the Real-Time PCR method. Additionally, bacterial loads in gastric tissue were quantified using culture-based colony-forming unit (CFU) analysis and expressed as log₁₀ CFU/g. The study also evaluated the effectiveness of combinations of these components in the rat gastritis model through microbiological and histopathological analyses. The postbiotics of S. thermophilus and L. casei have…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
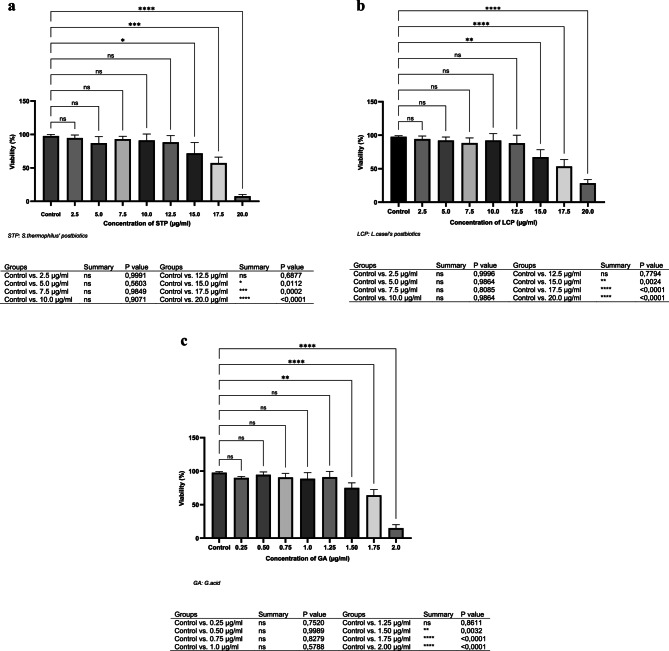 Figure 1
Figure 1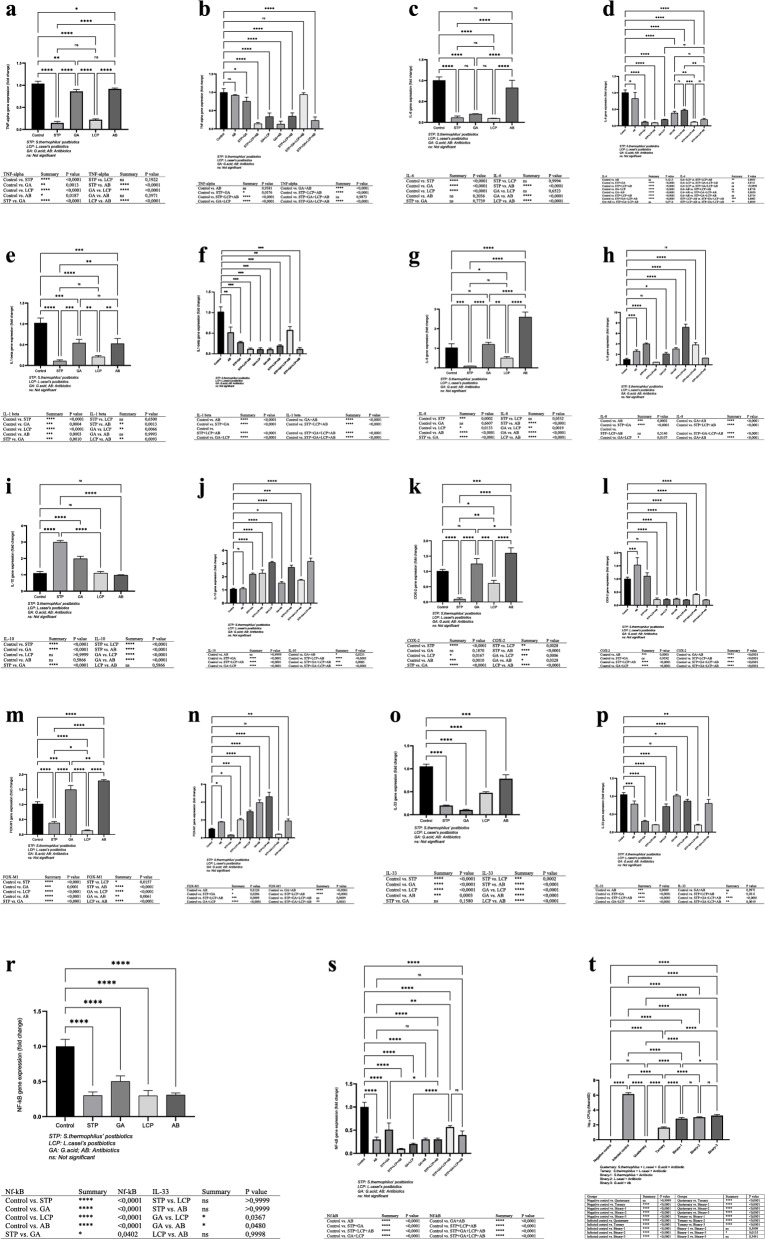 Figure 2
Figure 2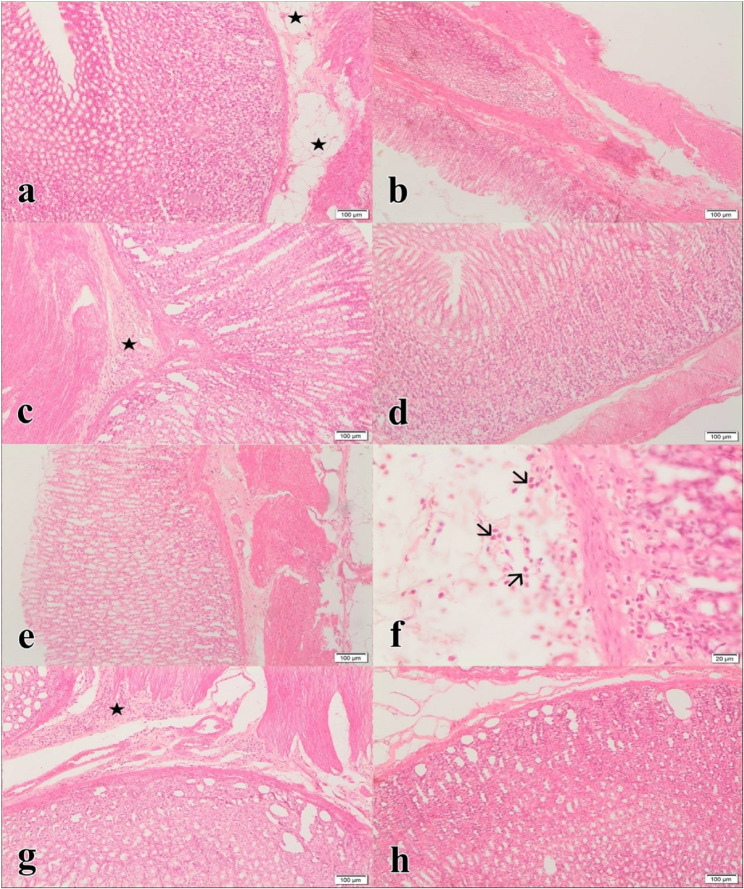 Figure 3
Figure 3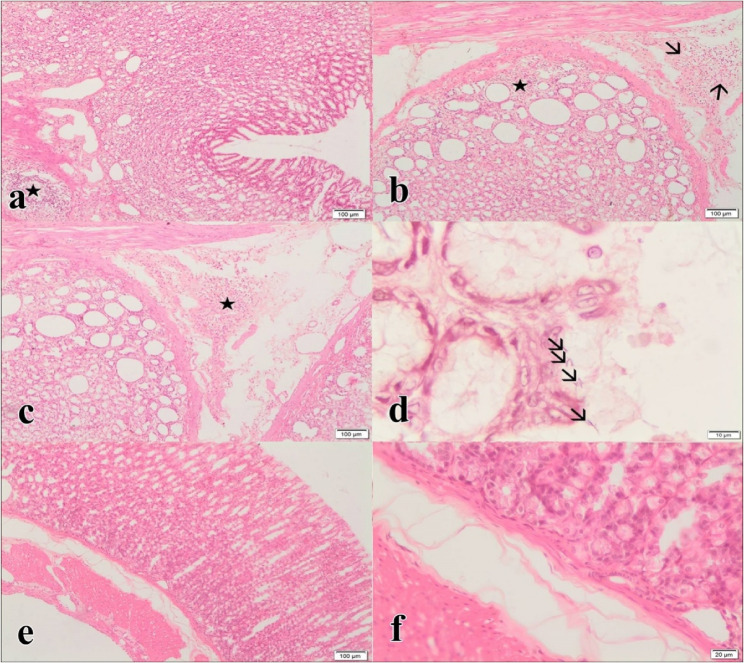 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHelicobacter pylori-related gastroenterology studies · Pharmacological Effects of Natural Compounds · Gut microbiota and health
Introduction
Helicobacter pylori is a Gram-negative, spiral-shaped, microaerophilic bacterium that infects over half of the global population, contributing to chronic gastritis, peptic ulcers, and stomach cancer [1]. H. pylori is treated with antibiotics and PPIs, but antibiotic overuse has led to resistance and treatment failures. Long-term antibiotic use also causes adverse effects, prompting research into alternative therapies and improved drug efficacy [2, 3].
Most drug research has recently been conducted on natural or herbal products due to their low cytotoxic effects. One such plant commonly used for treating stomach ailments is Glycyrrhiza glabra, which contains Glycyrrhizic acid, a potent ingredient that can be hydrolyzed into Glycyrrhetinic acid. Glycyrrhetinic acid has demonstrated antimicrobial and anti-inflammatory effects, and previous studies reported its efficacy against H. pylori in vitro. Its inclusion in this study aimed to explore its synergistic potential when combined with postbiotics and antibiotics. Furthermore, Glycyrrhetinic acid has been reported to suppress pro-inflammatory cytokines (e.g., IL-1β, TNF-α) and inhibit NF-κB pathway activation, both of which are critical in H. pylori-induced gastric inflammation and carcinogenesis. The rationale for including G. acid in this study was twofold: (1) to explore its potential synergistic effects when combined with postbiotics and antibiotics, aiming to enhance eradication rates while mitigating inflammation, and (2) to evaluate its therapeutic relevance in addressing antibiotic resistance by contributing a natural anti-H. pylori and anti-inflammatory agent to combination therapy. This approach aligns with recent literature emphasizing the role of phytochemicals in combating antibiotic-resistant pathogens and improving host immune response [4–7]. Various studies have reported that Glycyrrhiza glabra exhibits antibacterial [8], antiviral [4], anti-inflammatory [5], antiulcer [9], antiallergic [10], anti-oxidant [11], and anti-tumoral [12] properties. However, the effectiveness of G.acid against H. pylori, a bacteria responsible for causing stomach ulcers, has only been tested in a limited number of studies [13, 14]. In a previous study, G.acid showed significant antimicrobial activity against H. pylori in vitro [6]. Conducting in vivo studies will help clarify the effectiveness of G.acid against H. pylori and determine its potential for treating stomach ulcers [15, 16].
Some studies suggest that supplementing with certain strains of probiotics may help improve eradication rates when antibiotic therapy is not effective enough for treating H. pylori infection [13]. While some probiotics may have toxic effects, many are well-tolerated and produce bioactive compounds with antimicrobial and immune-modulating properties. They enhance host resistance to pathogens, regulate gut microbiota, and influence immune responses by modulating cytokine expression [17–21].
Probiotics produce bioactive substances such as short-chain fatty acids, enzymes, proteins, organic acids, vitamins, and amino acids. These substances are also known as postbiotics. The cell wall components of these bacteria also have similar health effects. Postbiotics are compounds that are produced by probiotic bacteria. They contain soluble factors such as various enzymes, proteins, polysaccharides, short-chain fatty acids, and peptidoglycans. These compounds have immunomodulatory and anti-inflammatory effects on the host, improving their health and well-being [22, 23].
Lactobacillus and Streptococcus postbiotics, composed of secreted metabolites such as short-chain fatty acids, exopolysaccharides, and bacteriocins, have been reported to be effective in boosting cell-mediated immunity and reducing inflammation. While both strains share core components, S. thermophilus-derived postbiotics are rich in exopolysaccharides, whereas L. casei postbiotics are known for their immunoregulatory proteins and higher peptidoglycan content, which may explain the variation in their anti-inflammatory activity. Lactobacillus species produce postbiotics that play a vital role in supporting the immune system of the intestinal mucosa. In one study, the potential effect of postbiotic fractions of intestinal L. rhamonosus and L. plantarum on the immune response induced by pro-inflammatory stimuli was investigated. The study reported that the presence of probiotic bacterial components on the mucosal surface during the initial and final stages of inflammatory conditions results from cellular interactions that regulate inflammation and prevent damage to the intestinal epithelium. Postbiotics of L.rhamnosus and L.plantarum have been shown to activate IL-10, an anti-inflammatory cytokine, and reduce inflammation by regulating the IL-18-related response [24, 25].
Studies have indicated that infants who consume infant formula containing postbiotic products derived from S.thermophilus experience less severe acute diarrheal attacks, an increase in thymus size, a decrease in fecal calprotectin, and an increase in secretory IgA. Furthermore, S.thermophilus postbiotics stimulate immunomodulatory reactions in dendritic cells by increasing the release of IL-10 and stimulating the Th1 immune system. Although the advantageous properties of postbiotics from probiotic bacteria are well established, the mechanisms underlying their interaction with immune cells and their ability to affect immune modulation require further investigation. Moreover, the precise molecular mechanism that enables postbiotics to produce positive effects on the host is yet to be fully comprehended [26–29].
Probiotics, mainly lactic acid bacteria like Lactobacillus and Streptococcus, modulate gut microbiota and exhibit antimicrobial activity. S. thermophilus exo-polysaccharides may reduce H. pylori-induced inflammation, but further studies are needed. Lactobacillus strains show anti-H. pylori effects and enhance immune responses by modulating T-cell balance and stimulating phagocytosis, chemokine, and cytokine production [18, 30–36].
Scientific research on the activities of probiotic microorganisms against H. pylori remains limited. This study aimed to investigate the role of postbiotics derived from probiotic microorganisms in improving H. pylori treatment outcomes. Following a comprehensive review of the literature, although some previous studies addressing this topic exist, to the best of our knowledge, this is the first study to evaluate the combined therapeutic potential of antibiotics, S. thermophilus and L. casei postbiotics, and glycyrrhetinic acid in both in vitro and in vivo models. The findings of this study may serve as a valuable reference for future research in this field.
The efficacies of combinations of G.acid, S.thermophilus, and L.casei’s postbiotics against H.pylori were evaluated in AGS cells (human gastric adenocarcinoma cell line) and a rat model of gastritis induced by H.pylori. The expression levels of NF-kB,* IL-1B*,* IL-6*,* IL-8*,* IL-10*,* TNF-α*,* COX-2*,* FOX-M1*, and IL-33 genes were determined in AGS cells. Then, the efficacies of these components in the rat gastritis model experimentally induced with H.pylori were compared with the standard antibiotic treatment groups. Microbiological and histopathological techniques were used for in-vivo evaluations.
Materials & methods
The effects of G.acid, S. thermophilus, and L. casei postbiotics on H. pylori were assessed in AGS cells and a rat gastritis model. Gene expression levels (NF-κB, IL-1B, IL-6, IL-8, IL-10, TNF-α, COX-2, FOX-M1, IL-33) were analyzed in AGS cells. In-vivo efficacy was compared with standard antibiotic treatment using microbiological and histopathological methods.
H. pylori culture
H. pylori (NTCC 11637) was obtained from Ankara Refik Saydam Hygiene Institute. Strains were incubated in Brucella Broth with 5% FBS under microaerophilic conditions for five days, then passaged onto 5% sheep blood agar and incubated at 37 °C for seven days. Strains were sensitive to Amoxicillin and Clarithromycin, with susceptibility tests performed before experiments [37].
Postbiotics of L. casei and S. thermophilus
L. casei and S. thermophilus were cultured on MRS agar at 37 °C under anaerobic conditions. Supernatants were collected by centrifugation (6000 rpm, 30 min, 4 °C) and filtered (0.45 μm). Blank MRS medium was processed similarly as a control.
Glycyrrhetinic acid and antibiotics
G.acid (Sigma-Aldrich, USA) was dissolved in 0.1% DMSO (Merck) for Vero cells. Amoxicillin (Deva, TR) and Clarithromycin (Abbott, USA) were used as standard drugs. H. pylori susceptibility was tested before the study (MIC: Amoxicillin 0.10 µg/mL, Clarithromycin 0.20 µg/mL) [37].
Cell culture
Vero (ATCC CCL-81) and AGS (ATCC CRL-1739) cell lines were used as healthy and H. pylori-infected models, respectively. Cells were cultured in RPMI-1640 with 10% FBS, HEPES, penicillin/streptomycin, and glutamine at 37 °C, 5% CO₂. Non-toxic doses of postbiotics and G.acid were determined in Vero cells. Cell density was set at 1 × 10⁶ cells/mL for experiments.
AGS cells were infected with H. pylori (1 × 10⁸ CFU/mL, MOI 100) for 6 h at 37 °C with 5% CO₂. Incubation continued for 96 h to assess the effects of postbiotics and G.acid on intracellular bacteria. Cells were collected at specific time points for PCR analysis.
Cytotoxicity tests
Cytotoxicity was evaluated using Vero cells in 96-well plates (1 × 10⁶ cells/mL) with RPMI 1640 + 10% FBS. Compounds were dissolved in 0.1% DMSO, which served as a negative control. Non-toxic concentrations of G.acid, S. thermophilus, and L. casei postbiotics were determined by the MTT assay [38].
MTT method
The MTT assay, a standard method for assessing cell viability and cytotoxicity, was used to evaluate the effects of postbiotics, G.acid, and drugs (Lansoprazole, Amoxicillin, Clarithromycin) on Vero cells. Cells (1 × 10⁵/well) were seeded in 96-well plates and incubated at 37 °C with 5% CO₂ for 24 h. Serial dilutions of postbiotics (2.5–20.0 µg/mL) and G.acid (0.25–2.0 µg/mL) were added. After incubation, MTT solution was applied for 2 h, followed by DMSO to dissolve formazan. Absorbance at 570 nm was measured using a spectrophotometer. IC50 values were determined from dose-response curves, and cytotoxicity was calculated as test/control absorbance ×100. Experiments were repeated three times per concentration.
Activity studies
Non-toxic doses of postbiotics (12.5 µg/mL) and G.acid (1.25 µg/mL) were determined using Vero cells. Cell viability and morphology were monitored daily under an inverted microscope. Cells (1 × 10⁵/mL) were incubated at 37 °C with 5% CO₂, treated with test compounds, and incubated for 96 h. After trypsinization and centrifugation, viability was assessed via hemocytometer, and mRNA expression levels were analyzed by RT-PCR.
Infection of AGS cells with H. pylori
AGS cells were cultured in DMEM with 4 mM L-glutamine, 10% FBS, and antibiotics. For gene expression studies, antibiotic-free medium was used. The effects of postbiotics, G.acid, and their combinations on NF-κB, IL-1B, TNF-α, FOX-M1, IL-6, IL-8, IL-10, and IL-33 gene expression were evaluated against an untreated control group (Table 1).
Table 1. Experimental groups and numbers in gene expression studyGroupsThe number of repetitionsGroup 1 (HP group)3Group 2 (STP group)3Group 3 (GA group)3Group 4 (LCP group)3Group 5 (AB group3Group 6 (STP + GA group)3Group 7 (STP + LCP group)3Group 8 (GA + LCP group)3Group 9 (GA + AB group)3Group 10 (STP + LCP + AB group)3Group 11 (STP + GA + LCP group)3Group 12 (STP + GA + LCP + AB group)3 HP: H.pylori
STP: S.thermophilus’ postbiotics
GA: Glysithetinic acid
LCP: L.casei’s postbiotics
AB: (Amoxicillin + Clarithromycin)
RNA isolation
Total RNA was isolated from AGS cells using High-Pure RNA Tissue Kits (Thermo Fisher Scientific, USA). DNase I treatment was applied to eliminate DNA contamination. RNA integrity was confirmed via agarose gel electrophoresis. cDNA was synthesized using the Thermo Scientific cDNA Synthesis Kit and stored at −20 °C until use.
RT-qPCR
Gene expression levels of NF-κB, IL-1B, IL-6, IL-8, IL-10, TNF-α, COX-2, FOX-M1, and IL-33 were analyzed using SYBR Green-based RT-qPCR on the Montana 4896 system (Anatolia, Türkiye). PCR conditions included initial denaturation at 95 °C for 10 min, followed by 35 cycles of 95 °C for 15 s and 60 °C for 60 s, with a final step at 99.9 °C for 10 min. GAPDH was used for normalization, and relative expression levels were calculated using the 2^−∆∆Ct^ method. Primer sequences are provided in Table 2.
Table 2. Oligonucleotide sequences were used in the present studyPrimersSequencesReferencesGAPDH5‘-TGCACCACCAACTGCTTAGC-3‘315‘-GGCATGGACTGTGGTCATGAG-3’IL-335’-GGAAGAACACAGCAAGCAAAGCCT-3’325’-TAAGGCCAGAGCGGAGCTTCATAA-3’FOX-M15’-TGCAGCTAGGGATGTGAATCTTC-3’335’-GGAGCCCAGTCCATCAGAACT-3’COX-25′-CTTGCTGTTCCCACCCATGTCAAA-3′345′-TGCACTGTGTTTGGAGTGGGTTTC-3′ TNF-alpha 5′-CGAGTGACAAGCCTGTAGC-3′355′-GGTGTGGGTGAGGAGCACAT-3′IL-105′-GTGATGCCCCAAGCTGAGA-3′365′-CACGGCCTTGCTCTTGTTTT-3′IL-65′-AAATGCCAGCCTGCTGA CGAAC-3′345′-AACAACAATCTGAGGTGCCCATGCTAC-3′IL-1 beta5’-AAGCCCTTGCTGTAGTGGTG-3’375’-GAAGCTGATGGCCCTAAACA-3’IL-85’-AGCACTCCTTGGCAAAACTG-3’385’-CGGAAGGAACCATCTCACTG-3’Nf-kB5’-AAAGACACATCCGGACCTCG-3’395’-TGTAAGAGTTCCCCTCCGGT-3’
In-vivo tests
This study was approved by the Hatay Mustafa Kemal University Animal Ethics Committee (27.12.2018, No: 2018/11 − 9) and conducted at the university’s Experimental Research Center. All animals used in this study were sourced directly from the Experimental Research Center of Hatay Mustafa Kemal University. The animals were bred and maintained under institutional care and were not privately owned by any external institution, individual, or farm. Therefore, no external informed consent was required.
Male albino Wistar rats were randomly divided into 13 groups (Table 3) and housed under standard conditions (23 ± 2 °C, 12 h light/dark cycle) with ad libitum access to standard feed and water. A total of 65 male Wistar albino rats (~ 300 g) were used. Rats were inoculated with H. pylori (5 × 10⁸ cfu/mL, 1 mL/rat, twice daily for 3 days). After 15 days, colonization and gastritis were confirmed by histopathological and microbiological analysis. Rats received a standard eradication regimen (amoxicillin 50 mg/kg, clarithromycin 25 mg/kg, lansoprazole 20 mg/kg). Safe, non-toxic doses of postbiotics (S. thermophilus,* L. casei* at 12.5 µg/mL) and G. acid (1.25 µg/mL) were determined based on rat weight. Daily doses were 3.75 mL/rat for postbiotics and 0.375 mL/rat for G. acid, administered via gavage [39].
Table 3. Animal experiments treatment groups and numbersExperimental groupsNumbers of ratsGroup 1 (HP group)5Group 2 (Healthy control group)5Group 3 (STP treatment group)5Group 4 (GA treatment group)5Group 5 (LCP treatment group)5Group 6 (AB group)5Group 7 (STP + GA group)5Group 8 (STP + LCP group)5Group 9 (GA + LCP group)5Group 10 (GA + AB group)5Group 11 (STP + LCP + AB group)5Group 12 (STP + GA + LCP group)5Group 13 (STP + GA + LCP + AB group)5Total number of animals65HP: H.pylori**STP: S.thermophilus’ postbioticsGA: Glysithetinic acid**LCP: L.casei’s postbioticsAB: (Amoxicillin + Clarithromycin)
The ulcer index was measured by considering the following criteria as stated in the literature [40]. For this, the number and severity of the lesions were evaluated. The following scores were used for evaluation: I: Mild; II: Medium, the presence of edema, hyperemia, and single petechiae; III: The presence of submucosal hemorrhagic lesions with small erosions; IV: The presence of hemorrhagic lesions with severe erosions. Ulcer Index (UI) = (nI) + (nII) × 2 + (nIII) × 3/Number of animals (“n” is the number of lesions). Then, the indicated treatments were applied to the following treatment groups for ten days. Following the development of gastritis, the compounds and their various combinations were administered at 1 mL per rat per day. The treatment was orally administered daily during the ten-day treatment period.
At the end of the experimental procedures, all animals, including those in the control groups, were humanely sacrificed under deep anesthesia. A combination of ketamine (150 mg/kg) and xylazine (9 mg/kg) was administered intraperitoneally (i.p.) to ensure adequate anesthesia and unconsciousness before sacrifice. The selected dosages and administration route were based on widely accepted guidelines for laboratory rodents, aiming to minimize animal pain and distress. Following confirmation of deep anesthesia (loss of righting reflex and pedal withdrawal reflex), rats were sacrificed by exsanguination through cardiac puncture. This euthanasia protocol was reviewed and approved by the Hatay Mustafa Kemal University Animal Ethics Committee (Approval No: 2018/11 − 9), by institutional guidelines, and international ethical standards for animal research. This ensured that all procedures complied with ARRIVE guidelines and relevant national legislation.
Following the sacrifice, the rats’ gastric tissues were removed. Then, the tissue samples from rats were placed in PBS for microbiological analysis, in formaldehyde for histopathological analysis, and sent to laboratories.
Microbiological analysis
Following euthanasia, gastric tissues were aseptically excised and immediately transferred into 10 mL of cold phosphate-buffered saline (PBS). One-gram tissue samples were homogenized and plated onto blood agar supplemented with 5% sheep blood. Plates were incubated at 37 °C for 72 h under microaerophilic conditions. Colonies were counted and expressed as log₁₀ CFU per gram of tissue. Each experimental group was analyzed in triplicate using independently treated animals.
In the infected control group, the mean bacterial load was 6.14 ± 0.18 log₁₀ CFU/g tissue (individual values: 6.32, 5.97, 6.13 log₁₀ CFU/g). No bacterial growth was detected in the negative control group (0.00 ± 0.00 log₁₀ CFU/g).
For treatment groups, the G. acid + Antibiotic (Binary-3) group had 3.24 ± 0.17 log₁₀ CFU/g (3.10, 3.20, 3.43), the L. casei + Antibiotic (Binary-2) group had 3.02 ± 0.07 log₁₀ CFU/g (3.02, 3.10, 2.95), and the S. thermophilus + Antibiotic (Binary-1) group had 2.83 ± 0.17 log₁₀ CFU/g (2.80, 3.02, 2.68). The ternary combination group (S. thermophilus + L. casei + Antibiotic) showed 1.63 ± 0.09 log₁₀ CFU/g (1.55, 1.73, 1.61), while the quaternary combination group (S. thermophilus + L. casei + G. acid + Antibiotic) exhibited complete eradication with no detectable colonies (0.00 ± 0.00 log₁₀ CFU/g).
Intermediate bacterial loads were observed in the binary and ternary treatment groups, while the quaternary group demonstrated total bacterial eradication. Infection rates were statistically evaluated using chi-square (χ²) tests (n = 5 per group), while CFU count comparisons among groups were analyzed using one-way ANOVA followed by Tukey’s post hoc test [41].
Pathological evaluation
Gastric tissues were fixed in 10% neutral buffered formalin, washed, dehydrated in graded alcohols, cleared with xylene, and embedded in paraffin. Section (5 μm) were cut, deparaffinized, rehydrated, and stained with Hematoxylin and Eosin (H&E). Tissue images were captured using a light microscope (Olympus CX31). Gastritis severity was assessed semi-quantitatively as mild, moderate, or severe [42].
Statistical analysis
Data were analyzed using GraphPad Prism v10. Results are expressed as mean ± SD from at least three experiments. Statistical comparisons were made using one-way ANOVA with Tukey’s post hoc test or unpaired two-tailed t-tests where appropriate. Differences were considered significant at p < 0.05. Significance levels: p < 0.05 (),* < 0.001 (), < 0.0001 (*), < 0.00001 (**), and ns for non-significant results.
Results
Vero cell experiments identified safe, non-toxic concentrations: G.acid (≤ 1.25 µg/mL) and postbiotics (≤ 12.5 µg/mL) (Fig. 1a-c). TNF-α expression remained unchanged in antibiotic and G.acid groups but was reduced 10-fold with S. thermophilus and 5-fold with L. casei postbiotics (Fig. 2a).Fig. 1a-c. Cytotoxicity results of the S.thermophilus' (a) and L.casei’s (b) postbiotics, and G.acid (c) on the viability of Vero cells by MTT methodFig. 2a. Effects of G. acid, S. thermophilus, and L.casei’s postbiotics on TNF-alpha expression on H.pylori-infected AGS cells. b. Synergistic effects of different combinations of G.acid,S. thermophilus, and L.casei’s postbiotics with the antibiotic group on TNF-alpha expression in H.pylori-infected AGS cells. c. Efficacy of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-6 expression in H.pylori-infected AGS cells. d. Synergistic effects of different combinations of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-6 expression in H.pylori-infected AGS cells. e. The activities of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-1**beta expression in H.pylori-infected AGS cells. f. Synergistic effects of different combinations of G.acid, S. thermophilus, and L.casei’s postbiotics on IL-1B expression in H.pylori-infected AGS cells. g. Efficacies of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-8 synthesis in H.pylori-infected AGS cells. h. Synergistic effects of different combinations of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-8 synthesis on H.pylori infected AGS cells. i. Effects of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-10 synthesis in H.pylori-infected AGS cells. j. Synergistic effects of different combinations of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-10 expression in H.pylori-infected AGS cells. k. Comparison of the effects of G.acid, S.thermophilus, and L.casei’s postbiotics on COX-2 synthesis with antibiotic treatment in H.pylori-infected AGS cells. l. Comparison of the synergistic effect of different combinations of G.acid, S.thermophilus, and L.casei’s postbiotics with antibiotic treatment on the COX-2 gene expression in H.pylori-infected AGS cells. m. Effects of G.acid, S. thermophilus, and L.casei’s postbiotics on FOX-M1 expression in H.pylori-infected AGS cells. n. Synergistic effects of different combinations of G.acid, S.thermophilus, and L.casei’s postbiotics on FOX-M1 expression in H.pylori-infected AGS cells. o. Effects of G. acid, S.thermophilus, and L.casei’s postbiotics on IL-33 expression in H.pylori-infected AGS cells. p. Synergistic effects of different combinations of G.acid, S.thermophilus, and L.casei’s postbiotics on IL-33 expression in H.pylori-infected AGS cells. r. Comparison of the effect of G.acid, S.thermophilus, and L.casei’s postbiotics on NF-kB expression level in the H.pylori-infected AGS cells with the antibiotic group. s. Comparison of the synergistic effect of different combinations of G.acid, S.thermophilus, and L.casei’s postbiotics with the antibiotic group on the level of NF-kB expression in H.pylori-infected AGS cells. t. Comparison of gastric H. pylori bacterial load among treatment groups based on CFU counts (log₁₀ CFU/g): Demonstrating the impact of postbiotics, Glycyrrhetinic acid, and antibiotics
The combination of S. thermophilus and L. casei postbiotics significantly inhibited TNF-α expression. A strong inhibitory effect was also observed in the G.acid plus antibiotic group (Fig. 2b).
IL-6 expression in H. pylori-infected AGS cells was reduced 10-fold with S. thermophilus and L. casei postbiotics and 5-fold with G.acid, while the antibiotic group showed no significant decrease (Fig. 2c).
Both individual and combined postbiotic treatments significantly reduced IL-6 gene expression. Notably, combinations including S. thermophilus with G. acid, L. casei, or both led to a 10-fold decrease compared to the control. Other combinations also showed significant reductions, while the antibiotic group showed no notable change (Fig. 2d).
S. thermophilus and L. casei postbiotics showed strong inhibitory effects, reducing gene expression by 10-fold and 5-fold, respectively, compared to the control (Fig. 2e).
Binary, triple, and quaternary combinations of S. thermophilus, L. casei postbiotics, G. acid, and antibiotics significantly reduced IL-1β expression-up to 10-fold-compared to the control, indicating strong synergistic effects (Fig. 2f).
S. thermophilus and L. casei postbiotics reduced IL-8 expression by 10-fold and 2-fold, respectively, while G. acid had no effect. Antibiotic treatment increased IL-8 expression by 2.6-fold (Fig. 2g).
S. thermophilus + L. casei postbiotics reduced IL-8 expression by 2-fold, while other combinations increased it. Notably, combining both postbiotics with antibiotics led to a 7.1-fold increase vs. control (Fig. 2h).
IL-10 expression did not change significantly with antibiotics or L. casei postbiotics but increased 2-fold with G. acid and 3.1-fold with S. thermophilus postbiotics (Fig. 2i).
Double, triple, and quadruple postbiotic combinations significantly increased IL-10 expression. Notably, G. acid + L. casei (3.1-fold) and the full combination with antibiotics (3.2-fold) showed the highest increases vs. control (Fig. 2j).
COX-2 expression decreased 10-fold with S. thermophilus and 1.67-fold with L. casei postbiotics, while antibiotics and G. acid showed no significant change vs. control (Fig. 2k).
All combined postbiotic groups showed significantly reduced COX-2 expression vs. control, except S. thermophilus + G. acid, which showed no change. The antibiotic group showed a 1.6-fold increase. Five-fold reductions were observed in combinations including L. casei postbiotics (Fig. 2l).
FOX-M1 expression decreased significantly with S. thermophilus (2.5-fold) and L. casei (10-fold) postbiotics, while non-significant increases were seen with antibiotics and G. acid (Fig. 2m).
S. thermophilus + G. acid and S. thermophilus + G. acid + L. casei postbiotics significantly reduced FOX-M1 expression by 3.3- and 2.5-fold, respectively, while all other groups, including the standard drug, showed increased expression (Fig. 2n).
IL-33 expression significantly decreased in S. thermophilus (5-fold) and G. acid (10-fold) groups, while the antibiotic group showed no change vs. control (Fig. 2o).
Combined treatments significantly reduced IL-33 expression: 3.3-fold with S. thermophilus + G. acid, and 5-fold with both S. thermophilus + L. casei and the triple combination (Fig. 2p).
S. thermophilus and L. casei postbiotics reduced NF-κB-1 expression 3.3-fold, similar to the antibiotic group, while G. acid led to a 2-fold decrease vs. control (Fig. 2r).
Combined postbiotics significantly inhibited NF-κB expression: S. thermophilus + L. casei showed a 10-fold, and G. acid + L. casei a 5-fold reduction vs. control, indicating strong synergistic effects (Fig. 2s).
H. pylori CFU levels in gastric tissue were quantitatively assessed by culture and reported as log₁₀ CFU/g. The highest bacterial burden was observed in the infected control group (6.14 ± 0.18 log₁₀ CFU/g), whereas the S. thermophilus + L. casei + G. acid + antibiotic (quaternary) group achieved complete eradication with undetectable CFUs (0.00 ± 0.00 log₁₀ CFU/g). The ternary combination group (S. thermophilus + L. casei + antibiotic) showed a mean bacterial load of 1.63 ± 0.09 log₁₀ CFU/g. In contrast, binary combinations exhibited varying levels: S. thermophilus + antibiotic group 2.83 ± 0.17 log₁₀ CFU/g, L. casei + antibiotic group 3.02 ± 0.15 log₁₀ CFU/g, and G. acid + antibiotic group 3.24 ± 0.17 log₁₀ CFU/g. All treatment groups demonstrated statistically significant reductions in bacterial load compared to the infected control (p < 0.01, chi-square test; Fig. 2t). Furthermore, infection rates among groups were statistically analyzed using the chi-square (χ²) test (n = 5 animals per group), and statistical significance was determined by comparing the number of infected and uninfected animals across different treatment groups.
Animal experiments and histopathological results
After H. pylori infection, gastric tissues were collected post-treatment and analyzed via histopathology and culture to assess the presence of H. pylori. All rats in the infection control group had H. pylori, with severe lymphocyte/leukocyte infiltration and marked mucosal-submucosal edema. In contrast, the S. thermophilus postbiotics group showed no immune cell infiltration and only mild submucosal edema, with negative culture results and no H. pylori-related pathology.
H. pylori was detected in 40% of rats in both the G. acid and L. casei groups. G. acid caused moderate lymphocyte and low leukocyte infiltration, while L. casei led to mild lymphocyte infiltration without edema. The antibiotic group had a 20% infection rate with only lymphocyte infiltration.
In the S. thermophilus + G. acid group, H. pylori was detected in 20% of rats, with lymphocyte infiltration but no edema. In contrast, the L. casei + G. acid group showed 60% infection, along with mucosal edema and immune cell infiltration. The G. acid + antibiotic group had 20% infection and lymphocyte infiltration. Triple treatment groups showed better protection and clearer histopathological improvement.
H. pylori was not detected in rats treated with S. thermophilus + L. casei + antibiotics or G. acid. Both groups showed moderate lymphocyte infiltration; the latter also showed mild edema and leukocyte infiltration.
Quadruple treatment groups also achieved complete H. pylori eradication. No edema or leukocyte infiltration was observed, with only mild lymphocyte infiltration present.
Samples from the S. thermophilus postbiotics group showed lymphoplasmacytic infiltration in the mucosal and submucosal layers, mild submucosal edema, and no eosinophil leukocyte infiltration (Fig. 3a).Fig. 3aS.thermophilus’ postbiotics treatment group: Mild edema of the submucosa (stars), H×E. bG.acid treatment group: Lymphoplasmocytic infiltration in the mucosa and submucosa, moderately severe infection, H×E. cL.casei’s postbiotics treatment group: Mild lymphoplasmacytic infiltration in the submucosal region (star), H×E. d Antibiotic treatment group, H×E. eS.thermophilus’ postbiotics plus G.acid treatment group H×E. fS.thermophilus’ postbiotics plus Lactobacillus casei’s postbiotics treatment group: Mild eosinophil leukocyte infiltration in the mucosal and submucosal region (arrows), H×E. gG. acid plus L.casei’s postbiotics treatment group: Mild lymphoplasmacytic infiltration in the submucosa (star, star), H×E. **h) **G.acid plus antibiotic treatment group, H×E
Samples from the G. acid treatment group showed moderate mucosal and submucosal lymphoplasmacytic infiltration, mild eosinophil leukocyte infiltration, and no submucosal edema (Fig. 3b).
In the L. casei postbiotics treatment group, mild lymphoplasmacytic infiltration was observed in the mucosal and submucosal regions, with no eosinophil infiltration or submucosal edema (Fig. 3c).
In the antibiotic treatment group, mild lymphoplasmacytic infiltration was observed in the mucosal and submucosal regions, with no eosinophil infiltration or submucosal edema (Fig. 3d).
No mucosal or submucosal lymphoplasmacytic infiltration, eosinophil infiltration, or edema was observed in the S. thermophilus postbiotics + G. acid treatment group (Fig. 3e).
The S. thermophilus + L. casei group showed mild lymphoplasmacytic and eosinophil infiltration with submucosal edema (Fig. 3f).
In the G. acid + L. casei postbiotics group, mild lymphoplasmacytic infiltration was observed in the mucosal and submucosal regions, with no eosinophil infiltration or submucosal edema (Fig. 3g).
In the G. acid + antibiotic group, mild mucosal and submucosal lymphoplasmacytic infiltration was observed, with no eosinophil infiltration or submucosal edema (Fig. 3h).
In the S. thermophilus + L. casei + antibiotic group, moderate lymphoplasmacytic infiltration was observed in the mucosal and submucosal regions, with no eosinophil infiltration or edema (Fig. 4a).Fig. 4aS.thermophilus’ postbiotics plus L.casei’s postbiotics plus antibiotic treatment group: Moderate lymphoplasmacytic infiltration in the mucosal and submucosal region (star), H×E. bS.thermophilus’ postbiotics plus G.acid plus L.casei’s postbiotics treatment group: Moderate lymphoplasmacytic infiltration in the mucosa (star) and submucosal region (arrows), H×E. **c) **G.acid plus L.casei’s postbiotics plus S.thermophilus’ postbiotics plus Antibiotic treatment group: Moderate lymphoplasmacytic infiltration in the mucosal and submucosal region (star), H×E. d Infection control group. H.pylori colonization (arrows), 100×, H×E.e Negative control group, H×E. f) Negative control group, H×E
In the S. thermophilus + L. casei + G. acid group, moderate lymphoplasmacytic infiltration was observed in the mucosal and submucosal regions, along with mild eosinophil infiltration and submucosal edema (Fig. 4b).
In the S. thermophilus + L. casei + G. acid group, moderate lymphoplasmacytic infiltration, mild eosinophil infiltration, and submucosal edema were observed (Fig. 4c).
In the infection control group, H. pylori colonization was detected with severe lymphocytic infiltration, mild edema, and eosinophil infiltration in the mucosal and submucosal regions (Fig. 4d).
In the negative control group (non-infected), no lymphoplasmacytic infiltration, eosinophil infiltration, or edema was observed in the mucosal and submucosal regions (Fig. 4e and f).
Discussion
Recently, combating drug resistance has focused on preventing antibiotic resistance and enhancing synergistic effects through compound combinations. Natural products, especially postbiotics-bioactive metabolites from probiotics, are widely used for their antimicrobial properties. Their potential to create synergistic effects offers a promising strategy against drug resistance [43–48].
Synergistic effects, where combined compounds exhibit more potent activity than individually, are promising alternatives amid rising antibiotic resistance and treatment challenges [49, 50].
Increasing antimicrobial resistance has boosted interest in natural bioactive compounds, which enhance antimicrobial effectiveness either directly or indirectly by modifying antibiotic resistance [7, 51]. Combinations of natural compounds may alter or block microbial resistance mechanisms, reducing or eliminating antibiotic resistance, thus representing a promising approach [52].
H. pylori infection promotes gastric inflammation by increasing proinflammatory cytokines (TNF-α, IL-1β, IL-6), favoring carcinogenesis. Thus, controlling or inhibiting these cytokines is crucial during treatment [53].
S. thermophilus and L. casei postbiotics individually inhibited TNF-α synthesis, while G. acid and antibiotics alone had no effect. Significant synergistic inhibition occurred only in the G. acid + antibiotic and quadruple (S. thermophilus + L. casei + G. acid + antibiotic) groups, suggesting potential improvement of conventional therapy.
IL-6 is rapidly produced during infections or injuries, but uncontrolled, prolonged IL-6 synthesis may lead to chronic inflammation and autoimmune pathology [54].
In this study, S. thermophilus, L. casei postbiotics, and G. acid significantly reduced IL-6 synthesis compared to control, indicating controlled inflammation, while antibiotics alone had no effect. Strong reductions occurred with combinations of S. thermophilus + G. acid, S. thermophilus + L. casei, and their triple combination. Quadruple combinations including antibiotics further reduced IL-6 expression, suggesting enhanced antibiotic efficacy against H. pylori. Controlling IL-1β, a key inflammatory mediator involved in carcinogenesis, is also crucial.
Elevated IL-1β is linked to cancer progression [55]. In our study, IL-1β expression significantly decreased with S. thermophilus and L. casei postbiotics, while G. acid and antibiotics alone showed no significant effect. Combined treatments showed greater efficacy.
The combined S. thermophilus + L. casei postbiotics and the quadruple combination (S. thermophilus + L. casei + G. acid + antibiotics) showed strong efficacy. G. acid alone did not significantly reduce IL-1β, but its combination with L. casei postbiotics and antibiotics exhibited notable synergistic activity.
IL-8 is overexpressed in H. pylori-infected gastric mucosa and associated with poor gastric cancer prognosis. It has chemotactic properties and promotes cancer cell proliferation and migration [56, 57].
The L.casei’s postbiotics and S.thermophilus’s postbiotics caused inhibition of IL-8 expression, which was significant in the S.thermophilus’s postbiotics treatment group compared to the control. While there was no increase in IL-8 expression in the G. acid treatment group, IL-8 expression was significantly higher in the antibiotic treatment group than in the control group. This situation can be interpreted as the combined antibiotic therapy used in the treatment of H.pylori today may pose a threat to cancer progression by increasing IL-8 expression. One of the most substantial findings in the study was the significant inhibition of IL-8 synthesis in the quadruple combined treatment group (S.thermophilus’s postbiotics plus L.casei’s postbiotics plus G.acid plus antibiotics), as with other cytokines, which was considered very important in creating new treatment protocols.
Alleviating the high inflammatory response in H.pylori infection is directly related to patient well-being and treatment success. Interleukin 10 (IL-10) is an anti-inflammatory cytokine involved in the down-regulation of human immune reactions. IL-8 is effective in reducing the increased inflammatory response in H.pylori infection [58]. We found a significant increase in IL-10 synthesis in the L.casei and antibiotic treatment group compared to the control group. There was a significant increase in IL-10 synthesis in the G.acid and especially S.thermophilus’s postbiotics treatment group. Interestingly, there were significant increases in IL-10 expression in all treatment groups with double, triple, and quadruple combinations of compounds compared to the standard antibiotic treatment group. Strong synergistic activity was found in the increase of IL-10 expression, especially in the quadruple combined treatment group. These findings were considered extremely important in the improvement of new treatment approaches or existing conventional therapy modalities.
COX-2, involved in converting arachidonic acid to prostaglandins, contributes to H. pylori-associated gastric carcinogenesis and is often overexpressed in gastric cancers. Thus, inhibiting COX-2 synthesis is crucial for preventing cancer progression [59].
Antibiotic therapy significantly increased COX-2 expression, potentially predisposing to cancer progression. S. thermophilus postbiotics significantly reduced COX-2 expression, whereas L. casei postbiotics caused a non-significant decrease. All combined treatments, except S. thermophilus + G. acid, significantly inhibited COX-2. Thus, combining these compounds with antibiotics may substantially reduce gastric cancer risk.
FOX-M1, a transcription factor commonly overexpressed in tumors, promotes cancer cell proliferation, migration, and angiogenesis. Thus, inhibiting FOX-M1 synthesis is crucial for preventing cancer progression [60, 61].
Postbiotics from S. thermophilus and L. casei significantly inhibited FOX-M1 expression (Fig. 2m). Combined treatments of S. thermophilus + G. acid and S. thermophilus + L. casei + G. acid also notably suppressed FOX-M1 (Fig. 2n).
Increased levels of IL-33 expression in the gastric mucosa of patients infected with H.pylori are considered a signal of the protumorigenic effect of this cytokine. In the present study, a decrease in IL-33 expression was observed in all groups compared to the control group. The decrease was significant in the S.thermophilus and L.casei’s postbiotics, and G. acid treatment groups. However, there was no significant decrease in the antibiotic treatment group, which was considered active inflammation. We believe this may be due to the lower antibacterial activity than other groups. There was no significant reduction in IL-33 expression in the groups treated with a combination of compounds and antibiotics. The absence of an increase in IL-33 expression in the treatment groups was considered the end of the acute phase of the disease and the transition to the recovery phase.
NF-κB, a proinflammatory pathway activated by H. pylori cagA during gastric carcinogenesis, promotes cytokine release, proliferation, angiogenesis, invasion, and apoptosis inhibition [62]. Chemotherapy-induced cellular stress activates NF-κB, potentially causing drug resistance. NF-κB is also a key factor in inflammation-driven carcinogenesis, including gastrointestinal cancers [63–65].
Postbiotics may exert their effects by regulating the NF-κB pathway. Mechanistically, postbiotic metabolites such as butyrate and lactate have been shown to inhibit NF-κB translocation by modulating IκB kinase activity or acting on upstream pattern recognition receptors such as TLRs. Relevant literature supports their influence on signal transduction pathways leading to suppressed cytokine expression [66–68]. To better explain the observed differences in biological activity between S. thermophilus and L. casei postbiotics, it is essential to consider their distinct compositional profiles. S. thermophilus-derived postbiotics predominantly contain exopolysaccharides, short-chain fatty acids, and specific bacteriocins, which have been associated with immunomodulatory and anti-inflammatory properties [69, 70]. In contrast, L. casei-derived postbiotics are rich in peptidoglycan fragments, lipoteichoic acids, and lactic acid, which play crucial roles in regulating cytokines and enhancing mucosal immunity [71, 72]. These compositional distinctions may explain the observed variations in their individual and combined immunomodulatory and anti-inflammatory effects observed in this study.
Therefore, inhibiting this gene transcription during H. pylori infection is of great importance in preventing cancer progression. NF-κB expression was inhibited in the single and combined treatment groups. Vigorous synergistic activity was detected in the S. thermophilus postbiotics plus L. casei postbiotics combined treatment group. We believe that the therapeutic activities of these compounds and combinations are very substantial in inhibiting NF-κB gene expression, both to prevent drug resistance and to inhibit cancer progression.
In addition to gene expression results, microbial load analysis revealed a significant reduction in H. pylori colonization across all treatment groups. Notably, the quadruple combination group exhibited complete eradication (0.00 ± 0.00 log10 CFU/g), further supporting the therapeutic potential of compound synergy.
After sacrifice, gastric tissues were analyzed microbiologically and histopathologically. Among single-treatment groups, S. thermophilus postbiotics showed complete H. pylori eradication and significant histopathological improvement, characterized by absence of lymphocyte infiltration, low leukocyte infiltration, and mild edema, indicating full recovery.
In the group treated with postbiotics of S.thermophilus, there was a noticeable improvement in both microbiological and histological conditions. This improvement is believed to be due to the regulatory effect of S.thermophilus postbiotics on gene expressions related to inflammation, specifically NF-kB. The expression levels of TNF-alpha, COX-2,* IL-1 beta*,* IL-6*,* IL-33*,* FOX-M1*, and IL-8 were significantly reduced in the S.thermophilus postbiotics group, while the IL-10 expression was significantly increased. We believe the effect is crucial for histopathological recovery, and recovery depends on gene expressions and their inhibition. Notably, this curative effect was not observed in the groups treated with single G.acid, L.casei’s postbiotics, or conventional antibiotics.
In the S. thermophilus postbiotics group, H. pylori was eradicated, unlike other single-treatment groups (G. acid: 40%, L. casei: 40%, antibiotics: 20% isolation rates). Culture findings matched histopathology: lymphoplasmacytic infiltration persisted in the antibiotic group, accompanied by leukocyte infiltration and edema in the L. casei group, and moderate lymphoplasmacytic with mild leukocyte infiltration in the G. acid group.
In combined groups (S. thermophilus + G. acid, S. thermophilus + L. casei, G. acid + antibiotic), H. pylori eradication was incomplete, but histological recovery was notable. The S. thermophilus + G. acid group showed no edema, leukocyte, or lymphocyte infiltration. The absence of edema and leukocyte infiltration in the G. acid + antibiotic group suggests enhanced antibiotic efficacy.
Triple combination therapies (S. thermophilus + L. casei + antibiotic and S. thermophilus + G. acid + antibiotic) were more effective against H. pylori, achieving complete eradication and notable histological improvement. Only moderate lymphocyte infiltration, mild edema, and minor leukocyte infiltration were observed.
Among tested regimens, quaternary combinations (four-component treatments; S. thermophilus + L. casei + G. acid + antibiotics) yielded complete eradication with undetectable CFUs (0.00 ± 0.00 log₁₀ CFU/g) and the most comprehensive histological recovery. Ternary combinations (three-component treatments; S. thermophilus + L. casei + antibiotics) showed significant but partial reductions in bacterial load (1.63 ± 0.09 log₁₀ CFU/g), indicating an intermediate level of efficacy. Binary combinations (two-component treatments: S. thermophilus + antibiotics, L. casei + antibiotics, or G. acid + antibiotics) exhibited variable effects, with CFU levels ranging between 2.8 and 3.1 log₁₀ CFU/g, reflecting modest reductions. This gradient in effectiveness underscores the synergistic value of multi-component therapies against H. pylori, highlighting their potential integration into future therapeutic protocols.
In the quadruple treatment (S. thermophilus + L. casei + G. acid + antibiotic), H. pylori was entirely eradicated, with no leukocyte infiltration or edema and only mild lymphocyte infiltration. Complete eradication and significant histological recovery were also observed in the single S. thermophilus group.
Eradicating H. pylori, a critical risk factor for gastric cancer is essential for cancer prevention. Our study presents a promising new treatment that effectively eliminates this bacterium, potentially preventing gastric cancer development and significantly impacting patient outcomes.
Compared to previous studies on probiotics [13, 33], the postbiotics utilized in this study offer several distinct advantages. While probiotics require live bacterial viability and successful colonization in the gastrointestinal tract to exert their beneficial effects, postbiotics function through their bioactive metabolites, such as short-chain fatty acids, bacteriocins, and polysaccharides, eliminating the dependency on live organisms. This not only enhances safety, especially in immunocompromised hosts, but also ensures greater stability, shelf-life, and batch-to-batch consistency. Moreover, unlike probiotics, whose colonization efficiency may be affected by host microbiota and antibiotic use, postbiotics provide a predictable and controlled biological response, as demonstrated by the consistent anti-inflammatory and anti-Helicobacter pylori effects were observed in both in vitro and in vivo models in this study. These attributes position postbiotics as promising next-generation therapeutic agents for managing H. pylori infection.
Quantitative culture-based CFU analysis strongly aligned with histological improvement. Groups receiving S. thermophilus postbiotics alone or in combination achieved significantly lower bacterial burdens, with full eradication in the quadruple group (S. thermophilus + L. casei + G. acid + antibiotic), indicating that bacterial load can serve as a reliable indicator of treatment efficacy.
G.acid combined with S.thermophilus and L.casei’s postbiotics, G.acid exhibited remarkable effectiveness against H.pylori in the AGS cell line and experimental rat gastritis model. The molecular mechanism underlying this efficacy involves the suppression of pro-inflammatory cytokines and the induction of anti-inflammatory cytokines, leading to the reduction of gastric inflammation and the prevention of H.pylori colonization. The findings of both in-vitro and in-vivo experiments have significant results for the obliteration of H.pylori. These results can cause the eradication of H.pylori and may be a breakthrough for its treatment, one of the most common infectious agents in the world. The success in treating H.pylori, as well as the treatment and prevention of H.pylori-related inflammatory disorders and gastric cancer, may be influential. These findings may have crucial consequences for exploring new treatment methods against H.pylori infections and improving existing treatment methods. However, it should be noted that a limitation of this study was the absence of cytokine protein level measurements in vivo, due to technical limitations in consistently isolating high-quality RNA or cytokine proteins from inflamed gastric tissue.
Supplementary Information
Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aiba Y, Ishikawa H, Tokunaga M, Komatsu Y. Anti-Helicobacter pylori activity of non-living, heat-killed form of lactobacilli including Lactobacillus johnsonii 1088. FEMS Microbiol Lett. 2017;364(11). 10.1093/femsle/fnx 102.10.1093/femsle/fnx 10228505287 · doi ↗ · pubmed ↗
- 2EUCAST. Breakpoint tables for interpretation of MI Cs and zone diameters. 2013; Version 3.1. Available from: http://www.eucast.org/fileadmin/src/media/PD Fs/EUCAST_files/Consultation/EUCAST_clinical_breakpoints_for_Helicobacter_pylori.pdf
- 3Cover TL. Perspectives on methodology for in vitro culture of Helicobacter pylori. Helicobacter Species. 2012;11–5. 10.1007/978-1-62703-005-2_3.10.1007/978-1-62703-005-2_3PMC 392188523015486 · doi ↗ · pubmed ↗