Resolving the taxonomic enigma of Nesocaryum stylosum (Boraginaceae): phylogenetic evidence for its reclassification as Cryptantha stylosa

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
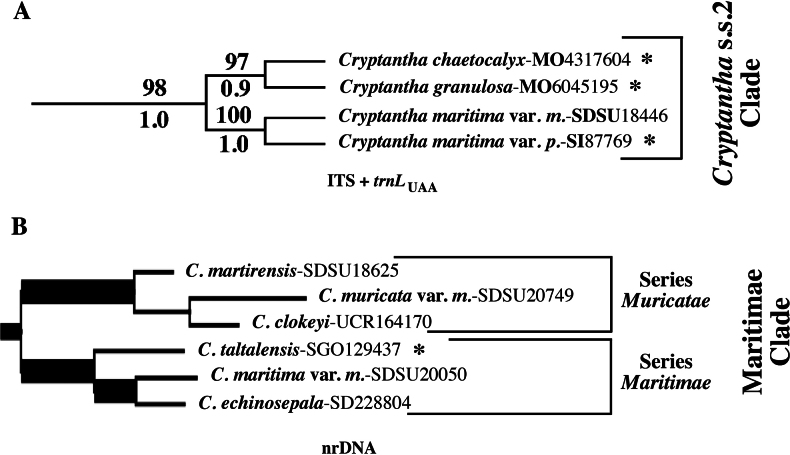 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4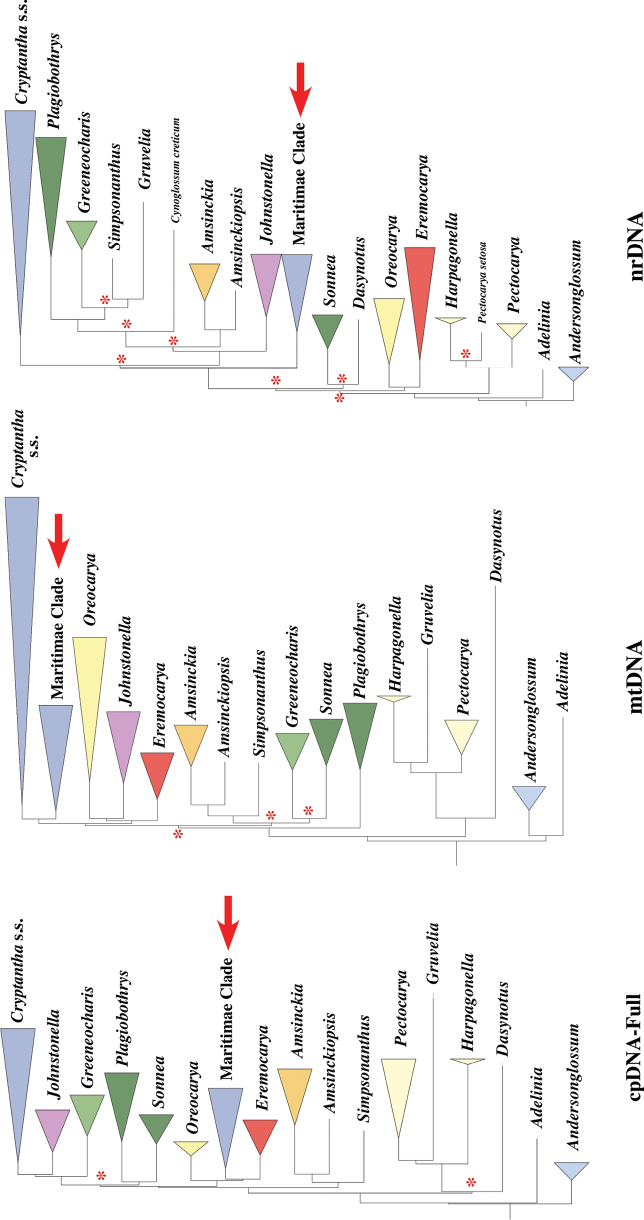 Figure 5
Figure 5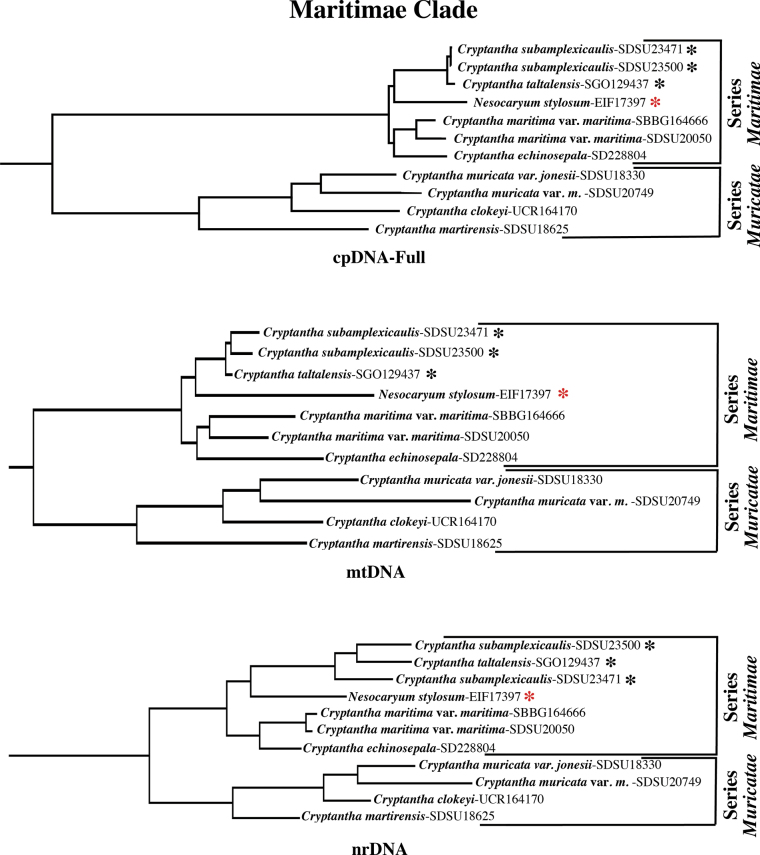 Figure 6
Figure 6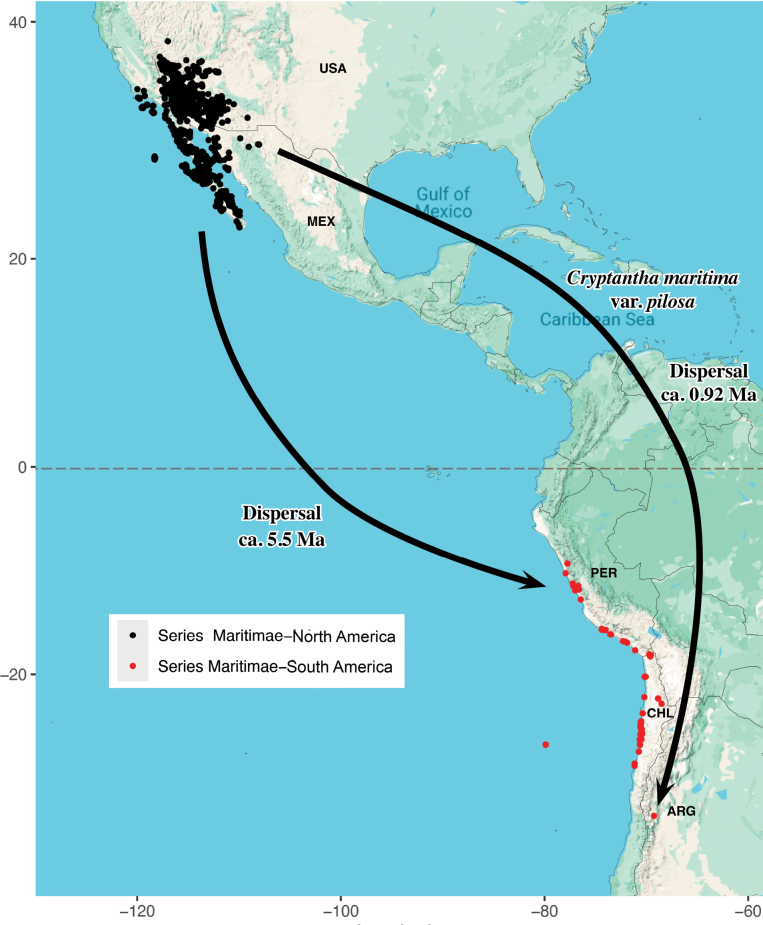 Figure 7
Figure 7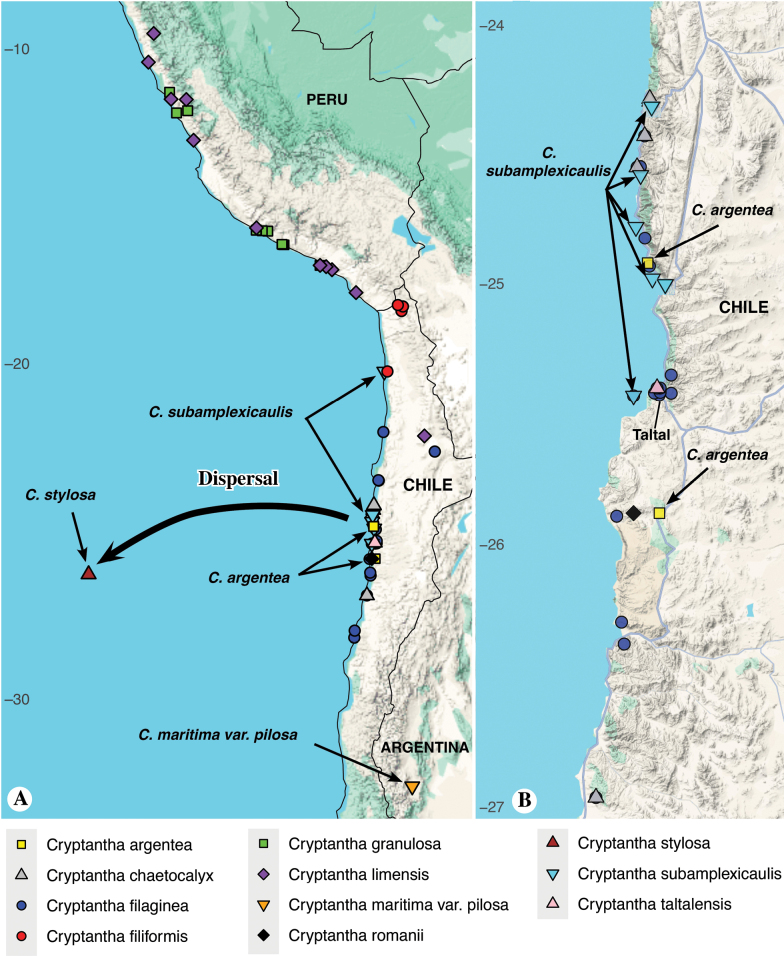 Figure 8
Figure 8|
| ||
| SAm | ||
| SAm | 1,4 | |
| NAm | 2,3,5 | |
| SAm | 4 | |
| SAm | ||
| SAm | 1 | |
| SAm | ||
| NAm | ||
| NAm | 1,2,3,4,5 | |
| NAm & SAm | 1 | |
| NAm | ||
| * | NAm | |
| SAm | ||
| SAm | 5 | |
| SAm | 5 | |
| SAm | 2,3,5 | |
| SAm | ||
|
| ||
| NAm | ||
| NAm | 2,3,5 | |
| ** | NAm | |
| NAm | 2,3,5 | |
| NAm | 2,3,5 | |
| NAm | ||
| NAm | ||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Botanical Research and Applications · Botany, Ecology, and Taxonomy Studies
Introduction
The genus Cryptantha Lehm. ex G.Don (Boraginaceae, subtribe Amsinckiinae, after Chacón et al. 2016), as currently delimited, consists of approximately 111 species and 126 minimum-rank taxa (the latter including the total number of accepted taxa, including varieties and subspecies, after Baldwin 2000), with 65 species native to North America, 47 species native to South America, and one of these species found on both continents (see Hasenstab-Lehman and Simpson 2012, Simpson et al. 2017a, Mabry and Simpson 2018, Amsinckiinae Working Group 2025). Even though the numerous species in the genus may be distinguished from one another by life history (perennial versus annual), stem habit and branching pattern and indument, inflorescence cymule number, leaf morphology, bract presence or absence and morphology, corolla limb width and fornix presence, absence, or color, and fruiting calyx shape, size, and indument, it is the morphology of the unit fruits, termed nutlets (also termed “eremocarps”; see Hilger 2014; Jeiter et al. 2018), that is often most important in delimiting taxa. The nutlets of Cryptantha species vary in number per fruit (1–4), heteromorphism (differences between nutlets within a fruit), size, shape, and sculpturing features, the latter including smooth, papillate, muricate or tuberculate, spinulose, and winged (see Simpson and Hasenstab 2009).
Cryptantha, as it has traditionally been circumscribed, has nutlets that bear a slit-like “ventral groove” positioned above and contiguous with the attachment scar. The molecular phylogenetic analyses of Hasenstab-Lehman and Simpson (2012), Simpson et al. (2017a), and Mabry and Simpson (2018) clarified generic circumscriptions within subtribe Amsinckiinae, to which Cryptantha belongs (see Chacón et al. 2016). These studies provided evidence for the segregation (and resurrection) of four genera that also have a ventral-groove attachment scar – Eremocarya Greene, Greeneocharis Gürke & Harms, Johnstonella Brand, and Oreocarya Greene – from the traditional circumscription of Cryptantha s.l. (note that a ventral groove evolved independently in some species of the genus Amsinckia). However, one additional clade of Cryptantha s.l. is phylogenetically separate from the bulk of Cryptantha taxa and from the four segregate genera mentioned above, a clade first reported by Hasenstab-Lehman and Simpson (2012). In that study, which used one chloroplast region and the nuclear cistron ITS genetic marker, the authors described what they termed the Cryptantha s.s.2 clade (Fig. 1A). The phylogenetic position of the Cryptantha s.s.2 clade varied depending on the type of analysis, ranging from being sister to Cryptantha s.s.1 (consisting of all other examined species in the genus) to being well separated and sister to a larger clade containing four other genera.
Cladograms, redrawn from original publications. A.Cryptantha s.s.2 clade from Hasenstab-Lehman and Simpson (2012), based on ITS and cpDNA data and both analyses; B.Maritimae clade from Simpson et al. (2017a), cistron analysis; chloroplast and mitochondrial analyses from that study are identical in topology. Series Muricatae and Maritimae are designated as in Table 1; see Discussion.
The phylogenetic study of subtribe Amsinckiinae by Simpson et al. (2017a) utilized sequence data from chloroplast DNA (cpDNA), mitochondrial DNA (mtDNA), and nuclear ribosomal DNA (nrDNA). This study also recovered a rogue group of Cryptantha species, which the authors informally termed the “Maritimae clade” (after Cryptantha maritima (Greene) Greene, a member of this clade). The authors interpreted this clade as equivalent to the Cryptantha s.s.2 clade of Hasenstab-Lehman and Simpson (2012) because of the shared membership of C. maritima var. maritima in both, but with expanded membership (Fig. 1B). However, in the Simpson et al. (2017a) analyses, three species corresponding to Johnston’s (1925) series Muricatae form a clade that is sister to a clade corresponding in part to Johnston’s series Maritimae. As in the earlier analysis, the Maritimae clade of Simpson et al. (2017a) also varied in phylogenetic position relative to other clades in subtribe Amsinckiinae depending on which of the three data types were analyzed; it is sister to Eremocarya with weak to mixed support in the mitochondrial and chloroplast DNA analyses, but sister to the remainder of the genus, i.e., Cryptantha s.s., with mixed support in the nuclear ribosomal (ITS) analysis.
Another member of Boraginaceae found in South America, the monotypic Nesocaryum stylosum (Phil.) I.M.Johnst., has puzzled taxonomists for nearly a century. This species is endemic to Isla San Ambrosio, a small (ca. 3.7 km^2^), rugged, uninhabited island located 900 km due west of the coast of central Chile (Fig. 2A, B). This plant species was originally described by Philippi (1870) as Heliotropium stylosum Phil. (see Appendix 1 for the protologue description). The species resembles many Heliotropium species in being a shrub with clustered (corymbose) white flowers (Figs 2C–F, 3A, B, D). Leaf surfaces and calyces are covered with whitish, densely strigose indument (Figs 2F, 3C, E, 4A). The epithet stylosum undoubtedly refers to the unique (“singularem,” in Philippi’s description) elongate style, described as twice as long as the ovary in the protologue (Appendix 1).
A, B. Map of Isla San Ambrosio, Chile, showing observations of Nesocaryum stylosum. A. Close-up satellite image showing the island and four localities of iNaturalist observations by Lukas Mekis (see Appendix 5); map from Google 2024, imagery 2024 Airbus, Maxar Technologies; B. Map showing the position of the island relative to South America, ca. 900 km west of the coast of Chile; map data Google 2024; C–F. Posted images of iNaturalist observations by Lukas Mekis, lukasmekis, some rights reserved (CC BY-NC); see Appendix 5 for full data: C. Whole plant, a small shrub (17 December 2021); D. Close-up of C (17 December 2021); E. Whole plant showing corymbose clusters of white flowers (17 December 2021); F. Close-up of vegetative shoots showing narrowly elliptic, gray-green, sericeous leaves (31 March 2022).
Nesocaryum stylosum herbarium sheet images. A. Holotype, E. [Enrique] Simpson s.n., 1869 (barcode SGO000004198 = accession SGO054406), showing a single whole plant; note woody stem; B–E. Specimen, N. Bahamande N. s.n., 21 August 1960 (accession SGO122886): B. One of several whole plants; C. Close-up of linear leaves; D. Inflorescence composed of two cymules arranged in a corymbose secondary unit; E. Developing fruits; note whitish, sericeous vestiture. All images were taken by Michael G. Simpson at the SGO herbarium, by permission.
Nesocaryum stylosum, material from specimen N. Bahamande N. s.n., 21 August 1960 (accession SGO122886). A. Close-up of fruiting calyces and three nutlets from one fruit; B. Fruiting calyx opened, showing two nutlets within; C. Nutlet close-up, shown (left to right) in dorsal, ventral, and lateral views; note smaller papillae and larger, rounded, whitish tubercles. Arrow indicates slightly developed expansion of the apical tip of the ventral groove. All images were taken by Lee M. Simpson, San Diego State University; material removed from specimens at the SGO herbarium, by permission.
In 1927, Johnston transferred this species of Heliotropium to its own genus, which he named Nesocaryum I.M.Johnst. (after neso-, island + caryum, fruit) (see Appendix 2 for the protologue description). Johnston (1927: 75) stated that N. stylosum was “evidently derived from Cryptantha,” given that the fruit (Fig. 4C) was “indistinguishable” from members of that genus. However, he placed the species in a monotypic genus because of its unique, even within Boraginaceae, calyx (Fig. 4A, B) and corolla morphology. The calyx is described as cylindrical with an elongate, abaxial adnate bract, and the corolla tube has “5 pairs of knife-like lamellae” at the base. Johnston considered Nesocaryum stylosum to be a derivative of a member of Cryptantha sect. Krynitzkia ser. Barbigerae (see Discussion).
In light of the long-standing uncertainty surrounding the affinities of Nesocaryum stylosum, the primary objective of this study is to analyze sequence data to evaluate its phylogenetic position and biogeographic history. We first test whether the species does indeed nest within the genus Cryptantha, as implied by nutlet morphology, or within another group of subtribe Amsinckiinae of Boraginaceae. If nested within Cryptantha, we then ask which taxa represent its closest relatives. Is Nesocaryum stylosum closely related to members of sect. Krynitzkia ser. Barbigerae, as suggested by Johnston (1927)? An overarching goal of this study is to evaluate the relationship of Nesocaryum stylosum to other members of this complex of plants and to address strongly supported results by updating the taxonomy of this species.
Methods
Sample collection and DNA extraction
Obtaining suitable material for DNA extraction from Nesocaryum proved challenging. An attempt to extract DNA from a specimen collected in 1960 was unsuccessful. However, using more recently collected material (Christian López Chamorro s.n., September 2018; EIF17397), we successfully extracted and sequenced DNA. Including Nesocaryum, we sequenced 94 new samples in this study across the Amsinckiinae. These new samples included several taxa not previously sequenced in Simpson et al. (2017a), including new members of the Maritimae clade. We relied on material available to us from herbarium specimens and silica-dried leaves from live material. To further expand these data, we downloaded sequence data for 142 samples from the Sequence Read Archive (SRA), representing data from Simpson et al. (2017a). These combined datasets were used for phylogenetic analysis, building upon the sequence data of Simpson et al. (2017a). Documentation of the new samples used for sequencing is provided in Appendix 3.
For DNA extraction, dried silica material was ground into a fine powder using a multi-sample Bead Beater tissue homogenizer and extracted using a modified cetyltrimethylammonium bromide (CTAB) protocol with the following modification: incubation in CTAB extraction buffer with proteinase K at 65 °C for 3–4 h, followed by overnight precipitations. Half of the samples underwent a final cleaning step using a Zymo DNA Clean and Concentrator-25 kit (Zymo Research, Irvine, CA, USA). DNA extractions were quantified on a Qubit fluorometer using the Qubit Double-Stranded High Sensitivity Assay Kit (Invitrogen, Carlsbad, CA, USA) to assess genomic DNA quantity. DNA quality was evaluated by visualization on an agarose gel following electrophoresis.
Library preparation, size selection, library quantification, and sequencing
A run of 94 samples was performed at the University of California at Riverside Genomics Core in a single lane on an Illumina NextSeq2000 platform, yielding an average of 1,275,636 reads per sample (range: 849–4,040,175). This produced single-end 100 bp reads with an average insert size of 250 bp. All reads are deposited in the Short Read Archive of the NCBI (https://www.ncbi.nlm.nih.gov; see Appendix 3). Using high-throughput genome skimming sequencing (see Ripma et al. 2014), we obtained chloroplast, cistron, and mitochondrial genome data.
Read recovery varied substantially among samples, likely due to differences in DNA input concentrations across libraries. In addition, many sequences were fragmentary, possibly reflecting the degraded nature of DNA from older herbarium specimens (Appendix 3).
Data analysis
We combined the sequence results of this study with those of Simpson et al. (2017a), yielding an initial total of 236 samples. One specimen of Cryptantha taltalensis I.M.Johnst. (SGO129437), included in the combined analyses, had previously been identified as C. subamplexicaulis (Phil.) Reiche in Simpson et al. (2017a) and Mabry and Simpson (2018); see Appendix 3. This name change resulted from a subsequent study of original material from that specimen and annotation by Pablo Moroni and Michael Simpson.
Raw sequencing reads were processed using CAPTUS (v1.4.8; Ortiz et al. 2023; https://github.com/edgardomortiz/Captus.git) to perform data cleaning, de novo read assembly, and targeted locus extraction. Initial quality control and adapter trimming were conducted using captus clean. Cleaned reads were subsequently assembled de novo using captus assemble. Following assembly, target regions of interest were extracted from assembled contigs using captus extract. This step employed a paralog tolerance of 1.5 and included mitochondrial targets specified by the parameter SeedPlantsMIT, as well as a custom full-chloroplast target derived from Echium plantagineum L. (Boraginaceae; Carvalho Leonardo et al. 2023), divided into 21 fragments designed to avoid breaks across gene boundaries. We also created a custom target for the cistron using assembly statistics to identify long contigs potentially spanning the complete region. For the sample Cryptantha decipiens (M.E.Jones) A.Heller (voucher SDSU20014 of Simpson et al. 2017a), a custom Python script was used to extract the cistron region from the contig NODE_57793_length_8014_cov_228.7687_k_95_flag_1, spanning positions 672–6471. Because this region was located on the negative strand, the extracted sequence was reverse-complemented. The resulting loci were subsequently aligned using captus align, which generated alignments for each marker type. For all analyses, we used the trimmed, naïve alignments produced by CAPTUS to reduce ambiguous regions and improve alignment quality. The 20 fragments of the full chloroplast genome and the 29 mitochondrial gene alignments were concatenated. Interactive HTML reports summarizing quality metrics and statistics for each step of data processing – cleaning, assembly, locus extraction, and alignment – along with target and tree files, have been deposited in Zenodo (https://doi.org/10.5281/zenodo.17246597).
Phylogenetic analyses were performed using IQ-TREE (v2.2.2.7; Minh et al. 2020). Maximum likelihood (ML) trees were inferred separately for the cistron, the full chloroplast genome, and concatenated mitochondrial genes. For the cistron alignment, IQ-TREE was run with the command-line options -nt AUTO to determine the optimal number of threads, -B 1000 to generate 1,000 ultrafast bootstrap replicates for node support, and -m MFP to enable ModelFinder Plus for automatic model selection. Analyses of the full chloroplast genome and concatenated mitochondrial genes were conducted under partitioned models. For these datasets, IQ-TREE was run with the alignment file in Nexus format and an accompanying partition file specifying gene or region boundaries. Commands included -nt AUTO, -B 1000 for ultrafast bootstrap support, —alrt 1000 for 1,000 SH-like approximate likelihood ratio test replicates, and -m MFP+MERGE to allow model selection with automatic merging of partitions based on similarity. Samples were excluded from phylogenetic analyses if they had 10 or fewer loci in the mitochondrial gene dataset or fewer than 10 recovered regions in the full chloroplast genome assembly (see Appendix 3). All scripts and code used for data processing and phylogenetic analyses are available in a public GitHub repository (https://github.com/mmabry/Nesocaryum_Boraginaceae.git). All trees were rooted using sequence data from the outgroup Myosotis laxa Lehm. (SBBG132391; see Simpson et al. 2017a).
Distribution mapping
To generate distribution maps, georeferenced data points for all members of the series Maritimae (see below) were downloaded from the Global Biodiversity Information Facility (GBIF.org 2024a–p). A total of 5,518 GBIF records were mapped and are listed by series and species in Appendix 4. These records were not taxonomically verified by the authors. In addition, iNaturalist (https://www.inaturalist.org) observations of Nesocaryum stylosum (Appendix 5) were mapped in detail. Maps were generated using R v4.3.1, with duplicate records removed and occurrence points plotted on a custom Google Map (Kahle and Wickham 2013).
Results
The three phylogenetic analyses, using sequences from the full chloroplast (cpDNA), mitochondria (mtDNA), and cistron (nrDNA), recovered the same genera and clades identified by Simpson et al. (2017a), including the Maritimae clade, all of which were individually well supported, with 100% bootstrap support values (Fig. 5). The only exception was the nrDNA analysis, in which Pectocarya is not monophyletic (with P. setosa A.Gray more closely related to Harpagonella A.Gray), Cynoglossum creticum Mill. is nested within the Amsinckiinae (being outside the subtribe in the other two analyses), and not all genera and clades are well supported. These nrDNA results mirror those of Simpson et al. (2017a), with interrelationships among genera and clades varying depending on the sequence data used. Thus, the backbone of the Amsinckiinae is weak in terms of bootstrap support values (Fig. 5). The full cladograms for each of the three sequence datasets are shown in Suppl. materials 1–3.
Cladograms of subtribe Amsinckiinae using the three DNA sequence datasets of this study: chloroplast (cpDNA), mitochondrial (mtDNA), and cistron (nrDNA). Genera and clades are represented by colored triangles, all robustly supported except for some taxa in the nrDNA analysis. Relatively short branches and weak support along the backbone of these analyses are indicated, with an asterisk representing nodal bootstrap support values <90%. Note differences in interrelationships among major clades and genera. The Maritimae clade (red arrows) is sister to Cryptantha s.s. only in the mtDNA analysis. All trees were rooted using sequence data from the outgroup Myosotis laxa Lehm. (SBBG132391).
This variability in genus and clade interrelationships is exemplified by the Maritimae clade, whose topological position shifts among the three analyses. The Maritimae clade is sister to Cryptantha s.s. in the mtDNA analysis but considerably more distant from Cryptantha s.s. in the cpDNA analysis, in which it is sister to Eremocarya with good support but a shallow branch, and in the nrDNA analysis, in which it is sister to a complex of genera with weak support (Fig. 5). Interestingly, in the analyses of Simpson et al. (2017a), the Maritimae clade is sister to Cryptantha s.s. only in the nrDNA analysis, not in the mtDNA analysis as observed here.
In all three analyses, however, the Maritimae clade comprises the same nine species and 10 minimum-rank taxa, all of which exhibit the same topology except for Cryptantha subamplexicaulis and C. taltalensis, which show a slightly different topology in the nrDNA analysis (Fig. 6). The Maritimae clade consistently consists of two sister clades (Fig. 6), which correspond to the modified series Maritimae and series Muricatae of Johnston (1925). Nesocaryum stylosum is part of series Maritimae (Fig. 6), where it is sister to the South American species Cryptantha subamplexicaulis and C. taltalensis. These three species are sister to C. maritima var. maritima (two samples) and C. echinosepala J.F.Macbr. from North America. Series Maritimae is sister to a clade composed of Cryptantha muricata (Hook. & Arn.) A.Nelson & J.F.Macbr., C. clokeyi I.M.Johnst., and C. martirensis M.G.Simpson & Rebman, this clade corresponding to the series Muricatae (Fig. 6; see Discussion).
*Phylogeny of the Maritimae clade in each of the three sequence analyses. Composition is identical across analyses, and topology is the same except for relationships of C. taltalensis and the two samples of C. subamplexicaulis, which differ in the nrDNA analysis. Cryptantha series Maritimae and series Muricatae are modified from Johnston (1925). =South American taxa. All nodes have maximal bootstrap support. Abbreviations of DNA sequence types are as in Fig. 5.
Given these sequence data and evidence from nutlet morphology, our results strongly support Nesocaryum stylosum as phylogenetically nested within series Maritimae of the Maritimae clade. In previous and current analyses, the Maritimae clade is either sister to Cryptantha s.s., representing the remainder of the genus, or more distantly related. We considered transferring the members of the Maritimae clade to another genus, but given the conflicting results, we preferred to defer this decision until additional sequence data—particularly nuclear DNA data—are available to better resolve the backbone relationships among clades of the Amsinckiinae. Thus, at this point, we consider it prudent to transfer Nesocaryum stylosum to the genus Cryptantha, given that: (1) the interrelationships of the Maritimae clade with other clades and genera vary depending on the sequence data analyzed; (2) the Maritimae clade is sister to Cryptantha s.s. in one analysis in the present study and in a separate analysis by Simpson et al. (2017a); (3) all other members of the Maritimae clade are currently classified in the genus Cryptantha (see Discussion); (4) the nutlet attachment scar of N. stylosum conforms to that of Cryptantha taxa; and (5) many members of the Maritimae clade still lack sequence data. We also note that members of the Maritimae clade lack an obvious morphological synapomorphy, other than a possibly elongate style, which is also present in some members of Cryptantha s.s.
Based on the total evidence and these considerations, we formally transfer Nesocaryum stylosum to Cryptantha as follows:
Taxonomic treatment
Cryptantha
stylosa
Taxon classificationPlantaeBoraginalesBoraginaceae
(Phil.) Scherson, Nic.García & M.G.Simpson comb. nov.
19D45778-6973-524A-90AB-732214EFDED3
Heliotropium stylosum Phil., Bot. Zeitung (Berlin) 28: 500 (1870). Basionym.
Type.
Chile. Isla San Ambrosio, 1869, Enrique Simpson s.n. Holotype: barcode SGO000004198!=accession SGO054406!
Discussion
As stated earlier, we confirm that this unusual plant is a member of Cryptantha, series Maritimae of the Maritimae clade of Simpson et al. (2017a) and Mabry and Simpson (2018) (Figs 5, 6). If future sequence data convincingly place the Maritimae clade apart from Cryptantha s.s., recognition of a separate genus for the clade may be warranted. For now, however, we retain the members of the Maritimae clade within the genus Cryptantha, treating this species as C. stylosa.
As noted above, the Maritimae clade in this study and in those of Simpson et al. (2017a) and Mabry and Simpson (2018) consists of two sister clades. One of these corresponds to the series Muricatae of Johnston (1925; Figs 1B, 6; Table 1). Johnston recognized only Cryptantha muricata (including its three varieties) in that series. We add four additional species to series Muricatae, resulting in a current composition of five species and seven minimum-rank taxa (Table 1; after Simpson et al. 2024). All of these taxa are morphologically similar in having ovoid fruiting calyces, ovate, papillate, and tuberculate to muricate nutlets, and a style extending to or beyond the nutlet apices (Simpson et al. 2024: Figs 4, 5).
**Table 1.: Taxa belonging to the Maritimae clade under the present classification, with eight species and 13 minimum-rank taxa in North America (NAm) and 12 species and minimum-rank taxa in South America (SAm), for a total of 19 species and 24 minimum-rank taxa (Cryptantha maritima var. pilosa occurs on both continents). *=Tentative placement in series after Simpson and Rebman (2021); =tentative placement in series after Simpson et al. (2024). Taxa are grouped within series Maritimae and Muricatae, as circumscribed here. Indicated are taxa sequenced or analyzed in molecular phylogenetic studies of: 1 = Hasenstab-Lehman and Simpson (2012); 2 = Simpson et al. (2017a); 3 = Mabry and Simpson (2018); 4 = Böhnert, Borda Tamayo, Mabry, Schwarzer, Simpson, and Weigend (unpublished data); 5 = this paper.
The second clade within the Maritimae clade corresponds in part to the series Maritimae of Johnston (1925). Johnston recognized five species in this series: Cryptantha dumetorum (Greene ex A.Gray) Greene, C. echinosepala, C. maritima, C. micromeres (A.Gray) Greene, and C. recurvata Coville. Of these five species, the current study and those of Simpson et al. (2017a) and Mabry and Simpson (2018) support only two – C. echinosepala and C. maritima – as belonging to the series Maritimae (see Simpson and Rebman 2021); the other three species are more closely related to taxa or clades of Cryptantha s.s., which comprises the bulk of the genus (Simpson et al. 2017a; Mabry and Simpson 2018). Based on morphological similarities, Simpson and Rebman (2021) provisionally added one additional North American species – Cryptantha pondii Greene – and included all varieties of C. maritima (Table 1).
The molecular phylogenetic studies of Hasenstab-Lehman and Simpson (2012), Simpson et al. (2017a), Mabry and Simpson (2018), Böhnert, Borda Tamayo, Mabry, Schwarzer, Simpson, and Weigend (unpublished data), and the present study provide phylogenetic evidence for the inclusion of seven additional South American species in series Maritimae (see list in Table 1, including the variety of C. maritima occurring on both continents). With respect to C. stylosa, although Johnston (1927) recognized the similarity of its nutlets to those of Cryptantha, he chose to classify the species in a monotypic genus based on its unique habit, a shrub, and its distinctive calyx morphology, described as cylindrical, mostly synsepalous, with a ribbed tube and a linear, adnate floral bract. We interpret these features as autapomorphies of Cryptantha stylosa.
Ongoing morphological observations, based on nutlet sculpturing, nutlet attachment scar, and style length, allow us to provisionally add five additional South American species to the series Maritimae, bringing the preliminary total across the two continents to 14 species and 17 minimum-rank taxa (Table 1). A more detailed revision of the infrageneric classification of Cryptantha is currently underway [Simpson, Moroni, and Mabry (unpublished data)].
Biogeographic history
The known distributions of members of series Maritimae are shown in Figs 7, 8 and exhibit a classic American amphitropic disjunct pattern (Guilliams et al. 2017; Simpson et al. 2017b). Although sequence data are lacking for many members of the series, the work of Guilliams et al. (2017) supports two past long-distance dispersal events within the series, both from western North America to western South America (Fig. 7). One dispersal event involved Cryptantha maritima var. pilosa I.M.Johnst., which is widespread in extreme southwestern North America, occurring in the Baja California Peninsula and Sonora, Mexico, as well as southeastern California, southern Nevada, and southern Arizona (see Simpson and Rebman 2021). The single known locality of this variety in northwestern Argentina (GBIF.org 2024i; see Figs 7, 8A) is estimated to have resulted from dispersal from North America approximately 0.92 million years ago (Guilliams et al. 2017).
Distribution map of members of Cryptantha series Maritimae in North and South America, based on GBIF data [GBIF.org (2024a–p); see References], representing a total of 5,518 GBIF records; see Appendix 4 for listings by species and series. Two separate long-distance dispersal events and estimated times of dispersal are indicated (after Guilliams et al. 2017).
A, B. Close-ups of the distribution map showing localities of South American members of Cryptantha series Maritimae. A. Possible dispersal event from coastal Chile to Isla San Ambrosio; B. Larger-scale view of coastal Chile. Localities of Cryptantha stylosa and the putative close relatives C. argentea and C. subamplexicaulis are labeled.
A second, independent dispersal event within the series Maritimae is estimated to have occurred approximately 5.53 million years ago (Fig. 7). This estimate is approximate, as it is based on sequence data from only three species. Our working hypothesis is that this dispersal event led to a radiation in South America, culminating in the 11 species of the series, aside from C. maritima var. pilosa, that are currently recognized in that region. We further hypothesize that C. stylosa evolved following a single westward dispersal event across the Pacific Ocean from an ancestral mainland population (Fig. 8A; see below).
With the exception of the Andean C. maritima var. pilosa and the insular Cryptantha stylosa, the South American species of series Maritimae occur primarily in coastal regions from central Peru to central Chile, with a few populations recorded at higher elevations (Figs 7, 8). These coastal regions are relatively dry and receive little annual rainfall. However, they do receive precipitation via condensation of coastal fog, allowing coastal plant communities to persist (Moreira-Muñoz 2011).
Taxonomic relationships of C.
stylosa within series Maritimae
Given that eight of the 17 taxa of the Maritimae clade remain unsequenced, we currently lack a sufficient sample size to evaluate the phylogenetic relationships of Cryptantha stylosa to all other members of series Maritimae; however, support values within series Maritimae for the taxa that were sequenced are all above 90%. Based on morphology, we hypothesize that two other members of the series are its possible closest relatives: C. argentea I.M.Johnst. and C. subamplexicaulis. Both of these taxa, like C. stylosa and unlike other members of series Maritimae, are perennial. Cryptantha subamplexicaulis is described as “perennial or persistent annual with a firm branching root, erect and subsimple at first but later with several or many trailing branches from a loose fruticulose caudex and forming a loose prostrate mat 3–12 dm broad and 1–2 dm tall, hispid or hispid-villous and usually appressedly canescent” (Johnston 1927: 41). Cryptantha argentea is described as “perennial, prostrate, densely silvery-striated, with very numerous ramified branches from a prostrate, densely branched, shrubby stem rising into a cushion 2–5 cm high” and as “notable for its silvery, very dense strigose pubescence and particularly because of its very densely pulvinate [cushion-like] habit. The root is strong, branched, and indubitably perennial” (Johnston 1927: 42–43). We note that C. argentea was sequenced but was not included in any of the three analyses because of insufficient sequence data quality; samples were excluded from phylogenetic analyses if they had fewer than 60 loci recovered for the chloroplast gene dataset, 10 or fewer loci for the mitochondrial gene dataset, or fewer than 10 recovered regions in the full chloroplast genome assembly (see Appendix 3).
Cryptantha stylosa, as noted earlier, is also perennial but differs from both species in having an erect, woody, shrubby habit. Cryptantha stylosa resembles C. subamplexicaulis in having relatively large corollas (limb 6–7 mm wide in the former, 5–6 mm wide in the latter). However, it more closely resembles C. argentea in vestiture, with both species having a densely silvery-strigose stem and leaf indumentum. Cryptantha stylosa resembles both of these species, and all other South American members of series Maritimae, in having an elongate style that protrudes well beyond the nutlet apices; however, this character also occurs in some members of series Muricatae.
The woody, shrubby habit of C. stylosa represents a derived condition within the series Maritimae and is unique within the genus Cryptantha. Five other South American species of Cryptantha are perennial, but these are herbaceous and evolved independently, being members of distantly related clades of Johnston’s sections Eucryptantha and Geocarya, which are nested within Cryptantha s.s. (Johnston 1927; see Mabry and Simpson 2018). We interpret the unique habit of C. stylosa as an example of the evolution of “gigantism” following dispersal to an island, a pattern commonly involving evolutionary transitions to woody perennials (Carlquist 1966). Examples of this phenomenon include, within Boraginaceae, Echium in the Cape Verde Archipelago (Romeiras et al. 2011) and Selkirkia in the Juan Fernández Islands (Holstein et al. 2016), and, within Asteraceae, the Hawaiian silversword alliance – Argyroxiphium, Dubautia, and Wilkesia (Baldwin 2003).
Conclusion
Based on the evidence presented here, Nesocaryum stylosum is placed in the genus Cryptantha as the new combination C. stylosa and is best classified within a revised and expanded series Maritimae of the Maritimae clade. Members of this series appear to have radiated in South America following two independent long-distance dispersal events: an early event resulting in diversification into 11 species of Cryptantha and a separate, relatively recent event involving C. maritima var. pilosa. The occurrence of Cryptantha stylosa on Isla San Ambrosio may represent yet another dispersal event from coastal Chile to the island, possibly involving ancestors related to the similarly perennial C. argentea and C. subamplexicaulis, although this hypothesis requires verification. Following dispersal to the island, the ancestor of C. stylosa underwent evolutionary changes in morphology, particularly the acquisition of a shrubby, woody habit that is unique within the genus and subtribe Amsinckiinae.
Although the current study provides a preliminary assessment of the composition and interrelationships of series Maritimae, additional work is needed to resolve this complex. Five South American species and three North American taxa remain unsequenced, and some South American samples were of limited quality. A more detailed molecular phylogenetic analysis of the group is required. Aside from Cryptantha maritima, the South American members of series Maritimae still require basic taxonomic study, including additional field collections and detailed analyses of morphological variation. Of the 14 species and 17 total taxa currently recognized in series Maritimae, only C. maritima var. pilosa has a published chromosome count (Las Peñas 2005); this count, 2n = 20, is aberrant within the genus. All other North American species have a chromosome number of 2n = 24, whereas non-hexaploid South American members of the genus have a chromosome number of 2n = 14 (see Amsinckiinae Working Group 2025). The reproductive biology and chemistry of members of the series remain unknown, with reproductive biology being of particular interest in understanding the adaptations of C. stylosa following its dispersal to an isolated island.
Future work incorporating expanded nuclear DNA sequence data will help determine whether the Maritimae clade is sister to Cryptantha s.s. and whether it should be recognized as a separate genus. Such studies will also provide the basis for a formal classification of the Maritimae clade and its infrageneric structure within the genus Cryptantha.
Supplementary Material
XML Treatment for Cryptantha stylosa
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amsinckiinae Working Group (2025) Systematics of Amsinckiinae (Boraginaceae): The Popcornflowers. https://plants.sdsu.edu/amsinckiinae [accessed 01.06.2025]
- 2Baldwin BG (2000) Roles for modern plant systematics in discovery and conservation of fine-scale biodiversity.Madroño 47: 219–229.
- 3Baldwin BG (2003) A phylogenetic perspective on the origin and evolution of Madiinae. In: Carlquist S Baldwin BG Carr GD (Eds) Tarweeds & Silverswords: Evolution of the Madiinae (Asteraceae).Missouri Botanical Garden Press, St. Louis, 193–228.
- 4Carlquist S (1966) The biota of long-distance dispersal. IV. Genetic systems in the floras of oceanic islands.Evolution; International Journal of Organic Evolution 20: 433–455. 10.2307/240658328562902 · doi ↗ · pubmed ↗
- 5Carvalho Leonardo I Alberti A Denoeud F Barreto Crespo MT Capelo J Bustos Gaspar F (2023) The complete plastome of Glandora prostrata subsp. lusitanica (Samp.) DC Thomas (Boraginaceae), the first chloroplast genome belonging to the Glandora genus. Mitochondrial DNA.Part B, Resources 8(2): 270–273. 10.1080/23802359.2023.2175976 PMC 993700836816053 · doi ↗ · pubmed ↗
- 6Chacón J Luebert F Hilger HH Ovcinnikova S Selvi F Cecchi L Guilliams CM Hasenstab-Lehman K SutorýK Simpson MG Weigend M (2016) The borage family (Boraginaceae s.str.): A revised infrafamilial classification based on new phylogenetic evidence, with emphasis on the placement of some enigmatic genera.Taxon 65: 523–546. 10.12705/653.6 · doi ↗
- 7GBIF.org (2024 a) GBIF Occurrence Download: Cryptantha argentea. 10.15468/dl.rxfvwz [accessed 16.07.2024] · doi ↗
- 8GBIF.org (2024 b) GBIF Occurrence Download: Cryptantha chaetocalyx. 10.15468/dl.h 3z 987 [accessed 16.07.2024] · doi ↗