Carbon and Nitrogen Stable Isotopic Profiling of Chimpanzees and Monkeys in Kalinzu Forest Reserve, Uganda
Takumi Tsutaya, Natsumi Aruga, Naoto F. Ishikawa, Yoko Sasaki, Haruka Kitayama, Minoru Yoneda, Nana O. Ogawa, Naohiko Ohkouchi, Chie Hashimoto

TL;DR
This study uses stable isotope analysis to compare the diets of chimpanzees and monkeys in Uganda, revealing unexpected dietary patterns and variability.
Contribution
The study introduces intra-individual dietary variability in chimpanzees using ultra-fine hair sectioning and compares their diet to sympatric primates.
Findings
Chimpanzees in Kalinzu have lower faunivory than sympatric monkeys based on isotope analysis.
Carbon and nitrogen isotope ratios in a chimpanzee's hair fluctuated by more than 1‰ within 10 days.
Plant foods and chimpanzees in Kalinzu showed lower carbon isotope ratios than expected for the region's rainfall.
Abstract
Stable isotope analysis is a widely used tool in primate ecology for investigating diet and environment, with numerous studies focusing on chimpanzees. However, few studies have used this method to explore the dietary niche of chimpanzees in comparison to other primates or examined intra‐individual dietary variability. This limitation hinders the understanding of the comparability of stable isotopic data with the wealth of behavioral observational data in primate ecology. We report the carbon and nitrogen stable isotope ratios of hairs from wild eastern chimpanzees (Pan troglodytes schweinfurthii) and four other primate species (Cercopithecus mitis, Cercopithecus ascanius, Allochrocebus lhoesti, and Colobus guereza) in the Kalinzu Forest Reserve, Uganda. Bulk analysis revealed that both plant foods and chimpanzees in Kalinzu exhibited lower carbon stable isotope ratios than expected for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
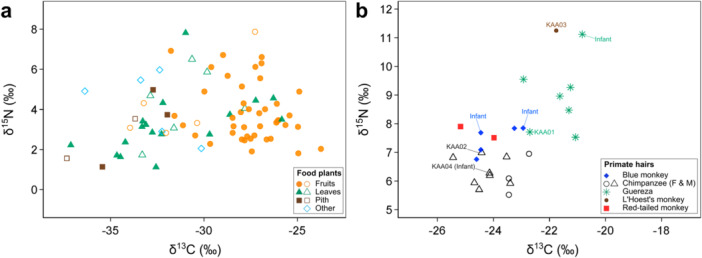 Figure 1
Figure 1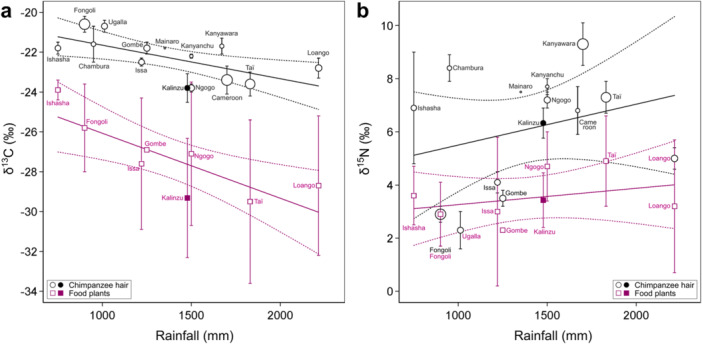 Figure 2
Figure 2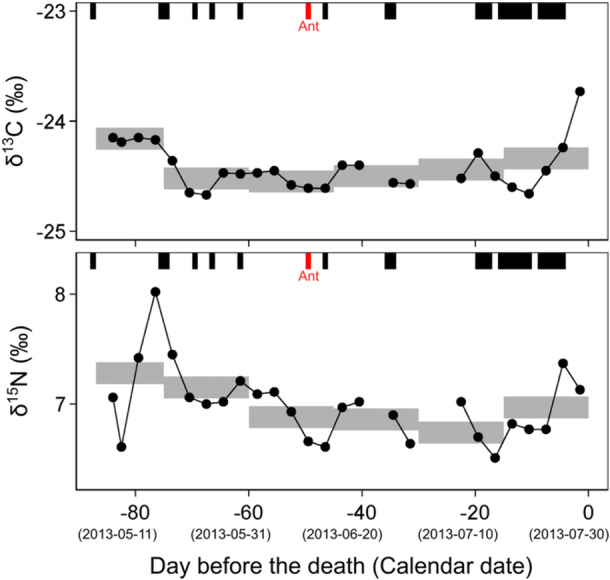 Figure 3
Figure 3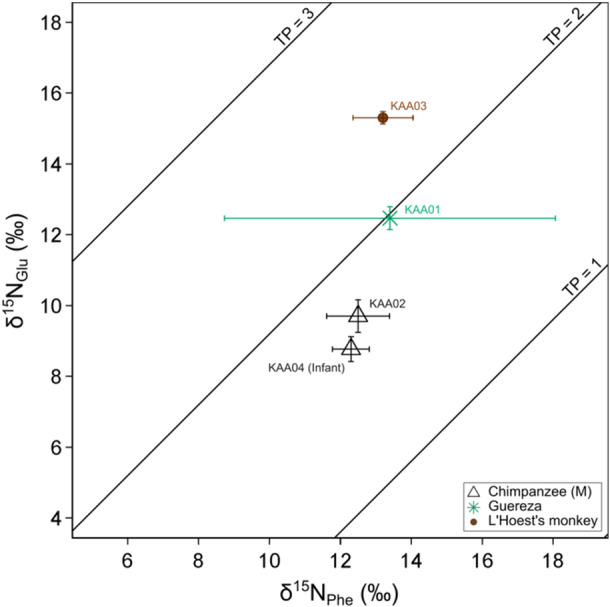 Figure 4
Figure 4| Consumer | Part | %C | %N | δ13C | δ15N |
| ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| Total | Fruits | 44.5 | 2.0 | 1.7 | 0.7 | −28.0 | 1.9 | 3.7 | 1.1 | 16 |
| Leaves | 44.1 | 3.2 | 3.3 | 1.2 | −31.8 | 2.6 | 3.3 | 1.2 | 12 | |
| Pith | 40.8 | 4.6 | 2.8 | 1.6 | −34.4 | 1.5 | 2.9 | 0.5 | 2 | |
| Other | 43.1 | 1.8 | 2.7 | 0.0 | −32.6 | 2.0 | 4.0 | 2.1 | 2 | |
|
|
|
|
|
|
|
|
|
|
| |
| Chimpanzee | Fruits | 44.6 | 2.1 | 1.6 | 0.8 | −27.5 | 1.5 | 3.6 | 1.0 | 14 |
| Leaves | 44.8 | 2.5 | 3.2 | 1.3 | −31.7 | 2.9 | 3.2 | 1.1 | 9 | |
| Pith | 44.0 | – | 1.7 | – | −33.4 | – | 3.3 | – | 1 | |
|
|
|
|
|
|
|
|
|
|
| |
| Blue monkey | Fruits | 45.0 | 1.8 | 1.7 | 0.6 | −27.7 | 1.4 | 3.7 | 1.3 | 9 |
| Leaves | 43.2 | 4.2 | 2.8 | 1.2 | −30.9 | 1.7 | 3.8 | 0.9 | 5 | |
|
|
|
|
|
|
|
|
|
|
| |
| Guereza | Fruits | 45.5 | 0.1 | 1.9 | 1.4 | −25.7 | 1.1 | 3.2 | 0.5 | 2 |
| Leaves | 44.4 | 2.4 | 3.5 | 1.7 | −31.7 | 3.9 | 3.0 | 1.1 | 5 | |
| Other | 41.8 | – | 2.7 | – | −31.2 | – | 2.5 | – | 1 | |
|
|
|
|
|
|
|
|
|
|
| |
| L'Hoest's monkey | Fruits | 45.1 | 1.9 | 1.8 | 0.6 | −28.2 | 2.1 | 3.8 | 1.2 | 9 |
| Leaves | 42.4 | 5.2 | 3.3 | 1.2 | −31.0 | 1.6 | 4.0 | 1.3 | 3 | |
| Pith | 40.8 | 4.6 | 2.8 | 1.6 | −34.4 | 1.5 | 2.9 | 0.5 | 2 | |
| Other | 43.1 | 1.8 | 2.7 | 0.0 | −32.6 | 2.0 | 4.0 | 2.1 | 2 | |
|
|
|
|
|
|
|
|
|
|
| |
| Red‐tailed monkey | Fruits | 45.0 | 1.8 | 1.7 | 0.6 | −27.7 | 1.4 | 3.7 | 1.3 | 9 |
| Leaves | 43.4 | 4.8 | 2.9 | 1.4 | −31.5 | 1.3 | 3.9 | 1.1 | 4 | |
|
|
|
|
|
|
|
|
|
|
|
| Response variables | Explanatory variables | Effect | SE |
|
| |
|---|---|---|---|---|---|---|
| δ13C | Fixed effect |
|
|
|
|
|
|
|
|
|
|
| ||
|
|
|
|
|
| ||
| Part: Pith | −5.50 | 3.19 | −1.73 | 0.097 | ||
| Stratif: Gap | 1.52 | 1.18 | 1.29 | 0.207 | ||
|
|
|
|
|
| ||
| Stratif: Understray | −1.17 | 2.90 | −0.40 | 0.690 | ||
| Season: Rainy | −0.94 | 0.59 | −1.58 | 0.119 | ||
| Random effect | SD of Species × Part | 3.40 | 1.84 | |||
| δ15N | Fixed effect |
|
|
|
|
|
| Part: Leaves | −0.68 | 0.46 | −1.45 | 0.157 | ||
|
|
|
|
|
| ||
|
|
|
|
|
| ||
|
|
|
|
|
| ||
| Stratif: Subcanopy | −0.20 | 0.51 | −0.39 | 0.703 | ||
|
|
|
|
|
| ||
| Season: Rainy | −0.25 | 0.42 | −0.60 | 0.554 | ||
| Random effect | SD of Species × Part | 1.80 | 1.34 | – | – | |
| Species | Category | %C | %N | δ13C | δ15N |
| ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| Blue monkey | Infant | 47.8 | 2.1 | 15.9 | 0.4 | −23.7 | 1.1 | 7.8 | 0.1 | 2 |
| Non‐infant | 45.5 | 3.5 | 15.8 | 1.7 | −24.1 | 0.7 | 7.2 | 0.6 | 3 | |
| Chimpanzee | Infant | 40.7 | — | 14.3 | — | −24.1 | — | 6.3 | — | 1 |
| Non‐infant F | 38.0 | 1.9 | 12.7 | 0.6 | −23.2 | 0.4 | 6.2 | 0.7 | 3 | |
| Non‐infant M | 42.7 | 2.5 | 14.1 | 1.0 | −24.1 | 0.6 | 6.4 | 0.5 | 5 | |
| Non‐infant total | 40.9 | 3.2 | 13.6 | 1.1 | −23.8 | 0.7 | 6.3 | 0.6 | 8 | |
| Guereza | Infant | 48.9 | — | 17.3 | — | −20.8 | — | 11.1 | — | 1 |
| Non‐infant | 41.0 | 2.5 | 14.0 | 0.9 | −21.8 | 0.8 | 8.6 | 0.8 | 6 | |
| L′Hoest's monkey | Non‐infant | 45.5 | — | 16.0 | — | −21.8 | — | 11.3 | — | 1 |
| Red‐tailed monkey | Non‐infant | 47.3 | 2.2 | 16.5 | 0.4 | −24.6 | 0.8 | 7.7 | 0.3 | 2 |
| Response variables | Explanatory variables | Effect | SE |
|
| |
|---|---|---|---|---|---|---|
| δ13C | Fixed effect |
|
|
|
|
|
| Species: Blue | −0.14 | 0.46 | −0.31 | 0.758 | ||
|
|
|
|
|
| ||
|
|
|
|
|
| ||
| Species: Red‐tailed | −0.63 | 0.62 | −1.03 | 0.318 | ||
| Age: Non‐infant | −0.37 | 0.46 | −0.81 | 0.430 | ||
| Random effect | SD of individual ID | 0.37 | 0.61 | — | — | |
| δ15N | Fixed effect |
|
|
|
|
|
| Species: Blue | 0.85 | 0.42 | 2.01 | 0.060 | ||
|
|
|
|
|
| ||
|
|
|
|
|
| ||
|
|
|
|
|
| ||
|
|
|
|
|
| ||
| Random effect | SD of individual ID | 0.12 | 0.35 | — | — |
| Species | ID | δ15NGlu | δ15NPhe | TP | |||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | Propagated error | ||
| Chimpanzee | KAA02 | 9.7 | 0.5 | 12.5 | 0.9 | 1.7 | 0.27 |
| KAA04 | 8.8 | 0.4 | 12.3 | 0.5 | 1.6 | 0.24 | |
| Guereza | KAA01 | 12.5 | 0.3 | 13.4 | 4.7 | 2.0 | 0.67 |
| L′Hoest's monkey | KAA03 | 15.3 | 0.2 | 13.2 | 0.9 | 2.4 | 0.33 |
- —Japan Society for the Promotion of Science10.13039/501100001691
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIsotope Analysis in Ecology · Primate Behavior and Ecology · Species Distribution and Climate Change
Introduction
1
Stable isotopes are used in primate ecological studies to investigate the diet and habitat use of various primate species (Crowley 2012; Sandberg et al. 2012). Among them, chimpanzees are probably the most well‐studied species, and various aspects of their behavior and ecology have been revealed mostly by carbon and nitrogen stable isotope analyses (Bădescu et al. 2017, 2022, 2025; Fahy et al. 2013; Loudon et al. 2016; Macho and Lee‐Thorp 2014; Oelze et al. 2020; Schoeninger et al. 1999, 2016; Sponheimer et al. 2006; Van Casteren et al. 2018). Because of the wealth of behavioral observational evidence, chimpanzees can serve as the model species to evaluate the precision of the stable isotopic methods by comparing the stable isotopic results with the known behavioral and environmental data.
Even in well‐studied chimpanzees, however, the scarcity of stable isotopic research on the two aspects of their diet hinders the accurate interpretations of their stable isotope ratios as a behavioral and ecological proxy. One aspect is the chimpanzee's dietary niche compared with other primate species in an ecosystem. Although their diet consists mainly of plants, the contribution of animal foods (faunivory), such as the consumption of invertebrate and vertebrate meat, has rarely been compared with other primate species in the same ecosystem (but see Cerling et al. 2004). While behavioral observation provides an estimate of the time spent feeding on certain food items, stable isotopes reveal the relative proportions of assimilated food sources to the tissue being analyzed, which is an ideal approach for quantitative inter‐species comparisons (Oelze et al. 2014; Schoeninger et al. 1998). The other understudied aspect is intra‐individual dietary variations (but see Fahy et al. 2014; Oelze et al. 2014, 2016). Although behavioral observations provide dietary data at a daily resolution, conventional stable isotope analysis only provides average data of months or longer periods, generating a gap in evidence between behaviors and stable isotopic data. By investigating how much variation in daily diet affects stable isotope ratios, it would be possible to expand the range of applications for stable isotope analysis in primate ecology.
In this study, profiling on carbon and nitrogen stable isotope ratios (δ^13^C and δ^15^N values, respectively) is performed on wild chimpanzees from Kalinzu Forest Reserve, Uganda, supplemented by that of food plants and other primate species. We hypothesized that the δ^13^C and δ^15^N values of the primate species in Kalinzu systematically differ with the diet and habitat of each species, and the inter‐species variations are greater than intra‐species variations. No previous study has reported stable isotopic data of chimpanzees in Kalinzu, and this study expands the understanding of the ecological determinants of stable isotope ratios of chimpanzees from various sites (Loudon et al. 2016; Schoeninger et al. 2016). The contribution of animal foods is compared between chimpanzees and other sympatric primate species, and compound‐specific nitrogen stable isotope analysis was preliminarily applied to further investigate faunivory. Because the trophic enrichment of ^15^N is more pronounced in non‐essential amino acids, the δ^15^N values of individual amino acids can be used to obtain more accurate estimates of faunivory (Chikaraishi et al. 2007, 2011; Ohkouchi et al. 2017). Daily to weekly dietary change is also investigated by applying ultra‐fine time‐series analysis of a chimpanzee's hair, using an ultra‐sensitive analyzer (Ogawa et al. 2010). This profiling deepens understanding of the stable isotopic ecology of wild chimpanzees and facilitates the connection between behavioral and stable isotopic data.
Materials and Methods
2
Study Site
2.1
Primate hair and food samples were collected from July 2013 to May 2017 in a moist, medium‐altitude evergreen forest in the Kalinzu Forest Reserve, western Uganda, covering an area of 137 km^2^ (30°07′ E, 0°17′ S; altitude 1000–1500 m above sea level) (Hashimoto et al. 1999). The annual rainfall from June 1997 to May 1998 and in 2015 was 1584 mm (Hashimoto et al. 1999) and 1370 mm (Matsuda et al. 2020), respectively. The mean minimum and maximum daily temperatures in each month from November 2013 to April 2016 were 14.0 ± 1.9°C and 27.2 ± 2.1°C, respectively (Matsuda et al. 2020). Two mild rainy seasons (late March–May and late September–December) and two mild dry seasons (January–early March and June–early September) exist. The four major types of vegetation characterize Kalinzu: mixed mature forest, Parinari‐dominated mature forest, Parinari‐dominated secondary forest, and Musanga‐dominated secondary forest (Hashimoto et al. 1999). Fruits of Ficus are abundant throughout the year, providing abundant food sources to primate species (Furuichi et al. 2001; Hashimoto et al. 2001).
The Kalinzu forest harbors six species of diurnal primates: eastern chimpanzee (P. t. schweinfurthii), blue monkey (Cercopithecus mitis), red‐tailed monkey (Cercopithecus ascanius), L′Hoest's monkey (Allochrocebus lhoesti), black and white colobus or guereza (Colobus guereza), and olive baboon (Papio anubis). Primates in Kalinzu have been studied since 1992 (Hashimoto 1993), and numerous behavioral and ecological studies have been conducted on primate feeding ecology (e.g., Aruga et al. 2015; Furuichi 2006; Hashimoto et al. 2001; Ihobe 2001; Matsuda et al. 2020; Tashiro 2006). However, stable isotopic studies on the diet of primate species in Kalinzu have never been reported.
Sample Collection
2.2
Hairs of chimpanzees, blue monkeys, red‐tailed monkeys, L′Hoest's monkeys, and guerezas were collected in Kalinzu opportunistically and non‐invasively during the study period (Supporting Table S1). Hairs of chimpanzees were mostly collected from the ground nest and the ground site where grooming was conducted. Hairs of other primate species were mostly collected from dead bodies found inside the forest. The cause of their death seemed to be physical trauma (e.g., falling from a tree or infanticide) in most cases, based on the behavioral observations or the appearance of the bodies and the surrounding situations. Chimpanzee hair used for ultra‐fine time‐series analysis was plucked and collected from the fresh dead body of an adult male (Kobo) on July 30, 2013, who was probably killed by lethal aggression. This is to exclude the time gap existing in the naturally shed hairs in the telogen phase. Each hair enters the telogen phase, where hair incremental growth stops for more than a few months, before shedding (Harkey 1993; Saitoh et al. 1970). Thus, the use of naturally shed hairs complicates the represented time window of the subject hair segments and the correspondence between behavioral and stable isotopic data of the host individual. After the collection, hair was stored in a dry condition at ambient temperatures or frozen condition at −20°C.
Food plant samples were collected during July and August of 2013 (dry season), February and March of 2015 (dry season), and November and December of 2015 (rainy season) (Supporting Table S2). Plant samples were cut into small pieces of less than 1 cm^3^, air‐dried overnight, and dried with silica gel. Vertical stratifications that potentially affect plant stable isotope ratios (Lowry et al. 2021; van der Merwe and Medina 1991) are defined as follows: “crown,” canopy crown structure of the forest; “understory,” the most basal flora below < 1.5 m from the ground where direct solar radiation does not reach; “subcanopy,” intermediate space between crown and understory where direct solar radiation does not reach but locates above 1.5 m from the ground; and “gap,” flora with direct solar radiation because of the clearance of canopy crown. Investigation into the baseline plant stable isotope ratios is important to accurately interpret the stable isotope ratios of primates (Wessling et al. 2019) and also benefits DNA‐based studies of diet.
The hair and plant samples were exported and analyzed under the material transfer agreement with the Uganda Wildlife Authority and the National Forest Authority. The primate hair samples were exported with the approval of the Uganda National Council for Science and Technology (NS 184) and the Ministry of Economy, Trade and Industry, Japan (17JP000003/TI), under the Convention on International Trade in Endangered Species (CITES). All research procedures followed the Code of Best Practices for Field Primatology (Riley et al. 2014) and were approved by the Ethics Committee for Animal Research of the Graduate University for Advanced Studies (SKD2025AR003).
Sample Processing
2.3
Hair samples were washed with Milli‐Q water under sonication and dried. Then, lipid‐extraction was performed by soaking the hairs in a hexane:dichloromethane (3:2 in volume) solution over 2 h. After the removal of organic solvent with washing by Milli‐Q water, the hair was air‐dried and used for stable isotope analyses.
Plant samples were freeze‐dried overnight and crushed into a fine powder with a stainless motor and pestle, if the matrix heterogeneity was high. The homogenized samples were used for stable isotope analyses.
Ultra‐Fine Sectioning
2.4
A single hair strand in the growing anagen phase of an adult male chimpanzee (Kobo) was cut into 1 mm sections and used for the timeseries analysis. The hair was wrapped in aluminum foil, wetted with ethanol to prevent electrostatics, and manually dissected in a 1 mm unit with a pair of scissors. To avoid the accumulation of small errors, the hair was first cut into long 5 mm segments, and each segment was cut into five 1 mm pieces. When the 1 mm segment was lost, data were not obtained in the corresponding period. Based on the previous data in humans (Harkey 1993; Saitoh et al. 1970), the hair growth rate was assumed to be 1 mm in 3 days (i.e., 1 cm in 1 month). Therefore, for example, the first, second, and third 1 mm segments from the hair root represent the 3‐day periods of the last 1–3 days (median 1.5 days), 4–6 days (median 4.5 days), and 7–9 days (median 7.5 days) from the estimated date of the death (July 30, 2013).
The microscopic observations on the hair follicle were performed on the hair strands to assess the hair growth cycle of the typical hairs obtained from Kobo. Keyence VHX‐2000 was used for the microscopic observation. The observed follicles are transparent white in color with an elongated, straight form, which are the typical features of hairs in the anagen phase (Supporting Figure S1). This means that typical hairs of Kobo are not hairs that have ceased growing and retain a record of past dietary intake until they eventually fall out. Rather, the typical hairs of Kobo continuously recorded dietary changes up until the time of his death. Therefore, there was no gap between the assigned time window to the segments and the actual timing of hair growth that reflects dietary stable isotopic signals at the time.
Bulk Stable Isotope Analysis of Plants
2.5
Bulk carbon and nitrogen stable isotope ratios of plant samples were measured using an elemental analyzer–isotope ratio mass spectrometry (Thermo Flash 2000 elemental analyzer, Finnigan ConFlo III interface, and Thermo Delta V mass spectrometer) at the University Museum, University of Tokyo, Japan. Carbon and nitrogen stable isotope ratios were expressed in δ notation relative to standards, δ^13^C (‰) and δ^15^N (‰), with Vienna Pee Dee Belemnite (VPDB) and atmospheric nitrogen (AIR) as the standard for carbon and nitrogen, respectively. The δ^13^C and δ^15^N values were calibrated against the laboratory working standard (l‐alanine: δ^13^C = −19.6 ± 0.2‰; δ^15^N = 8.7 ± 0.2‰) provided by SI Science (Saitama, Japan), whose values were determined by the NBS 19 and the International Atomic Energy Agency (IAEA) Sucrose ANU (calibrated against Pee Dee Belemnite) and IAEA N1 and IAEA N2 (calibrated against AIR) international standards, respectively. Based on repeated measurements of the calibration standards, analytical errors were determined to be less than ±0.1‰ for both δ^13^C and δ^15^N. Based on the carbon and nitrogen atomic contents obtained during the measurements, the carbon and nitrogen weight proportions of the samples (%C and %N) were calculated, from which the atomic carbon to nitrogen (C/N) ratio was further determined.
Conventional and Ultra‐Sensitive Bulk Stable Isotope Analyses of Hairs
2.6
Bulk carbon and nitrogen stable isotope ratios (δ^13^C_bulk_ and δ^15^N_bulk_) of hair samples were measured using a modified elemental analyzer/isotope mass spectrometer at the Japan Agency for Marine‐Earth Science and Technology (JAMSTEC). Instrumentation consisted of an elemental analyzer (FlashEA 1112, Thermo Finnigan) coupled to an isotope ratio mass spectrometer (Delta plus XP, Thermo Finnigan) through a continuous flow interface (ConFloIII, Thermo Finnigan) with modification specifically made to improve the sensitivity of the analysis (Ogawa et al. 2010). This system achieved increased sensitivity via optimized chromatography, reduced blank concentrations, and increased sample amount introduced to the ion source for mass spectrometry (Ogawa et al. 2010). Stable isotope ratios were calibrated using three laboratory standards: l‐tyrosine (BG‐T: δ^13^C = −20.83 ± 0.10‰, δ^15^N = 8.74 ± 0.04‰), l‐proline (BG‐P: δ^13^C = −10.27 ± 0.04‰, δ^15^N = 13.51 ± 0.02‰), and dl‐alanine (CERKU‐01: δ^13^C = −25.36 ± 0.08‰, δ^15^N = −2.89 ± 0.04‰). These stable isotopic compositions were calibrated with authentic standards (Tayasu et al. 2011). The analytical errors determined by the repeated measurements of l‐tyrosine (BG‐T) at the same timing of sample analysis were± 0.2–0.7‰ and ± 0.2–0.6‰ for δ^13^C and δ^15^N, respectively.
Compound‐Specific Stable Isotope Analysis of Amino Acids
2.7
Compound‐specific stable isotope analysis (CSIA) of amino acid δ^15^N (δ^15^N_AA_) was performed for selected hair samples. Samples for CSIA were prepared based on the amino acid derivatization procedures described in Chikaraishi et al. (2015). Briefly, samples (~1 mg) were hydrolyzed with 12 M HCl at 110°C overnight, and then washed with n‐hexane:dichloromethane (3:2 in volume) to remove the remaining hydrophobic compounds. After the hydrolysis, samples were derivatized using thionyl chloride:2‐propanol (1:4 in volume) at 110°C for 2 h and then pivaloyl chloride:dichloromethane (1:4 in volume) at 110°C for 2 h. Then, the amino acid derivatives were extracted with n‐hexane:dichloromethane (3:2 in volume). The δ^15^N values of each amino acid were determined using a gas chromatograph (6890N, Agilent Technologies) coupled to an isotope ratio mass spectrometer (Delta plus XP, Thermo Finnigan) via a GC‐combustion III interface (Thermo Finnigan) at JAMSTEC (Ishikawa 2018; Ishikawa et al. 2022). Reference mixtures of seven amino acids (alanine, valine, leucine, norleucine, aspartic acid, glutamic acid, and phenylalanine, provided by Indiana University and Shoko Science Co. Ltd.) with known δ^15^N values (ranging from +1.7‰ to +45.7‰) were analyzed every six sample runs. Duplicate or triplicate measurements were performed for each amino acid. The analytical error of the standards was typically smaller than 1.0‰.
Behavioral Observation
2.8
Behavioral observations of chimpanzees were performed daily by four local research assistants. The scan sampling of behaviors of chimpanzees in the observer's sight was obtained every 10 min from 8:00 to 17:00 from Monday to Saturday. The species and parts of food items consumed were also recorded during the scans. The presence of Kobo, the chimpanzee individual whose hair was used for ultra‐fine time‐series analysis, and his feeding items were evaluated from these group‐scan‐based behavioral observation records.
Based on the feeding behavioral data, each combination of plant species and part was scored on whether each primate species consumes it. Since no published food list is available for species other than the guereza (Matsuda et al. 2020), scoring was conducted based on the personal observations of researchers who have been studying the relevant species for several years in Kalinzu. The scoring results were reviewed by local research assistants and other researchers working on these primate species in Kalinzu. Results reported by Matsuda et al. (2020) were used to identify food items in guereza.
Data Analysis
2.9
Statistical analyses were performed in R software environment, version 4.2.3 (R Core Team 2023). Linear mixed models (LMMs) were applied using lme4, version 1.1–31 (Bates et al. 2015) and lmerTest, version 3.1–3 (Kuznetsova et al. 2017) packages. The statistical significance level was set as α < 0.05. Linear mixed models were applied with the following conditions:
- To reveal the drivers of stable isotopic variations in plants, plant δ^13^C_bulk_ or δ^15^N_bulk_ values were set as the response variable, and part (fruits, leaves, pith, or others), stratification (crown, subcanopy, understory, or gap), and season (dry or rainy) were set as the explanatory variables. The combination of species and part was set as a random effect.
- To reveal the stable isotopic difference of the primate species, hair δ^13^C_bulk_ or δ^15^N_bulk_ values were set as the response variable, and primate species and age class (infant or non‐infant) were set as the explanatory variables. The individual ID of primates was set as a random effect.
To reduce the sampling bias, mean values of samples in the same category (i.e., the combination of species and part in plants and individuals in hairs) were used to calculate summary statistics, such as means and standard deviations, when applying statistical tests. On the other hand, bias in the number of samples was considered in LMMs by incorporating them into random effects. Also, to control the effect of breast milk consumption on the stable isotope ratios (Tsutaya and Yoneda 2015), only non‐infant data were used for comparison.
The trophic position (TP) was calculated based on the stable nitrogen stable isotope ratios of glutamic acid and phenylalanine with the following equation proposed in Chikaraishi et al. (2011, 2014):
Uncertainty in the TP was calculated by propagation of errors by combining the uncertainties in the stable isotopic difference between glutamic acid and phenylalanine in reference mixtures of amino acids (σ = 1.5‰), the trophic discrimination factors of glutamic acid and phenylalanine during each trophic transfer (σ = 1.2‰ and 0.5‰, respectively: Chikaraishi et al. 2009), and the measured analytical reproducibility of δ^15^N_Glu_ and δ^15^N_Phe_ values in each sample, by using the equation shown in Ishikawa et al. (2022). It should be noted that the propagated errors might overestimate the TP estimate errors.
Results
3
Bulk Stable Isotope Ratios of Plants
3.1
The total averages of δ^13^C_bulk_ and δ^15^N_bulk_ values of each combination of plant species and part that were consumed by any of the primate species were −30.1 ± 3.1‰ and 3.5 ± 1.1‰, respectively (Table 1; Figure 1a). The average δ^13^C_bulk_ (from −30.2‰ to −28.8‰) or δ^15^N_bulk_ (from 3.0‰ to 3.8‰) values of consumed food items were similar among each primate species (Table 1).
Distributions of carbon and nitrogen stable isotope ratios of (a) food plants and (b) primate hairs from Kalinzu. Plant species and part eaten by chimpanzees are shown in solid points (a). Samples used for CSIA are shown with its ID (KAA) and those from infants are also shown (b).
The results of LMMs showed significantly lower δ^13^C_bulk_ values of leaves compared with fruits, significantly lower δ^13^C_bulk_ values in samples obtained from the subcanopy compared with those from the crown, significantly lower δ^15^N_bulk_ values in piths and other parts compared with fruits, and significantly higher δ^15^N_bulk_ values of samples from the understory compared with those from the forest crown (Table 2). The average δ^13^C_bulk_ value of leaves was 3.8‰ lower than that of fruits (Table 1). Compared with plant samples obtained from the crown, the average δ^13^C_bulk_ value of plant samples obtained from the subcanopy was 2.5‰ lower, and the average δ^15^N_bulk_ value of plant samples obtained from the understory was 0.3‰ higher (Supporting Table S3).
Bulk Stable Isotope Ratios of Primate Hairs
3.2
The average bulk stable isotope ratios of five primate species ranged from −24.6‰ to −21.8‰ in carbon and from 6.2‰ to 11.3‰ in nitrogen in adults (Table 3; Figure 1b). The results of LMMs showed significantly higher δ^13^C_bulk_ values of guereza and L′Hoest's monkey compared with chimpanzees, significantly higher δ^15^N_bulk_ values of guereza, L′Hoest's monkey, and red‐tailed monkey compared with chimpanzees, and significantly higher δ^15^N_bulk_ values of infants (Table 4).
Mann–Whitney U‐tests showed no significant sex difference in δ^13^C_bulk_ (U = 13, p = 0.134) and δ^15^N_bulk_ (U = 6, p = 0.786) values of chimpanzees.
The average δ^13^C_bulk_ or δ^15^N_bulk_ values of the non‐infant Kalinzu chimpanzees were −23.8 ± 0.7‰ and 6.3 ± 0.6‰, respectively (Table 3). The stable isotopic differences between diet and hair were 5.5‰ and 2.9‰ for carbon and nitrogen, comparable to those estimated in captive chimpanzees in an experimental setting (Tsutaya et al. 2017). Comparison of these values with the stable isotope ratios of chimpanzees and their food plants from other study sites across Africa indicated lower δ^13^C_bulk_ values for both hair and food plants but similar δ^15^N_bulk_ values compared with those predicted by the relationships between annual rainfall and stable isotope ratios (Figure 2; Supporting Table S4). Although the δ^13^C_bulk_ values of hair and plants were 1.7‰ and 1.3‰ lower than the predicted values, respectively, the δ^15^N_bulk_ values were both only 0.1‰ higher than the predicted values (Figure 2).
The relationship between mean annual rainfall and (a) carbon or (b) nitrogen stable isotope ratios of food plants and hairs of chimpanzees across different sites. The 1 SD ranges of each data point are shown for stable isotope ratios. The diameters of plots for hairs correspond to the number of analyzed samples in each study. The predicted relationship and its error ranges are shown with solid and dotted lines, respectively. Please see Supporting Table S4 for the information on each site and Supporting Text S1 for the details of the linear regression.
Time‐Series Analysis of a Chimpanzee Hair
3.3
Ultra‐fine time‐series analysis of Kobo's hair revealed rapid fluctuations of δ^13^C_bulk_ or δ^15^N_bulk_ values in daily to weekly scales (Figure 3; Supporting Table S5). The δ^13^C_bulk_ or δ^15^N_bulk_ values of 1 mm segments corresponding approximately 3 days during 84 days before the death ranged from −24.7‰ to −23.7‰ and from 6.5‰ to 8.0‰ (Supporting Table S5). Although most parts of the hair segments have a relatively constant stable isotope ratio, a 1.0‰ increase occurred during 9 days just before the death in δ^13^C_bulk_ values, and a 1.4‰ increase and subsequent 1.0‰ decrease occurred during 12 days in δ^15^N_bulk_ values (Figure 3). If the stable isotope ratios of 1 mm segments (~3 days) were averaged, based on the length, into hypothesized 5 mm segments (half a month), such rapid fluctuations were not seen (Figure 3), and the difference between the minimum and maximum values decreased to 0.4‰ and 0.6‰ for δ^13^C_bulk_ or δ^15^N_bulk_ values, respectively (Supporting Table S6).
Carbon and nitrogen stable isotope ratios of the ultra‐fine sections of the hair of a male chimpanzee (Kobo), plotted against the days before his death. The average values of 5 mm segments are calculated and shown in gray horizontal bars with the height of typical analytical uncertainty of ±0.1‰. Days with behavioral observation records are shown in rectangular blocks above the stable isotopic trajectories, and days with the observation of faunivory are shown in red.
Among the 29 days in which Kobo was observed during the 87‐day period before his death, animal food consumption (i.e., ant‐eating) was only seen once at 49 days before his death, and no rapid δ^13^C_bulk_ or δ^15^N_bulk_ fluctuation can be seen after this ant‐eating event. Kobo was relatively well observed during the 19‐day period before his death, in which the rapid increase in hair δ^13^C_bulk_ values occurred, but no animal food consumption was recorded.
Compound‐Specific Stable Isotope Analysis of Primate Hairs
3.4
The measured δ^15^N_Phe_ values of chimpanzees, guereza, and L′Hoest′s monkey were similar (from 12.3‰ to 13.4‰), indicating that they share the same nitrogen source (Figure 4; Table 5; Supporting Table S7). It should be noted that phenylalanine for guereza and L′Hoest′s monkey coeluted with unknown impurities in one of their three chromatograms. Consequently, their δ^15^N_Phe_ values were determined in duplicate, and guereza δ^15^N_Phe_ particularly showed larger uncertainty than others (Figure 4). The δ^15^N_Glu_ values, which typically increase with the increase in TP, were the lowest in chimpanzees (8.8‰ and 9.7‰) and larger in guereza (12.5‰) and in L′Hoest′s monkey (15.3‰) (Table 5).
Nitrogen stable isotope ratios of Glutamic acid and Phenylalanine of primate hairs from Kalinzu. Sample IDs and estimated TPs calculated based on the equation mentioned in the main text are also shown.
The TPs calculated from the results of CSIA were 1.6–1.7, 2.0, and 2.4 for chimpanzees, guereza, and L′Hoest's monkeys, respectively (Table 5; Supporting Figure S2). The TPs are expected to be 2.0 for obligate plant consumers and 3.0 for faunivorous animals that exclusively consume plant‐eating animals. This preliminary result indicates that the major dietary nitrogen sources were plants in chimpanzees and guereza, but certain amounts of foods in higher TP, such as insects, were incorporated into the diet of L′Hoest's monkey, although more measurement data are needed to reach a robust conclusion.
Discussion
4
Stable Isotopic Variation Among Ecosystems
4.1
The hair and plant food stable isotope ratios of Kalinzu chimpanzees were generally consistent with those predicted based on the relationship between rainfall and stable isotope ratios, although the hair and plant δ^13^C_bulk_ values were lower than those predicted from the general trend across various chimpanzee study sites (Figure 2; Supporting Table S4). The δ^13^C_bulk_ values of terrestrial C3 plants show a negative correlation with the amount of rainfall (Kohn 2010), and chimpanzee hairs also reflect this trend (Loudon et al. 2016; Schoeninger et al. 2016). Because the chimpanzee hair and the plants the chimpanzees consumed in Kalinzu show lower δ^13^C_bulk_ values, the deviation from the general trend was not caused by the distinct dietary habits of Kalinzu chimpanzees but by environmental factors affecting the baseline plant stable isotope ratios. Although the cause of this baseline change is unknown, this result emphasizes the importance of analyzing the stable isotope ratios of food plants to compare those of primate tissues (Wessling et al. 2019).
Multiple factors affect plant δ^15^N_bulk_ values, such as climate, type of mycorrhizal association, and nitrogen sources (Amundson et al. 2003; Craine et al. 2009, 2015; Szpak 2014), and the relationship between plant δ^15^N_bulk_ values and environmental factors is complex. Reflecting such complexity, chimpanzee δ^15^N_bulk_ values vary by baseline change and dietary content, such as meat (Loudon et al. 2016; Schoeninger et al. 2016). Despite this variation, plant and chimpanzee δ^15^N_bulk_ values did not deviate from typical values previously reported, suggesting that animal meat, typically showing higher δ^15^N values than plants, was not the major dietary component of Kalinzu chimpanzees.
Stable Isotopic Variations Among Primate Species
4.2
The comparison of hair stable isotope ratios of five primate species in Kalinzu indicated the lower δ^13^C_bulk_ and the lowest δ^15^N_bulk_ values in chimpanzees (Figure 1b, Tables 3 and 4). This result suggests little dietary contribution of animal protein, which typically shows higher δ^13^C_bulk_ and δ^15^N_bulk_ values because of higher TPs, in chimpanzees' diet. The CSIA results also support the relatively low contribution of animal protein in Kalinzu chimpanzees (Figure 4; Table 5). Previous studies have reported the successful reconstruction of meat consumption using bulk stable isotope analysis of bone collagen in Taï chimpanzees (Fahy et al. 2013), although not from the bulk hairs (Oelze et al. 2020). Our results, on the contrary, demonstrated that in Kalinzu chimpanzees, both bulk and CSIA analyses of hair were effectively employed to assess the relatively lower dietary contribution of animal proteins. Nevertheless, it should be acknowledged that research on meat‐eating behaviors has been scarce, and previous systematic studies have focused on the extent of ant‐eating behaviors in Kalinzu (Hashimoto et al. 2000, 2015; Koops et al. 2015; Tashiro et al. 1999; Hashimoto et al. 2001; Ihobe 2001; Shirasawa et al. 2024).
A L′Hoest′s monkey showed the highest δ^13^C_bulk_ and δ^15^N_bulk_ values among the primate species in Kalinzu (Figure 1b; Table 3). The highest δ^13^C_bulk_ value is partly inconsistent with the visually observed feeding behaviors of L′Hoest′s monkeys, whose daily activities are mostly spent in the lower strata of the forest (Tashiro 2006). Although the trend was not statistically significant in this study (Table 2), the typical δ^13^C values of plants from the understory are lower than those from the forest canopy (Lowry et al. 2021; van der Merwe and Medina 1991), and such lower δ^13^C values are reflected in consumers. On the other hand, the highest δ^15^N_bulk_ value is consistent with the fact that a large proportion of L′Hoest′s monkeys' diet consists of terrestrial invertebrates, such as insects (Tashiro 2006). Considering that the δ^15^N_bulk_ values of plants from undertray are significantly high in this site (Table 2) and the fact that insects generally show higher δ^15^N values than plants (e.g., Hyodo et al. 2010), the δ^15^N values in L′Hoest′s monkey were consistent with their behavioral traits. The TP of 2.4, a slight faunivory, calculated in L′Hoest′s monkey in Kalinzu (Table 5) also suggests their consumption of terrestrial invertebrates. Relatively higher δ^15^N_bulk_ values in blue monkeys and red‐tailed monkeys compared with chimpanzees (Figure 1b; Table 3) agree with the higher contributions of terrestrial invertebrates in their diet (Struhsaker 2017; Tashiro 2006). Nevertheless, given that only one data point is available for L′Hoest′s monkeys, it is necessary to increase the sample size for a more robust discussion.
The TP 2.0 measured by CSIA accurately reflects that guerezas are obligate leaf‐eaters. On the other hand, guerezas' δ^13^C_bulk_ and δ^15^N_bulk_ values are significantly higher than those of chimpanzees (Table 4) and are the second highest after L′Hoest′s monkeys (Figure 1). Considering their heavy reliance on a limited variety of plant leaves (Matsuda et al. 2020), unknown physiological mechanisms might affect guerezas' relatively higher bulk stable isotope ratios. The investigation into the dietary, ecological, and physiological determinants of stable isotope ratios in guerezas is an interesting theme for future stable isotopic studies.
Across the primate species in Kalinzu, hairs from infants showed significantly higher δ^15^N_bulk_ values (Table 4). Infants show higher δ^15^N_bulk_ values (2–3‰) during the breastfeeding period because of breast milk consumption (Fahy et al. 2014; Tsutaya and Yoneda 2015), which caused the significantly higher δ^15^N_bulk_ values in infant primate individuals in Kalinzu.
Intra‐Individual Variations
4.3
Rapid fluctuations of stable isotope ratios (> 1‰) within several days were observed in the ultra‐fine time‐series analysis of a single hair employed in this study, which was not previously observed in the sequential analysis of non‐human primate hairs (e.g., Oelze et al. 2014, 2016). In particular, δ^15^N_bulk_ values showed a fluctuation (1.4‰) over a period of only 6 days (Figure 4; Supporting Table S5) that was comparable to the largest difference (1.5‰) between chimpanzee individuals analyzed in Kalinzu (Table 3). If the fine‐scale values over a period of several weeks are clumped, the variation in stable isotope ratios from a single individual becomes smaller (Figure 3; Supporting Table S6). Therefore, it should be noted that such average results can mask rapid potential fluctuations in stable isotope ratios, as shown in this study. Similar issues have been reported in other tissues with incremental growth, such as shells (Burchell et al. 2013) and teeth (Tsutaya 2020). As recent analyses with higher temporal resolution are increasingly applied, the temporal resolution of stable isotope analysis may come closer to the daily resolution of behavioral observations, revealing previously unanticipated intra‐individual variations in stable isotope ratios and nutrient intake.
When using behavioral observation, collecting dietary data continuously over several months for a single individual can be difficult, and ultra‐fine time‐series stable isotope analysis of hairs offers a new solution. Continuous analysis of tooth dentine offers a sequential record, but the temporal resolution of each increment is relatively coarse, on the order of several months (Fahy et al. 2014). Fecal samples reflect short‐term dietary variation over a period of a few days (Bădescu et al. 2017), but it is not always practical to collect them continuously for several months. Our analysis revealed a stable isotopically fluctuating diet in Kobo, on the order of a few days, even for the periods without behavioral observational data. In our dataset, 64 days (73.6%) of the 87‐day period corresponding to the hairs analyzed were not accompanied by an observational record of Kobo (Supporting Table S8), and the diet during this period was not observed. Further investigations into the factors affecting intra‐individual stable isotopic variation, such as the rate and extent to which body nitrogen pools reflect inputs, physiological factors that can deviate hair growth, and fine‐scale time‐series changes in baseline stable isotope ratios, enable more nuanced interpretations of the ultra‐fine time‐series data. Such detailed knowledge of primate diet also contributes to a better interpretation of results obtained from DNA‐based studies, such as fecal metagenomics (Mallott et al. 2017) and chemosensory gene evolution (Hayakawa et al. 2012; Itoigawa et al. 2021), by providing quantitative baseline data of their feeding ecology.
Stable isotope analysis of primate hair has traditionally focused on dietary differences between individuals, but by applying ultra‐fine time‐series analysis, we can add a detailed temporal dimension to this analysis. This enables the investigation of interspecific and intraspecific dietary niche competition and differentiation in relation to the seasonality of food resources and social interactions among individuals. Such investigations are applicable to subjects that are well‐suited for stable isotope analysis, such as unhabituated populations and museum specimens, where direct behavioral observation is difficult. Improving the precision and the throughput of sectioning will be important in the future to implement these applications more effectively. Because the length of the hair segments to be sampled in ultra‐fine sectioning is less than one‐fifth to one‐tenth the length compared with the conventional analysis, even the same degree of uncertainty in the length of the hair segments results in a relatively larger difference in the assigned time window. Because of the limited precision of manually cutting hairs into segments, it is necessary to use a microtome or other precise machine to section the hairs to finely control the time window represented by the hair segment (Matsubayashi et al. 2017). Such a partial automation of sectioning would also improve the throughput and enable a large‐scale analysis.
Conclusions
5
Carbon and nitrogen stable isotope analyses were performed on the hair and food plants of primates in Kalinzu to profile the stable isotopic variations. The baseline δ^13^C values in Kalinzu were > 1‰ lower than those predicted from the rainfall. The δ^13^C and δ^15^N values of chimpanzees were generally lower than those of other primate species, and CSIA showed their plant‐based diet, indicating the relatively small contribution of animal foods in the protein intake in Kalinzu chimpanzees. Ultra‐fine time‐series analysis of an adult male chimpanzee's hair revealed rapid intra‐individual variations. This study showed that by improving and carefully implementing stable isotopic methods, detailed data comparable to behavioral observations can be obtained in primate ecology.
Author Contributions
Conceptualization: Takumi Tsutaya, Natsumi Aruga, Chie Hashimoto. Methodology: Takumi Tsutaya. Software: Takumi Tsutaya. Investigation: Takumi Tsutaya, Naoto F. Ishikawa, Yoko Sasaki, Minoru Yoneda, Nanako O. Ogawa, Naohiko Ohkouchi, Chie Hashimoto. Resources: Natsumi Aruga, Haruka Kitayama, Chie Hashimoto. Writing – Original Draft: Takumi Tsutaya. Writing – Review and Editing: Takumi Tsutaya, Naoto F. Ishikawa, Haruka Kitayama, Nanako O. Ogawa, Naohiko Ohkouchi, Chie Hashimoto. Visualization: Takumi Tsutaya. Funding acquisition: Takumi Tsutaya, Chie Hashimoto.
Supporting information
kalinzuhair2_si.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Amundson, R. , A. T. Austin , and E. A. G. Schuur , et al. 2003. “Global Patterns of the Isotopic Composition of Soil and Plant Nitrogen.” Global Biogeochemical Cycles 17: 31. 10.1029/2002 GB 001903. · doi ↗
- 2Aruga, N. , H. Matsuo , T. Furuichi , and C. Hashimoto . 2015. “Root Eating by Wild Chimpanzees (Pan troglodytes) in the Kalinzu Forest, Uganda: Possible Medicinal Plant Use.” Pan Africa News 22: 7–10. 10.5134/198895. · doi ↗
- 3Bădescu, I. , C. Curteanu , D. W. Sellen , D. P. Watts , and M. A. Katzenberg . 2025. “Investigating Infant Feeding Development in Wild Chimpanzees Using Stable Isotopes of Naturally Shed Hair.” American Journal of Primatology 87: e 23552. 10.1002/ajp.23552.37779353 PMC 11650929 · doi ↗ · pubmed ↗
- 4Bădescu, I. , M. A. Katzenberg , D. P. Watts , and D. W. Sellen . 2017. “A Novel Fecal Stable Isotope Approach to Determine The Timing of Age‐related Feeding Transitions in Wild Infant Chimpanzees.” American Journal of Physical Anthropology 162: 285–299. 10.1002/ajpa.23116.27768227 · doi ↗ · pubmed ↗
- 5Bădescu, I. , D. P. Watts , M. A. Katzenberg , and D. W. Sellen . 2022. “Maternal Lactational Investment is Higher For Sons in Chimpanzees.” Behavioral Ecology and Sociobiology 76: 44. 10.1007/s 00265-022-03153-1. · doi ↗
- 6Bates, D. , M. Mächler , B. Bolker , and S. Walker . 2015. “Fitting Linear Mixed‐Effects Models Using lme 4.” Journal of Statistical Software 67: 1–48. 10.18637/jss.v 067.i 01. · doi ↗
- 7Burchell, M. , A. Cannon , N. Hallmann , H. P. Schwarcz , and B. R. Schöne . 2013. “Refining Estimates for the Season of Shellfish Collection on the Pacific Northwest Coast: Applying High‐Resolution Stable Oxygen Isotope Analysis and Sclerochronology.” Archaeometry 55: 258–276. 10.1111/j.1475-4754.2012.00684.x. · doi ↗
- 8Cerling, T. E. , J. A. Hart , and T. B. Hart . 2004. “Stable Isotope Ecology in the Ituri Forest.” Oecologia 138: 5–12. 10.1007/s 00442-003-1375-4.14530961 · doi ↗ · pubmed ↗