Social evaluation of skilfulness in Tonkean macaques (Macaca tonkeana) and brown capuchins (Sapajus apella)
Marie Hirel, Michele Marziliano, Hélène Meunier, Hannes Rakoczy, Julia Fischer, Stefanie Keupp

TL;DR
This study explores whether Tonkean macaques and brown capuchins can evaluate the skilfulness of others based on their actions.
Contribution
The study investigates how nonhuman primates form social evaluations about others' competence.
Findings
Subjects did not consistently choose the skilful actor over the unskilled one.
Initial preferences for the skilful actor showed a significant shift.
Subjects looked preferentially at the skilful actor during simultaneous container manipulation.
Abstract
For optimal decision-making, social animals can benefit from evaluating others’ behaviours. Some species seemingly consider the skills of others when deciding who to interact with in different contexts. Yet, whether and how nonhuman animals form impressions about others’ competence is still unclear. In this study, we investigated whether Tonkean macaques (Macaca tonkeana) and brown capuchins (Sapajus apella) can evaluate the skilfulness of others. Subjects observed two human actors (one skilful, one unskilled) trying to open several food containers. Only the skilful actor successfully opened the containers and released food so the experimenter could give it to the subjects. Our results revealed that subjects did not choose the skilful actor significantly more frequently than the unskilled one. Their choices for the skilful actor did not increase through trials nor were they based on the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
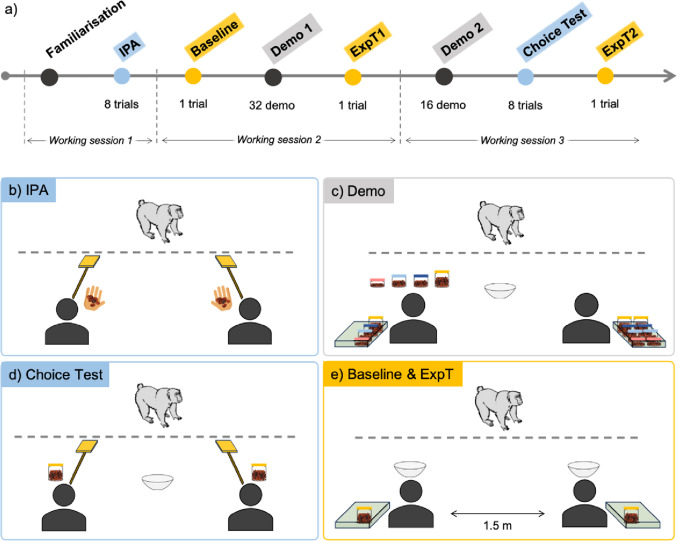 Figure 1
Figure 1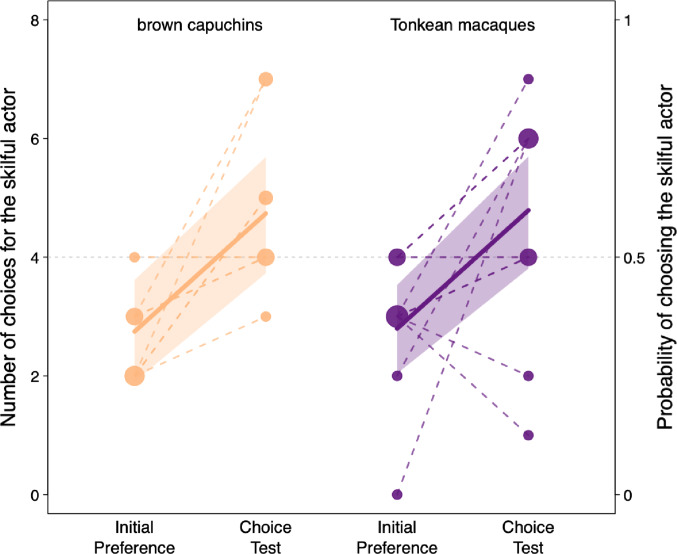 Figure 2
Figure 2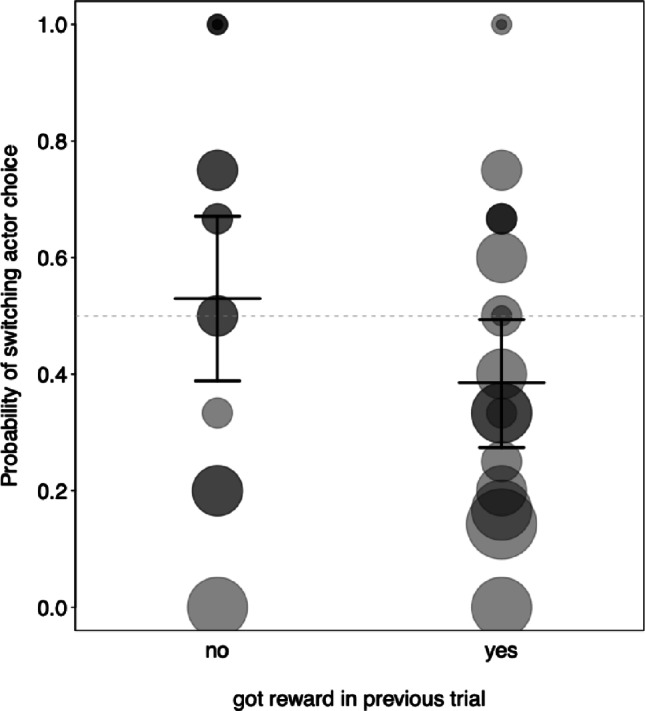 Figure 3
Figure 3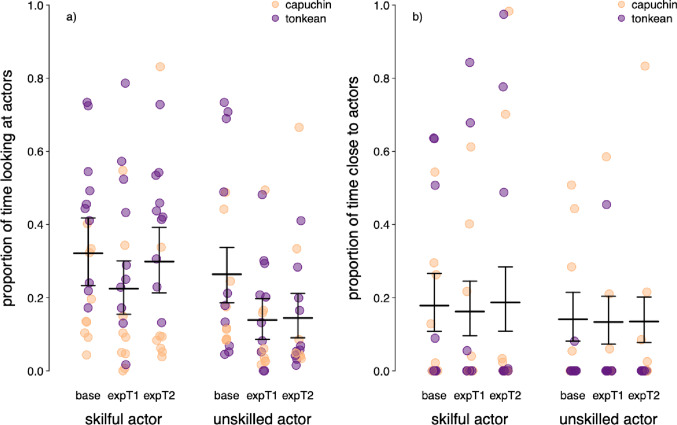 Figure 4
Figure 4- —Georg-August-Universität Göttingen (1018)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPrimate Behavior and Ecology · Animal Vocal Communication and Behavior · Hemispheric Asymmetry in Neuroscience
Introduction
Forming impressions about others from their past behaviours is a valuable cognitive skill, for example, in social contexts that require choosing interaction partners or role models (Corriveau et al. 2009; Rakoczy et al. 2009; Fusaro et al. 2011; Kushnir et al. 2013; Hermes et al. 2016, 2017; Wu et al. 2016). Most research has focused on evaluating others’ prosocial characteristics in nonhuman primates (Russell et al. 2008; Subiaul et al. 2008; Herrmann et al. 2013; Anderson et al. 2013a, b, 2016; Kawai et al. 2014) and a few other animal species (dogs and wolves: Kundey et al. 2011; Jim et al. 2022; fishes: Bshary and Grutter 2006; Vail et al. 2014; elephants: Jim et al. 2021; cats: Leete et al. 2020). Social evaluation of others’ skills (i.e., the performance of an individual in a particular task; Sih et al. 2019) is also essential social information to consider when deciding who to interact with. Individuals might benefit from observing and/or interacting with the skilful foragers of their group to access food resources or to learn new foraging skills (although skilful foragers may also represent a threat or competitor for food access, making interactions risky). Similarly, recruiting the most skilful partners seems advantageous for successful cooperation.
Some field experiments show that several nonhuman primates increased their affiliative behaviours toward the only group members skilled at providing food in a novel experimental foraging situation (Stammbach 1988; Fruteau et al. 2009; O’Hearn et al. 2025b; Karakoç et al. 2025). Others preferentially observed (Ottoni et al. 2005; Tan et al. 2018) or copied (Horner et al. 2010; Kendal et al. 2015; Canteloup et al. 2021) the most skilful group members at a foraging technique. However, only a few experimental studies investigated the ability to evaluate the skills of others in nonhuman species. After observing two human actors repeatedly trying to open containers filled with food, long-tailed macaques (Macaca fascicularis) preferentially chose (Placì et al. 2019), and dogs preferentially looked at or approached (Piotti et al. 2017; Chijiiwa et al. 2022) a skilful over an unskilled actor. Chimpanzees (Pan troglodytes) and coral trout (Plectropomus leopardus) strategically recruited the most skilful conspecific partners for cooperation (Melis et al. 2006; Vail et al. 2014; Keupp and Herrmann 2024).
These previous studies suggest that some animal species can consider the skills of others and behave differently towards individuals who differ in their degree of skilfulness. Yet, the cognitive processes underlying these behaviours are still unclear. Individuals can adapt their behaviour toward skilful foragers because they make inferences about the partner’s traits and attribute competence to them (i.e., impression formation process) or because they learn to associate the foragers’ actions with the resulting outcome (i.e., outcome-based process; for a review on social evaluation and underlying cognitive mechanisms, see O’Hearn et al. 2025b). For example, when recruiting conspecifics for cooperative tasks, chimpanzees appeared to engage in a Win-stay/Lose-shift strategy, making decisions based on the outcome of the previous test trial rather than on an evaluation of their partners’ skills (Melis et al. 2006). In addition, confounding factors such as individual differences other than skills, familiarity, or pre-existing relationships between the subjects and the individuals being assessed were not always considered in previous studies. To broaden our understanding of social evaluation in nonhuman primates, we investigated whether two monkey species, Tonkean macaques (Macaca tonkeana) and brown capuchins (Sapajus apella), can evaluate the skilfulness of others at making food accessible.
Both species live in multi-male multi-female societies characterised as tolerant (Thierry et al. 1994, 2004; De Waal 2000; Fragaszy et al. 2004; Thierry 2007; Riley 2010). They demonstrated social cognitive skills such as social relationships knowledge, attention and goal-directed understanding, and visual perspective-taking (Fragaszy et al. 2004; Phillips et al. 2009; Defolie et al. 2015; Canteloup et al. 2016, 2017; Canteloup and Meunier 2017; Whitehouse and Meunier 2020). They exhibit tool-using in foraging contexts (Anderson 1985; Fragaszy et al. 2004; Ducoing and Thierry 2005; Ottoni and Izar 2008), and brown capuchins even showed preferential attention toward skilful conspecific foragers (Ottoni et al. 2005). Tonkean macaques had never been tested on social evaluation before (except in Hirel et al. 2025a) while brown capuchins already demonstrated social evaluation of human actors’ helpfulness and reciprocity (Anderson et al. 2013a, b). Therefore, testing brown capuchins, a species for which we already have observational and experimental evidence of potential social evaluation abilities, allows us to better gauge the strengths and limits of our experimental paradigm by assessing both species’ results (e.g., methodological explanations or species-specific differences). While we hypothesised that both species could evaluate others’ skills and did not aim to compare species directly, any observed differences can provide valuable theoretical and methodological insights for future research.
In this experiment, subjects could sample information about two human actors’ skills from observation during two demonstration sessions. One actor (skilful) always succeeded at opening transparent containers filled with food, while the other actor (unskilled) always failed. Subjects received a food reward from a third person (the experimenter) once a container was successfully opened by the skilful actors, whereas they went empty-handed after each unsuccessful attempt by the unskilled actor. Then, the subjects completed a Choice Test and two Expectation Tests. In the Choice Test, we measured the proportion of choosing the skilful actor. We predicted that (1) both species would choose the skilful actor more often than the unskilled actor, and (2) shift their initial actor’s preference before the experimental manipulation in favour of the skilful actor as this became the better option. We also examined whether the subjects engaged in a Win-stay/Lose-shift (WS/LS) strategy, i.e., basing their decisions on the outcome of the previous trial. In the two Expectation Tests, we measured subjects’ anticipatory behaviours while the two actors simultaneously manipulated a food container. Preferential looking or spatial proximity towards the skilful actor would indicate anticipation of the successful task completion and that the subjects expected the outcome of the actors’ actions. We predicted that the subjects’ anticipatory behaviours would increase toward the skilful actor while decreasing toward the unskilled actor relative to the pre-demonstration phase. We further expected this effect to strengthen from Expectation Test 1 (conducted after the first demonstration session) to Expectation Test 2 (run after both demonstrations and the Choice Test), as subjects obtained more information about the actors’ skills.
Methods
Subjects and study site
This study was conducted at the Centre de Primatologie – Silabe de l’Université de Strasbourg (France) between July and September 2023. Nineteen subjects participated in the study including ten Tonkean macaques (four females; 13.7 ± 5.0 years old) from two different social groups and nine brown capuchins from one social group (five females; 8.3 ± 4.4 years old; Table S1). All of them were born and raised in captivity and live in wooded outdoor enclosures with constant access to indoor rooms (more details on subjects and housing can be found in the Supplementary Materials). Individuals were fed once per day with dry pellets and once per week with fruits and vegetables, and had access to water ad libitum. Subjects were tested individually in experimental rooms situated next to their outdoor enclosure (Figure S1). They participated in the experiment on a voluntary basis, which may have biased our sample of subjects towards dominants, individuals who are more comfortable in the experimental rooms and/or experienced in experimental testing (see STRANGE framework; Webster and Rutz 2020). The subjects had participated in several ethological studies, but they had never been tested with social evaluation experiments before (except for one Tonkean macaque in Hirel et al. 2025a; Table S1). One brown capuchin only completed Expectation Test 1 and three trials of the Choice Test but was included in the analyses as specified below.
Fig. 1(a) Chronological order of the experimental steps; Materials’ and actors’ disposition at the start of (b) each trial of the Initial Preference Assessment (IPA): the two actors presented a target in one hand and raisins on their palm of their other hand; (c) the demonstration sessions: the two actors sat in front of the mesh, each with a basket containing identical sets of baited containers next to them. The bowl was in the middle, the active demonstrator had placed four containers in front of them, which they would attempt to open in a row during one demo trial. In Demo 1, the actors alternately manipulated eight containers (i.e., two demo trials) on one side before switching position to manipulate eight containers on the opposite side. In Demo 2, the actors alternately manipulated four containers on each side; (d) Choice Test: the two actors sat in front of the mesh and presented their target, one container was placed in front of each actor, the bowl was in the middle; (e) Expectation Tests (Baseline, ExpT1, ExpT2): the two actors sat in front of the mesh with their basket and one container next to them, a bowl was placed in front of each actor, each actor manipulated one container. For each step, the actors sat at the same two predetermined locations in front of the mesh of the testing room, 1.5 m apart and with a distance from the mesh adjusted for the arm length of each subject
Table 1. Information about the different experimental stepsStepTypeMeasurePurposeAnalysesFamiliarisation1 session–Let subjects experience themselves the possible states of the containers and how they can be opened and closed–DemonstrationDemo 1 (8 obs/actor), Demo 2 (4 obs/actor)Looking timeGive subjects the opportunity to obtain information about the actors’ skills from observation–Initial Preference Assessment (IPA)1 session(8 trials)Actor choicesAssessing subjects’ initial preferences for the actors and assigning the roles of the actorsEstimation of the change in preference for the skilful actor from the IPA to the Choice TestChoiceTest1 session(8 trials)Actor choicesMeasuring the number of choices of the subjects for the skilful actorEstimation of the probability to choose the skilful actor according to trial numberExpectationTest3 trials of 20 s each: Baseline, ExpT1, ExpT2Looking & spatialproximity timeMeasuring the subjects’ time of looking at and being close to the actors before the demonstrations (Baseline), after Demo 1 (ExpT1), and after both demonstrations and Choice Test (ExpT2)Estimation of the effect of trial (Baseline, ExpT1, ExpT2) on the subjects’ time of looking at and/or in proximity to both actors
Procedure and design
The same two familiar humans played the roles of the skilful and the unskilled actors for all subjects. The task was to open and empty transparent containers filled with raisins into a transparent bowl. The skilful actor was always successful whereas the unskilled actor always failed. The actors’ roles were constant for each subject throughout the experimental procedure and were assigned according to subjects’ choices in the Initial Preference Assessment (IPA; see below). For each experimental session, both actors were present and sat at two predetermined locations in front of the experimental room, with their locations pseudo-randomised (i.e., an equal number of times on each side) within subjects and counterbalanced between subjects (for more details on the materials, actors’ locations, and procedure explained below, please refer to the Supplementary Materials).
The experiment consisted of eight steps divided into three ‘working’ sessions for each subject (Fig. 1a; Table 1). Each subject underwent up to one working session per half day (i.e., maximum two sessions per day). To increase the possibility for the subjects to evaluate the actors’ skills at manipulating the containers, the subjects experienced the possible states of the containers (i.e., open or closed) and the outcomes in their food intake (Kuroshima et al. 2014), and how a human can open and close the containers, during a familiarisation session before the test phase. In addition, as touching the target was the measure of subjects’ choices for the actors during the IPA and Choice Test, the experimenter ensured that each subject was able to choose the better of two options by touching targets before the experiment. Only subjects who met the learning criterion (i.e., choosing the better food option on at least ten out of 12 trials in a session) could participate in the study.
Initial preference assessment (IPA)
We conducted eight IPA trials before the experimental manipulation to assess whether subjects had a spontaneous initial preference for one of the actors, i.e., a personal preference unrelated to any experimental manipulation. First, the two actors fed the subject a few raisins, one after the other, to show the subject that both were willing to give food. Then, for each IPA trial, both actors held one raisin in the palm of their hand and a target in the other. Once the experimenter lured the subject to the central position equidistant to the actors, the actors presented their target simultaneously in front of them, and the experimenter stepped back. The subject could approach one of the actors to touch their target and obtain the food; this target touch was considered a choice for this actor (video S1). If the subjects chose each actor in four out of eight trials, the actors’ roles were randomly assigned and counterbalanced between subjects. If a subject chose one actor five times or more out of the eight trials, we assigned to this ‘preferred’ person the role of the unskilled actor for this subject. We chose this conservative role assignment rule to avoid a confound of initial preference for an actor and choice based on skill demonstrations of the actor.
Baseline
We measured the baseline for subjects’ anticipatory behaviours before they acquired any information about the skilfulness of the actors during the demonstration sessions. The two actors had one identical closed baited container next to them and one transparent bowl in front of them (Fig. 1e). Once the experimenter lured the subject to the central position, the actors started simultaneously manipulating their container while the experimenter stepped back (video S5). After 20 s of manipulation, the skilful actor successfully opened their container and emptied it into their bowl while the unskilled actor tried in vain to empty their still closed container into their bowl. Both actors then simultaneously put their containers into their baskets and looked down. The experimenter first took the empty bowl in front of the unskilled actor to show it to the subject, then took the full bowl in front of the skilful actor to feed the subject with a few raisins. During the duration of the trial, the actors focused only on the containers and never looked at the subject.
Demonstration sessions
Each actor alternately demonstrated their skill to open four containers in a row in one location (subsequently referred to as one demo trial). One transparent bowl was in the middle equidistant from the actors (Fig. 1c). In Demo 1 session, each actor had eight closed baited containers and alternately demonstrated twice in the same location (e.g., one skilful-left demo trial, one unskilled-right, one skilful-left, and one unskilled-right). The actors then repeated the same procedure but at the opposite location. In Demo 2 session, the procedure was the same except that the actors held four containers and only one demo trial per location was performed by each actor. We limited Demo 2 session to two demo trials so that subjects could be tested immediately afterward with the Choice Test and Expectation Test 2, while keeping the working session short enough to maintain subjects’ motivation. Therefore, the subjects observed each actor manipulating 16 containers (eight containers per side) during Demo 1 and eight containers (four containers per side) during Demo 2, resulting in a total of 24 observations per actor for each subject.
A demo trial started with an actor placing four containers aligned in front of them and then presenting their target. In the meantime, the other actor stayed head down without moving until the end of the demo trial. Once the subject had touched the target, the actor started manipulating the four containers. After trying for around five seconds, the skilful actor successfully opened and emptied each container whereas the unskilled actor failed and tried in vain to empty the closed container into the transparent bowl. After manipulating their fourth container, the actor stopped moving and looked down. The experimenter took the transparent bowl which was either filled with food (after a skilful actor’s manipulation) or empty (after an unskilled actor’s manipulation). The experimenter then fed the subject some of the food from the bowl or showed the empty bowl to the subject, before replacing the bowl with a new identical empty one (video S2). We deemed it necessary to reward the animals with small quantities of food, but not directly from the skilful actor; otherwise, they might have lost the motivation to participate in the experiment.
Choice test
The subjects chose in eight trials which of the two actors could manipulate a container to release food. A trial began with the two actors placing one identical closed baited container in front of them and holding a target in their hand. One transparent bowl was in the central position, equidistant to the actors. Once the experimenter lured the subject to the central position, the actors presented their target simultaneously while the experimenter stepped back (Fig. 1d). The subjects indicated their choice by touching the target of one actor. The unchosen actor removed their target, put their container back, and waited with their head down until the end of the trial. Once chosen, the skilful and unskilled actors followed the same actions with their container as during the demonstration trials. The experimenter then fed the subject some of the food from the bowl (after the skilful actor’s manipulation) or showed the empty bowl to the subject (after the unskilled actor’s manipulation; videos S3 and S4). The subjects were given a maximum of 20 s to make a choice, otherwise the trial was aborted. A trial (with the same actors’ location configuration) was repeated a maximum of three times before aborting the session (which never happened).
Expectation test
We measured subjects’ anticipatory behaviours toward the actors in two trials, after subjects had information about the actors’ skills from Demo 1 (ExpT1) and from both demonstrations and the Choice Test (ExpT2; Fig. 1a). The procedure for ExpT1 and ExpT2 was exactly the same as for the Baseline (videos S5 and S6). The actors’ location was the same for the three trials for one subject, but counterbalanced between subjects.
Data coding
All experimental steps were videotaped with three GoPro9 cameras to obtain different views of the subjects, the actors, setups, and experimental rooms. All the videos were coded frame by frame by an observer using Behavioral Observation Research Interactive Software (BORIS v.8.20; Friard and Gamba 2016). A second observer, who was unaware of the study design and hypothesis, coded independently 38 videos which were pseudo-randomly selected to include 20% of the sessions for each combination of phase, species, and subjects, giving a relatively representative sample of each behaviour coded. Inter-coder reliability scores ranged from 63% to 100% (see Supplementary Materials for details). For the IPA and Choice Test, videos were coded for: (a) the choices of the subjects toward either actor and (b) the synchronicity of targets’ presentation by the actors. For the Expectation Tests and the demonstration sessions, videos were coded for: (a) the duration of looking at either actor and (b) the amount of time subjects spent in close range to either actor (1 m radius), in nearby range, or far away (Figures S1 and S2). The exact definitions of the coded behaviours are reported in Table S2. Both Tonkean macaques and brown capuchins paid attention to the actors’ actions during the demonstrations around half the time (Table S3). As a measure of the subjects’ attention to both actors during the demonstration sessions that we included in the analyses, we chose the minimum looking time per subject to the skilful and unskilled actor, i.e., the time during which each subject observed each actor.
Data analyses
Choices
We assessed whether subjects chose the skilful actor more often than the unskilled actor in the Choice Test and whether this probability of choosing the skilful actor increased with direct experience (i.e., trial number), by fitting a Generalized Linear Mixed model (GLMM; Baayen 2008) with logit link function (McCullagh and Nelder 1989) and a binomial error structure. The sample included 147 observations (i.e., trials) from 19 subjects (including the brown capuchin subject who completed only the first three trials of the Choice Test session). The response variable for each trial was 1 (choice of the skilful actor) or 0 (choice of the unskilled actor). The fixed-effect predictors we included in this model were trial number (main predictor), species (capuchin, Tonkean), the attention directed at the actors during the demonstration, and the three-way interaction between these predictors. This three-way interaction was included to account for the possibility that species differences, as well as individual variation in attention during the demonstration, could influence trial learning curves, and that the two species may require different amounts of time to acquire comparable levels of information about the actors’ skills. To control for their potential effects, we included the sex of the subjects, the side location (left, right), and the identity (A, B) of the skilful actor. Although we did not have a specific expectation regarding potential sex differences in social evaluation abilities, we decided to include sex as a control factor, given previous findings showing sex differences in chimpanzees and dogs (Watson et al. 2018; Chijiiwa et al. 2022). An additional control predictor, synchronicity of targets’ presentation (−1: unskilled actor first, 0: synchronous, + 1: skilful actor first), was added to the model. During video coding, we noticed that the two actors were not always fully synchronous which could have affected the choices of the subjects.
To account for individual differences, avoid overconfident model estimates, and keep type I error rate at the nominal level of 5%, we included subject ID as a random intercept effect and all identifiable random slopes within subject, which were trial number, side location of the skilful actor, and synchronicity of targets’ presentation (Schielzeth and Forstmeier 2009; Barr et al. 2013). As an overall test of the fixed effects and to avoid cryptic multiple testing (Forstmeier and Schielzeth 2011), we compared this full model with a null model lacking the effect of trial number and its interactions but being otherwise identical, using likelihood ratio tests (Dobson and Barnett 2018). To test the impact of individual fixed effects, we conducted likelihood ratio tests (Dobson and Barnett 2018) that compared the full models with reduced models, each lacking one fixed effect at a time (Barr et al. 2013). We obtained confidence intervals of model estimates and fitted values using a parametric bootstrap (N = 1000 bootstraps). We checked all the relevant model assumptions and transformed some variables when needed to ease the interpretation of the model estimates and convergence. This procedure of assumption checks applies to all analyses (see Supplementary Materials for more details on each model analysis).
Shift in preference for the skilful actor
Regardless of whether the probability of choosing the skilful actor during the Choice Test differed from chance (50%), subjects might have chosen the skilful actor more often in the Choice Test than during the IPA. Therefore, we fitted a similar model except that we estimated the change in preference for the skilful actor from the IPA to the Choice Test. The sample included 288 observations (i.e., trials) from 18 subjects (excluding the brown capuchin subject who did not complete the Choice Test). The response variable was a matrix comprising the number of choices for the skilful and unskilled actors for each subject and phase. This model included the test phase (IPA, Choice Test) as the main predictor and species (capuchin, Tonkean). To account for random individual differences, avoid overconfident model estimates, and keep the type I error rate at the nominal level of 5%, we included subject ID as a random intercept effect and the only theoretically identifiable random slope phase within the subject.
Alternative WS/LS strategy
The subjects could have used alternative strategies rather than sampling and evaluating the actors’ skills. Like chimpanzees in the study by Melis et al. (2006), they could engage in a WS/LS strategy by tracking their success with each of the actors and basing their decisions on the outcome of the previous trial. We ran a post-hoc GLMM analysis to estimate the effect of positive outcomes on the probability of staying with or switching actor choices during the Choice Test. For each trial, we coded whether the subjects stayed with or switched their choice of actors from the previous trial (variable ‘strategy’) and whether they had chosen the skilful actor and obtained a food reward in the previous trial (variable ‘reward’). The sample included seven trials per subject (trials 2 to 8), resulting in 126 trials in total. In the model, we included strategy (stay, switch) as the response variable, reward (yes, no) as a fixed effect, and species as a control predictor. We also included subject ID as a random intercept effect to account for random individual differences, to avoid overconfident model estimates, and to keep the type I error rate at the nominal level of 5%. If subjects follow the WS/LS strategy, they should base their decisions on the outcomes in the previous trial. We would observe more “stay” decisions after successes than failures and more “switch” decisions after failures than successes.
Anticipatory behaviours
We assessed whether subjects would preferentially look or be close to the skilful actor while both actors simultaneously manipulated a baited container for 20 s. We wanted to estimate the effect of trial (Baseline, ExpT1, and ExpT2) on the subjects’ looking time and their proximity to both actors, as subjects gained more indirect and direct information about the actors’ skills. We define direct information as information obtained through direct interactions with the actors, and indirect information as information acquired solely through observation of the actors. As the response variables were bound between 0 and 20 s, we turned them into proportions to rule out fitted values and confidence intervals of fitted values extending beyond the possible response range. We fitted two GLMMs with a beta error distribution (McCullagh and Nelder 1989) and with identical structure except for the response variable, which was either the proportion of time looking at or the proportion of time spent in proximity to either actor. The sample for each model included 112 observations from 19 subjects and 56 levels of trial nested in subject (including the brown capuchin subject who completed only the Baseline and ExpT1). In these models, we included actor (skilful, unskilled), trial (Baseline, ExpT1, ExpT2) and their interaction as the main predictors. To control for their potential effects, we also included species, the sex of the subjects, and the identity of the skilful actor.
To account for random individual differences, avoid overconfident model estimates and keep the type I error rate at the nominal level of 5%, we included subject ID as a random intercept effect and all theoretically identifiable random slopes (Schielzeth and Forstmeier 2009; Barr et al. 2013), which were trial and actor within subject. In addition, we included the random intercept effect of trial nested in subject to account for the fact that the data for the skilful and unskilled actor for any given combination of subject and trial were not independent. Following the same model comparison procedure as before, we compared these full models with null models lacking the effects of trial, actor, and their interaction but being otherwise identical, and with reduced models, each lacking one fixed effect at a time, using likelihood ratio tests.
R functions and packages
We conducted statistical analyses and created plots using R (version 4.3.2; R Core Team 2022). We used the function glmer of the package lme4 (version 1.1–35.1; Bates et al. 2015) to fit the logistic models, and the function glmmTMB of the homonymous package (version 1.1.8; Brooks et al. 2017) for the models with beta distribution. Parametric bootstraps were obtained using the functions bootMer (package lme4) and simulate (package glmmTMB).
Results
Choices
In the Choice Test, subjects chose the skilful actor 85 times out of 144 (brown capuchins: 39 choices, 60.9%; Tonkean macaques: 46 choices, 57.5%) and they chose the unskilled actor 59 times out of 144 (brown capuchins: 25 choices, 39%; Tonkean macaques: 34 choices, 42.5%; excluding the brown capuchin who did not complete the session). Twelve subjects (six out of nine brown capuchins and six out of ten Tonkean macaques) chose the skilful actor in the first trial (Table S1). Overall, their probability of choosing the skilful actor in the Choice Test was not influenced by any of the predictor terms (full-null model comparison: χ^2^ = 7.345, df = 4, p = 0.119; Table 2). This result indicates that subjects did not choose the skilful actor more often than expected by chance, nor did they increase their choices for the skilful actor over the trials.
Table 2. Results of the subjects’ choices full model in the Choice Test (estimates together with standard errors, 95% confidence limits, significance tests, and the estimates range obtained when dropping levels of grouping factors one at a time)TermEstimateSECL_lower_CL_upper_χ^2^df P minmax(Intercept)−2.7450.999−7.788−1.196 ^1^ −3.2−2.269Species1.5571.107−0.736 ^1^ 0.9351.882Trial2.6021.3120.1379.169 ^1^ 2.0733.506Attention−1.9650.863−6.074−0.455 ^1^ −2.148−1.32Sex1.2390.6860.2293.9773.25610.0710.7452.054Side skilful1.8080.7140.4765.3736.51210.0111.4582.083Target sync0.170.284−0.5391.0730.36210.5470.0180.306ID skilful0.1880.72−1.2582.1860.07010.791−0.250.577Species × Trial−2.1481.689−9.0741.598 ^1^ −3.172−1.447Species × Attention2.1561.124−0.0436.837 ^1^ 1.552.531Trial × Attention2.5681.3040.0038.72 ^1^ 1.8863.263Species × Trial × Attention−3.8751.765−11.708−0.5645.26510.022−4.677−2.976Trial number was transformed to range from 0 to 1 (originally from 1 to 8). Attention and target synchronicity were z-transformed to a mean of 0 and a standard deviation of 1 (original means and standard deviations were 98.9 ± 30.56 s and 0.39 ± 0.57 s, respectively). Species, sex, side location, and identity of the skilful actor were dummy coded with brown capuchins, female, left, and actor A being their respective reference level^1^ not indicated because of very limited interpretability
The model revealed a significant three-way interaction between trial, species, and the subjects’ attention during the demonstration phase (p = 0.022; Table 2 and S3, Figure S3), possibly due to multiple testing. There were no significant effects of sex (p = 0.071), synchronicity in target presentation (p = 0.547), or the identity of the skilful actor (p = 0.791; Table 3). Success probability was higher when the skilful actor was on the right-hand side (p = 0.011; Table 2). In the Choice Test, subjects chose the left side 34.7% of the time and the right side 65.3% of the time, and four subjects chose the right side in at least seven out of the eight trials. During the IPA, subjects chose the left side 30.9% of the time and the right side 69.1% of the time. These results indicate a general preference for the right side from the start of the experiment, which may have interfered with the subjects’ choices during the Choice Test.
Shift in preference for the skilful actor
Thirteen out of the 18 subjects increased their number of choices for the skilful actor from the IPA to the Choice Test, with seven of them choosing the skilful actor in at least six out of the eight trials of the Choice Test (Table S1; Fig. 2). The model revealed a strong effect of phase (p < 0.001; Table 3), with the probability of choosing the skilful actor increasing from 34.7% in the IPA to 59% in the Choice Test (excluding the brown capuchin’s choices who did not complete the Choice Test; Fig. 2). The model revealed no species differences in this shift in actors’ preference (p = 0.917).
Table 3. Results of the full model for the comparison of subjects’ choices between the IPA and the Choice Test (estimates together with standard errors, 95% confidence limits, significance tests, and the estimates range obtained when dropping levels of grouping factors one at a time)TermEstimateSECL_lower_CL_upper_χ^2^df P minmax(Intercept)−0.6480.23−1.162−0.193−0.714−0.571Phase1.0210.2860.4821.59110.22810.001*0.8911.127Species0.0280.266−0.5220.5860.01110.917−0.0640.11The reference levels for phase and species were respectively IPA and brown capuchins
Fig. 2. Probability of choosing the skilful actor during the Initial Preference Assessment (IPA) and the Choice Test for brown capuchins and Tonkean macaques. The continuous lines and the shaded areas depict the fitted model and its 95% confidence intervals. Points depict observations; the area of the point is proportional to the number of observations (range: 1 to 5). Observations from the same subject are connected by dashed lines
Alternative WS/LS strategy
After an unsuccessful trial in the Choice Test (receiving no reward), the subjects stayed with the same actor choice in 23 out of 50 trials (46%) while switched in 27 out of 50 trials (54%). After a successful trial in the Choice Test (receiving a reward), the subjects stayed with the same actor choice in 46 out of 76 trials (60.5%) while switched in 30 out of 76 trials (39.5%). The model revealed a non-significant effect of reward (p = 0.115) and no species differences in the probability of using the WS/LS strategy (p = 0.709; Table 4). Even though the subjects tended to stay with the same actor more often after a win than after a loss, they did not switch more often after a loss than after a win (Fig. 3). These results indicate that our subjects did not base their decisions on the outcomes they experienced in previous trials with a WS/LS strategy.
Table 4. Results of the full model for subjects’ Win-Stay/Loose-Switch (WS/LS) strategy (estimates together with standard errors, 95% confidence limits, and the estimates range)TermEstimateSECL_lower_CL_upper_χ^2^df P minmax(Intercept)0.1980.356−0.5220.9220.0310.437Reward−0.5850.374−1.3750.1462.48810.115−0.905−0.402Species−0.1420.380−0.8980.6860.14010.709−0.3010.041Reward was dummy coded with its reference level being no reward obtained in previous trial. The reference level for species was brown capuchins
Fig. 3. Probability of subjects switching their actor choices depending on the outcome in previous trial, i.e., whether the subjects received a reward or not. Points depict observations; the area of the point is proportional to the number of observations
Anticipatory behaviours
Overall, the proportion of time looking at the actors during the Expectation Tests was significantly influenced by trial and the actors’ role (full-null model comparison: χ^2^ = 18.5, df = 5, p = 0.002; Table 5). During ExpT1 and ExpT2, with all other factors being averaged, subjects looked longer at the skilful (ExpT1: 4.99 s; ExpT2: 6.44 s) than the unskilled actor (ExpT1: 2.93 s; ExpT2: 2.91 s; Fig. 4 and Table S4). They looked less at the unskilled actor during ExpT1 and ExpT2 than in the Baseline (5.42 s) but did not spend more time looking at the skilful actor than in the Baseline (6.52 s; Fig. 4 and Table S4). The model revealed significant effects of species (p = 0.005; Table 5 and S4; mean looking time: 12.3 s for Tonkean macaques and 6.81 s for brown capuchins), and of the skilful actor’s identity (accounting for a difference of about 1.3 s in looking time; p = 0.031), while the effect of sex was not significant (p = 0.097; Table 5 and S4).
Overall, the proportion of time spent in proximity to the actors during the Expectation Tests was not influenced by any of the predictors (full-null model comparison: χ^2^ = 2.28, df = 5, p = 0.808; Table S5). The proportion of time spent close to the actors was generally low and only slightly higher close to the skilful actor (ExpT1: 2.99 ± 5.44 s; ExpT2: 4.43 ± 7.47 s) than close to the unskilled actor (ExpT1: 1.38 ± 3.35 s; ExpT2: 1.29 ± 3.98 s; Table S4). However, subjects did not spend time close to each actor equally at random in the Baseline (skilful: 3.28 ± 4.78 s, unskilled: 1.44 ± 3.15 s), which may have interfered with the test results afterwards.
Table 5. Results of the full model for the subjects’ looking time during the Expectation Test (estimates together with standard errors, 95% confidence limits, significance tests, and the estimates range obtained when dropping levels of grouping factors one at a time)TermEstimateSECL_lower_CL_upper_χ^2^df P minmax(Intercept)−1.0720.271−1.638−0.585 ^1^ −1.204−0.968Phase ExpT1−0.4910.255−1.023−0.008 ^1^ −0.595−0.365Phase ExpT2−0.1060.250−0.5950.391 ^1^ −0.2800Actor role−0.2770.317−0.9410.310 ^1^ −0.431−0.112Species0.6220.1970.2591.0487.84210.005**0.5080.821Sex0.3290.199−0.0500.7462.75710.0970.1720.423ID skilful−0.4520.196−0.841−0.0674.66010.031*−0.614−0.362Phase ExpT1 × actor role−0.3090.365−1.0010.4383.00720.222−0.448−0.167Phase ExpT2 × actor role−0.6500.369−1.3620.115 ^2^ −0.876−0.442Phase, actor role, species, sex and ID of the skilful actor were dummy coded with their reference level being respectively baseline, skilful, brown capuchins, female, and actor A^1^ not indicated because of very limited interpretability^2^only one p-value because the interaction was tested as a whole
Fig. 4. Anticipatory behaviours of the subjects during the Expectation Tests. Plot (a) shows the effect of the interaction between trials and actors’ roles on the proportion of time looking at the skilful and unskilled actors, while plot (b) shows the effect of the interaction between trials and actors’ roles on the proportion of time spent in proximity to the actors. All proportions were calculated from a total duration of 20 s. Points depict single observations (n = 112). Observations for brown capuchins and Tonkean macaques are shown in yellow and purple, respectively. The horizontal lines and error bars depict fitted values and their 95% confidence intervals for all non-plotted predictors being averaged. Base: Baseline; expT1: Expectation trial 1; expT2: Expectation trial 2
Discussion
We investigated whether Tonkean macaques and brown capuchins can evaluate the skilfulness of human actors. During two demonstration sessions, subjects observed a skilful actor who consistently succeeded in opening transparent food containers and an unskilled actor who consistently failed, with rewards provided by the experimenter only following the skilful actor’s manipulations. Then, subjects completed a Choice Test and two Expectation Tests designed to assess their evaluation of the actors’ skills through different behavioural measures. When choosing which of the two actors they wanted to manipulate a container to extract food (Choice Test), subjects’ choices for the skilful actor did not differ from chance as a group. However, Tonkean macaques and brown capuchins successfully shifted their initial preference for the actors in favour of the skilful actor when this switch allowed them to maximise reward outcomes in the Choice Test. When both actors simultaneously manipulated their containers (Expectation Tests), subjects looked significantly longer at the skilful actor than the unskilled actor. They did not increase their time spent looking at the skilful actor relative to the baseline session but significantly decreased their time looking at the unskilled actor. This decrease in looking time did not become more pronounced after subjects completed the two demonstration sessions and the Choice Test (Expectation Test 2), compared with after only one demonstration session (Expectation Test 1). Unlike their looking pattern, however, subjects did not spend more time near the skilful actor than the unskilled actor, nor did their proximity patterns change across the Expectation Tests.
Impression formation or outcome-based process?
Like other primate species (Stammbach 1988; Fruteau et al. 2009; Kendal et al. 2015; Canteloup et al. 2021; Keupp and Herrmann 2024; O’Hearn et al. 2025b; Karakoç et al. 2025), Tonkean macaques and brown capuchins in our study adapted their behaviour in relation to others’ competence levels. The experimental manipulation – i.e., the actors’ demonstrations, which subjects observed around half of the time – induced a shift in their initial choice preference toward the skilful actor and a decrease in attention to the unskilled actor. The looking pattern in the Expectation Tests is unlikely to be explained by a food-driven attention, as the two actors had the same amount of food in their containers, and we measured the subjects’ looking time during the 20 s before the skilful actor released the food. In addition, the absence of an increase in choices for the skilful actor through trials and a WS/LS strategy suggests that these monkeys did not base their decisions on the outcomes they experienced during the Choice test. These findings may reflect an ability to assess the actors’ skills based on prior observation.
Yet, our design does not allow us to disentangle impression formation from outcome-based processes. During the demonstrations, subjects received small amounts of food from the experimenter following the skilful actor’s manipulations, but not after the unskilled actor’s manipulations. This procedure was a compromise between not giving them any food at all, which would facilitate a clear interpretation of the results but considerably reduce the subjects’ level of motivation, and allowing the actors to give food. Having the experimenter, rather than the skilful actor, provide the food helped maintain the subjects’ motivation and attention, although it resulted in a different reward history following one type of demonstration. Consequently, subjects may have learned to associate the actors’ behaviours with the outcomes they experienced (i.e., receiving food or not), without necessarily forming impressions of the actors’ skills (associative learning; Heyes 2012). Social evaluation based on impression formation involves cognitive inferences – forming abstract representations of others’ characteristics and using these to infer their future behaviours (O’Hearn et al. 2025a). In addition, obtaining rewards indirectly from the experimenter may have weakened the perceived contingency between the actors’ actions and resulting outcomes, thereby making social evaluation more difficult. This aspect of the procedure may also have biased subjects’ attention during both the demonstration and test phases toward the experimenter rather than the actors. Therefore, in our study, as in previous work in which nonhuman animals experienced differential outcomes through direct interaction with others (i.e., direct information sampling; Melis et al. 2006; Carballo et al. 2015; Tan et al. 2018), the underlying mechanisms remain unclear.
Which behavioural measures to assess social evaluation?
The discrepancies observed across our behavioural measures raise questions about which behaviours best capture social evaluation and whether all measures equally reflect it. Previous studies showed preferential attention of primates and dogs toward skilful than unskilled individuals (Ottoni et al. 2005; Tan et al. 2018; Chijiiwa et al. 2022). In our study, we observed a differential attention toward the actors characterised by a decrease in looking time at the unskilled actor. Both Tonkean macaques and brown capuchins paid less attention to the unskilled actor but did not increase attention to the skilful one. This pattern parallels the ‘negativity bias’ reported in brown capuchins, dogs, and children who showed an avoidance of antisocial over preferential choices to prosocial individuals (see Chijiiwa 2021 for a review). As in previous studies, testing choices between skilled and unskilled versus neutral actors (e.g., an actor with containers but not trying to open them) would help clarify whether this bias similarly drove our subjects’ shift in choices. However, alternative explanations for this decrease in looking time, such as habituation to the task or a decline in motivation over time, cannot be excluded. Importantly, whether social attention is a driver, a consequence of social evaluation, or both, and whether it is directly linked to social decisions, remains unclear at this point. Notably, recent large-scale research with infants evidenced that active measures, such as choice behaviour, may more reliably capture social evaluation than looking time (Lucca et al. 2025).
Our findings on choice and spatial proximity contrast with previous results. Chimpanzees preferred to approach and spent time near a prosocial than an antisocial human actor (Russell et al. 2008), and dogs show similar preferences for skilful or prosocial experimenters (Kundey et al. 2011; Marshall-Pescini et al. 2011; Carballo et al. 2015; Chijiiwa et al. 2022). While choice behaviour is the most common measure of social evaluation, results across studies are mixed. Some studies report preferences for prosocial or skilful individuals (Subiaul et al. 2008; Anderson et al. 2013b, 2016; Placì et al. 2019; Keupp and Herrmann 2024), whereas others, including ours, find that choices did not differ from chance (Kawai et al. 2019; Jim et al. 2021, 2022). Methodological factors may contribute to this variability. The number of choice trials, for instance, ranges widely across studies (e.g., 8 in ours vs. 144 trials in Anderson et al. 2013b, 2016). Our subjects also exhibited a right-side bias from the start of the experiment, which may have interfered with their choice probability. This bias could reflect the testing environment (e.g., the trapdoor to access the experimental room from the park of the brown capuchins was on the right side) or reduced interest in the task. Although given that the side of the successful choice alternated pseudo-randomly within sessions (i.e., an equal number of times on each side), a side bias represents a very efficient and less cognitively demanding strategy to obtain half of the rewards.
In addition, the social agents to evaluate are sometimes conspecifics, sometimes humans – with both significant and null effects reported. Fully controlled conspecific-based paradigms are methodologically challenging, e.g., controlling pre-existing social relationships, standardising demonstrations, and counterbalancing roles. Alternatively, testing nonhuman animals with human actors may reduce social relevance and their engagement in the task. Evaluating heterospecifics may also require distinct or more demanding cognitive processes, limiting interpretations of social evaluation. Spatial proximity measure may be particularly irrelevant in this case and not a reliable proxy for social expectation. Furthermore, several studies lacked controls for initial preferences and may have then wrongly assumed that subjects’ initial choices before the sampling phase were at chance level – particularly with demonstrators familiar to the subjects. Initial preference assessment and baseline tests also allow very informative and direct measurement of preference and behavioural change – an approach potentially more comparable to observed behavioural shifts toward skilled conspecifics in wild populations (O’Hearn et al. 2025b; Karakoç et al. 2025).
Finally, the extent to which nonhuman animals use evaluations of others’ characteristics to guide social decisions remains unclear. Social evaluation could inform both active choices (e.g., interaction, approach, or avoidance) and more indirect behaviours (e.g., observation, spatial proximity). However, across species, social evaluation may guide behaviours depending on the species-specific relevance of the context and the trait being assessed. For instance, successful cooperation can require selecting partners based on perceived willingness to cooperate, competence, or prosociality (Wu et al. 2016; Manrique et al. 2021) and social learning can benefit from merely observing competent or knowledgeable models (Laland 2004; Camacho-Alpízar and Guillette 2023). Our study context and task, which did not involve learning or cooperation with conspecifics, did affect subjects’ anticipatory-looking but not their active choices for the actors. Whether this difference in our behavioural measures reflects a context-specific effect on behavioural display of social evaluation warrants further investigation.
Species differences in social evaluation
During both the Demonstration and Expectation Test sessions, Tonkean macaques generally looked longer at the actors than the brown capuchins. Interspecific differences in preferential choices for prosocial over antisocial agents have also been reported previously (Russell et al. 2008; Herrmann et al. 2013; Kawai et al. 2019). Although we did not predict differences between the two species we tested, variation in how species acquire and use social information may explain these results and raises questions about the relevance of behavioural measures across taxa. Strategies for social information acquisition and use likely vary between species with different social organisations (e.g., Scheid et al. 2007; Range et al. 2009; Faraut and Fischer 2019). In large groups, direct information sampling can be time-consuming – interacting with each group member requires a lot of time that is not devoted to other activities – while indirect information sampling (e.g., looking, hearing) may be more efficient or safer for less tolerant species, as direct interactions can be associated with a high risk of getting injured. These social factors can similarly affect how species use their social evaluation (e.g., by preferentially observing rather than directly interacting with others). However, since both Tonkean macaques and brown capuchins live in similar group sizes and are characterised as socially tolerant species (Fragaszy et al. 2004; Thierry et al. 2004), group size or tolerance alone are unlikely to account for the observed differences in looking behaviour.
Differences in cognitive processing may offer an alternative explanation, with some species or individuals picking up on informational cues faster than others. The amount of social information needed to form impressions of others is unknown. In our study, looking time toward the unskilled actor decreased after a single demonstration session (32 container manipulations, 16 per actor), whereas no choice preference for the skilful actor (but a shift in preference) was observed. Notably, some species – including Tonkean macaques – did not demonstrate the use of social information about others’ skills or prosociality without direct experience (Nitzschner et al. 2012; Jim et al. 2021; Hirel et al. 2025a, b), whereas chimpanzees and orangutans did (Russell et al. 2008; Subiaul et al. 2008; Herrmann et al. 2013; Keupp and Herrmann 2024). These findings suggest possible species differences in social evaluation abilities, underlying cognitive mechanisms, or strategies for acquiring social information.
Conclusion
Tonkean macaques and brown capuchins did not choose the skilful actor more often than the unskilled actor, but they successfully shifted their initial preference in favour of the skilful actor. They neither learned through choice trials nor based their choices on previous trials outcomes. Both species looked more at the skilful over the unskilled actor when both actors simultaneously manipulated a container, seemingly due to a decrease in attention to the unskilled actor, but did not change their time spent near either actor. These results suggest that these monkeys may have used social information about the actors’ actions acquired from the demonstration sessions to adapt their behaviour. Yet, because subjects experienced differential rewarding during demonstrations, it remains unclear whether they formed impressions of the actors’ skills or based their decisions on previous outcomes. The mechanisms underlying looking behaviour and the measures that most accurately reflect social evaluation in nonhuman animals also deserve to be clarified. We encourage further research across diverse taxa to explore the cognitive processes underlying social information acquisition in nonhuman animals and the social environments that promote its emergence.
The methodological conundrum in balancing the absence of rewards with maintaining subjects’ motivation, which we faced in this study, highlights one of the challenges of investigating social evaluation in nonhuman animals, as sometimes it is not possible to rely exclusively on indirect information sampling to seed the experimental manipulations. Future studies should also carefully consider methodological design, for instance by comparing direct versus indirect information sampling, using conspecifics rather than human actors, controlling for initial preferences or behavioural baselines, and testing across multiple social contexts (e.g., cooperation, competition, social learning) using different behavioural measures (e.g., choice, looking, proximity). The approaches developed by Hirel et al. (2025a) and Keupp and Herrmann (2024) in Tonkean macaques and chimpanzees offer promising paradigms in this regard, adaptable to different species and contexts.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.