Molecular identification of Borrelia and Rickettsia in hard ticks infesting domestic and wild animals in Cameroon
Archile Paguem, Pierre Kamtsap, Kingsley Tanyi Manchang, Alfons Renz, Sabine Schaper, Gerhard. Dobler, Robert E. Rollins, Lidia Chitimia-Dobler

TL;DR
This study identifies Borrelia and Rickettsia in ticks from Cameroon, highlighting potential public health risks from these tick-borne pathogens.
Contribution
First detection of Borrelia spp. in ticks from wild animals in Cameroon and identification of three Rickettsia species in domestic animal ticks.
Findings
18.01% of tick pools tested positive for Rickettsia DNA.
10.38% of tick pools tested positive for Borrelia DNA.
Borrelia spp. in Cameroon are closely related to species from China and the Ivory Coast.
Abstract
Ticks are blood-sucking arthropods which can vector various, pathogenic microorganisms between humans and domestic or wild animal hosts. In Cameroon, little is still known about the diversity of ticks and tick-borne pathogens found feeding on these various hosts. This study investigates the frequency of positive pools of Borrelia spp. and Rickettsia spp. in 415 DNA pools arising from 1148 collected ticks belonging to five genera and twenty-five tick species collected from both domestic and wild animals in Cameroon. Tick species were identified morphologically and confirmed molecularly when necessary. All tick pools were tested for Rickettsia spp. and Borrelia spp. using molecular methods of which 18.01 % and 10.38 % of tick pools tested positive for Rickettsia or Borrelia DNA, respectively. This is the first Borrelia spp. detection in ticks collected from wild animals in Cameroon. Three…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
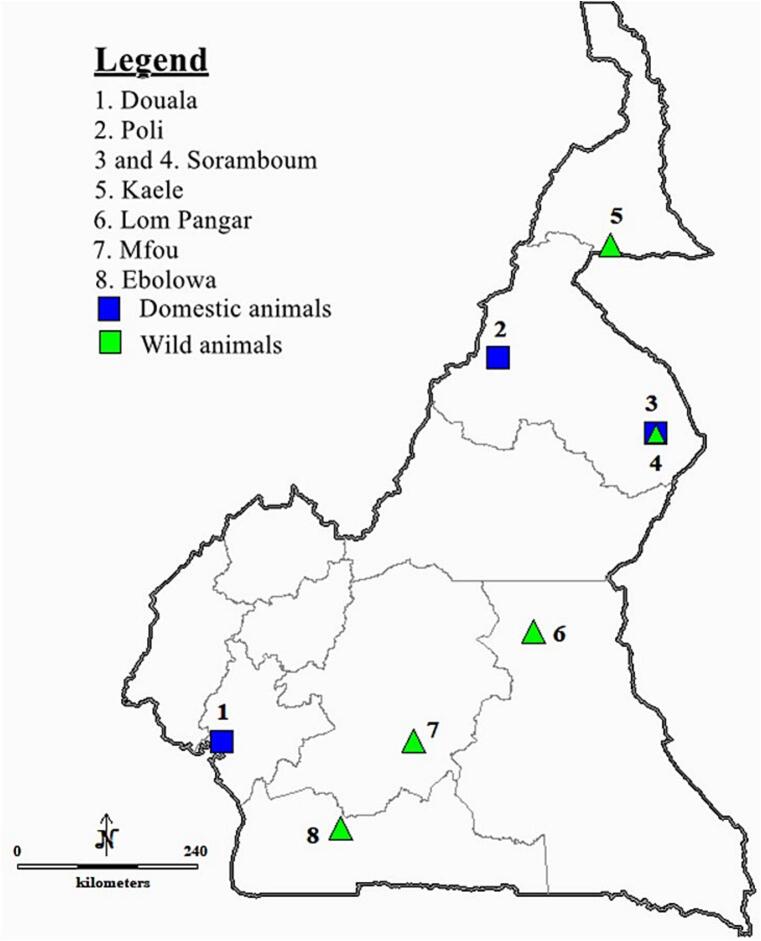 Figure 1
Figure 1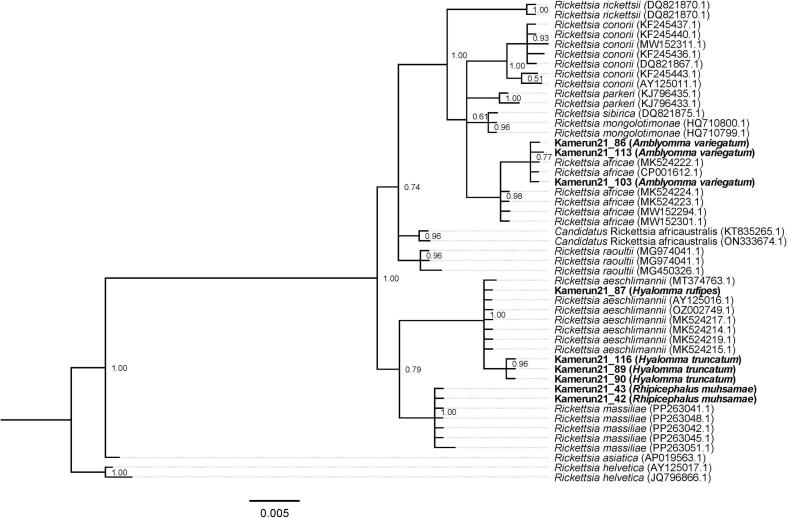 Figure 2
Figure 2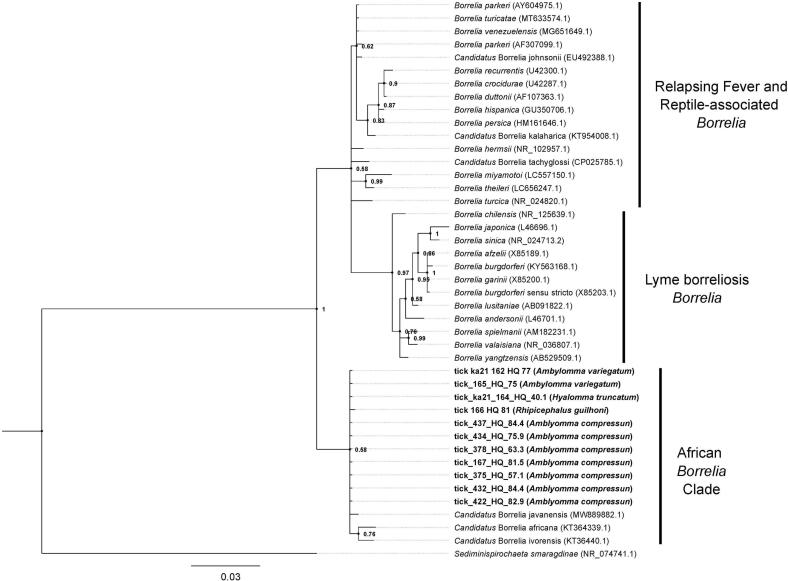 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Bartonella species infections research · Vector-Borne Animal Diseases
Introduction
1
Ixodid ticks are obligate blood-feeding arthropods, which parasitize every class of terrestrial vertebrate in the world (Sonenshine and Roe, 2013). They primarily parasitize wild animals, with only about 10 % of species feeding on domestic animals (Sonenshine and Roe, 2013; Lane and Crosskey, 1993). There are currently 762 recognized species with 17 extant genera in the family Ixodidae which are divided into major two groups, the Prostriata and the Metastriata (Guglielmone et al., 2023; Barker et al., 2024; Kelava et al., 2024). Ticks are also known to harbour, and in some cases vector, a multitude of microorganisms such as viruses, bacteria, protozoa, and helminths (Jongejan and Uilenberg, 2004). Wild animals play a significant role as reservoirs for many pathogens, which can be easily spread to domestic animals or humans via infected tick bites (Liyanaarachchi et al., 2015; Ruiz-Fons and Gilbert, 2010). The resurgence of several tick-borne diseases continues to pose public health concerns and economic impacts worldwide (Abanda et al., 2019; Kuehn, 2019; Wesołowski et al., 2014; Balinandi et al., 2020; Adams, 2016). Although the significance of tick species as disease vectors infesting livestock, pet and humans is well-established, there are still much unknown with respect to tick diversity and host distribution on wildlife (Liyanaarachchi et al., 2015; Schulz et al., 2021) especially in many African countries.
Throughout Africa, different tick species are known to transmit tick-borne pathogens such as piroplasmoses caused by the protozoans belonging to the genera Babesia and Theileria, bacterial infections caused by species belonging to various genera (Anaplasma, Borrelia, Ehrlichia, Rickettsia), and also many viral diseases such as Crimean-Congo haemorrhagic fever (Abanda et al., 2019; Schulz et al., 2021). Members of the genus Rickettsia, such as the species Rickettsia africae, Rickettsia aeschlimannii, and Rickettsia massiliae, are known as the causative agent of African tick bite fever or spotted fever rickettsiosis. The most prevalent species in sub-Saharan Africa is R. africae, where certain tick species of the genus Amblyomma act as the main reservoirs and vectors (Kelly et al., 1996; Paguem et al., 2023; Parola et al., 2001; Vanegas et al., 2018). This infection has been reported with high seroprevalence in sub-Saharan African countries including Cameroon (11.9 %–51.8 %) (Ndip et al., 2004; Mediannikov et al., 2010). Rickettsia aeschlimannii was first identified in a patient returning from Morocco (Raoult et al., 2002). In this country, it was first isolated from Hyalomma marginatum ticks (Parola et al., 2005) but has also been reported by PCR in other Hyalomma species including Hy. rufipes and Hy. truncatum ticks collected from livestock in North Africa (Kernif et al., 2012). In West Africa, R. aeschlimannii was also detected in 15 % to 95 % of Hy. rufipes from Mali, Niger, Senegal, Nigeria, and Cameroon (Mediannikov et al., 2010, Mediannikov et al., 2012, Mediannikov et al., 2013; Parola et al., 2001; Parola, 2006; Vanegas et al., 2018; Diarra et al., 2017; Ngnindji-Youdje et al., 2022). Since its description in 2005, R. massiliae infections in humans have been confirmed in Europe and South America (Parola, 2006; Parola et al., 2008, Parola et al., 2013; Garcia-Garcia et al., 2010; Cascio et al., 2013). Rickettsia massiliae is thought to be associated with Rhipicephalus tick species and was detected by PCR in Rhipicephalus spp. from Côte d'Ivoire (Berrelha et al., 2009), Rhipicephalus guilhoni from Senegal (Mediannikov et al., 2010), Rhipicephalus senegalensis from Guinea (Mediannikov et al., 2012), Rhipicephalus eversti eversti from Nigeria (Reye et al., 2012) as well as Rh. lunulatus and Rh. muhsamae from Cameroon (Ngnindji-Youdje et al., 2022).
In addition to rickettsioses, ixodid ticks can act as the vectors of various Borrelia species which, in some cases, can lead to diseases in humans. They are traditionally classified into the Lyme borreliosis (LB) group, the relapsing fever (RF) group and recently described third group of reptile/monotreme-associated borreliae (Margos et al., 2018). The causative agents of LB are ecologically associated with the ticks of the genus Ixodes and are predominantly found in the temperate northern hemisphere (Barbour, 1998). In contrast, RF group Borrelia are mostly associated with soft ticks and found in subtropical regions worldwide (Cutler et al., 2009; Trape et al., 2013). Relapsing fever is one of the most common diseases in several African regions including Senegal (Vial et al., 2006; Parola et al., 2011) and east African countries (Cutler et al., 2010). It is caused by different Borrelia species such as Borrelia hispanica, Borrelia duttonii, and Borrelia crocidurae. Borrelia hispanica was recently detected in 11.6 % to 20 % of Ornithodoros ticks from northern Africa (Trape et al., 2013; Sarih et al., 2009; Trape et al., 1996). Borrelia crocidurae is responsible for tick-borne relapsing fever in West Africa. In Ethiopia, a Borrelia spp. was identified by PCR in 7.3 % of Amblyomma cohaerens (Mediannikov et al., 2013). Phylogenetically, this Borrelia sp. was placed in an intermediate position between Lyme disease and relapsing fever groups and suggested to potentially belong to the reptile associated Borrelia (Mediannikov et al., 2013). The reptile-associated borreliae were described so far only from Amblyomma, Hyalomma, Bothriocroton, and Ixodes genera, mainly associated with reptiles and echidna (Takano et al., 2010; Takano et al., 2011; Jiang et al., 2021; Loh et al., 2017). The extent of these species' distribution though in other African countries, such as Cameroon, remains unclear.
In Cameroon, so far, 53 ixodid tick species are known from samples collected on domestic and wild animals (Ragenau, 1951; Ragenau, 1953; Morel and Mouchet, 1958; Morel and Mouchet, 1965). Most studies have focused on ticks parasitizing livestock (Bayemi, 1991; Awa et al., 2015; Silatsa et al., 2019; Vanegas et al., 2018; Ngnindji-Youdje et al., 2022) with fewer studies though reporting tick species from wildlife (Paguem et al., 2023; Morel and Mouchet, 1958). To date, however, the existence and/or prevalence of tick-borne associated pathogens (e.g., Rickettsia spp., Borrelia spp.) arising from ticks feeding on wild/domestic animals still remains poorly understood in this country. Our study aims to provide the first data screening for one such potential pathogen (Borrelia) in Wild animals and to provide complementary data on Rickettsia in ticks collected from both domestic and wild animals in Cameroon to provide a better understanding of the epidemiology of these pathogens in Cameroon.
Material and methods
2
Study sites, tick sampling, and species identification
2.1
Between June 2020 and November 2021, ticks were collected from domestic animals (cattle, goats, sheep, donkeys, dogs, and pigs) in three areas (Poli, Soramboum, and Douala) and from wild animals (Phataginus tricuspis (white-bellied pangolin), Varanus niloticus (Nile monitor), Atelerix albiventris (four-toed hedgehog), Cephalophus rufilatus (red flanked duikers), antelopes, Lepus victoriae (African savanna hare), Cercopithecus sp., Caracal aurata (African golden cat), Mastomys natalensis, Civettictis civetta (African civet), Phacochoerus africanus (warthog), Python sebae (African rock python), and Atherurus africanus (African brush-tailed porcupine)) sold in the bush meat markets in five localities (Kaele, Soramboum, Lom Pangar, Mfou, and Ebolowa) (Fig. 1). The information on tick species infesting wild animals and the Rickettsia species detected in the respective ticks has been previously published (Paguem et al., 2023). These samples are considered herein as at that time they were not screened for Borrelia spp. Selected animals were thoroughly examined for attached ticks and all the visible ticks were collected using pointed forceps in a manner as not to damaged them, and kept in vials containing 70 % ethanol until identification. All ticks were identified to the species level using morphological characteristics described by (Hoogstraal, 1956; Matthysse and Colbo, 1987; Apanaskevich et al., 2008; Voltzit and Keirans, 2003; Walker, 2014; Walker et al., 2000; Tomlinson and Apanaskevich, 2019; Arthur, 1965; Bakkes et al., 2020). Some tick species were confirmed by amplifying the 16S rRNA gene as described by Halos et al. (2004). Tick sequence data generated from wild animals was previously published in Paguem et al. (2023).Fig. 1. Map of the study area. The sampling areas (blue square and green triangle) were located in different areas of Cameroon. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)Fig. 1
RNA/DNA extraction
2.2
Total RNA/DNA was extracted using the MagNA Pure LC RNA/DNA Kit (Roche, Mannheim, Germany) in a MagNA Pure LC instrument according to the manufacturer's instructions. DNA was extracted from individual ticks (if a single tick was found on a single host or a single life stage of a respective tick species) or pools (2–10 ticks per pool, if ticks belonged to the same species, developmental stage and were collected from the same animal). From the 1148 ticks collected this corresponded to 415 DNA pools which were screened for microorganisms in the PCR analyses described below. The extracted total DNA was stored at -80 °C until PCR analysis.
PCR amplification of Rickettsia spp and Borrelia spp
2.3
Rickettsia spp. DNA was analysed using a pan-Rickettsia real-time RT-PCR to amplify part of the gltA gene (Wölfel et al., 2008), ticks tested positive for rickettsiae were identified down to Rickettsia species level by PCR amplification of 23S—5S intergenic spacer region (Chitimia-Dobler et al., 2018).For this purpose, primers 23S and 23S rev and the thermoprofile of a previously published method (Jado et al., 2006) were modified to achieve optimum sensitivity. Briefly, 5 μl DNA, 0.5 μM Primer 23S for and 23S rev (Table 1), 1 U Platinum® Taq DNA Polymerase High Fidelity (Invitrogen), 1× reaction buffer, and a final concentration of 4 mM MgSO4 were added to a final volume of 50 μl per reaction. Initial denaturation at 95 °C for 2 min was followed by 45 cycles at 95 °C for 30 s, 30 s at 58 °C, and 30 s at 68 °C and a final extension at 68 °C for 10 min. The obtained DNA amplicons were visualized by gel electrophoresis.Table 1. Primers and probes used for molecular investigation of tick species and their pathogens.Table 1. GenusPrimerTarget genePrimer sequence (5′-3′)Annealing T (°C)Amplicon size (bp)ReferencesTicksTQ16S + F116S rRNACTGCTCAATGATTTTTTAAATTGCTGTGG320Halos et al. (2004)TQ-16S-2RACGCTGTTATCCCTAGAGRickettsiaRH314:gltAAAACAGGTTGCTCATCATTCWölfel et al. (2008)RH654:AGAGCATTTTTTATTATTGGNot applicableRickettsia23S for23S–5S intergenic spacer regionGATAGGTCGGGTGTGGAAGCACChitimia-Dobler et al. (2018)23S revGGGATGGGATCGTGTGTTTCAC378–532Borrelia outer16S1A16S rRNACTA ACG CTG GCA GTG CGT CTT AAG631205Richter and Matuschka (2006); Abanda et al. (2019)16S1BAGC GTC AGT CTT GAC CCA GAA GTTBorrelia inner16S2A16S rRNAAGT CAA ACG GGA TGT AGC AATAC56600–720Richter and Matuschka (2006); Abanda et al. (2019)16S2BGTT ATT CTT TCT GAT ATC AACAG
For all PCR, standard procedures for PCR testing (three room concept, inclusion of positive and negative controls, extraction controls) were included in each run (Chitimia-Dobler et al., 2018).
Additionally, to detect Borrelia DNA, generic primers were used in a nested PCR targeting the 16S rRNA as described previously (Richter and Matuschka, 2006; Abanda et al., 2019). Briefly, the first reaction (25 μL final volume) contained 2 μM of each outer primer (Table 1), 0.2 mM dNTP mix, 0.5 U Go Taq DNA polymerase (Promega, Germany), 1× Go Taq buffer, and 1 μL of extracted DNA. Nuclease-free water was used as negative control and plasmid containing 10 copies of 16 s rDNA of Borrelia burgdorferi s.l kindly shared by colleagues from Freie Universität Berlin, Germany (Abanda et al., 2019) was used as positive control. The corresponding gene loci, primer pairs and annealing temperatures are shown in Table 1. PCR amplification was carried out as follows: initial denaturation step at 95 °C for 3 min, followed by 35 amplification cycles at 95 °C for 60 s, at 63 °C for 60 s, at 72 °C for 30 s, and final extension at 72 °C for 10 min (Master Cycler EP S Thermal Cycler®, Eppendorf, Hamburg, Germany). Thereafter, the second PCR reaction was carried out with 1 μL of first PCR product as template under the same cycling conditions as described above, except for an annealing temperature of 56 °C, and using the inner primer pairs (Table 1). All samples were visualized through electrophoresis on a 1.5 % agarose gel stained with Midori Green (Nippon Genetics Europe, Düren, Germany).
sequencing
2.4
The Rickettsia 23S—5S amplicon obtained were sequenced by Sanger sequencing (GATC Biotech, Konstanz, Germany) and Selected positive reactions were prepared following manufacturer's recommendations (Macrogen, Amsterdam, Netherlands) and sent for sequencing.
BLAST and phylogenetic analysis
2.5
All the sequences obtained were screened with BLASTn analysis (Altschul et al., 1990) and representative related sequences downloaded from GenBank (https://www.ncbi.nlm.nih.gov/ nucleotide).
Sequence data for Rickettsia 23S—5S intergenic spacer obtained from ticks collected in this study (n = 9) were compiled with reference data from GenBank and aligned using MUSCLE v3.8.425 (Edgar, 2004a, Edgar, 2004b) as implemented in Aliview v1.28 (Larsson, 2014). Phylogenetic reconstruction was performed in MrBayes v. 3.2.6 (Huelsenbeck and Ronquist, 2001; Ronquist et al., 2012) with ploidy set to haploid and a GTR (Tavaré, 1986) substitution model with inverse gamma-distributed rate variation. Three independent runs were launched and ran for two million generations. Convergence was checked with Tracer v. 1.7.1 (Rambaut et al., 2018). Consensus trees were built using the sumt command from MrBayes using a respective burn-in of 25 %. Convergence to a single topology in all three independent runs was checked manually in FigTree v. 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/). The Rickettsia tree was rooted on the branch leading to the clade containing both Rickettsia helvetica reference sequences.
Borrelia species identity was determined through phylogenetic reconstruction based on the partial 16S rRNA sequences produced during this study. Sequences were compiled with GenBank references representing members of the known Borrelia clades (Lyme borreliosis, relapsing fever, reptile/monotreme-associated) and recently described Borrelia found in ticks from Africa. Sequences were then aligned using MUSCLE v3.8.425 (Edgar, 2004a, Edgar, 2004b) as implemented in Aliview v1.28 (Larsson, 2014). Phylogenetic reconstruction was performed in MrBayes v. 3.2.6 (Huelsenbeck and Ronquist, 2001; Ronquist et al., 2012) with ploidy set to haploid and a GTR (Tavaré, 1986) substitution model with inverse gamma-distributed rate variation. Three independent runs were launched and ran for five million generations. Convergence was checked with Tracer v. 1.7.1 (Rambaut et al., 2018). Consensus trees were built using the sumt command from MrBayes using a respective burn-in of 25 %. Convergence to a single topology in all three independent runs was checked manually in FigTree v. 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/). The 16S rRNA sequence from Sediminispirochaeta smaragdinae was included as an outgroup to root the tree.
Nucleotide sequence data reported in this paper are available in the National Center for Biotechnology Information (NCBI) GenBank™ databases under the accession numbers: PQ844815- PQ844825 for Borrelia and PV110811-PV110819 for Rickettsia sequences.
Results
3
Tick species and host range
3.1
In total, 1148 ticks (557 males, 471 females, 106 nymphs and 14 larvae) were collected from 117 domestics animals including cattle (n = 49), goats (n = 35), sheep (n = 7), donkeys (n = 1), dogs (n = 11), and pigs (n = 14) and from 166 out of 2000 wild animals examined. Infested wild animals belonged to thirteen species: Phataginus tricuspis (white-bellied pangolin, n = 48), Varanus niloticus (Nile monitor, n = 31), Atelerix albiventris (four-toed hedgehog, n = 25), Cephalophus rufilatus (red flanked duikers, n = 15), antelopes (n = 15), Lepus victoriae (African savanna hare, n = 9), Cercopithecus sp. (monkey, n = 7), Caracal aurata (African golden cat, n = 5), Mastomys natalensis (rodent, n = 4), Civettictis civetta (African civet, n = 3), Phacochoerus africanus (warthog, n = 2), Python sebae (African rock python, n = 1), and Atherurus africanus (African brush-tailed porcupine, n = 1) (all information about ticks collected from the 166 wild animals have already been published, see Paguem et al., 2023).
Based on morphological characteristics and 16S rDNA sequencing, 25 different tick species belonging to Amblyomma, Haemaphysalis, Hyalomma, Ixodes, and Rhipicephalus genera were identified. Overall, Amblyomma compressum (286/1148, 24.91 %) was the most common tick species collected and, although it was only found on wild animals, it represented almost half of the ticks exclusively collected from wild animals (286/686, 41.70 %, highly common on pangolins). The other two major tick species collected where Rhipicephalus microplus (197/1148, 17.16 %) and Amblyomma variegatum (192/1148, 16.72 %). The host ranges of the 27 species of ticks belonging to the family Ixodidae collected from domestic animals are shown in the Table 2 and information on host ranges for wild animals can be found in Paguem et al. (2023). Of the 25 tick species infesting 24 species of domestic and wild animal, then species, namely Rh. guilhoni, Rhipicephalus annulatus, Rh. microplus, Rh. decoloratus, Rh. linnaei, Hy. truncatum, Hy. rufipes, Hy. nitidum, Rh. afranicus and A. variegatum were common on domestic and wild animals. Eight tick species were only found in wild animals.Table 2. Prevalence of Rickettsia spp. in tick collected from domestic animals. Rickettsia spp. detected in the ticks collected from wild animals are reported in Paguem et al. (2023).Table 2. Tick speciesTick life stagesHostRickettsia spp.MalesFemalesNymphsPan*-Rickessia* positive/tick pool23S–5S intergenic spacer regionAmblyomma variegatum52143Cattle, goat,unidentified wild animals14/20R. africae (n = 3)Haemaphysalis leachi1Dog0/1Hyalomma nitidum1Dog0/1Hyalomma rufipes74Cattle, sheep1/6R. aeschlimannii (n = 1)Hyalomma truncatum3220Pig, dog, cattle, unidentified wild animals, goats5/13R. aeschlimannii (n = 3)Rhipicephalus afranicus45Goats0/6Rhipicephalus annulatus64Sheep, goat, cattle, dog, unidentified wild animals0/17Rhipicephalus cuspidatus1unidentified wild animals0/1Rhipicephalus decoloratus22Goat, cattle, dog, unidentified wild animals0/8Rhipicephalus guilhoni83Dog, goat, unidentified wild animals0/7Rhipicephalus lunulatus12Goats, unidentified wild animals0/3Rhipicephalus microplus25140Sheep, goat, donkey, cattle, dog, unidentified wild animals0/30Rhipicephalus muhsamae33121Pig, dog, goat, cattle, unidentified wild animals2/18R. massiliae (n = 2)Rhipicephalus senegalensis33Goats0/3Rhipicephalus sp.1Cattle0/1Total165292522/135
Frequency of positive pools of Rickettsia and Borrelia spp. in ticks collected from domestic and wild animals
3.2
Briefly, sequencing results were obtained for 66 out of the 114 Rickettsia-positive ticks pools collected from wild animals as reported in Paguem et al. (2023). Three different Rickettsia species were identified in these samples with the most common species being Rickettsia africae 61/66, 92.4 %), with very few samples identified as Rickettsia aeschlimannii (3/66, 4.5 %) and Candidatus Rickettsia africaustralis (2/66, 3.0 %). Rickettsia sequences were obtained for nine of the 22 Rickettsia-positive ticks collected from domestic animals in this study (Table 2). These were identified as R. aeschlimannii (4/9, 44.4 %) in Hy. rufipes (n = 3) and Hy. truncatum (n = 1), Rickettsia africae (3/9, 33.3 %) in A. variegatum (n = 3), and R. massiliae was detected in Rh. muhsamae males (Table 2; Fig. 2). Rickettsia africae sequences were 99 % identical with the sequence MK524222 and MK524224 isolated from A. lepidum in Sudan*. Rickettsia aeschlimannii (PV110813, PV110814 and PV110817) was 99.68 % identical with the sequences MT374764 and MK524216 from Hy. rufipes in Italy and Sudan respectively. R. massiliae* (PV110818.1) was 100 % identical with PP263045, PP263050 and PP263048 from ticks in Greece*.*Fig. 2. Phylogenetic reconstruction based on the 23S—5S intergenic spacer sequences for Rickettsia spp. amplified from ticks collected from wild/domestic animals in Cameroon. Samples sequenced in this study are shown in bold and GenBank accession numbers are included for all reference sequences. Phylogenetic reconstruction was performed in MrBayes v. 3.2.6 (Huelsenbeck and Ronquist, 2001; Ronquist et al., 2012) with ploidy set to haploid and a GTR (Tavaré, 1986) substitution model with inverse gamma-distributed rate variation. Three independent runs were launched and ran for 5 million generations. Convergence was checked with Tracer v. 1.7.1 (Rambaut et al., 2018). The phylogeny was rooted on the branch leading to the monophyletic clade containing both reference sequences for R. helvetica.Fig. 2
Screening of all ticks for Borrelia spp. using nPCR, detected 49/415 (10.38 %) positive tick pools (Table 3). Ten out of 27 tick species tested positive to Borrelia spp.: namely A. compressum 30/111 (27.03 %), A. flavomaculatum 6/17 (35.29 %), A. variegatum 1/49 (2.04 %), Hy. rufipes 1/8 (12.5 %), Hy. truncatum 2/18 (11.11 %), Rh. annulatus 1/17 (5.88 %), Rh. afranicus 1/5 (20.00 %), Rh. guilhoni 2/2 (100 %), Rh. lunulatus 2/3 (66.67 %), Rh. senegalensis 3/3 (100 %). We succeeded in amplifying a fragment of the 16S rRNA in 11 ticks pools. Phylogenetic reconstruction based on the 16S rRNA fragment supported two major clades one containing all relapsing fever, reptile/monotreme-associated, and Lyme borreliosis species with a second clade containing newly described African Borrelia and all ticks sequenced in this study (Fig. 3). The split between these groups is highly supported with a node probability of 1 (Fig. 3). All sequenced Borrelia reported in this study show highest similarity to Candidatus Borrelia javanensis (GenBank: MW889882.1) isolated from pangolins in China.Table 3. Prevalence of Borrelia spp. detected in ticks.Table 3. Tick speciesTotal tick poolsBorrelia spp. percentage of tick pools (%)HostAmblyomma compressum11130/111 (27.03 %)White-bellied pangolins, Warthogs, Nile monitors, African brush-tailed, Porcupines, Monkeys, African golden cat, Antelopes, African civetsAmblyomma flavomaculatum186/18 (33.33 %)Nile monitors, Four toed hedgehogs, White bellied pangolins, Monkeys, WarthogsAmblyomma variegatum471/47 (2.13 %)Nile monitor, Four toed hedgehog, Pigs, Dogs, Cattle, Goats, Un-identified wild animalsHaemaphysalis leachi9White-bellied pangolins, African civetsHaemaphysalis camicasi1Red flanked duikerHaemaphysalis houyi65Nile monitors, Red flanked duikers, Monkeys, African savanna hares, Four toed hedgehogs,Antelopes, African rock pythonHaemaphysalis parmata2Nile monitors, AntelopeHyalomma nitidum2Red flanked duiker, DogsHyalomma rufipes81/8 (12.5 %)Four-toed hedgehogs, Sheep, Cattle,Hyalomma truncatum182/18 (11.11 %)Four-toed hedgehogs, Nile monitors, Pigs, Dogs, Cattle, Un-identified wild animalsIxodes moreli1AntelopeIxodes rasus10White-bellied pangolins, Four-toed hedgehogs, Warthogs, MonkeysRhipicephalus microplus41Four-toed hedgehogs, Nile monitors,Rhipicephalus decoloratus8Cattle, Goat, Un-identified wild animalsRhipicephalus annulatus171/17 (5.88 %)Cattle, Sheep, Goat, Un-identified wild animalsRhipicephalus afranicus191/19 (5.3 %)Goat, Monkeys, African rock pythons, Red flanked duikers, Antelopes, Nile monitors, Four-toed hedgehogsRhipicephalus guilhoni92/2 (100 %)African savanna hares, Four-toed hedgehogs,Nile monitorsRhipicephalus lunulatus32/3 (66.67 %)Goat and Un identified wild animalsRhipicephalus moucheti11Red flanked duikers, Antelopes, Monkeys, Four-toed hedgehogs, African savanna hares, Nile monitorsRhipicephalus muhsamae10Rats, Nile monitorsRhipicephalus linnaei2Red flanked duikers, African savanna hares, Nile monitorsRhipicephalus senegalensis33/3 (100 %)GoatRhipicephalus sp.1CattleTotal41549/415 (11.81 %)Fig. 3. Phylogenetic reconstruction based on partial nucleotide sequences of the 16S rRNA (573 bp) genes of Borrelia spp. identified in ticks collected from domestic and wild animals in Cameroon. Samples sequenced in this study are shown in bold and GenBank accession numbers are included for all reference sequences. Phylogenetic reconstruction was performed in MrBayes v. 3.2.6 (Huelsenbeck and Ronquist, 2001; Ronquist et al., 2012) with ploidy set to haploid and a GTR (Tavaré, 1986) substitution model with inverse gamma-distributed rate variation. Three independent runs were launched and ran for 5 million generations. Convergence was checked with Tracer v. 1.7.1 (Rambaut et al., 2018). The 16S rRNA sequence from Sediminispirochaeta smaragdinae was included as an outgroup to root the tree. Numbers next to the nodes reports node probability and the scale bar is in substitutions per base pair.Fig. 3
Two co-infections of Rickettsia and Borrelia (2/415; 0.42 %) were detected by nPCR. Co-infections involved the presence of an unidentified as Borrelia spp. closely related to Candidatus Borrelia javanensis with R. aeschlimannii in a single Hy. rufipes collected from a four-toed hedgehog and with R. africae in a single A. flavomaculatum collected from a Nile monitor.
Discussion
4
Borrelia and Rickettsia species in ticks collected from domestic and wild animals in Cameroon were investigated. The present study provides the first molecular proof for the presence of Borrelia spp. closely related to Candidatus Borrelia javanensis in ticks from wild animals in Cameroon.
Three Amblyomma species were recorded from domestic and wild animals, with A. compressum and A. flavomaculatum exclusively found on wild animals, whereas A. variegatum were found on both domestic and wild animals. Further description of tick diversity and host range in wild animals can be found in Paguem et al. (2023). The additional results from domestic animals show, that nine species, namely Rh. guilhoni, Rh. annulatus, Rh. microplus, Rh. decoloratus, Rh. linnaei, Hy. truncatum, Hy. rufipes, Hy. nitidum, and A. variegatum infested both domestic and wild animals. In this study, and as previously shown on cattle and small ruminants, three tick species were predominant: Rh. decoloratus, Rh. microplus, and A. variegatum (Awa et al., 2015; Silatsa et al., 2019; Ngnindji-Youdje et al., 2022). This finding could suggest that wild animals that cohabit with domestic animals could additionally act as hosts of tick classically found on livestock (Pegram et al., 1987).
There are many reports about A. variegatum carrying R. africae from different African countries such as Sudan (Springer et al., 2020; Shuaib et al., 2020), Ethiopia (Tufa et al., 2021), and Cameroon (Ndip et al., 2004; Vanegas et al., 2018; Ngnindji-Youdje et al., 2022; Paguem et al., 2023). In the present study, R. africae was detected in A. variegatum collected from domestic animals and in A. compressum and A. flavomaculatum collected from wild animals (see Paguem et al., 2023). The high proportion of infested ticks from our data and previous reports could suggest that tick species from the genus Amblyomma play an important role as vector and reservoir of R. africae in the Afrotropical region*. Rickettsia aeschlimannii* was identified in two Hy. rufipes and one Hy. truncatum in the screened tick samples. Samples from this study however formed two distinct clades within the phylogeny. Even so, upon sequence comparison the sequences were over 99 % similar suggesting this structure could be due to local sequence polymorphisms instead of representing a novel Rickettsia lineage or species. Rickettsia aeschlimannii is a recognized human pathogen causing spotted fever and has been detected in different countries in sub-Saharan Africa in A. variegatum, Rh. annulatus, Rh. evertsi evertsi, Rh. appendiculatus, H. rufipes, and H. truncatum (Parola et al., 2013). Rickettsia massiliae was detected in two Rh. muhsamae males (one male tested individual and in one 10 male pool) collected from pigs in Sorrambum in the Mayo-Rey region. It was previously reported in ticks collected from cattle in the western region of this area. Rickettsia massiliae was detected in (16/27) Rh. lunulatus, Rh. muhsamae (1/10) (Ngnindji-Youdje et al., 2022) and in one Rh. lunulatus from Cameroon (Vanegas et al., 2018). Rickettsia massiliae was reported from Côte d'Ivoire in Rh. senegalensis (33 %) and in Rh. guilhoni (22 %) from Senegal (Sarih et al., 2009; Trape et al., 1996; Georges, 2005). Further investigations are necessary regarding the species identity and phylogeny of these unidentified Rickettsia species.
The association of Borrelia spirochetes with ticks in Africa in general and in Cameroon in particular is still poorly understood. Until now only Borrelia theileri, a member of the tick-borne relapsing fever group was reported in one Rh. microplus tick collected in western region of Cameroon (Ngnindji-Youdje et al., 2022) and detected in cattle blood in the country (Abanda et al., 2019). In this study, Borrelia sp. was detected in 10.38 % of tick pools in three Amblyomma species (A. compressum, A. flavomaculatum, A. variegatum), two Hyalomma species (Hy. rufipes, Hy. truncatum), and five Rhipicephalus species (Rh. annulatus, Rh. afranicus, Rh. guilhoni, Rh. lunulatus, and Rh. senegalensis) collected from domestic and wild animals across different geographical areas. The BLAST and phylogenetic analysis of the successfully sequenced samples (n = 11), showed that these Borrelia sp. shared similarity and clustered with Candidatus B. javanensis detected in 3 % (12/227) of Amblyamma javanense ticks collected from pangolins (Manis javanica) in China (Jiang et al., 2021). Interestingly, most of the Borrelia spp. closely related to Candidatus B. javanensis positive samples were from A. compressum ticks collected feeding on pangolins. This could suggest that a potential association between Borrelia spp. closely related to Candidatus B. javanensis and pangolins could exist, although this would need to be stringently tested in lab-based studies*.* The clade within the phylogeny also contained other recently described Borrelia, namely, Candidatus B. africana and Candidatus B. ivorensis from A. variegatum in Côte d'Ivoire (Ehounoud et al., 2016). We detected Borrelia spp. closely related to Candidatus B. javanensis in A. compressum, A. flavomaculatum, in one A. variegatum, two Hy. truncatum and one Rh. guilhoni from Cameroon*.* Our study represents, to the best of our knowledge, the first report of Borrelia spp. closely related to Candidatus B. javanensis in A. compressum and A. flavomaculatum. Blast analysis of the 16S rRNA gene did show variation between the reference sequence for Candidatus B. javanensis from screened ticks collected in China although all sequences reported in this study clustered together with this reference sequence in the phylogenetic reconstruction. Our phylogenetic reconstruction differs slightly from previous reports of Candidatus B. javanensis suggesting that this and other African Borrelia may be separate from the known Borrelia clades (Jiang et al., 2021) although analysis including additional genetic markers is needed to support this. Further molecular investigation is needed, however, to better resolve the phylogeny by including analysis of additional marker genes (e.g., flaB, gyrB) genes or utilizing whole genome sequencing. Our results suggest the possibility of more complex vector pathogen-reservoir host interactions which need to be investigated in more detail. Additionally, the pathogenicity and zoonotic potential of the Borrelia reported here is not known and will need to be addressed in future studies.
Conclusions
5
The present study was carried out to determine which tick species infest domestic and wild animals in Cameroon and also to determine which are positive for Borrelia and Rickettsia. As far as it is known, this is the first kind of tick survey both on domestic and wild animals and their associated Borrelia and Rickettsia pathogens performed in Cameroon. In total, twenty-five tick species belonging to Amblyomma, Haemaphysalis, Hyalomma, Ixodes, and Rhipicephalus genera were collected from six domestic and thirteen wild animal species. Three species of Rickettsia were detected in ticks collected from domestic animals namely R. africae, R. aeschlimannii, and R. massiliae. Additionally, Borrelia spp. are reported for the first time. Borrelia species detected in ticks from wild and domestic animals in Cameroon are closely related to Candidatus B. javanensis from China, and from a clade with Candidatus B. africana and Candidatus B. ivorensis from Ivory Coast. Although this study is limited to the analysis of partial 16S fragment sequences Borrelia spp., robust multilocus (e.g., flaB, glpQ, gyrB) or whole genomic sequencing is needed for confirmation. Further studies should be encouraged to investigate the risk of tick-borne diseases, especially in relation to potential bovine borrelioses, in order to reduce potential risks in the region.
CRediT authorship contribution statement
Archile Paguem: Writing – review & editing, Writing – original draft, Visualization, Validation, Investigation, Formal analysis, Data curation, Conceptualization. Pierre Kamtsap: Writing – review & editing, Writing – original draft, Visualization, Validation, Formal analysis. Kingsley Tanyi Manchang: Writing – review & editing, Writing – original draft, Methodology, Formal analysis. Alfons Renz: Writing – review & editing, Writing – original draft, Validation, Supervision, Conceptualization. Sabine Schaper: Writing – review & editing, Writing – original draft, Methodology, Formal analysis. Gerhard. Dobler: Writing – review & editing, Writing – original draft, Resources, Methodology, Conceptualization. Robert E. Rollins: Writing – review & editing, Writing – original draft, Software, Methodology, Formal analysis, Data curation. Lidia Chitimia-Dobler: Writing – review & editing, Writing – original draft, Visualization, Validation, Supervision, Resources, Methodology, Formal analysis, Conceptualization.
Ethical statement
The study was approved by the National Institute of Agricultural Research for Development (IRAD) in Cameroon, which is the country's government institution for animal health and husbandry improvement. The ethical clearance number is CEIUD/371/01/2016/M.
Funding statement
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Declaration of competing interest
None.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abanda B.Paguem A.Abdoulmoumini M.Kingsley M.T.Renz A.Eisenbarth A.Molecular identification and prevalence of tick-borne pathogens in zebu and taurine cattle in North Cameroon Parasit. Vectors 12201944810.1186/s 13071-019-3699-x 31511038 PMC 6737592 · doi ↗ · pubmed ↗
- 2Adams D.A.Summary of notifiable infectious diseases and conditions—United States, 2014 MMWR Morb. Mortal. Wkly Rep.63542016115210.15585/mmwr.mm 6354 a 127736829 · doi ↗ · pubmed ↗
- 3Altschul S.F.Gish W.Miller W.Myers E.W.Lipman D.J.Basic local alignment search tool J. Mol. Biol.2151990403410223171210.1016/S 0022-2836(05)80360-2 · doi ↗ · pubmed ↗
- 4Apanaskevich D.A.Schuster A.L.Horak I.G.The genus Hyalomma: VII. Redescription of all parasitic stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae)J. Med. Entomol.45200881783110.1603/0022-2585(2008)45[817:tghvro]2.0.co;218826023 · doi ↗ · pubmed ↗
- 5Arthur D.R.Ticks of the Genus Ixodes in Africa 1965 The Athlone Press London, UK
- 6Awa D.N.Adakal H.Luogbou N.D.D.Wachong K.H.Leinyuy I.Achukwi M.D.Cattle ticks in Cameroon: is Rhipicephalus (Boophilus) microplus absent in Cameroon and the Central African region?Ticks Tick Borne Dis.6201511712210.1016/j.ttbdis.2014.10.00525575435 · doi ↗ · pubmed ↗
- 7Bakkes D.K.Chitimia-Dobler L.Matloa D.Oosthuysen M.Mumcuoglu K.Y.Mans B.J.Matthee C.A.Integrative taxonomy and species delimitation of Rhipicephalus turanicus (Acari: Ixodida:Ixodidae)Int. J. Parasitol.50202057759410.1016/j.ijpara.2020.04.00532592812 · doi ↗ · pubmed ↗
- 8Balinandi S.Chitimia-Dobler L.Grandi G.Nakayiki T.Kabasa W.Bbira J.Lutwama J.J.Bakkes D.K.Malmberg M.Mugisha L.Morphological and molecular identification of Ixodid tick species (Acari: Ixodidae) infesting cattle in Uganda Parasitol. Res.11920202411242010.1007/s 00436-020-06742-z 32533261 PMC 7366568 · doi ↗ · pubmed ↗