Exploring the role and therapeutic potential of RNA N6-methyladenosine modification in abortion disease pathology: a comprehensive review
Zhuo Chang, Lu-Hao Li, Liang-Zhen Lv, Zhao-Di Wang, Qing-yi Wang, Hui Zhu, Bei Jiang, Xue-Ming Zhou, Ya-Peng Han, Xue Pan, Li Ren, Sen Cheng, Zi-Meng Lei

TL;DR
This review explores how RNA modifications, specifically m6A, may contribute to recurrent miscarriages and could lead to new treatments.
Contribution
The paper provides a comprehensive review of m6A's role in RSA, highlighting its potential as a therapeutic target.
Findings
m6A modification influences gametogenesis, embryo quality, and placental development.
Dysregulation of m6A regulators is linked to RSA pathophysiology in human and animal studies.
Targeting m6A machinery presents therapeutic potential but requires further research.
Abstract
Recurrent spontaneous abortion (RSA), defined as two or more consecutive pregnancy losses, affects 1%–5% of couples and poses a significant challenge to reproductive health. Despite its prevalence, the underlying etiology remains elusive in approximately half of all cases, hindering the development of targeted therapies. The emerging field of epitranscriptomics, particularly the dynamic and reversible N6-methyladenosine (m6A) RNA modification, offers a novel lens through which to investigate the complex gene-environment interactions underlying RSA. This review systematically synthesizes current knowledge on the pivotal roles of m6A modification in key processes essential for a successful pregnancy: gametogenesis and early embryo quality, placental development and function, and the establishment of immune tolerance at the maternal-fetal interface. We critically evaluate the direct and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
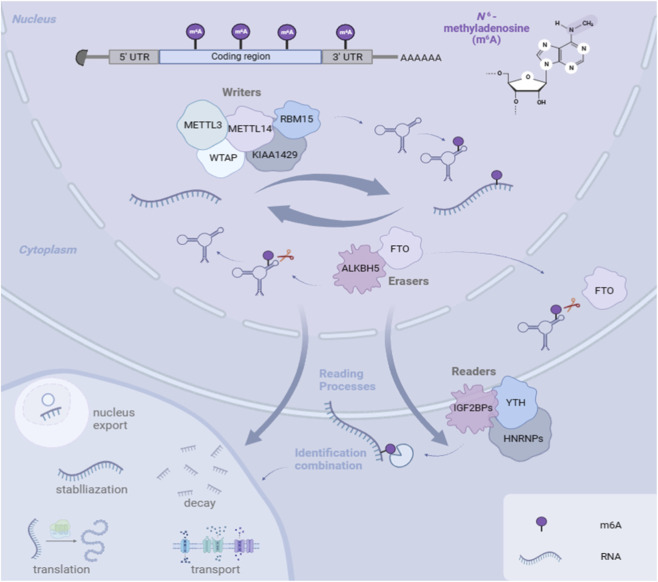 FIGURE 1
FIGURE 1- —National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA modifications and cancer · Metalloenzymes and iron-sulfur proteins · Chromatin Remodeling and Cancer
Background
1
Recurrent spontaneous abortion (RSA) is a devastating disorder, classically defined as three or more consecutive losses before 28 weeks (Practice Committee of the American Society for Reproductive, 2012), with modern guidelines recognizing two or more losses as clinically significant. It affects 1%–5% of couples and inflicts profound physical and psychological burdens (Rai and Regan, 2006), (Turesheva et al., 2023). Alarmingly, the pathogenesis remains unclear in over 60% of cases (Jaslow et al., 2010),creating a major barrier to effective treatment. This underscores the urgent need to explore novel molecular pathways governing pregnancy establishment and maintenance.
Successful pregnancy requires flawless execution of interconnected events: competent embryo development, implantation, placentation via trophoblast invasion and decidualization, and establishment of local immune tolerance. Epigenetic regulation, particularly RNA modification, has emerged as a key mechanism controlling these processes (Zhao et al., 2020; Sendinc and Shi, 2023). Among these,N6-methyladenosine (m6A)—the most abundant internal mRNA modification in eukaryotes—stands out for its dynamic and reversible nature, enabling precise post-transcriptional control of gene expression (Roundtree et al., 2017; Huang N. et al., 2022; Dominissini et al., 2012; Gilbert et al., 2016).
The m6A modification is installed by methyltransferase complexes (“writers” like METTL3/METTL14/WTAP), removed by demethylases (“erasers” like FTO/ALKBH5), and interpreted by binding proteins (“readers” like the YTHDF family) that direct RNA fate, influencing splicing, stability, and translation (Shi et al., 2019; Jia et al., 2011; Zhang et al., 2010; Wang and Lu, 2021; Shi et al., 2017).While m6A’s role in fields like oncology is well-established (Willyard, 2017; Lee et al., 2014), its importance in reproduction is increasingly recognized (Huang and Chen, 2023).It is integral to key reproductive events, from gametogenesis to placental development and immune regulation (Zhao et al., 2022).
However, research directly investigating m6A in RSA-specific pathogenesis remains nascent and fragmented. Insights are often extrapolated from other pregnancy complications or non-reproductive models. Therefore, this review aims to provide a comprehensive and critical analysis of m6A’s role in RSA. We will synthesize physiological functions, evaluate direct and indirect pathogenic evidence, identify conflicts and gaps, and discuss therapeutic potential and challenges.
The mechanisms and regulation of m6A RNA methylation
1.1
Epigenetics plays a pivotal role in regulating gene expression, which is crucial for both disease development and prevention. Among the known epigenetic mechanisms—including DNA modification, histone modification, RNA modification, and chromatin remodeling—RNA modification has become a major research focus (Zhao et al., 2020). RNA molecules undergo approximately 160 types of chemical modifications, with methylation being the most prevalent (Sendinc and Shi, 2023).
In eukaryotes, N6-methyladenosine (m6A) is the most abundant internal modification on messenger RNA (mRNA) and long non-coding RNAs, occurring at the sixth nitrogen position of adenosine (Roundtree et al., 2017; Huang N. et al., 2022; Dominissini et al., 2012). This modification is enriched near stop codons and within 3′untranslated regions. m6A dynamically influences multiple aspects of RNA metabolism, including splicing, translational efficiency, nuclear export, stability, and decay. Importantly, m6A modification is reversible, which underpins its significant regulatory potential and therapeutic prospects (Gilbert et al., 2016; Berulava et al., 2020).
The m6A modification process is regulated by three classes of proteins: methyltransferases (“writers”), demethylases (“erasers”), and binding proteins (“readers”) (Figure 1).
The mechanism of m6A readers, writers, and erasers.
Methyltransferases
1.1.1
Methyltransferases, or writers, catalyze the addition of a methyl group to adenosine on RNA. The core N6-adenosine methyltransferase complex (MTC) in mammals includes methyltransferase-like protein 3 (METTL3), methyltransferase-like protein 14 (METTL14), and Wilms’ tumor 1-associating protein (WTAP) (Roundtree et al., 2017). METTL3 is the catalytic core, directly binding to the methyl donor S-adenosylmethionine (Bokar et al., 1997; Xiong et al., 2022). METTL14, while lacking independent catalytic activity, forms a stable heterodimer with METTL3, enhancing RNA binding and complex stability (Wang et al., 2016; Lin et al., 2017). WTAP does not possess catalytic activity but is essential for localizing the METTL3-METTL14 complex to nuclear speckles, thereby guiding site-specific methylation (Huang Q. et al., 2022; Ping et al., 2014). Recent studies have identified additional regulatory components, such as RNA binding motif protein 15 (RBM15), Vir-like m6A methyltransferase associated (VIRMA/KIAA1429), and zinc finger CCCH domain-containing protein 13 (ZC3H13), which contribute to the specificity and efficiency of m6A deposition (Jia et al., 2021; Wang Y. et al., 2021; Wang et al., 2020).
Demethylation enzymes
1.1.2
Demethylases, or erasers, remove m6A marks, making this modification reversible. The two primary m6A demethylases are fat mass and obesity-associated protein (FTO) and AlkB homolog 5 (ALKBH5), both belonging to the α-ketoglutarate-dependent dioxygenase family (Jia et al., 2022). The discovery of FTO’s demethylase activity in 2011 overturned the long-held view of static RNA modification, confirming the dynamic nature of m6A (Fu et al., 2013). FTO is distributed in both the nucleus and cytoplasm, whereas ALKBH5 is primarily nuclear. Both enzymes remove m6A through an Fe(II)- and α-ketoglutarate-dependent oxidative process (Aik et al., 2014).
Binding proteins
1.1.3
Or readers, specifically recognize m6A-modified RNAs and mediate their functional outcomes. The YT521-B homology (YTH) domain family is a major class of readers, including cytoplasmic members (YTHDF1, YTHDF2, YTHDF3) and nuclear members (YTHDC1, YTHDC2) (Zhang et al., 2010). They perform distinct functions: YTHDF1 and YTHDF3 promote translation, while YTHDF2 facilitates mRNA decay (Wang and Lu, 2021; Shi et al., 2017). YTHDC1 regulates mRNA splicing and nuclear export, and YTHDC2 can promote translation while also affecting RNA stability (Xiao et al., 2016). Other important readers include insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs), which stabilize target mRNAs (Huang et al., 2018; Duan et al., 2024), and heterogeneous nuclear ribonucleoproteins (HNRNPs), which are involved in splicing regulation (Zhou et al., 2019; Liu et al., 2015; Alarcon et al., 2015).
The coordinated actions of writers, erasers, and readers establish m6A as a precise and reversible layer of post-transcriptional regulation, influencing diverse biological processes and disease states.
Analyzing RSA pathogenesis from the perspective of the potential role of m6A methylation modification
1.2
A successful pregnancy involves establishing and maintaining pregnancy throughout its stages, culminating in labor. Failure can lead to adverse pregnancy outcomes. RSA, as an early pregnancy failure, has complex and multiple causes. Understanding it involves examining crucial stages of normal pregnancy maintenance, including normal embryogenesis, maternal-fetal interface establishment, and immune microenvironment regulation at this interface. These stages require several cytokines, each regulated at the RNA transcriptional level. Therefore, m6A methylation modifications affecting RNA transcription are crucial in gestation and influence pregnancy outcomes.
Biological functions and pathological effects of m6A methylation modifications in pregnancy establishment and RSA
1.3
Embryogenesis and development
1.3.1
Embryogenesis and development are critical for establishing pregnancy, as embryo quality is a major determinant of early pregnancy outcome. This process encompasses oogenesis, spermatogenesis, fertilization, and subsequent embryonic cell proliferation and differentiation. Evidence indicates that m6A methylation regulates oogenesis, spermatogenesis, and embryonic development, thereby affecting the quality of the fertilized egg and the early embryo.
Oogenesis
1.3.1.1
In mammals, the development of a mature, fertilization-competent oocyte follows a defined pathway: primordial germ cells differentiate into oogonia, which enter meiosis to form primary oocytes. These become enclosed by granulosa cells to form primordial follicles. After puberty, selected follicles mature and release a fertilizable oocyte. The quality of this process is paramount for female fertility and successful pregnancy (Laisk et al., 2019; Feng et al., 2014; Li and Albertini, 2013).
Previous studies have demonstrated that m6A methylation regulates various cellular processes in the female reproductive system, including cell proliferation, differentiation, metabolism, and the cell cycle (Huang and Chen, 2023). Direct evidence confirms its essential role in oogenesis. Reader proteins such as YTHDC1, YTHDC2, YTHDF1, and YTHDF2 participate in oocyte formation, development, and maturation by modulating RNA transcription and translation, which is critical for early embryogenesis (Kasowitz et al., 2018; Qi et al., 2022; Zhao et al., 2021; Ivanova et al., 2017). Nuclear YTHDC1 promotes pre-mRNA splicing; its deletion leads to oocyte arrest at the primary follicle stage and infertility (Kasowitz et al., 2018). Cytoplasmic YTHDF2 mediates maternal mRNA degradation during meiotic maturation, ensuring proper transcriptome construction in mature oocytes and facilitating early embryonic cleavage (Kasowitz et al., 2018; Zhao et al., 2021).
Methyltransferases (writers) such as METTL3 and METTL14 are also key for follicular development and oocyte maturation (Lin et al., 2017). Demethylases (erasers) such as ALKBH5 regulate maternal mRNA degradation by removing m6A marks. ALKBH5 deficiency leads to abnormal m6A accumulation, excessive translation, meiotic defects, chromosomal misalignment, spindle anomalies, and ultimately infertility (Bai et al., 2023). Another demethylase, FTO, maintains chromatin stability and accessibility by acting on long-interspersed element-1 (LINE1) RNA; loss of FTO causes LINE1 RNA degradation, chromatin compaction, and abnormal oocyte development (Wei et al., 2022).
Furthermore, differentially expressed genes involved in steroidogenesis, granulosa cell proliferation, and follicular development are highly enriched in m6A modifications, indicating that m6A not only regulates mRNA fate within oocytes but also indirectly influences the follicular microenvironment via granulosa cells (Cao et al., 2020). Dynamic changes in m6A modifications within granulosa cells across different follicular stages suggest involvement in regulating the oocyte-supporting environment (Cao et al., 2020).
Additionally, m6A modification is closely associated with long non-coding RNAs (lncRNAs) in reproductive disorders. lncRNAs are themselves highly enriched in m6A marks and play crucial roles in embryo implantation. In the endometrium of patients with recurrent implantation failure (RIF), m6A modification patterns of lncRNAs are significantly altered, affecting the expression of key implantation-related factors such as LIF, thereby disrupting the implantation window (Zhang L. et al., 2021; Liu HT. et al., 2022; Wang et al., 2024).
Spermatogenesis
1.3.1.2
In mammals, spermatogenesis not only involves the stages of proliferation, meiosis, and spermiogenesis but is also critically regulated by epigenetic mechanisms, particularly N6-methyladenosine (m6A) modification (Tang et al., 2018). As the most prevalent modification of eukaryotic mRNA, m6A plays an essential role in maintaining the homeostasis, proliferation, and differentiation of spermatogonial stem cells (SSCs) into spermatocytes (Tang et al., 2018). The loss of m6A-regulating methyltransferases METTL3 and METTL14 disrupts post-transcriptional regulation in SSCs, leading to impaired spermatogenesis, indicating that m6A is deeply involved in the early stages of sperm development (Lin et al., 2017).
As SSCs transition to spermatocytes and enter meiosis, m6A reader proteins such as YTHDC2 begin to play a key role at the molecular level (Hsu et al., 2017). YTHDC2 regulates the translation and degradation of target mRNAs by binding to m6A-modified sites. Its expression is significantly upregulated prior to the onset of meiosis in spermatocytes, and its deletion results in meiotic arrest at the zygotene stage, blocking further spermatogenesis and ultimately inducing spermatocyte apoptosis (Hsu et al., 2017). Additionally, YTHDC2 interacts with multiple mRNA metabolic complexes, including XRN1, UPF1, and MOV10, to control transcript fate and maintain mRNA homeostasis during spermatogenesis (Hsu et al., 2017).
In the later stages of spermatogenesis, particularly during the haploid phase, the m6A demethylase ALKBH5 primarily functions in round spermatids (Tang et al., 2018). By removing m6A marks from long 3′-UTR transcripts, ALKBH5 ensures proper splicing and stable storage of these early-reserved mRNAs, thereby supporting late-stage sperm assembly (Tang et al., 2018). However, the loss of ALKBH5 function leads to aberrant splicing, with transcripts being incorrectly processed into truncated fragments and prematurely degraded. This results in disrupted expression of proteins essential for spermiogenesis and causes defective sperm development and infertility (Tang et al., 2018).
Even after the completion of spermatogenesis, the subsequent functional performance of sperm remains influenced by m6A regulation. Processes such as the acrosome reaction, capacitation, and fusion with the oocyte rely not only on structural maturation but also on the regulated expression of sperm-specific mRNAs and proteins. m6A-mediated regulation of mRNA stability and translational efficiency during these stages may indirectly affect sperm viability and fertilization potential.
Moreover, the regulatory role of m6A extends into early embryonic development (Wang et al., 2023). From the zygote to the blastocyst stage, m6A modification is crucial for maternal mRNA degradation, zygotic genome activation (ZGA), and embryonic fate determination. Studies have revealed that key developmental transcription factors such as Nanog, Sox2, and Oct4 carry m6A marks, which are closely associated with their mRNA stability and translational efficiency. In addition, m6A enhances miRNA-mediated degradation of maternal transcripts and regulates the expression of retrotransposons, playing a vital role in maintaining epigenetic homeostasis during embryogenesis (Wang et al., 2023).
Therefore, from SSC fate determination and the successful progression of meiosis to the maturation of haploid sperm and their roles in fertilization and embryonic development, m6A modifications and their regulatory factors—such as METTL3/METTL14, YTHDC2, and ALKBH5—form an epigenetic regulatory axis that spans the entire process of spermatogenesis and extends into early embryogenesis. This provides critical insights into the multilayered mechanisms underlying sperm development and the molecular basis of male infertility (Liu HT. et al., 2022; Wang et al., 2024).
Establishment and development of the maternal-fetal interface
1.3.2
The maternal-fetal interface, located between the mother and the fetus, includes maternal meconium and fetal placenta components. This interface is crucial for maintaining pregnancy by providing nutritional support, acting as a protective barrier for the fetus, and ensuring maternal immune tolerance (Tang et al., 2018). Inadequate development of this interface can lead to adverse pregnancy outcomes, significantly RSA (Wang et al., 2023). Placental tissue predominantly contains trophoblast cells, while endometrial tissue comprises stromal cells undergoing endometrial metamorphosis. Recent studies increasingly show the significant role of RNA m6A modification in cell proliferation, differentiation, invasion, and endometrial metamorphosis, affecting the establishment and development of the maternal-fetal interface.
Trophoblast invasion
1.3.2.1
During early implantation, trophoblast cells adhere to the maternal endometrium, proliferate, differentiate, and invade the decidua and spiral artery walls. In normal pregnancy, this invasion induces vascular remodeling to ensure adequate blood supply. In contrast, restricted trophoblast invasion is observed in conditions like early-onset preeclampsia (ePE) and intrauterine growth restriction (IUGR), leading to failed spiral artery remodeling and placental insufficiency (Kadyrov et al., 2006).
Aberrant invasion is closely associated with dysregulated expression of m6A-modifying enzymes in trophoblasts (Huang N. et al., 2022). For instance, WTAP expression is downregulated in ePE. Its loss reduces m6A methylation on downstream target HMGN3 mRNA, compromising its stability and weakening its promotive effect on trophoblast migration and invasion. This effect is mediated through reader protein IGF2BP1 (Bian et al., 2022).
In RSA villous tissues, the core writer METTL3 is significantly downregulated. One of its targets, ZBTB4—a transcription factor that inhibits migration and proliferation—exhibits increased mRNA stability and expression when m6A is reduced, thereby suppressing trophoblast invasion. METTL3-mediated m6A modification shortens ZBTB4 mRNA half-life; METTL3 insufficiency allows ZBTB4 to escape degradation (Huang N. et al., 2022).
Conversely, the demethylase ALKBH5 is abnormally upregulated in RSA. High ALKBH5 expression reduces the m6A level and stability of target gene CYR61 (an extracellular matrix protein critical for placentation), weakening its supportive function in trophoblast migration and invasion (Li et al., 2019). ALKBH5 overexpression is also associated with ferroptosis regulation, potentially influencing trophoblast survival by modulating genes like ferritin light chain (FTL) (Xie et al., 2016).
Excessive trophoblast apoptosis is another key factor in ePE and IUGR. Interestingly, in maternal anemia, trophoblast invasiveness is enhanced despite hypoxia, suggesting that apoptosis, rather than hypoxia per se, may be the primary mechanism affecting vascular remodeling in some pathologies (Kadyrov et al., 2006). m6A modification is involved in regulating programmed cell death pathways, including ferroptosis (Yang and Stockwell, 2016). ALKBH5 expression changes may influence FTL expression, modulating cellular iron storage and trophoblast sensitivity to ferroptosis (Gentric et al., 2019).
Furthermore, circular RNAs (circRNAs) are extensively involved in trophoblast function. Multiple circRNA–miRNA–mRNA axes have been confirmed in pregnancy complications. For example, circ_0017068 induces M1 macrophage polarization and trophoblast apoptosis via the miR-512-5p/MITA pathway and is implicated in RSA (Liu et al., 2023). Others like circ_0074371, circ_0011460, and circ_0037078 play roles in fetal growth restriction and preeclampsia by regulating trophoblast behavior (Zhou et al., 2018; Fan et al., 2021; Yao et al., 2022; Zou and Mao, 2022).
Decidualization
1.3.2.2
Before embryo implantation, the endometrium must undergo decidualization, a process where endometrial stromal cells (ESCs) transform into decidual stromal cells (DSCs) under specific hormonal cues (Kagawa et al., 2022; Lee and DeMayo, 2004; Gellersen and Brosens, 2014; Owusu-Akyaw et al., 2019). Impaired decidualization can lead to reduced uterine receptivity, inadequate vascular remodeling, and insufficient fetal nutrient supply, all contributing to recurrent miscarriage (Ng et al., 2020; Salker et al., 2010; Lucas et al., 2020; Chang et al., 2022).
Various m6A-regulating enzymes modulate decidualization. METTL3, as an m6A writer, suppresses the expression of key decidualization genes such as progesterone receptor (PGR) (Zheng et al., 2023), forkhead box O1 (FOXO1) (Li et al., 2023), and homeobox A10 (HOXA10) (Xue et al., 2021) through m6A-mediated mRNA degradation. METTL3-mediated m6A enhances the binding of reader YTHDF2 to target mRNAs, promoting their decay and weakening DSC functional transformation (Li et al., 2023). Increased enhancer of zeste homolog 2 (EZH2) expression in ESCs from endometriosis patients is linked to elevated m6A levels and repression of decidual genes like IGFBP1, suggesting crosstalk between m6A and histone modifications (Lin et al., 2024).
A clear connection exists among m6A imbalance, decidualization defects, and miscarriage. Biological samples from miscarriage patients show decreased FTO and increased METTL3 expression, accompanied by downregulation of decidual markers (IGFBP1,PRL,FOXO1), supporting that m6A imbalance disrupts DSC formation (Qiu et al., 2021). FTO downregulation increases m6A burden on key mRNAs (e.g.,,HLA-G,VEGFR), affecting immune tolerance and angiogenesis at the maternal-fetal interface (Qiu et al., 2021). The reader YTHDF2 regulates EZH2 mRNA stability, epigenetically suppressing decidualization-related genes and aggravating decidual defects (Lin et al., 2024).
CircRNA expression is also abnormal in decidua of patients with pathological pregnancy loss and is closely linked to m6A modifications. Certain circRNAs may regulate m6A modulator expression via competing endogenous RNA (ceRNA) mechanisms, indirectly influencing methylation levels of key decidual mRNAs (Cui et al., 2023).
Immune microenvironment regulation at the maternal-fetal interface
1.3.3
The maintenance of a successful pregnancy relies on a specialized immune microenvironment that facilitates maternal immune tolerance toward fetal alloantigens, preventing immunological conflict. A substantial body of research has established a close association between immune microenvironment imbalance and recurrent spontaneous abortion (RSA), particularly in unexplained cases where immune rejection is often a primary cause (Recurrent abortion, 1993; Alijotas-Reig et al., 2014; Zenclussen et al., 2007; La Rocca et al., 2014). This microenvironment comprises various immune cells, including decidual natural killer (dNK) cells, decidual macrophages (dMΦ), and decidual T cells, which collectively maintain a dynamic equilibrium balancing tolerance and defense (Xu et al., 2021).
In normal pregnancy, dNK cells exhibit a low-cytotoxicity, high-secretory phenotype (CD56^∧^brightCD16^∧^-), assisting in spiral artery remodeling and receiving inhibitory signals from fetal HLA molecules to maintain tolerance (Xu et al., 2021). dMΦ are predominantly polarized toward the M2 phenotype, contributing to angiogenesis and tissue repair (Erlebacher, 2013). Regulatory T cells (Tregs) migrate to the interface, suppressing effector T cell activity through cytokines like TGF-β and IL-10, thereby preventing fetal rejection (Alijotas-Reig et al., 2014).
As a post-transcriptional epigenetic regulator, RNA m6A methylation plays a crucial role in maintaining immune homeostasis at the maternal-fetal interface. “Writer” enzymes such as METTL3, METTL14, and WTAP add m6A modifications to specific transcripts, affecting the development and function of T cells, NK cells, and macrophages. For example, METTL3 can promote the degradation of SOCS family mRNAs in CD4^+^ T cells, activating the IL-7–STAT5 axis to maintain T cell homeostasis (Wu et al., 2023). In Tregs, m6A modification enhances their suppressive function (Wu et al., 2023). Conversely, “eraser” enzymes such as FTO and ALKBH5 remove m6A marks to modulate the expression of key factors like VEGF and STAT1, influencing macrophage polarization and NK cell function (Wu et al., 2023).
In RSA, immune microenvironment imbalance is closely linked to aberrant m6A modification patterns (Tsui et al., 2006). Inflammatory stimuli such as IL-6 and IL-1β can drive naïve CD4^+^ T cells toward pro-inflammatory Th17 differentiation while suppressing Treg formation, intensifying local inflammation and impairing implantation (Bansal, 2010). Studies have shown that uterine tissues from RSA patients exhibit decreased expression of METTL3 and WTAP, along with elevated expression of FTO (Tsui et al., 2006). These changes may interfere with T cell differentiation, decidualization, trophoblast invasion, and vascular remodeling, increasing the risk of pregnancy failure (Tsui et al., 2006).
Moreover, m6A modification is involved in regulating inflammatory responses. During pathogenic invasion or inflammation, METTL3/14 expression can be upregulated, increasing m6A methylation and activating pro-inflammatory pathways. Melatonin, through its receptor MTNR1B, may modulate this process by restoring FTO function and reducing METTL3 expression, thereby helping to maintain the immunosuppressive environment essential for early pregnancy (Zhao et al., 2024).
Correlation between m6A methylation modifications in risk factors and RSA occurrence
1.4
Successful pregnancy is a long-term and complex process with several risk factors that can lead to miscarriage. Addressing these risks at their source is crucial for treating recurrent miscarriages, making it a significant aspect of RSA exploration. Accepted factors, such as autoimmune diseases, obesity, and endocrine imbalances, are widely recognized. Recent studies have found the biological role of m6A methylation in these risk factors, offering new insights into RSA.
Autoimmune dieases
1.4.1
The immune environment is critical for preventing maternal-fetal conflict. Dysregulated immune responses are closely associated with an increased risk of RSA (Alecsandru et al., 2021; Vomstein et al., 2021). The human leukocyte antigen (HLA) system, including allelic polymorphisms and maternal-fetal incompatibility, may contribute to RSA pathogenesis (Prins et al., 2014; Meuleman et al., 2015; Meuleman et al., 2018). Specific combinations of fetal HLA-C and maternal killer-cell immunoglobulin-like receptors (KIRs) can lead to either insufficient placental invasion or heightened immune aggression, triggering miscarriage (Vomstein et al., 2021). HLA class II genes (e.g.,,DRB1,DQB1) influence immune tolerance; susceptible alleles may enhance pro-inflammatory T cell activation and impair regulatory T cell (Treg) differentiation, increasing fetal rejection risk (Aimagambetova et al., 2019). Maternal immunological memory against fetal HY antigens (from a male fetus) can also contribute to miscarriage in subsequent pregnancies (Aruna et al., 2011).
The transcription factor FOXP3 is pivotal for Treg differentiation and function. Polymorphisms in the FOXP3 gene may affect its expression and influence RSA susceptibility (Abdukassimova et al., 2021). Importantly, m6A modification plays a key role in regulating FOXP3 expression by modulating mRNA stability and translational efficiency (Bahia et al., 2022). Specifically, METTL14-catalyzed m6A helps sustain FOXP3 expression in Tregs and maintains their immunosuppressive function (Liu Y. et al., 2022). Loss of METTL14 reduces Treg numbers and function, diminishing maternal immune tolerance and elevating miscarriage risk (Liu Y. et al., 2022). Additionally, m6A regulatory enzymes such as ALKBH5, YTHDF3, and YTHDC1 are involved in HLA gene expression regulation. Imbalances in their expression can disrupt maternal-fetal immune homeostasis and increase RSA risk (Zhang X. et al., 2021; Sun et al., 2021).
Endocrine disorders
1.4.2
Approximately 20% of RSA cases are associated with endocrine dysfunction (Quenby et al., 2021; Khalife et al., 2019), most commonly involving progesterone deficiency and thyroid dysfunction.
Progesterone
1.4.2.1
Progesterone is central to embryo implantation and early gestation. Reduced progesterone levels are found in many RSA patients (Coomarasamy et al., 2021; Duane et al., 2022). Its action is mediated through the progesterone receptor (PGR). Studies show that PGR mRNA expression is regulated by METTL3-mediated m6A methylation, and METTL3 deficiency significantly reduces PGR levels (Zheng et al., 2023). YTHDF1 is also involved in PGR mRNA translation. Luteal phase insufficiency and low progesterone impair endometrial support, leading to implantation failure and early loss (Khalife et al., 2019). In autoimmune RSA models, exogenous progesterone supplementation improves outcomes by upregulating the Treg/NK cell ratio and reducing pro-inflammatory cytokines (Chen et al., 2021).
Thyroid function
1.4.2.2
Thyroid autoimmunity (TAI), indicated by positivity for thyroid peroxidase antibody (TPO-Ab) and elevated thyroid-stimulating hormone (TSH), is significantly associated with early miscarriage in RSA patients (Wang L. et al., 2021). Polymorphisms in the FTO gene are closely associated with TSH levels, with certain genotypes identified as risk factors for elevated TSH (Wang L. et al., 2021). FTO, highly expressed in the hypothalamus and pituitary, regulates TSH synthesis and secretion by modulating mRNA methylation, influencing the hypothalamic-pituitary-thyroid axis (Dwivedi et al., 2012).
Obesity
1.4.3
Obesity is a significant risk factor for miscarriage, involving oxidative stress, systemic inflammation, reduced uterine receptivity, and impaired oocyte metabolism (Eapen et al., 2021). This risk is heightened in women with a history of RSA (Dimitriadis et al., 2020; Cavalcante et al., 2019). The development of obesity is regulated by methylation mechanisms. The FTO gene is a well-known susceptibility gene for adipogenesis, regulating adipocyte differentiation through m6A demethylation (Frayling et al., 2007; Locke et al., 2015). Its risk allele (e.g., rs9939609 A) increases body mass index (BMI) and indirectly raises the risk of type 2 diabetes and recurrent miscarriage. FTO’s high hypothalamic expression also suggests a role in central energy intake regulation, potentially disrupting ovulation and endometrial preparation (Frayling et al., 2007).
Other m6A-associated enzymes affect lipid synthesis. Increased METTL3 expression can suppress adipocyte proliferation, while METTL3 deficiency reduces m6A on fatty acid synthase (Fasn)mRNA, impairing fatty acid metabolism (Liu et al., 2019; Xie et al., 2019). In hepatic tissue, METTL3 promotes lipid synthesis by stabilizing lipogenic genes; its overexpression exacerbates hepatic fat accumulation and insulin resistance (Xie et al., 2019). In hyperlipidemic mouse livers, METTL3 deficiency reduces global m6A and hepatic lipid accumulation by increasing the stability of metabolic genes like Lpin1 (Li et al., 2020). The reader YTHDC2 is upregulated in hepatocytes in obesity and non-alcoholic fatty liver disease (NAFLD), associated with triglyceride accumulation. YTHDC2 recognizes and degrades m6A-modified lipogenic gene mRNAs, inhibiting excessive lipid synthesis (Zhou et al., 2021; De Jesus et al., 2019).
Thrombosis
1.4.4
Thrombophilia is a major cause of miscarriage, disrupting the balance between coagulation and fibrinolysis essential for placental perfusion. m6A RNA methylation may participate in thrombosis by regulating gene expression. During venous thromboembolism (VTE), activated neutrophils release neutrophil extracellular traps (NETs), which disrupt endothelial barrier function and promote iron-dependent lipid peroxidation (ferroptosis), fostering thrombosis (Li et al., 2024).
NETs markedly increase m6A RNA modification levels in endothelial cells, particularly by upregulating METTL3, which enhances m6A methylation and stability of TLR4 mRNA. This activates the TLR4/NF-κB pathway, promoting pro-inflammatory cytokine release, oxidative stress, and ferroptosis—a vicious cycle of inflammation-ferroptosis-thrombosis. Inhibition of METTL3 can reverse TLR4/NF-κB activation, alleviating endothelial damage and thrombogenesis (Li et al., 2024; Zhang et al., 2024).
Bioinformatics analyses reveal aberrant expression of various m6A regulatory factors (e.g., METTL3, RBM15, FTO, YTHDF1/3, HNRNPC) in the peripheral blood of VTE patients, associated with altered immune cell infiltration and autophagy. Among these, YTHDF3 is a central node linking m6A modification, immune responses, and autophagy, exacerbating inflammation and thrombosis (Zhang et al., 2024).
Conclusion
2
RSA poses a significant threat to women’s reproductive health, imposing substantial burdens on patients, families, and society. Its etiology is more complex and less understood than that of sporadic abortion, and treatment options remain limited. Therefore, investigating novel, multifaceted therapeutic strategies targeting its pathogenesis is imperative.
Epigenetics, particularly the reversible m6A RNA methylation which does not alter the genetic code, stands out as a promising field. It offers fresh insights and potential targets for developing diverse therapeutic approaches aimed at addressing the pathogenesis of RSA. In this review, we have examined the biological functions of m6A in key stages of pregnancy establishment and maintenance, and explored its potential involvement in pathways influencing established risk factors for RSA.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abdukassimova M. Kanabekova P. Bauyrzhanova Z. Ukybassova T. Kaldygulova L. Imankulova B. (2021). Association of human forkhead box protein 3 (FOXP 3) gene polymorphisms with idiopathic recurrent pregnancy loss among kazakhstani women. Gene 801, 145835. 10.1016/j.gene.2021.145835 34274475 · doi ↗ · pubmed ↗
- 2Aik W. Scotti J. S. Choi H. Gong L. Demetriades M. Schofield C. J. (2014). Structure of human RNA N(6)-methyladenine demethylase ALKBH 5 provides insights into its mechanisms of nucleic acid recognition and demethylation. Nucleic Acids Res. 42 (7), 4741–4754. 10.1093/nar/gku 085 24489119 PMC 3985658 · doi ↗ · pubmed ↗
- 3Aimagambetova G. Hajjej A. Malalla Z. H. Finan R. R. Sarray S. Almawi W. Y. (2019). HLA-DQ, and HLA-DP loci are linked with altered risk of recurrent pregnancy loss in Lebanese women: a case-control study. Am. J. Reprod. Immunol. 82 (4), e 13173. 10.1111/aji.13173 31339184 · doi ↗ · pubmed ↗
- 4Alarcon C. R. Goodarzi H. Lee H. Liu X. Tavazoie S. Tavazoie S. F. (2015). HNRNPA 2B 1 is a mediator of m(6)A-Dependent nuclear RNA processing events. Cell. 162 (6), 1299–1308. 10.1016/j.cell.2015.08.011 26321680 PMC 4673968 · doi ↗ · pubmed ↗
- 5Alecsandru D. Klimczak A. M. Garcia Velasco J. A. Pirtea P. Franasiak J. M. (2021). Immunologic causes and thrombophilia in recurrent pregnancy loss. Fertil. Steril. 115 (3), 561–566. 10.1016/j.fertnstert.2021.01.017 33610320 · doi ↗ · pubmed ↗
- 6Alijotas-Reig J. Llurba E. Gris J. M. (2014). Potentiating maternal immune tolerance in pregnancy: a new challenging role for regulatory T cells. Placenta 35 (4), 241–248. 10.1016/j.placenta.2014.02.004 24581729 · doi ↗ · pubmed ↗
- 7Aruna M. Nagaraja T. Andal Bhaskar S. Tarakeswari S. Reddy A. G. Thangaraj K. (2011). Novel alleles of HLA-DQ and -DR loci show association with recurrent miscarriages among South Indian women. Hum. Reprod. 26 (4), 765–774. 10.1093/humrep/der 024 21325036 · doi ↗ · pubmed ↗
- 8Bahia W. Zitouni H. Kanabekova P. Bauyrzhanova Z. Shaimardanova M. Finan R. R. (2022). Human forkhead box protein 3 gene variants associated with altered susceptibility to idiopathic recurrent pregnancy loss: a retrospective case-control study. Am. J. Reprod. Immunol. 88 (2), e 13551. 10.1111/aji.13551 35452532 · doi ↗ · pubmed ↗