Tongue microstructure physically constrains division of labor in bumblebee foraging
Zexiang Huang, Shumeng Wu, Qinglin Wu, Tianyu Mai, Jieliang Zhao, Bo Wang, Jianing Wu

TL;DR
This paper shows that the physical structure of bumblebee tongues explains why queens stop foraging when workers appear.
Contribution
The study provides physical evidence linking tongue microstructure to the division of foraging labor in bumblebees.
Findings
Bumblebee tongue microstructure affects foraging efficiency.
Physical tongue traits explain caste-specific foraging behavior shifts.
Queens' foraging decline is tied to their less efficient tongue structure.
Abstract
Bumblebee (Bombus terrestris) is a primitively eusocial species that sustains its colonies by dividing foraging duties between queens and workers. Previous research emphasizes hormonal, metabolic, and behavioral contrasts between the castes but seldom considers why queens relinquish foraging once workers emerge. Here, we link foraging efficiency to tongue microstructure, providing physical evidence for the shift in bumblebee foraging behavior.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
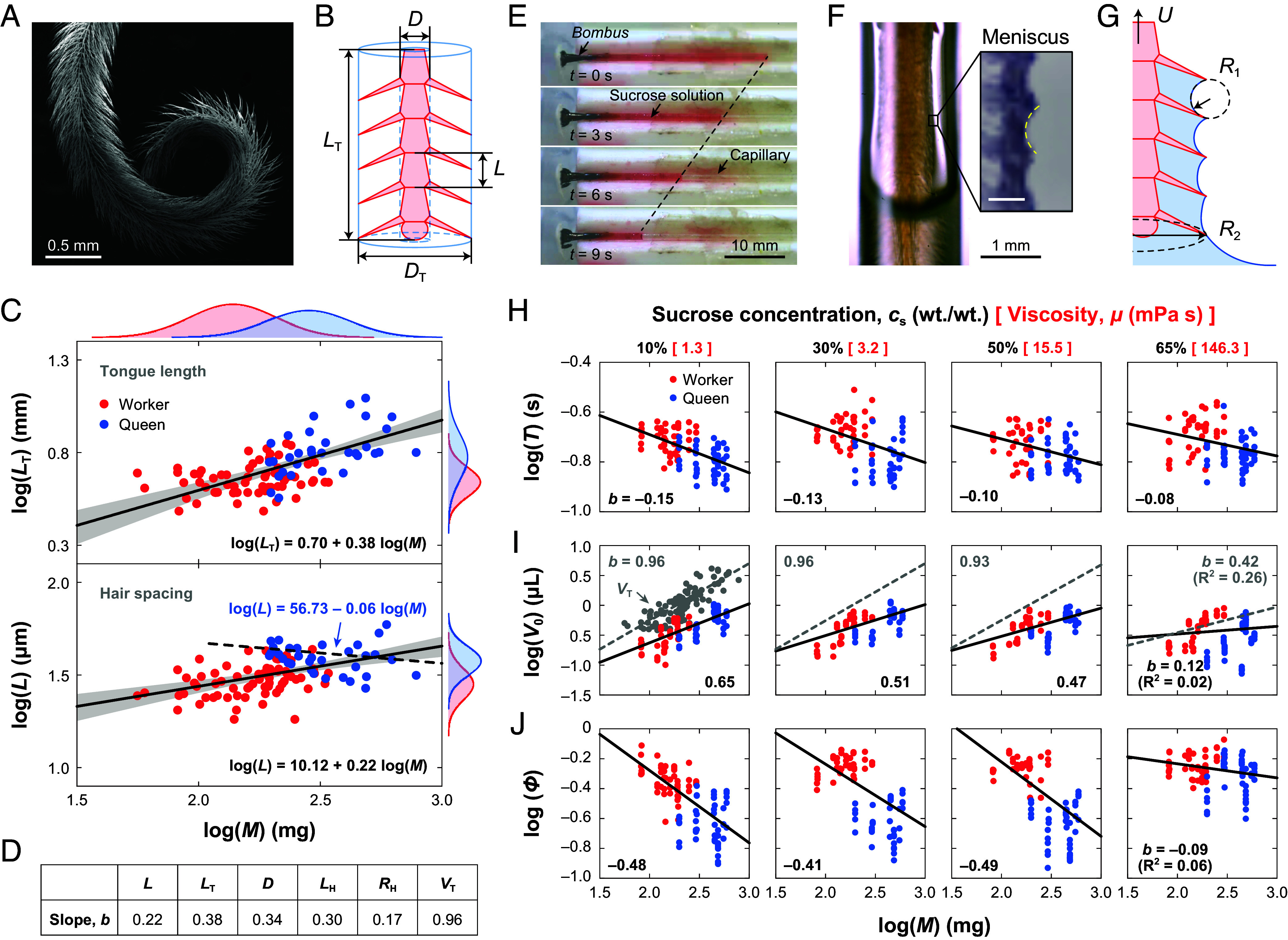 Figure 1
Figure 1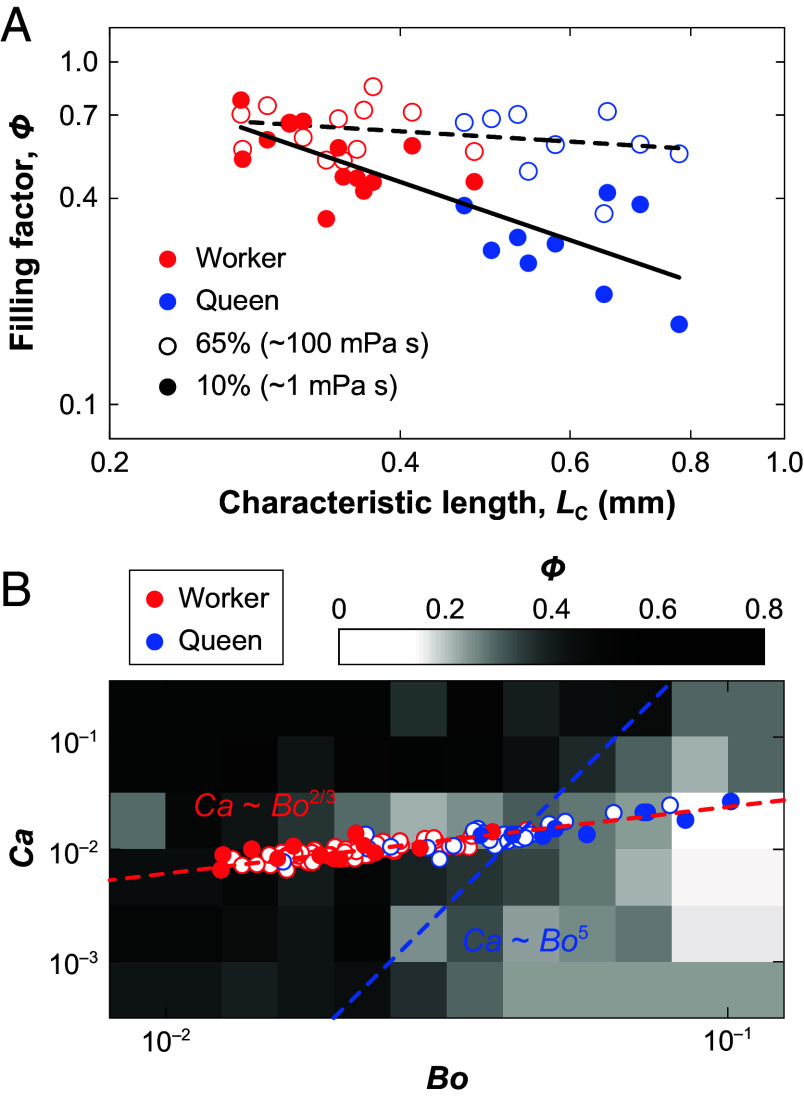 Figure 2
Figure 2- —MOST | National Natural Science Foundation of China (NSFC)501100001809
- —MOST | National Natural Science Foundation of China (NSFC)501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Arachnid Ecology and Behavior · Animal Behavior and Reproduction · Neurobiology and Insect Physiology Research
Understanding how social-insect castes allocate labor has long been central to behavioral ecology, especially regarding how caste-specific morphological and behavioral differences translate into variation in foraging performance (1). Bumblebee (Bombus terrestris) is an influential model because it straddles the transition from primitively to highly eusocial life histories while remaining amenable to both field and laboratory study (2). In bumblebee species, adult queens are always larger than workers, but show virtually no external morphological differences (3). However, queens forage intensively only during nest founding and cease once workers take over, whereas workers continue to collect nectar throughout the season (4). Existing evidence attributes this caste divergence to endocrine regulation, energy metabolism, and task specialization, but no study has yet examined whether tongue microstructure constrains foraging efficiency (5?–7). Here, we integrate postmortem scanning electron microscopy (SEM) with in vivo high-speed filming to establish a form–function relationship of the bee tongue, providing morphological and physical evidence for queen-worker divergence. We show that, although hair spacing increases more slowly than tongue length under allometric growth, their positive scaling still renders the tongues of large queens excessively porous, limiting nectar retention and making queens less efficient nectar foragers than workers.
To sustain efficient nectar intake, bumblebees have evolved an elongated proboscis made up of a pair of galeae and labial palpi, which form a feeding tube surrounding a hairy tongue (Fig. 1A). The tongue, or glossa, partitioned into segments by transverse cuticular rings, each decorated by slender hairs, is the primary feeding structure for extracting nectar from the floral tube. We dissected 99 B. terrestris adults (nworker = 67 and nqueen = 32; body mass M = 65 to 810 mg) to examine their tongue morphology (SI Appendix). Here, we report two critical parameter that capture tongue architecture at distinct scales, namely the macroscale measure of tongue length LT and the microscale measure of hair spacing L. As shown in Fig. 1C, Log-log regressions showed near-isometric scaling of LT but shallow allometric scaling of L. Kernel-density ridges show queens cluster at higher body mass with longer tongues and wider hair spacing. We hypothesize that these geometric differences underpin the caste-specific division of labor during nectar collection.
Lapping, the common nectar-feeding mechanism of bumblebees, involves back-and-forth motion of the tongue to collect nectar (8). To test how hair spacing influences nectar uptake, we recorded videos (nvideo = 3 to 6 per bee) of bumblebees (nworker = 13 and nqueen = 9) capturing artificial nectars with different sugar contents, namely different viscosities (Movie S1). Tracking the liquid–air meniscus inside the capillary allowed us to measure the liquid intake volume per lap V0 and lapping time T of each bee, and compute the ingestion rate Q (Fig. 1E). Across three orders of magnitude in viscosity, lapping time was nearly constant in both castes and varied only weakly with body mass (T ~ M^−0.1^; Fig. 1H), indicating that bees have the ability to adjust retraction force in response to nectar viscosity (8). Queens lapped slightly faster than workers, but resultant differences in retraction speed (U = LT/T) were minor compared to those driven by tongue geometry.
Previous work assumed that the entire space inside the outer envelope of the tongue hairs fills completely with liquid per lap (8, 9). We modeled this space as a cylindrical sleeve with negligible hair volume and defined theoretical nectar-holding capacity as VT = π(DT^2^ − D^2^)LT/4, where DT is outer diameter of the tongue considering hairs, and D is the diameter of tongue base (Fig. 1B). Across individuals, VT scaled approximately in direct proportion to body mass. However, the measured per-lap volume V0 increased with size more slowly than VT (Fig. 1 D and I). Given that lapping time varied little across body sizes (Fig. 1H), the ingestion rate Q = V0/T exhibited the same subisometric scaling as V0. This aligns our laboratory results with field observations showing that larger workers achieve greater absolute nectar loads and intake rates, but that these gains do not increase proportionally with body size (10, 11).
To account for differences in tongue size, we normalized the observed intake volume V0 by VT and defined a filling factor Φ = V0/VT. We found that Φ declined with body mass across viscosities, indicating that size-related differences in nectar retention. In addition, when individuals were matched for body mass or tongue length, queens consistently exhibited lower Φ than workers (Fig. 1J). This difference arises because queens consistently possess wider hair spacing than workers at a given body size (Fig. 1C). Given that each colony provided only a single queen, who was invariably the largest individual (7), queens consistently represented the upper extreme of hair-spacing variation within colonies, thereby exhibiting size-independent hair spacing within queens. Overall, the reduced filling efficiency in queens compared to equally sized workers, along with their consistently wider hair spacing, demonstrates that differences in tongue microstructure drive variation in nectar-uptake performance. Consequently, queens cannot fully exploit the internal tongue volume to capture nectar, which may make them less suited to foraging and promote caste-based division of labor.
Previous theory typically focused on viscous entrainment by the hairy structures, overlooking how the same microstructure provides additional support for the liquid (8, 12, 13). High-speed films recorded a curved meniscus spanning the gaps between hairs during tongue withdrawal, whose curvature imposes additional negative Laplace pressure on the liquid surface (14) (Fig. 1F and Movie S2). Thus, capillary pressure provides a driving force for entrainment here rather than resistance. The result is a positive pressure difference along the tongue, ΔPc ≈ 2γ/L, where γ is surface tension (Fig. 1G and SI Appendix). Accordingly, increasing hair spacing weakens the supportive pressure generated by the liquid bridge, especially in dilute nectar. Our statistical results show that hair spacing increases with body mass with an exponent of ~0.22, below isometry and shallower than the scaling of tongue length LT (Fig. 1C). This underscaling reflects a compromise that partly, though not fully, optimizes microgeometry as body size increases.
To capture the coeffects of viscosity and capillarity, we propose a characteristic length LC = (LLT)^1/2^, which merges the macro- and microscale of hairy tongue and clearly separates the castes (SI Appendix). Workers have small LC that keeps them in the capillary regime, making nectar uptake largely insensitive to viscosity and enabling flexible foraging across nectar types. Queens, with much larger LC, sit closer to the viscous regime and therefore perform better with high-viscosity nectar (Fig. 2A). We farther map the data into a dimensionless space with Bond number Bo = ρgLC^2^/γ and Capillary number Ca = μU/γ = 2μLTf/γ, revealing clear differences between queens and workers. Specifically, constant Φ corresponds to steep isolines in this Bo-Ca space (Ca ~ Bo^5^), indicating that maintaining high nectar retention requires rapidly increasing viscous entrainment at larger scales. However, morphology imposes a much shallower scaling (Ca ~ Bo^2/3^). Consequently, as body size increases, bees cannot sustain the required steep Bo-Ca relationship to maintain high filling factor (Fig. 2B and SI Appendix). This mismatch between physical demands and morphological constraints explains why queens become less efficient nectar foragers.
In summary, our integrative morphological and kinematic analysis reveals that a scaling mismatch between tongue length and hair spacing in bumblebee queens limits their nectar capture. This physical constraint offers a mechanistic explanation for why queens relinquish foraging once workers emerge, underscoring how subtle deviations in microstructure can influence the division of labor in social insects. Our work offers a simple framework for pairing individual-level structure and function to predict performance. Beyond bumblebees, it can be extended across bee taxa by shifting the bee-flower problem from purely geometric matching (tongue length versus corolla depth) to physical matching between tongue porosity and nectar properties (viscosity, surface tension). This framework could apply to other hairy fluid-capture systems and inform bio-inspired porous interfaces.
Material and Methods
The detailed methods are described in SI Appendix. Briefly, B. terrestris queens and workers were offered sucrose solution in a glass capillary, while their feeding behaviors were recorded. The structural parameters of their tongues were measured from the SEM images. For the theoretical analysis, a Bo-Ca framework was used to link tongue structure and function.
Supplementary Material
Appendix 01 (PDF)
Dataset S01 (XLSX)
Movie S1.Feeding behavior of a queen bumblebee with a 50% (wt./wt.) sugar solution.
Movie S2.High-speed video of a queen bumblebee feeding from a glass capillary containing 50% (wt./wt.) sucrose solution.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1C. R. Smith, A. L. Toth, A. V. Suarez, G. E. Robinson, Genetic and genomic analyses of the division of labour in insect societies. Nat. Rev. Genet. 9, 735–748 (2008).18802413 10.1038/nrg 2429 · doi ↗ · pubmed ↗
- 2G. Ghisbain, L. Chittka, D. Michez, Bumblebees. Curr. Biol. 35, R 206–R 211 (2025).40132549 10.1016/j.cub.2025.01.041 · doi ↗ · pubmed ↗
- 3J. J. M. Pereboom, W. C. Jordan, S. Sumner, R. L. Hammond, A. F. G. Bourke, Differential gene expression in queen–worker caste determination in bumble-bees. Proc. R. Soc. B. Biol. Sci. 272, 1145–1152 (2005).10.1098/rspb.2005.3060 PMC 155981016024376 · doi ↗ · pubmed ↗
- 4D. Goulson, Bumblebees: Behaviour, Ecology, and Conservation (Oxford University Press, 2010).
- 5C. Grüter, C. Menezes, V. L. Imperatriz-Fonseca, F. L. W. Ratnieks, A morphologically specialized soldier caste improves colony defense in a neotropical eusocial bee. Proc. Natl. Acad. Sci. U.S.A. 109, 1182–1186 (2012).22232688 10.1073/pnas.1113398109 PMC 3268333 · doi ↗ · pubmed ↗
- 6K. M. Kapheim, Nutritional, endocrine, and social influences on reproductive physiology at the origins of social behavior. Curr. Opin. Insect Sci. 22, 62–70 (2017).28805640 10.1016/j.cois.2017.05.018 · doi ↗ · pubmed ↗
- 7M. Ayasse, S. Jarau, Chemical ecology of bumble bees. Annu. Rev. Entomol. 59, 299–319 (2014).24160431 10.1146/annurev-ento-011613-161949 · doi ↗ · pubmed ↗
- 8A. Lechantre , Essential role of papillae flexibility in nectar capture by bees. Proc. Natl. Acad. Sci. U.S.A. 118, e 2025513118 (2021).33931548 10.1073/pnas.2025513118 PMC 8126835 · doi ↗ · pubmed ↗