Molecular characterization of a clinical ST145 Klebsiella oxytoca strain co-producing KPC-2 and IMP-96 carbapenemases
Hao Liu, Chao Yan, Sibo Wang, Juntian Jiang, Meiling Jiao, Fupin Hu, Xuesong Xu

TL;DR
This study reports a rare Klebsiella oxytoca strain that produces two carbapenem resistance genes, which are located on transferable plasmids, highlighting the risk of spreading antibiotic resistance.
Contribution
The first report of a clinical ST145 Klebsiella oxytoca isolate co-producing blaKPC-2 and blaIMP-96 resistance genes on transferable plasmids.
Findings
The isolate K. oxytoca K31 is resistant to multiple antibiotics including carbapenems and ceftazidime-avibactam.
blaKPC-2 and blaIMP-96 genes are located on IncFIB(K)-like and IncM1 plasmids, respectively, and both are transferable.
94.2% of 104 ST145 K. oxytoca isolates globally carry at least one carbapenem resistance gene.
Abstract
The objective of this study was to investigate the antimicrobial resistance phenotype and genetic characteristics of a clinical ST145 Klebsiella oxytoca isolate co-producing KPC-2 and IMP-96 carbapenemases. The isolate was first identified by MALDI-TOF MS. PCR and Sanger sequencing were used to detect carbapenem resistance genes. Antimicrobial susceptibility testing was performed using broth microdilution. Whole genome sequencing was carried out using Illumina and Nanopore platforms. Conjugation experiments and comparative genomic analysis were used to assess plasmid transferability and the genetic context of resistance genes. A total of 103 K. oxytoca genome sequences were retrieved from public databases and, together with the isolate from this study, used to construct a core genome single nucleotide polymorphism (SNP)-based phylogenetic tree. The antimicrobial resistance genes carried…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
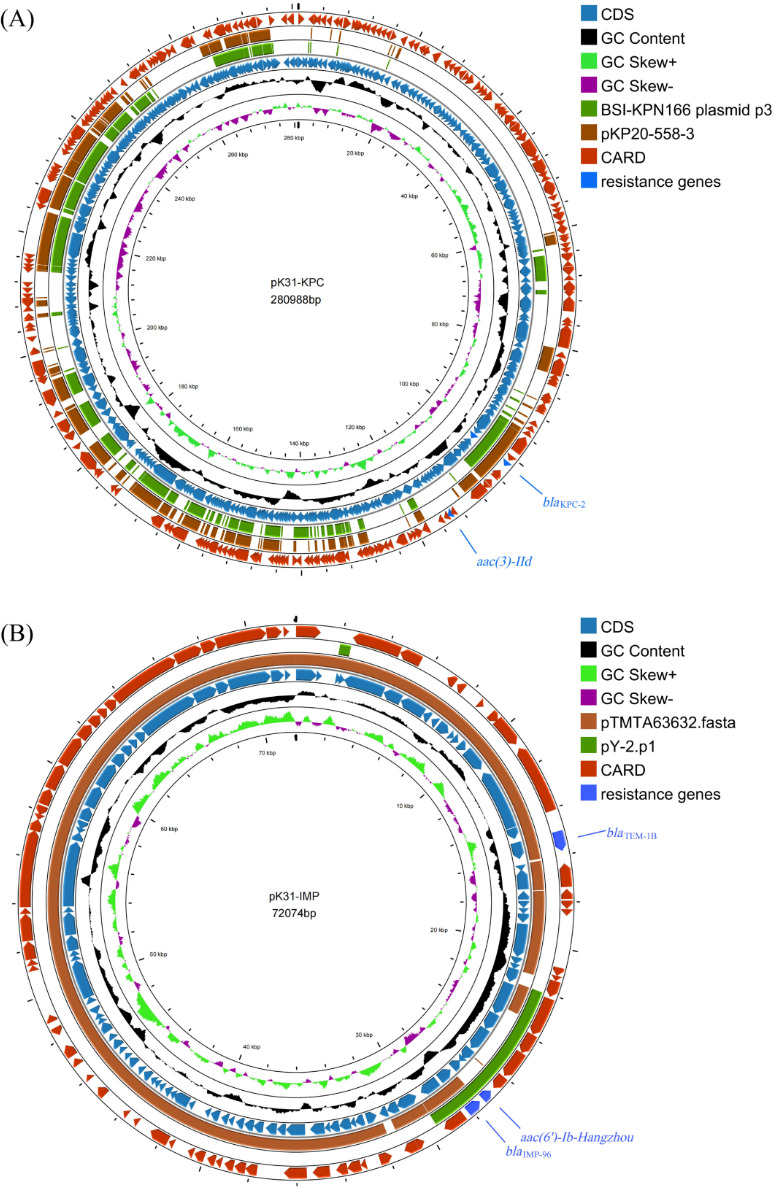 Fig 1
Fig 1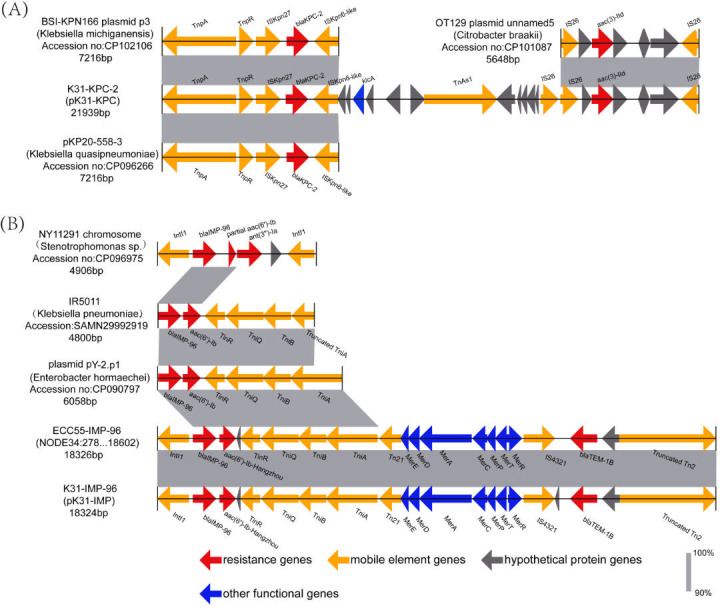 Fig 2
Fig 2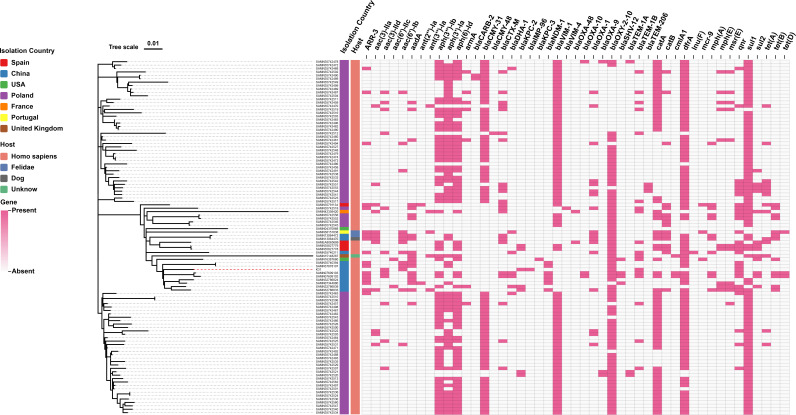 Fig 3
Fig 3| Antibiotic | MIC (μg/mL) | |||||
|---|---|---|---|---|---|---|
| Imipenem | 16 | 2 | 0.5 | 0.125 | ≤0.06 | 1 |
| Meropenem | 8 | 4 | 4 | ≤0.03 | ≤0.03 | 0.125 |
| Meropenem-vaborbactam | 4 | ≤0.03 | 4 | ≤0.03 | ≤0.03 | 0.06 |
| Aztreonam-avibactam | 0.25 | 0.25 | 0.125 | 0.06 | ≤0.03 | 2 |
| Ceftazidime-avibactam | >64 | 0.5 | >64 | 0.25 | 0.06 | 1 |
| Piperacillin-tazobactam | >256 | 128 | 64 | 4 | ≤2 | ≤2 |
| Ceftazidime | >32 | 16 | >32 | 0.5 | ≤0.25 | 1 |
| Ceftriaxone | >32 | 32 | >32 | ≤0.25 | ≤0.25 | 8 |
| Cefepime | 32 | 8 | 32 | 0.5 | ≤0.25 | 0.5 |
| Aztreonam | >128 | 128 | ≤1 | ≤1 | ≤1 | 2 |
| Amikacin | ≤1 | 2 | 2 | 2 | ≤1 | 2 |
| Ciprofloxacin | >8 | ≤0.06 | ≤0.06 | ≤0.06 | ≤0.06 | 0.25 |
| Sitafloxacin | 2 | ≤0.06 | ≤0.06 | ≤0.06 | ≤0.06 | 0.125 |
| Colistin | 0.25 | 0.5 | 0.25 | 0.125 | 0.25 | 1 |
| Tigecycline | 0.25 | 0.125 | 0.125 | 0.125 | ≤0.06 | 4 |
| Eravacycline | 0.25 | 0.125 | 0.125 | 0.06 | ≤0.06 | 2 |
| Chromosome/ | Size (bp) | GC content (%) | Plasmid incompatibility group | Antibiotic resistance genes | Virulence factor genes |
|---|---|---|---|---|---|
| Chromosome | 6,023,973 | 55% |
|
| |
| pK31-KPC | 280,988 | 53% | IncFIB(K)-like | ||
| pK31-IMP | 72,074 | 53% | IncM1 | ||
| pK31-3 | 58,767 | 52% | repB(R1701) |
| |
| pK31-4 | 8,613 | 53% | – | ||
| pK31-5 | 7,066 | 51% | – | ||
| pK31-6 | 5,890 | 46% | – | ||
| pK31-7 | 5,413 | 51% | – | ||
| pK31-8 | 4,048 | 30% | – |
- —Department of Science and Technology of Jilin Provincehttp://dx.doi.org/10.13039/501100011789
- —National Natural Science Foundation of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Bacterial Identification and Susceptibility Testing · Neonatal and Maternal Infections
INTRODUCTION
Klebsiella oxytoca is a non-motile, encapsulated, Gram-negative bacterium that can colonize the gastrointestinal tract of healthy individuals (1) and is also found in other human body sites, such as the skin and oropharynx (2). The pathogenicity of K. oxytoca is closely related to the immune status of the host and is currently considered an opportunistic pathogen associated with nosocomial infections (3, 4). K. oxytoca can cause a range of infections, including antibiotic-associated hemorrhagic colitis, bloodstream infections, urinary tract infections, pneumonia, and intra-abdominal infections (5–9). According to the 2024 data from the China Antimicrobial Surveillance Network (CHINET) (https://www.chinets.com/), the overall detection rate of K. oxytoca among the 458,271 clinical isolates is approximately 1%. In recent years, with the widespread use of carbapenems, carbapenem-resistant K. oxytoca strains have been increasingly reported (10, 11), posing a new threat to public health.
Carbapenem resistance in K. oxytoca is primarily mediated by the production of carbapenemases. According to the Ambler classification system, the most common class A carbapenemase is Klebsiella pneumoniae carbapenemase (KPC), while class B enzymes include Verona integron-encoded metallo-β-lactamase (VIM) and imipenemase (IMP), and class D enzymes are OXA-48-like (2). KPC, the most common class A serine carbapenemase, can hydrolyze carbapenems, penicillins, cephalosporins, and aztreonam through a serine-based active site (12). However, its activity can be inhibited by novel β-lactamase inhibitors, such as avibactam (13). To date, Gram-negative bacteria harboring the blaKPC gene are widely disseminated throughout the world (14). Among the class B metallo-β-lactamases, the IMP-type carbapenemase is encoded by the blaIMP gene, and its enzymatic activity is mediated by zinc ions to hydrolyze the β-lactam ring. Unlike KPC, these enzymes can be inhibited by metal chelators such as EDTA (15). Molecular studies have shown that blaIMP is frequently transferred horizontally between different Gram-negative bacterial species via class 1 and class 3 integrons (16). Infections caused by carbapenem-resistant bacteria are associated with higher mortality rates and represent a major challenge for clinical management.
Strains producing multiple carbapenemases pose significant challenges in clinical treatment, particularly those co-producing class B metallo-β-lactamases and KPC, as they render most β-lactam antibiotics and β-lactam/β-lactamase inhibitor combinations ineffective (17, 18). In 2011, Chinese researchers first reported a clinical case of K. oxytoca co-producing KPC-2 and IMP-8 carbapenemases (19). Notably, the novel blaIMP mutant, blaIMP-96, was first identified in the chromosomal genome of Stenotrophomonas spp. (20). In this study, we describe a clinical isolate of K. oxytoca co-producing KPC-2 and IMP-96 carbapenemases. Molecular epidemiological tracking suggested that the blaIMP-96 gene might have been horizontally transferred across species, possibly originating from Stenotrophomonas spp. and disseminated to Enterobacter hormaechei and K. oxytoca through class 1 integron-based mobile genetic elements. Using whole genome sequencing, conjugation experiments, and antimicrobial susceptibility profiling, we have elucidated the genetic context of these resistance genes and their potential for dissemination. This finding not only reveals the cross-species transmission pathway of a novel carbapenemase gene but also provides crucial molecular epidemiological evidence for the control and prevention of hospital-acquired infections caused by multidrug-resistant pathogens.
MATERIALS AND METHODS
Strains and case information
The K. oxytoca K31 strain was isolated from the puncture drainage fluid of an epidural abscess in a 72-year-old male patient. This patient was admitted to the emergency department of the China-Japan Union Hospital of Jilin University in 2020 for acute onset of cerebral hemorrhage, with concomitant bilateral pneumonia. The patient underwent craniectomy with decompression, hematoma evacuation, and ventriculostomy for drainage. Prophylactic antimicrobial treatment was started with piperacillin-sulbactam (5.0 g Q12h). On postoperative day 2, the patient underwent a tracheotomy due to worsening pneumonia and sputum retention. By day 7, the patient developed fever, and cerebrospinal fluid analysis revealed a significantly elevated white blood cell count, consistent with secondary intracranial infection. Vancomycin (1.0 g Q12h) and meropenem (1.0 g Q6h) were added to control the infection. However, on postoperative day 11, repeat lumbar puncture indicated unresolved infection, with a maximum body temperature of 39.2°C and the development of an epidural abscess around the cranial incision. K. oxytoca K31 was isolated from the abscess drainage. Antimicrobial susceptibility testing showed that the strain was resistant to imipenem, meropenem, and ceftazidime-avibactam. The patient eventually discharged himself due to critical illness. The bacterial species was initially identified using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) (BioMérieux, France), and confirmed by whole genome sequencing.
Antimicrobial susceptibility testing and carbapenemase detection
According to the guidelines of the Clinical and Laboratory Standards Institute (CLSI) (21), the minimum inhibitory concentrations (MICs) of antimicrobial agents were determined using the broth microdilution method. The tested antimicrobial agents included imipenem, meropenem, meropenem-vaborbactam, aztreonam-avibactam, ceftazidime-avibactam, piperacillin-tazobactam, ceftazidime, ceftriaxone, cefepime, aztreonam, amikacin, ciprofloxacin, levofloxacin, colistin, tigecycline, and eravacycline. Quality control procedures and interpretation of results were performed according to the CLSI 2024 breakpoint criteria (21). Antimicrobial susceptibility results for colistin and aztreonam-avibactam were interpreted according to the guidelines of European Committee on Antimicrobial Susceptibility Testing (EUCAST) (22). The MICs of tigecycline and eravacycline were interpreted according to the Enterobacterales breakpoints defined by the U.S. Food and Drug Administration (FDA) (23). Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853 were used as quality control strains. Carbapenemase phenotypic detection was performed using meropenem (10 µg) susceptibility disks alone, and in combination with phenylboronic acid (PBA), EDTA, or both PBA and EDTA, as previously described (24).
Whole genome sequencing and data analysis
Genomic DNA was extracted from the bacterial strain using a DNA extraction kit according to the manufacturer’s instructions. Whole genome sequencing was performed using both short-read sequencing technology (Illumina, San Diego, CA, USA) and long-read sequencing technology (Nanopore, Oxford, UK). Raw sequencing data were filtered for quality and trimmed for read length using Sickle (GitHub). Hybrid assembly was performed with SPAdes version 3.12.0 to generate a high-quality draft genome. Multi-locus sequence typing (MLST) typing was performed using the BIGSdb-Pasteur MLST database (https://bigsdb.pasteur.fr/), while plasmid replicon types and antimicrobial resistance genes were identified using the PlasmidFinder, Pmlst, and ResFinder databases available at the Center for Genomic Epidemiology (http://www.genomicepidemiology.org/services/). Capsule serotypes and virulence genes were analyzed using Kleborate (http://github.com/katholt/Kleborate) (25) and VFDB (https://www.mgc.ac.cn/VFs/) (26). Open reading frames (ORFs) were predicted and annotated using RAST version 2.0 and BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Conjugation experiment and plasmid sequencing
Escherichia coli J53, a strain resistant to sodium azide, was used as the recipient strain to evaluate plasmid transferability using a standard liquid conjugation method. During the experiment, donor and recipient strains were mixed at a 1:1 cell ratio and co-cultured in pre-warmed lysogeny broth (LB), Luria-Bertani formula, at 37°C, followed by co-incubation at 37°C for 18 h. Transconjugants were selected on Mueller–Hinton (MH) agar plates containing dual selective agents (aztreonam 8 mg/L + sodium azide 100 mg/L, or ceftazidime-avibactam 4 mg/L + sodium azide 100 mg/L). Positive colonies growing on the selective media were rapidly identified using MALDI-TOF MS. PCR and PCR-based Sanger sequencing were used to confirm the presence of blaKPC-2 and blaIMP-96 genes in the transconjugants. Plasmid DNA from the transconjugants was extracted using the Qiagen Midi Kit (Qiagen, Hilden, Germany) and further verified by short-read sequencing on the Illumina platform (San Diego, CA, USA).
Phylogenetic analysis
To further investigate the evolutionary relationships among ST145 K. oxytoca isolates, 103 publicly available whole genome sequences were downloaded from the Bacterial and Viral Bioinformatics Resource Center (BV-BRC). These genomes, together with the isolate characterized in this study, were used to construct a single nucleotide polymorphism (SNP)-based phylogenetic tree. SNP distances between bacterial genomes were calculated using Snippy v4.6.0 (https://github.com/tseemann/snippy), run with default parameters. The phylogenetic tree was constructed using FastTree and subsequently visualized and refined via the ChiPlot platform (27). Antimicrobial resistance genes were screened using ABRicate v1.0.1(https://github.com/tseemann/abricate) against the ResFinder database (≥80% identity, ≥90% coverage).
RESULTS
Characteristics of K. oxytoca K31 clinical strain
Antimicrobial susceptibility testing revealed that K. oxytoca K31 was highly resistant to carbapenems, β-lactams, β-lactam/β-lactamase inhibitor combinations, and quinolones. Specifically, the strain was resistant to imipenem (MIC = 16 µg/mL), meropenem (MIC = 8 µg/mL), aztreonam (MIC >128 µg/mL), ceftazidime (MIC >32 µg/mL), ceftriaxone (MIC >32 µg/mL), cefepime (MIC = 32 µg/mL), piperacillin-tazobactam (MIC >256 µg/mL), ceftazidime-avibactam (MIC >64 µg/mL), ciprofloxacin (MIC >8 µg/mL), and sitafloxacin (MIC = 2 µg/mL). However, amikacin (MIC ≤1 µg/mL), meropenem-vaborbactam (MIC = 4 µg/mL), aztreonam-avibactam (MIC = 0.25 µg/mL), eravacycline (MIC = 0.25 µg/mL), tigecycline (MIC = 0.25 µg/mL), and colistin (MIC = 0.25 µg/mL) exhibited good in vitro activity against K31 (Table 1).
Detection of carbapenemase genes and conjugation experiment
PCR-based sequencing results revealed that K. oxytoca K31 simultaneously carries two carbapenemase-encoding genes, blaKPC-2 and blaIMP-96. Conjugation experiments confirmed that plasmids carrying carbapenem resistance genes were successfully transferred to E. coli J53. Transconjugants grown on selective medium containing aztreonam (8 mg/L) and sodium azide (100 mg/L) were verified by PCR to be blaKPC-2-positive, whereas colonies growing on medium containing ceftazidime-avibactam (4 mg/L) and sodium azide (100 mg/L) were confirmed to be blaIMP-96-positive. The conjugation frequency of the blaKPC-2-positive plasmid was 3.6 × 10⁻⁶, while that of the blaIMP-96-positive plasmid was 5.6 × 10⁻⁵ (calculated according to the number of conjugants per initial donor bacteria). Antimicrobial susceptibility testing showed that KPC-positive transconjugants were resistant to meropenem (MIC = 4 µg/mL), intermediate to imipenem (MIC = 2 µg/mL), and sensitive to ceftazidime-avibactam (MIC = 0.5 µg/mL). IMP-positive transconjugants were resistant to meropenem (MIC = 4 µg/mL) and ceftazidime-avibactam (MIC >64 µg/mL) but sensitive to imipenem (MIC = 0.5 µg/mL). Compared with the recipient strain E. coli J53, the MICs of imipenem, meropenem, and ceftazidime-avibactam in KPC-2-positive transconjugants increased by at least 16, 128, and 2-fold, respectively. For blaIMP-96-positive transconjugants, the MICs of imipenem, meropenem, and ceftazidime-avibactam increased by at least 4, 128, and 256-fold, respectively (Table 1).
Whole genome sequencing analysis of plasmid sequences harboring blaKPC-2 and blaIMP-96 and their genetic context
Whole genome sequencing analysis revealed that the K. oxytoca K31 strain (accession number: GCA_051123605.1) belongs to ST145, with a predicted capsular serotype of KL128. The total genome size is 6,466,832 bp, including a 6,023,973 bp chromosome and eight circular plasmids of sizes 280,988, 72,074, 58,767, 8,613, 7,066, 5,890, 5,413, and 4,048 bp. Several common antimicrobial resistance genes were identified, including the carbapenemase genes blaKPC-2 and blaIMP-96, the β-lactam resistance genes blaTEM-1B and blaOXY-2-9, the aminoglycoside resistance genes aac (3)-IId and aac(6')-Ib-Hangzhou, and the quinolone resistance gene qnrS1. Furthermore, several virulence factors were detected, including fyuA, ybtE, ybtT, ybtU, irp1, irp2, ybtA, ybtP, ybtQ, ybtX, ybtS, ompA, entA, entB, and yagZ/ecpA (Table 2). Sequencing analysis identified the conjugative plasmid pK31-KPC, which is 280,988 bp in size and contains three potential origins of replication. On comparison with the PlasmidFinder database, we found that pK31-KPC had the highest similarity to IncFIB(K) plasmids (98.75% identity). Therefore, it was classified as an IncFIB(K)-like plasmid. pK31-KPC contains 346 ORFs, with the blaKPC-2 and aac (3)-IId genes responsible for carbapenem and aminoglycoside resistance, respectively (Fig. 1A). Blast comparison results showed that the multidrug-resistant region containing blaKPC-2 and aac (3)-IId was not highly homologous to any known plasmids in existing databases. The genetic environment surrounding blaKPC-2 showed 99% identity and 99% query coverage with the plasmid p3 from Klebsiella michiganensis (accession number: CP102106) and plasmid pKP20-558-3 from Klebsiella quasipneumoniae (accession number: CP096266). The surrounding genetic structure was tnpA-tnpR-ISKpn27-blaKPC-2-ISKpn6-like. The genetic environment of aac (3)-IId showed high similarity (99% identity and query coverage) to the plasmid OT129 unnamed5 from Citrobacter braakii (accession number: CP101087) (Fig. 2).
(A) Circular comparison of plasmid pK31-KPC with plasmids BSI-KPN166 plasmid p3 (CP102106) and pKP20-558-3 (CP096266). (B) Circular comparison of plasmid pK31-IMP with plasmids pY-2.p1 (CP090797) and pTMTA63632 (AP019667). The different colors in the rings represent different plasmids. Resistance genes are labeled in blue.
Genetic context of plasmid-borne resistance genes.(A) Comparison of the genetic environment of blaKPC-2 on plasmid pK31-KPC with that of plasmids BSI-KPN166 plasmid p3 (CP102106) and pKP20-558-3 (CP096266). The genetic environment of the Aac (3)-IId gene on plasmid pK31-KPC is compared with that of plasmid OT129 plasmid unnamed5 (CP101087). (B) Comparison of the genetic environment of blaIMP-96 on plasmid pK31-IMP with that of the chromosomal region of NY11291 (CP096975), IR5011(SAMN29992919), plasmid pY-2.p1 (CP090797), and ECC55-IMP-96 (NODE34). Genes are represented by arrows and are color-coded based on their functional categories.
Plasmid pK31-IMP, an IncM1-type plasmid of 72,074 bp, contains 98 ORFs. This plasmid harbors the blaIMP-96, blaTEM-1B, and aac(6')-Ib-Hangzhou resistance genes (Fig. 1B), which confer resistance to carbapenems, β-lactams, and aminoglycosides, respectively. Blast comparison analysis showed that plasmid pK31-IMP shares 99% identity and 93% query coverage with plasmid pTMTA63632 from Klebsiella pneumoniae strain TA6363 (accession number: AP019667). Comparative genetic structure analysis revealed that the blaIMP-96 gene is located on the chromosome of Stenotrophomonas spp. strain NY11291 (accession number: CP096975) isolated in Beijing, China, flanked on both sides by integron IntI1. In Klebsiella pneumoniae strain IR5011 (accession number: SAMN29992919), also isolated in Beijing in the same year, blaIMP-96 was identified with the right-side intI1 replaced by a transposon-associated gene cluster (TniR–TniQ–TniB–truncated TniA). A similar structural arrangement was subsequently observed on plasmid pY-2.p1 (accession number: CP090797) of Enterobacter hormaechei Y-2 isolated in Hohhot, China. Notably, we also found that Enterobacter hormaechei strain ECC55, which also carries blaIMP-96, shares an identical multidrug-resistant genetic region with K31. This region has the genetic structure: IntI1-blaIMP-96-aac(6')-Ib-Hangzhou-TniR-TniQ-TniB-TniA-Tn21-MerE-MerD-MerA-MerC-MerP-MerT-MerR-IS4321-blaTEM-1B-Truncated Tn2 (Fig. 2).
Phylogenetic tree and resistance genes of ST145 K. oxytoca
A total of 103 whole-genome sequences of ST145 K. oxytoca isolates were retrieved from public databases and, together with the clinical isolate K31 from this study, used to construct a core genome SNP-based phylogenetic tree and to analyze the antimicrobial resistance gene profiles. The 104 isolates included in the analysis originated from seven countries: Poland (n = 83), China (n = 12), Spain (n = 4), the United States (n = 2), the United Kingdom (n = 1), France (n = 1), and Portugal (n = 1). Among them, 100 isolates were derived from human hosts, three from animals, and one had unknown host information. Phylogenetic analysis revealed that the strains most closely related to K31 were two isolates from China (BioSample: SAMN37878107 and SAMN07609103); however, all three strains differed by more than 60 SNPs, indicating a notable degree of genetic divergence. Resistance gene analysis showed that 94.2% (98/104) of the ST145 isolates carried carbapenemase genes, including blaVIM-1 (n = 85), blaKPC-2 (n = 7), blaNDM-1 (n = 7), blaOXA-48 (n = 2), blaKPC-3 (n = 1), blaVIM-4 (n = 1), and blaIMP-96 (n = 1). In addition, the isolates harbored a wide array of other resistance determinants, including genes associated with resistance to rifampicin (ARR-3), aminoglycosides (aac (3)-IIa, aac (3)-IId, aac(6′)-IIc, aadA, aph(3′)-Ia, aph (6)-Id, etc), other β-lactams (blaCMY, blaCTX-M, blaTEM, blaDHA, etc), chloramphenicol (catA, catB, cmlA1), trimethoprim (dfrA), macrolides (mph(A), mph(E), msr(E)), quinolones (qnr), sulfonamides (sul1, sul2), tetracyclines (tet(A), tet(B), tet(D)), and colistin (mcr-9) ( Fig. 3).
Phylogenetic tree of 104 ST145 K. oxytoca isolates and the distribution of their associated antimicrobial resistance genes.
DISCUSSION
In recent years, there has been a marked increase in the clinical isolation rate of carbapenem-resistant Enterobacterales (28, 29). Data from the SENTRY Global Antimicrobial Surveillance Programme (2013–2023) indicate that among 8962 K. oxytoca isolates, the overall resistance rates to imipenem (1.0%) and meropenem (0.9%) remain relatively low, but the resistance trend is steadily increasing. The imipenem resistance rate increased from 0.4% in 2013 to 1.5% in 2023, while the meropenem resistance rate increased from 0.5% to 1.1% over the same period. In contrast to these global trends, data from the China Antimicrobial Resistance Surveillance System (CARSS) in 2019 indicated significantly higher non-susceptible rates among K. oxytoca isolates, with 6.4% and 5.5% non-susceptible to imipenem and meropenem, respectively (2). Carbapenem-resistant K. oxytoca was first reported in 2003, with the strain harboring the blaKPC-2 gene and exhibiting high resistance to both cephalosporins and carbapenems (10). In 2010, a nosocomial infection outbreak in a Spanish intensive care unit was associated with nine K. oxytoca isolates carrying blaIMP-8, which had no or low resistance to carbapenems (30). In the present study, we identified K. oxytoca K31, which co-harbors the blaKPC-2 and blaIMP-96 resistance genes. This strain demonstrated high resistance not only to imipenem and meropenem, but also to several cephalosporins (ceftazidime, ceftriaxone, cefepime), β-lactam/β-lactamase inhibitor combinations (piperacillin-tazobactam, ceftazidime-avibactam), and quinolones (ciprofloxacin, sitafloxacin). Only amikacin, meropenem-vaborbactam, aztreonam-avibactam, tigecycline, eravacycline, and colistin showed good in vitro activity against this strain.
Carbapenemase production is the primary mechanism of carbapenem resistance in carbapenem-resistant Enterobacterales (CRE). Zhang et al. (31) studied 1105 non-duplicate CRE strains from China and found that NDM and KPC-2 were the most prevalent carbapenemase types. Among the 24 K. oxytoca isolates, carbapenemase genes were detected in all strains, including KPC (29%, 7/24), NDM (42%, 10/24), IMP (25%, 6/24), and KPC + NDM (4%, 1/24) (31). There are also significant regional differences in the resistance genes carried by K. oxytoca strains. For example, strains in Poland predominantly produce VIM (32), whereas those in the United States are predominantly KPC producers (33). In a multicenter study conducted by Wan et al. in 2023, 25 carbapenem-resistant K. oxytoca isolates were collected from five hospitals in Beijing, Chengdu, Guangzhou, Yinchuan, and Suzhou. Carbapenemase genes were detected in 92.0% (23/25) of the isolates, with two strains lacking carbapenemase genes (34). Interestingly, K. oxytoca strains carrying blaIMP-8 isolated in Spain did not show high-level resistance to carbapenems (30), and conjugation experiments in our study further revealed the influence of carbapenemase production on the resistance phenotype. KPC-2-positive transconjugants showed low-level resistance to meropenem and intermediate resistance to imipenem. The blaIMP-96-positive transconjugants also showed a low level of resistance to meropenem but were sensitive to imipenem. These findings suggest that bacterial resistance is a multifactorial process and that carbapenemase production may be only one component. Alternations in porin structures, overexpression of efflux pumps, and high expression of other non-carbapenem β-lactamases may act synergistically to enhance resistance to carbapenems. In addition, K. oxytoca complex strains often carry the chromosomally encoded blaOXY gene, which confers low-level resistance to penicillin (35). In our study, strain K. oxytoca K31 also carried the chromosomally encoded blaOXY-2-9 gene and plasmid-borne β-lactam resistance genes, including blaTEM-1B, the aminoglycoside resistance genes aac (3)-IId and aac(6')-Ib-Hangzhou, and the quinolone resistance gene qnrS1.
Since the first report of the class A serine carbapenemase KPC-1 in Klebsiella pneumoniae in 2001 (12), Gram-negative bacteria carrying the blaKPC-1 gene and its variants have become widespread worldwide, with blaKPC-2 being the most common subtype. It has been identified in several pathogens, mainly Enterobacterales (such as Klebsiella spp., Escherichia coli, Citrobacter spp.) and non-fermenting bacteria (such as Acinetobacter spp., Pseudomonas spp.) (36, 37). Previous studies have suggested that mobile genetic elements, such as transposons may play an important role in the rapid dissemination of blaKPC (38, 39). blaKPC-2 is often located within the transposon Tn4401, which consists of the transposase gene (tnpA), the resolvase gene (tnpR), and two insertion sequences, ISKpn6 and ISKpn7 (40). In previously reported K. oxytoca isolates, the genetic environment surrounding blaKPC-2 was characterized as ISKpn27-ΔblaTEM-1-blaKPC-2-ΔISKpn6, in which ISKpn6 is arranged in parallel with the blaKPC-2 gene. This structure is presumed to be derived from the Tn4401 transposon (11). In the current study, the K. oxytoca K31 isolate also carried the blaKPC-2 gene. The surrounding genetic structure of blaKPC-2 was similar to that of the classical Tn4401, but with differences in the insertion sequences. Specifically, blaKPC-2 was flanked by ISKpn27 and an ISKpn6-like sequence. This structure shared 99% homology with the plasmids BSI-KPN166 plasmid p3 and pKP20-558-3, suggesting a possible common evolutionary origin. In addition, downstream of blaKPC-2, we identified an aminoglycoside resistance gene aac (3)-IId flanked by the insertion sequence IS26. The genetic structure of this region was very similar to that found in the Citrobacter braakii plasmid OT129. The combined presence of blaKPC-2, aac (3)-IId, and surrounding genes formed a novel multidrug resistance gene island. Existing studies have also confirmed that ISs play a crucial role in mediating plasmid sequence rearrangements and horizontal transfer of resistance genes (41, 42). IMP genes were the first metal-type carbapenemase genes to be discovered (43). Members of the IMP gene family are typically integrated into the genetic structure of class one integrons as gene cassettes (44, 45). The blaIMP-96 gene, which is the focus of this study, is a mutant of IMP-8 that differs by a single nucleotide mutation resulting in a serine-to-glycine substitution at position 214 (S214G). This mutation disrupts the hydrogen bonding with the catalytic residues, thereby enhancing resistance to meropenem. This mutation alters the enzyme’s conformation and disrupts the hydrogen bonding interactions with the catalytic residues, thereby enhancing resistance to meropenem (20). This amino acid substitution has also been observed in E. coli, where it confers increased resistance to certain carbapenem antibiotics (46). The blaIMP-96 gene was first identified on the chromosome of Stenotrophomonas spp. isolated in Beijing, China, in 2011. Sequence comparison revealed that similar genetic structures surrounding blaIMP-96 were also present in Klebsiella pneumoniae strain IR5011, which was isolated in the same city and year, and later in plasmid pY-2.p1 of Enterobacter hormaechei Y-2 recovered in Hohhot, China. Compared with the original structure in the Stenotrophomonas spp. chromosome, the IntI1 element to the right side of the blaIMP-96 gene is replaced by a transposase-associated gene cluster. In IR5011, no IntI1 element was detected upstream of blaIMP-96, which may be attributed to limitations of sequencing coverage or assembly gaps. Interestingly, although blaIMP-96 is located at the beginning of the plasmid sequence in pY-2.p1, a complete IntI1 gene is detected at the other end of the circular structure of the plasmid. Due to the peculiar circular nature of plasmids, IntI1 is actually located upstream of blaIMP-96, suggesting that it may play a role in the transfer of the resistance gene. Furthermore, in our study, Enterobacter hormaechei strain ECC55, which also carries the blaIMP-96 gene, showed a genetic environment that was very similar to the genetic structure of IR5011, plasmids pY-2.p1, and pK31-IMP. These findings indicate that this conserved genetic structure may have remained relatively stable during the vertical dissemination of blaIMP-96 among different bacterial species. Related studies have also reported similar gene structures for plasmids carrying blaIMP-8, such as IntI1-blaIMP-8-aacA4-TniR-TniQ-TniB-TniA (47). Thus, it remains unclear whether the IntI1 element carrying blaIMP-96 originated from a transposition event involving the IntI1 gene cassette from the chromosome of Stenotrophomonas spp. or from a mutation in plasmid-borne blaIMP-8 sequences during adaptive evolution. However, based on existing research findings, we believe that the IntI1-mediated transfer mechanism of blaIMP gene cassettes plays an important role in the spread of resistance genes in Gram-negative bacteria. These results suggest that horizontal gene transfer and homologous recombination mediated by mobile genetic elements are key mechanisms for the widespread transfer of carbapenem resistance genes, significantly accelerating the spread of carbapenem-resistant phenotypes in clinical pathogens. In addition, blaIMP often coexists with other resistance genes within integrons, which, besides aminoglycoside resistance genes, may also include the class D β-lactamase gene blaOXA and the phenicol resistance gene catB3(20), leading to multidrug resistance and complicating the selection of effective clinical antimicrobial therapies.
Genomic analysis of global ST145 K. oxytoca isolates revealed that 94.2% of strains carried carbapenem resistance genes, and all isolates carried multiple additional classes of antimicrobial resistance genes. These findings indicate that ST145 K. oxytoca has emerged as a high-risk, multidrug-resistant clonal lineage. The majority of isolates (79.8%, 83/104) originated from Poland, of which 82 carried blaVIM-1 and one carried blaVIM-4. Although only four isolates were reported from Spain, three of them harbored blaVIM-1, suggesting that blaVIM-1-positive ST145 strains may be widely disseminated across Europe. Among the 12 isolates from China, the most prevalent carbapenemase gene was blaNDM-1 (58.3%, 7/12), consistent with findings reported by Zhang et al. (28). Notably, six ST145 isolates were found to co-harbor both class A and class B carbapenemase genes. Among them, four isolates carried both blaVIM-1 and blaKPC-2, originating from Poland (n = 2) and Spain (n = 2). The remaining two isolates were from China, including the K31 strain described in this study and another strain harboring both blaNDM-1 and blaKPC-2. In addition, three isolates carried the mobile colistin resistance gene mcr-9: one from the United States (harboring blaKPC-3), one from Spain (harboring blaVIM-1), and one from Portugal (with no detectable carbapenemase gene). The co-occurrence of carbapenemase and colistin resistance genes in these isolates poses a significant challenge to clinical management.
Conclusion
In conclusion, our study confirmed the simultaneous presence of the blaKPC-2 and blaIMP-96 resistance genes in a K. oxytoca strain. These two carbapenemase genes were located on IncFIB(K)-like and IncM1-type plasmids, respectively, both of which possess conjugative transfer capabilities. We also analyzed the genetic structures surrounding the blaKPC-2 and blaIMP-96 genes. These results suggest that resistance-encoding genes can be captured by mobile genetic elements, forming stable, heritable composite transposons, integrons, or resistance gene islands, which in turn facilitate horizontal gene transfer between bacterial species. The widespread dissemination of resistance genes carried by mobile genetic elements between bacterial species not only accelerates the evolution of resistant bacteria but also poses a serious challenge to current antimicrobial treatment strategies. In particular, the emergence and potential spread of Enterobacterales strains co-producing both blaKPC-2 and blaIMP-96 could have serious implications for clinical treatment. We strongly recommend the urgent implementation of effective surveillance systems and stringent infection control measures to prevent potential outbreaks of such pathogens in healthcare settings across China. Based on a global genomic analysis of ST145 K. oxytoca isolates, we found that this sequence type exhibits an exceptionally high carriage rate of carbapenem resistance genes and has emerged as a high-risk epidemic clonal lineage with significant public health implications. Enhanced surveillance and screening measures should therefore be implemented in clinical settings to monitor the spread of this lineage.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Claesson MJ, Jeffery IB, Conde S, Power SE, O’Connor EM, Cusack S, Harris HMB, Coakley M, Lakshminarayanan B, O’Sullivan O, et al.. 2012. Gut microbiota composition correlates with diet and health in the elderly. Nature 488:178–184. doi:10.1038/nature 1131922797518 · doi ↗ · pubmed ↗
- 2Yang J, Long H, Hu Y, Feng Y, Mc Nally A, Zong Z. 2022. Klebsiella oxytoca complex: update on taxonomy, antimicrobial resistance, and virulence. Clin Microbiol Rev 35:e 0000621. doi:10.1128/CMR.00006-2134851134 PMC 8635272 · doi ↗ · pubmed ↗
- 3Disse SC, Meyer S, Baghai-Arassi A. 2018. Sepsis-associated purpura fulminans due to Klebsiella oxytoca. Dtsch Arztebl Int 115:784. doi:10.3238/arztebl.2018.0784 a PMC 632936830602411 · doi ↗ · pubmed ↗
- 4Suthers E, Rosenstengel A, Hart J, Lewis JR, Kay I, Waterer G, Lee YCG, Brims F. 2015. Pleural empyema caused by Klebsiella oxytoca: a case series. Respirology 20:507–509. doi:10.1111/resp.1247625677866 · doi ↗ · pubmed ↗
- 5Ghasemian A, Mohabati Mobarez A, Najar Peerayeh S, Talebi Bezmin Abadi A, Khodaparast S, Mahmood SS. 2019. Expression of adhesin genes and biofilm formation among Klebsiella oxytoca clinical isolates from patients with antibiotic-associated haemorrhagic colitis. J Med Microbiol 68:978–985. doi:10.1099/jmm.0.00096531136296 · doi ↗ · pubmed ↗
- 6Hareza DA, Bergman Y, Jacobs E, Lu J, Hanson ND, Conzemius R, Cosgrove SE, Harris AD, Simner PJ, Tamma PD. 2025. Molecular epidemiology of β-lactamases in ceftriaxone-resistant Enterobacterales bloodstream infections in the mid-Atlantic United States. Antimicrob Agents Chemother 69:e 0125824. doi:10.1128/aac.01258-2439868794 PMC 11881560 · doi ↗ · pubmed ↗
- 7Mohammedkheir MIA, Gaafar EM, Abd Alla EGE. 2024. Assessment of Bla TEM, Bla SHV, and Bla CTX-M genes of antibiotic resistance in Gram-negative bacilli causing urinary tract infections in Khartoum State: a cross-sectional study. BMC Infect Dis 24:141. doi:10.1186/s 12879-024-09023-738287256 PMC 10826001 · doi ↗ · pubmed ↗
- 8Lee KM, Morris-Love J, Cabral DJ, Belenky P, Opal SM, Jamieson AM. 2018. Coinfection with influenza A virus and Klebsiella oxytoca: an underrecognized impact on host resistance and tolerance to pulmonary infections. Front Immunol 9:2377. doi:10.3389/fimmu.2018.0237730420852 PMC 6217722 · doi ↗ · pubmed ↗