Impact of invasive weed Parthenium hysterophorus (Asteraceae) on mosquito abundance and plant-feeding behavior in an arboviral endemic region in Kenya
Tasneem Osman, Tatenda Chiuya, Eric M. Fèvre, Sandra Junglen, Christian Borgemeister

TL;DR
An invasive weed in Kenya boosts mosquito numbers and their plant-feeding behavior, increasing the risk of disease spread.
Contribution
Shows how an invasive plant affects mosquito populations and feeding behavior in a disease-prone region.
Findings
Mosquito abundance was significantly higher in Parthenium-infested villages, especially during the dry season.
Important vector species showed a clear preference for feeding on Parthenium hysterophorus.
Parthenium acts as a sustainable nutritional source, potentially increasing disease transmission risks.
Abstract
Invasive alien species (IAS) are rapidly altering ecosystems, undermining biodiversity, ecosystem processes, and interspecies interactions. Although IAS ecological and economic effects are well recognised, their impact on mosquito populations and the dynamics of infectious diseases is poorly understood. Plant-derived sugars are crucial for mosquito biology, supporting nectarivorous male survival and enhancing female blood feeding. In this study, we investigated how Parthenium hysterophorus, a rapidly proliferating invasive weed, shapes the population structure and nectar-feeding behaviour of the mosquito vector in the Rift Valley area of Kenya. Across six villages, three heavily infested with P. hysterophorus and three uninfested controls, we collected 48,489 mosquitoes representing 35 species from two subfamilies (Anophelinae and Culicinae) and nine genera, including Anopheles, Aedes,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
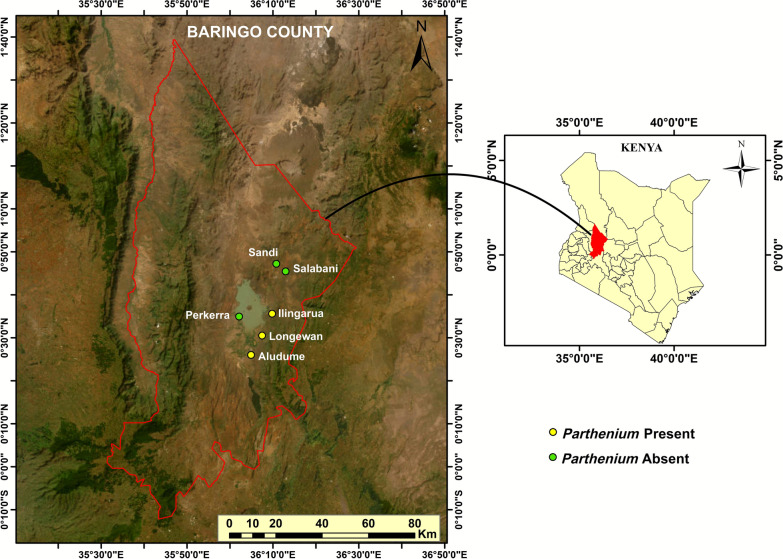 Figure 1
Figure 1 Figure 2
Figure 2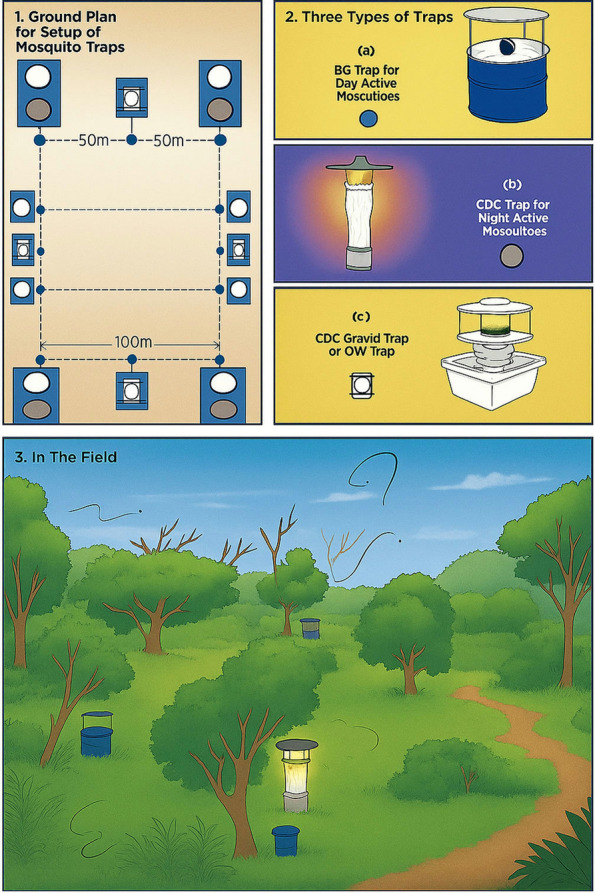 Figure 3
Figure 3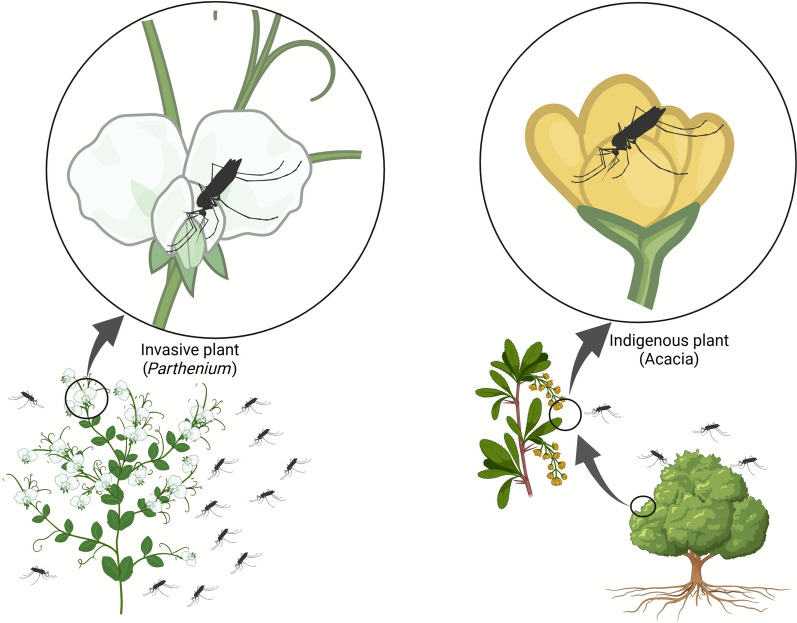 Figure 4
Figure 4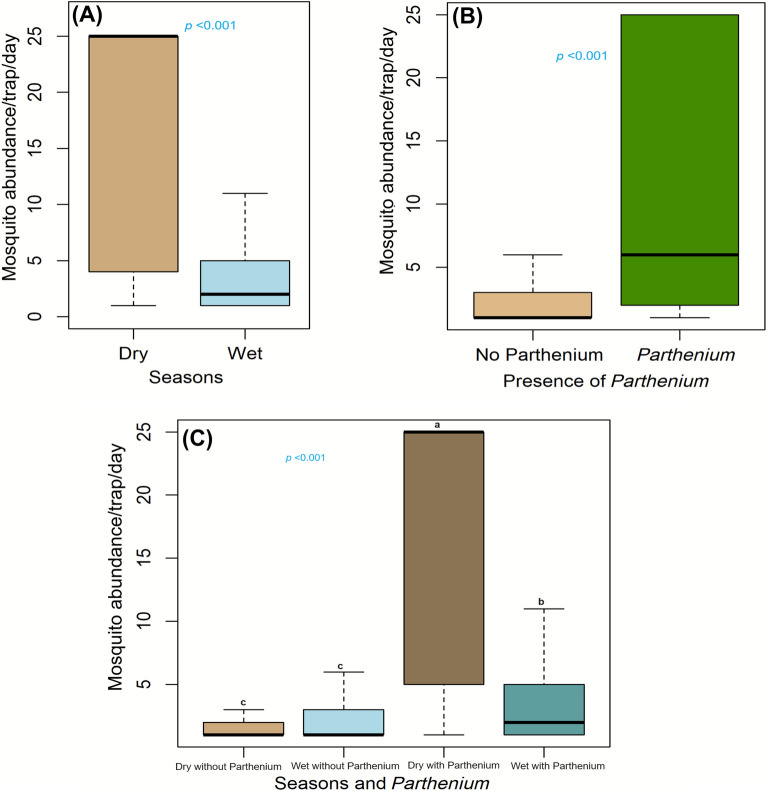 Figure 5
Figure 5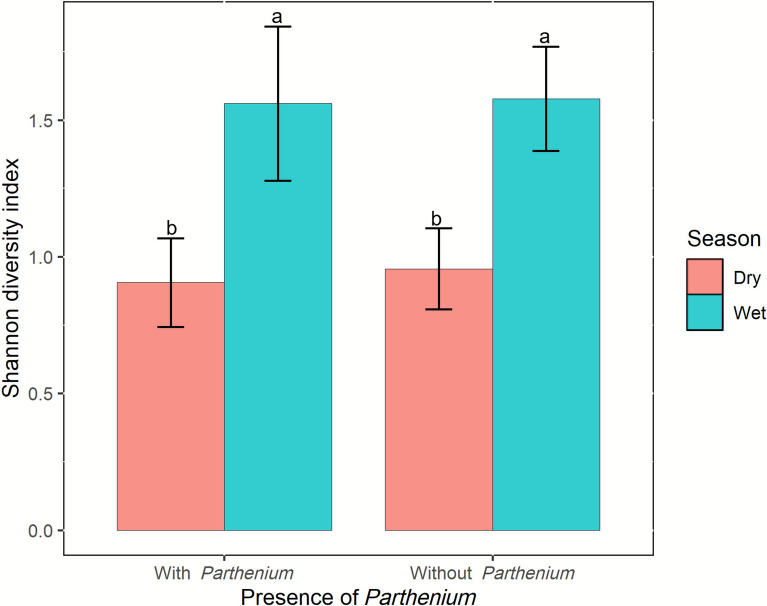 Figure 6
Figure 6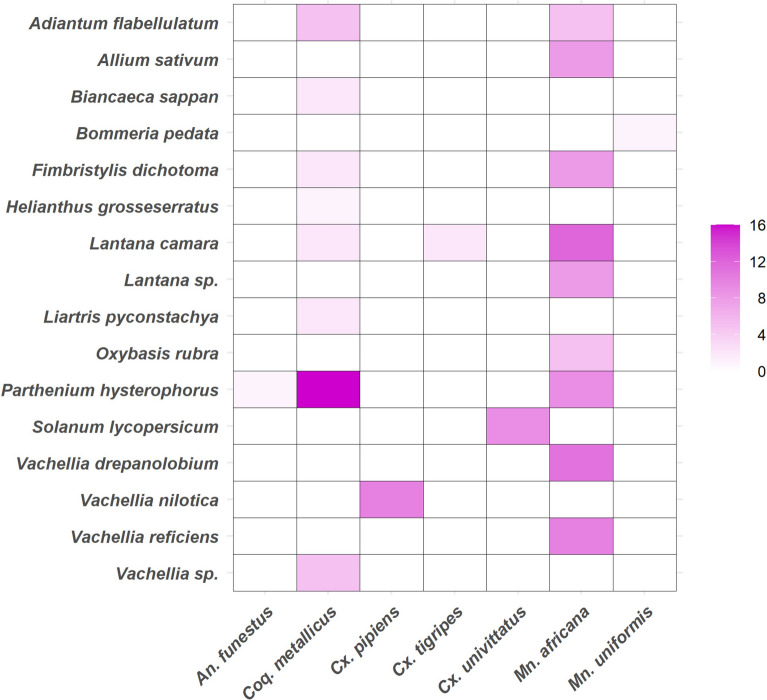 Figure 7
Figure 7 Figure 8
Figure 8- —https://doi.org/10.13039/501100001659Deutsche Forschungsgemeinschaft
- —https://doi.org/10.13039/100002322Schlumberger Foundation
- —Rheinische Friedrich-Wilhelms-Universität Bonn (1040)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAllelopathy and phytotoxic interactions · Insect Pest Control Strategies · Plant Parasitism and Resistance
Background
Invasive alien species (IAS) threaten the integrity of ecosystems throughout the world, affecting native species diversity and biological integrity [1]. Various factors, such as climate change, globalization, and reduced agricultural productivity, contribute to the spread and establishment of IAS [2]. With the increasing movement of people and goods around the world, fueled by new trade routes and enhanced transportation, the number of species accidentally introduced into new areas is rising constantly [3, 4]. Among the most aggressive and ecologically disruptive invasive species is Parthenium hysterophorus L. (Asteraceae). This rapidly expanding nonnative plant aggressively occupies regions outside its indigenous habitat in Central and South America, mainly Mexico and the Caribbean region, presenting significant risks to ecological diversity and farming practices, while also demonstrating harmful effects on both livestock and humans [5].
Mosquitoes depend on plant sugars as their primary source of energy. Males are exclusively herbivorous, while females enhance their blood meals with plant sugars to aid in flight, survival, and reproduction [6, 7]. The plant-feeding behavior of mosquitoes plays a crucial role in their biology, affecting their lifespan, reproductive success, and overall ability to transmit diseases. Nectar-rich plants can enhance mosquito populations, thereby facilitating the transmission of mosquito-borne diseases [8]. Mosquitoes demonstrate a distinct and selective preference for specific plant species while seeking sugar sources, and these preferred plants play a crucial role in mosquito survival by supplying essential sugar sources that boost energy reserves, extend lifespan, and improve reproductive success [9, 10].
Some IAS are highly attractive to mosquitoes as they tend to be more widespread and often have longer active flowering periods, thus guaranteeing a reliable supply of sugar meals and thereby enhancing the mosquito lifespan [11]. A particularly important example is the invasive neotropical weed P. hysterophorus, which has become one of Africa’s greatest threats to biodiversity [12]. In addition, feeding on P. hysterophorus negatively affects animal health [13], and its pollen and allelopathic substances can cause respiratory problems and skin allergies in humans [13, 14]. Parthenium hysterophorus deteriorates soil quality, exacerbates competition for water resources, and may facilitate the dissemination of vector-borne diseases via habitat modification [15], resulting in major ecological and public health issues in impacted areas [16, 17].
Previous studies have shown that female Anopheles gambiae Giles (Diptera: Culicidae), the most important malaria vector in sub-Saharan Africa (SSA), is highly attracted to P. hysterophorus plants [10]. Feeding on this plant improves its fitness compared with feeding on other, noninvasive plants [15, 18], enhancing its ability to transmit malaria [15, 19]. Moreover, P. hysterophorus can also influence the oviposition behavior of mosquito females, as its plant litter can alter the microhabitat conditions, making them more suitable for breeding [18, 20, 21]. Overall, these factors can potentially lead to an increase in mosquito populations in areas infested with Parthenium, subsequently affecting the transmission dynamics of mosquito-borne diseases.
Previous investigations on plant feeding have concentrated mainly on Anopheles gambiae concerning malaria transmission. Nonetheless, there remains a significant gap in our understanding of how plant feeding, particularly the impact of IAS such as P. hysterophorus, affects the transmission dynamics and ecological interactions of mosquito vectors linked to arboviral diseases. These mosquitoes, including species of the genera Aedes, Culex, and Mansonia, play a significant role in the spread of viruses such as dengue, Zika, chikungunya, Rift Valley, and yellow fever viruses [18]. Considering that elevated populations of mosquitoes are typically linked to a rise in pathogen spread, such as arboviruses [22], it is essential to comprehend the relationships between floral resources and these disease-carrying insects. We therefore investigated how floral diversity in Baringo County, Kenya, an arboviral endemic region, affects mosquito proliferation and their plant-feeding behavior, with a particular focus on P. hysterophorus and arboviral mosquito vectors.
Methods
Study site
The study was conducted in the Kenyan Rift Valley, specifically in Baringo County (Fig. 1), which lies between latitudes 0° 13′ S and 1° 40′ N and longitudes 35° 36′ and 36° 30′ E. The county is crossed by the Equator. The area experiences annual temperatures ranging between 21 and 32 °C, with precipitation between 300 mm and 1500 mm distributed over two rainy seasons, i.e., March/April to May/June, termed the long rains, and October to early December, termed the short rains [23]. The vegetation is diverse, with key zones that include Acacia–Commiphora bushlands in low-lying semi-arid areas, dominated by Vachellia (Acacia) tortilis (Forssk) Galasso & Banfi (Fabaceae) and Commiphora africana (A. Rich) Endl. (Burseraceae). Baringo’s dry montane forests, found in the Tugen Hills, host species such as Juniperus procera Hochst ex Endl. (Cupressaceae) [24], while the riverine vegetation along rivers such as the Perkerra includes Ficus sycomorus L. (Moraceae) and V. (Acacia) xanthophloea (Benth.) P. J. H. Hurter [25]. Grasslands and shrublands in the lowlands of the county support grasses such as Chloris gayana Kunth and Cynodon dactylon (L.) Pers. (both Poaceae) [25]. IAS such as Neltuma (Prosopis) juliflora (Sw.) (Fabaceae) [26], and P. hysterophorus L. (Asteraceae) have spread in areas such as Marigat, displacing native plants and causing ecological and economic damage [27]. Parts of the county are prone to frequent flooding associated with outbreaks of Rift Valley fever virus, which the area has experienced in the recent past [28–30]. The villages selected for this study included Aludume, Ilngarua, and Longewan, which are infested with P. hysterophorus, alongside Perkerra, Salabani, and Sandai, which are generally P. hysterophorus free (Fig. 1).Fig. 1. Map of Baringo County, Kenya, showing study sites, which include villages with a known presence of (Aludume, Ilingarua, and Longewan) and those without (Perkerra, Salabani, and Sandai). Map generated in ArcGIS Pro version 3.0 (ESRI, Redlands, CA, USA)
Study design and sampling
Mosquito sampling was conducted in six villages within Baringo County, Kenya, to assess the influence of P. hysterophorus infestation on mosquito abundance and diversity. To enable comparison, the study design comprised three heavily *P. hysterophorus-*infested locations and three ecologically comparable, control sites that were not infested to facilitate robust comparisons. Sampling occurred from October 2019 to March 2020, encompassing both dry and wet seasons to address seasonal variations in mosquito populations. Each site was sampled once during each season, with a duration of five consecutive days per site. This approach ensured that we caught seasonal fluctuations while collecting data consistently across all sites. At each site, three different types of traps were used to capture adult mosquitoes: Biogents (BG)-Sentinel traps, Centers for Disease Control and Prevention (CDC) small light traps, and CDC gravid traps (Figs. 2 and 3). Twenty trap placements were created, including eight BG-Sentinel traps, eight CDC small light traps, and four CDC gravid traps. All traps were positioned close to vegetation and zones of human activities. BG-Sentinel targeted day-active species, such as Aedes aegypti, while CDC small light traps, situated at a height of 1.5 m above the ground, were utilized to catch night-active mosquitoes, including Anopheles and Culex spp. CDC gravid traps, located near water sources, were used to attract gravid mosquitoes.Fig. 2. Field setup and sampling activities during mosquito collection in Baringo County, Kenya. A The invasive weed Parthenium hysterophorus in bloom. B P. hysterophorus-infested agricultural field, showing typical moist furrows that provide suitable mosquito breeding habitats. C BG-Sentinel trap placed near vegetation for capturing day-active mosquitoes. D CDC light trap positioned among P. hysterophorus vegetation to target nocturnal mosquito species. E Field technician inspecting a CDC gravid trap during evening collectionsFig. 3Strategic layout of BG-Sentinel traps, CDC small light traps, and CDC gravid traps in the field to capture mosquitoes of different species and behaviors.
The traps were supplied with dry ice to release carbon dioxide (CO_2_), acting as a mosquito attractant [31]. To minimize the overlap in mosquito captures, the BG-Sentinel and CDC small light traps were situated 10 m apart at each location. Trap locations were additionally separated by 100 m to reduce the potential interaction between collections.
Moreover, CDC gravid traps were carefully positioned approximately halfway between the BG-Sentinel and CDC small light traps to enhance the probability of capturing gravid mosquitoes. Trapping was performed over five consecutive days at each site. BG-Sentinel traps functioned during daytime hours (06:00–17:00), whereas CDC small light traps and CDC gravid traps were utilized overnight (18:00–06:00). This method facilitated a thorough evaluation of mosquito populations, encompassing individuals with diverse activity patterns and reproductive status. The detailed trapping schedule was the following: 8 BG-Sentinel traps × 5 days = 40 trap-days, 8 CDC mall light traps × 5 days = 40 trap-days, and 4 CDC gravid traps × 5 days = 20 trap-days. Thus, in total, we had 100 trap-days per location.
Mosquito sample preparation and processing
The samples were delivered to a temporary field laboratory at the Kenya Forestry Research Institute (KEFRI) in Baringo County for processing. Non-mosquito specimens were discarded, while the mosquitoes were initially preserved in liquid nitrogen and then transported in cryovials to the Kenya Medical Research Institute (KEMRI) in Nairobi for morphological identification using taxonomic keys [32, 33]. Mosquitoes were grouped into pools containing no more than 25 individuals according to site, species, sex, trap type, and fed status. Following identification, the mosquitoes were transferred to ILRI, where they were stored at −80 °C for long-term preservation.
Plant feeding assay
Anthrone sugar test
The anthrone test is a biochemical assay to detect the presence of sugar (fructose) [34] and can be an indicator of recent sugar or plant feeding in mosquitoes. Before the test, each mosquito was rinsed in phosphate-buffered saline (PBS) to remove plant residues from their exoskeletons before further analysis. The mosquito’s abdomen was then dissected with a scalpel and macerated in 50 μL absolute ethanol using a sterilized pestle. An aliquot of 25 μL of each homogenate was transferred to the wells of a flat-bottomed 96-well microplate for analysis. The assay is based on the original procedure by Handel and Day [35], with modifications by Matheson et al. [36] to improve the accuracy and sensitivity of detecting fructose. Briefly, 200 μL of anthrone solution (0.15% anthrone (Sigma-Aldrich GmbH, Germany) w/v in 71.7% sulfuric acid) was added to each well containing homogenate. The plate was incubated at room temperature (25 °C) for 60 min. A positive result was indicated by a color change from yellow to blue, indicating the presence of fructose in the sample.
DNA extraction
For plant DNA extraction from fructose-positive mosquito samples, the insect homogenate was further homogenized using ceramic beads and a tissue lyser. DNA was then extracted using the ISOLATE II Plant DNA Kit (QIAGEN, Germany) according to the manufacturer’s instructions with modifications. These included the extension of the incubation period with the lysis buffer PA1 and RNase by 4–6 h and elution with Buffer PG for 10 min. The extracted DNA was further analyzed on 1% agarose gel under ultraviolet (UV) light and subsequently quantified by a spectrophotometer NanoDrop™ 2000 (ThermoScientific, USA) and the fluorometer Qubit^®^ 2.0 (Invitrogen, USA). DNA was stored at −20 °C until further analysis, which included polymerase chain reaction (PCR) screening and sequencing.
PCR screening and sequencing
PCR was conducted to identify the plant species upon which the mosquitoes had presumably fed, for each pool that tested positive for fructose. Three different sets of primers, i.e., MatK (MatK-413f-1 and MatK-1227r-1), rbcL (rbcL-1F and rbcL-724R), and ITS (ITS1 and ITS4), were used to amplify both conserved rbcL and polymorphic MatK genes of the chloroplast and the 5.8S ITS sequence as described by Kress et al. [37]. These markers are widely recognized in plant barcoding studies [38]. The PCR used MyTaq Polymerase DNA polymerase kit (Bioline, London, UK) in a reaction volume of 10 μL, which contained 10 μM of each forward and reverse primer, 0.0625 U MyTaq DNA polymerase, 5× MyTaq reaction buffer, and 1–2 μL of DNA template (1–10 ng). The reactions were performed in a thermal cycler (Eppendorf Mastercycler Nexus, Germany), following a thermal profile that included an initial denaturation at 94°C for 4 min, followed by 35 cycles of 94°C for 30 s, 52–55 °C for 30 s, and 72 °C for 1 min, concluding with a final extension at 72 °C for 10 min [39]. Molecular-grade water was included as a negative control in all PCRs, while DNA extracted directly from the tissue of selected plant species located within a 100-m radius of each trap site served as a positive control. The PCR products were resolved by 1.5% agarose gel electrophoresis, stained with SYBR Green (Sigma-Aldrich GmbH, Germany), and compared against a 100-bp DNA ladder (HyperLadder, Bioline, London, UK). PCR products were purified and then Sanger sequenced to generate both forward and reverse reads at the BecA-ILRI (The Biosciences eastern and central Africa-International Livestock Research Institute) Segoli laboratory in Nairobi, Kenya. They were visually inspected, aligned, and edited using Geneious Prime version 2023.1 [40]. Each sequence was compared with a reference sequence in the GenBank database using BLASTn (www.ncbi.nlm.nih.gov) and the Barcode of Life Database (BOLD) system database (www.boldsystem.org). Sequences were assigned to specific plant species based on a match threshold of >98%.
Statistical analysis
Total mosquito species-specific abundance was analyzed using general linear models (GLMs), considering the impact of seasons (dry and rainy) and the presence of P. hysterophorus as contributing factors. The Shannon diversity index was calculated to examine mosquito diversity, with analyses conducted using the “vegan” package in R [41]. In this regard, the Shapiro–Wilk test was performed to assess the normality of the data distribution. The extent of plant feeding (fructose-positive samples) was computed as a percentage of the total number of mosquitoes tested. A chi-squared test of independence was utilized to compare the fructose positivity rates between male and female mosquitoes, where P. hysterophorus was present or absent and across seasons. A pairwise comparison of proportions was conducted using the pairwise nominal independence test. All statistical tests were performed using R software, with a significance level of P < 0.05.
Results
Abundance and distribution of mosquitoes
During the sampling period from October 2019 (rainy season) to February 2020 (dry season), a total of 48,489 mosquitoes were captured: 37,181 in the rainy season (October 2019) and 11,272 in the dry season (February 2020) (Table 1). The collected mosquitoes were classified into two subfamilies: Anophelinae, represented by the Anopheles genus, and Culicinae, which included the eight genera: Aedomyia, Aedes, Coquillettidia, Culex, Ficalbia, Mansonia, and Uranotaenia. In total, 35 mosquito species were morphologically identified. Mansonia africana was the predominant species collected (n = 31,692/53,258; 59.5%) followed by Coq. metallicus (n = 4906/53,258; 9.2%) and Cx. pipiens (n = 4199/53,258; 7.9%) in decreasing order (Table 1). Table 1. Distribution and abundance of mosquito species collected across six locations in Baringo County, KenyaSpecies namePresence of PartheniumAbsence of PartheniumTotalIlngaruaLongewanAludumePerkerraSalabaniSandaiAe. aegypti^^5210014465Ae. furcifer*^^0011001Ae. hirsutus*^^2225865146243Ae. mcintoshi*^^37794662014271Ae. metallicus0000101Ae. simpsoni0000303Ae. sudanensis0001001Ae. tarsalis0110002Ae. tricholabis321921010181Aedes* spp.^^04323215Aedomyia furfurea1033105An. coustani2051171223139An. funestus*^^16145463286An. gambiae*^^229261992391353An. malculipalpis0000101An. nili*^^0010066An. pharoensis*^^1100014An. squamosus0000000Anopheles* spp.^^6016212736Coq. aurites2103633604282466Coq. fuscopennatus601112398025226Coq. mettallicus*^^173466442734110057355143Coquillettidia* spp.^^0101003Cx. annulioris1224935864123433Cx. bitaeniorhynchus15320114Cx. cinereus*^^40542212Cx. ethiopicus141211504083Cx. pipiens ^^3323487019046418503846Cx. poicilipes37380833Cx. tarsalis^^0200004Cx. tigripes201611016868144Cx. univittatus ^^288127573273867961697Cx. vansomereni^^20039014Cx. zombaensis3001004Culex* spp^^6013403527122267Ficalbia mediolineata47730122F. splendens44350219Mansonia africana*^^23,8305353239170139717236,806Mn. uniformis*^^13763608432888972,609Mansonia* spp.^^695077295Uranotaenia* spp.0120002Total28,2787222245341941889445348,489^*^Medically important
Effect of seasonal variation and Parthenium on mosquito abundance
The GLM analysis with Poisson distribution revealed a significant seasonal effect, with mosquito abundance being significantly higher during the dry compared with the wet season (P < 0.001) (Fig. 5A). The estimated marginal means indicated that the dry season provided more favorable conditions for mosquito proliferation in the study area. The effect of the presence of P. hysterophorus on mosquito abundance was also evaluated using a GLM with Poisson distribution (Figs. 4, 5B). The model showed that sites infested with P. hysterophorus had significantly higher mosquito abundance than non-infested sites (odds ratio (OR) = 1.8031, P < 0.001), suggesting that P. hysterophorus creates an environment conducive to higher mosquito densities. Further analysis revealed that the abundance of medically important mosquito species was significantly higher in areas with P. hysterophorus (P < 0.001). This includes key vectors, particularly An. gambiae, An. funestus, Culex pipiens, Culex univittatus, and Coquillettidia metallicus.Fig. 4. Differential mosquito visits to invasive and native plant species. The graphic illustrates mosquito abundance in relation to the invasive species Parthenium hysterophorus, with the native plant Acacia spp. An elevated quantity of mosquitoes is recorded engaging with P. hysterophorus, indicating its possible function in augmenting mosquito longevity via enhanced sugar availability, with ramifications for vector ecology and disease transmission.Source: authors with BioRender.com (www.biorender.com)Fig. 5. Box plots illustrating the daily mosquito abundance per trap in various environments, during dry and wet seasons (A), in the presence and absence of Parthenium (B), and accounting for both factors (season and Parthenium) (C). Asterisks (*) and different letters denote significant differences at P < 0.001
The combined effect of season and presence of Parthenium
To investigate the interactive effect of season and presence of P. hysterophorus on mosquito abundance, a GLM was fit with an interaction term between season and P. hysterophorus presence (Fig. 5C), revealing a significant interaction effect (P < 0.001), with the highest mosquito abundance observed during the dry season in P. hysterophorus-infested areas. This abundance was significantly higher than in any other combination of season and P. hysterophorus. Conversely, the lowest mosquito abundance was recorded during the rainy season in areas without P. hysterophorus. Pairwise comparisons, adjusted using Tukey's method, revealed distinct groupings a, b, and c (Fig. 5), indicating significant differences in mosquito abundance between the interaction groups.
Mosquito species diversity
In sites free of P. hysterophorus, the average Shannon diversity index was higher during the wet (mean = 1.58) compared with the dry season (mean = 0.96) (t = −4.89, P = 0.021). Similarly, in sites with P. hysterophorus, species diversity was also higher during the wet (mean = 1.56) than the dry season (mean = 0.91) (t-value of −2.55, P-value = 0.012) (Fig. 6).Fig. 6. Mean Shannon diversity index (± standard error (SE)) of mosquito populations during the dry and rainy seasons, comparing villages with and without P. hysterophorus presence. Different letters indicate significant differences between the seasons in villages with and without P. hysterophorus (t-test, P < 0.05)
Fructose positivity rates and dynamics of sex ratio
Female mosquitoes had significantly higher fructose positivity rates in areas with P. hysterophorus (28.13%; n = 299/1,063) compared with areas without P. hysterophorus (18%; n = 38/209) (χ^2^ = 8.3689, P = 0.0038), indicating a correlation between the presence of P. hysterophorus and increased herbivorous activity in females. The fructose positivity rates for male mosquitoes showed no significant difference between P. hysterophorus-infested areas (23.07%; n = 27/117) and non-infested areas (15%; n = 17/110) (χ^2^ = 1.5676, P = 0.2106). The chi-squared test revealed no significant difference in overall fructose positivity across sexes in P. hysterophorus-infested areas (χ^2^ = 1.2122, P = 0.2709). Similarly, there was no significant difference between females and males in areas lacking P. hysterophorus (χ^2^ = 0.20886, P = 0.6477). This indicates that the impact of P. hysterophorus on fructose positivity was significant solely for female mosquitoes. A thorough chi-squared analysis of all mosquitoes (combined sexes) between P. hysterophorus-infested and non-infested regions demonstrated a significant correlation, indicating heightened fructose positivity in P. hysterophorus-infested areas (χ^2^ = 3.980, P = 0.046).
Diversity of plants identified in sugar-fed mosquitoes
A total of 476 fructose-positive mosquito samples underwent PCR analysis, providing a 19.7% (94/476) amplification success rate. In total, 47 samples were successfully sequenced from 12 males and 35 females of various mosquito species. The sequenced fragments varied in length from 450 to 660 base pairs. Analysis of the 47 sequences revealed 16 different plant species as potential sources of plant sugars (Fig. 7). The heatmap analysis indicated specific feeding preferences among mosquito species for different plant species. Mansonia africana had the most extensive spectrum of plant consumption, with high counts recorded for Lantana camara L. (Verbenaceae) (n = 12), V. drepanolobium (Harms ex. Sjöstedt) P.J.H. Hurter (n = 11), and Vachellia reficiens (Wawra) Kyal. & Boatwr. (n = 10). Importantly, P. hysterophorus was recognized as a key plant source for Mn. africana (n = 9). Other plant species found in Mn. africana were Fimbristylis dichotoma (L.) Vahl (Cyperaceae) (n = 8), Allium sativum (n = 8), and Lantana sp. (n = 8), while Oxybasis rubra (L.) S. Fuentes, Uotila & Borsch (Amaranthaceae) and Adiantum flabellulatum (L.) (Pteridaceae) were consumed by n = 5 each. In Coq. metallicus, notable numbers tested positive for P. hysterophorus (n = 16), A. flabellulatum (n = 5), Vachellia sp. (n = 5), L. pycnostachya, Biancaeca sappan (L. 1753) Tod. 1875 (Fabaceae), F. dichotoma, and L. camara, all of which were consumed by two individuals (n = 2) each, while Helianthus grosseserratus M. Martens (Asteraceae) (n = 1) was also detected in the same species. Anopheles funestus tested positive only for P. hysterophorus (n = 1). In Cx. pipiens, V. nilotica was detected (n = 10), while in Cx. tigripes, L. camara was found (n = 2). In Cx. univittatus, S. lycopersicum (n = 9) was found. Mn. uniformis tested positive only for Bommeria pedata Sw. (Pteridaceae) (n = 1) (Fig. 7).Fig. 7. Heatmap displaying plant–host relationships among seven mosquito species from which DNA sequences of fructose-positive samples were successfully amplified. The color intensity in each cell signifies the frequency of identified plant DNA in the corresponding mosquito species, with darker hues representing stronger feeding connections
Discussion
The impact of invasive plant species on mosquito diversity and abundance, as well as on the potential consequences for transmission of vector-borne diseases, has gained increasing attention [8, 42]. We trapped considerably higher numbers of Mn. africana, Coq. metallicus, and Cx. pipiens during the dry season in villages where P. hysterophorus was present compared with Parthenium-free sites, suggesting a potential positive ecological association between the mosquitoes and the IAS. The high abundance of Mansonia spp. is likely driven by the favorable larval habitat provided by wetland vegetation, as these mosquitoes attach to aquatic plants for respiration through modified syphons. Furthermore, M. africana exhibited a significant preference for P. hysterophorus nectar and elevated fructose positivity, suggesting that the proliferation of the species in infested areas is significantly influenced by the availability of floral sugar and the suitability of the larval habitat [43, 44]. However, additional factors certainly contributed to the elevated mosquito populations. These may include humans and livestock as suppliers of blood and water storage containers as breeding locations. Mosquitoes may also receive sugar from other flowering plants [10]. The epidemiological importance of these mosquito species is highlighted by their roles as vectors for several arboviruses. Mansonia, Coquillettidia, and Culex spp. transmit, among others, the Rift Valley fever virus (RVFV), which can affect both humans and livestock [45, 46]. Additionally, Cx. pipiens is a key vector for the West Nile virus, lymphatic filariasis, Rift Valley fever virus, and encephalitic viruses, which have serious implications for human health [47, 48]. The high prevalence of these vector species in Baringo County emphasizes their potential influence on public health, warranting continuous surveillance and vector control measures. Plant sugar assays and DNA barcoding also showed that most of the mosquitoes were feeding on P. hysterophorus and L. camara, suggesting that these plants potentially serve as a crucial source of sugar for mosquitoes.
Our findings indicate a complex and species-specific relationship between the presence of P. hysterophorus and the abundance of mosquitoes. Interestingly, An. gambiae a key malaria vector, was found to be more prevalent in sites devoid of Parthenium (Table 1), contradicting previous studies that identified P. hysterophorus as an essential sugar source boosting the survival and activity of An. gambiae [10, 49]. The prior findings, mainly derived from controlled experiments, indicated a significant preference for P. hysterophorus over other plant species such as Senna occidentalis (L..) Link (Fabaceae) [10, 50, 51]. Conversely, our field data reveal that, in natural settings, An. gambiae populations might be more affected by various ecological factors, including host availability, microhabitat traits, or alternative sugar sources. Our research highlights that field conditions may influence mosquito–plant interactions differently from laboratory settings.
Other IAS have also been found to enhance the abundance of mosquitoes; for example, in Mali, a drastic drop in Anopheles spp. populations and sugar feeding was observed in areas where the flowering parts of Neltuma (Prosopis) juliflora (Sw.) Raf. (Fabaceae) were removed compared with areas with intact stalks. Notably, most of these studies focused on Anopheles mosquitoes; hence, our study is crucial in providing information on plant foraging in arboviral mosquito vectors such as Mansonia and Culex spp. In this regard, Aedes aegypti mosquitoes were found to be frequently feeding on Fabaceae plants in the coastal town of Kilifi; however, some fed on P. hysterophorus [52]. Typically, mosquito abundance is higher during the rainy season because of the greater availability of breeding habitats and favorable humidity and temperature [53, 54]. Yet, we found the exact opposite in areas where Parthenium was present, with significantly higher mosquito trap catches during the dry than the wet season, something that we did not observe in areas where the IAS was absent. One of the contributing factors could have been that farmers in Baringo County do not control weeds during the dry season, thereby allowing P. hysterophorus in affected areas to flourish. In contrast, during the rainy (growing) season, fields are cleared for crop cultivation, resulting in the removal of Parthenium. Hence, the abundance of P. hysterophorus during the dry season in affected areas is likely to provide favorable microhabitats and sugar-rich food sources for mosquitoes, thereby enabling their survival and reproduction even when traditional breeding sites are in short supply [27, 42, 55]. Parthenium hysterophorus thus helps to sustain mosquito populations during periods of low to no precipitation, highlighting its potential to affect mosquito dynamics at the ecological level.
In our study, we found P. hysterophorus to be considerably more preferred by mosquitoes than other invasive plant species such as L. camara and also Vachellia reficiens (Wawra) Kyal. & Boatwr. (Fabaceae). Mosquitoes demonstrate selective plant-feeding preferences, and while studies show that P. hysterophorus has poor nutritional content, it is likely to attract mosquitoes through release of volatile compounds [15, 17, 56]. It is also likely to outcompete native flora, especially during the dry season, resulting in a reduction in the availability of native nectar sources and compelling mosquitoes to adapt their feeding habits [57, 58]. Mosquitoes that frequently fed on P. hysterophorus included Coq. metallicus, Mn. africana, and An. funestus. Mansonia spp. are recognized vectors of RVFV and lymphatic filariasis [59] (and in the last major RVF outbreak in Kenya, the virus was frequently detected in Mn. africana and Mn. uniformis mosquitoes in Baringo County [30]. Anopheles funestus is one of the principal malaria vectors in SSA [60]. Their high trap catches in areas infested with P. hysterophorus suggest that the IAS may elevate the risk of arboviral disease transmission.
We found a significant correlation between the presence of P. hysterophorus and elevated fructose positivity in female mosquitoes, suggesting that the IAS acts as a crucial sugar source. Nyasembe et al. [15] observed that P. hysterophorus enhances An. gambiae longevity by providing ample sugar sources, and P. hysterophorus was favored above other invasive species such as L. camara, indicating its distinct appeal. While the presence of IAS such as P. hysterophorus can potentially increase disease risk by increasing mosquito abundance, other studies have shown that sugar intake prior to blood feeding may protect Ae. aegypti females against infection with Semiliki Forest and Zika viruses [61]. On the basis of previous studies, this effect can also be extrapolated to Anopheles and Culex spp. mosquitoes [62, 63] The protective effect is thought to be mediated through increased melanization and stimulation of the mosquito immune system [61]. This suggests that plant sugar in mosquitoes could reduce their capacity to harbor and transmit viruses. Further field and in vitro experiments are therefore required to disentangle these contradictory dynamics in plant feeding and arboviral transmission.
Conclusions
Our study indicates a distinct association between the presence of P. hysterophorus and increased mosquito populations, particularly under unfavorable dry conditions, stressing the crucial role of IAS-like P. hysterophorus as both a sugar source and possible breeding microhabitat for mosquitoes, enabling vector persistence during adverse environmental conditions. Furthermore, temperature, humidity, and land-use patterns can shape mosquito populations by driving resource availability and the suitability of breeding sites. DNA barcoding and sugar analyses identified P. hysterophorus as a significant sugar source for various medically important mosquito species, including Mn. africana and Cx. pipiens. Importantly, mosquito diversity remained consistent throughout the year in P. hysterophorus-infested areas, in contrast to control areas that displayed seasonal variations. These findings indicate that the effective management of P. hysterophorus could function as a strategic approach for controlling mosquito proliferation, thereby reducing the risk and spread of vector-borne diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hollis L. Weeding invasive alien species—Africa’s economic burden. 2021 [Internet]. [cited 2025 Jul 1]. https://blog.invasive-species.org/2021/05/25/weeding-invasive-alien-species-africas-economic-burden/
- 2Almire F, Terry S, Mc Farlane M, Sziemel AM, Terhzaz S, Varjak M, et al. Sugar feeding enhances gut immunity and protects against arboviral infection in the mosquito vector Aedes aegypti. 2021 [Preprint]. 10.1101/2021.01.05.42537510.1371/journal.ppat.1009870 PMC 841234234473801 · doi ↗ · pubmed ↗