Genetic Diversity of Polymyxin Resistance Genes in Klebsiella pneumoniae Clinical Isolates
Daniel Miceli Serwy, Maria Eduarda Rocha Conde, Ana Luiza Carneiro Alencar, Roberto Leonan Morim Novaes, Josué da Costa Lima‐Junior, Fabio Faria da Mota, Ana Paula Carvalho‐Assef, Teca Calcagno Galvao, Viviane Zahner

TL;DR
This study explores how Klebsiella pneumoniae develops resistance to polymyxin antibiotics through genetic changes in specific genes.
Contribution
The study reveals that polymyxin resistance in Klebsiella pneumoniae is driven by polygenic adaptation and positive selection, with mgrB insertions playing a key role.
Findings
High genetic diversity and positive selection were observed in polymyxin-resistant Klebsiella pneumoniae isolates.
mgrB insertions dominate resistant populations and are linked to rapid haplotype spread via horizontal gene transfer.
Abstract
This study investigates the genetic diversity and evolutionary mechanisms driving polymyxin resistance in Klebsiella pneumoniae , a critical priority pathogen. By analysing mgrB, phoPQ and pmrAB genes in susceptible (PM‐S) and resistant (PM‐R) populations through neutrality tests (Tajima D, Fu & Li's D) we uncovered polygenic adaptation and positive selection as a key driver of resistance. High genetic diversity was observed across all loci, with mgrB insertions dominating PM‐R populations. Negative Tajima and Fu & Li's D values and excess rare alleles revealed recent population expansions linked to the reintroduction of polymyxins in the 2010s. Positive selection via selective sweeps was detected in PM‐R isolates, exemplified by the rapid spread of haplotype 27, which presents mgrB insertions, the major determinant of LPS modification pathway hyperactivation. The expansion of this…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
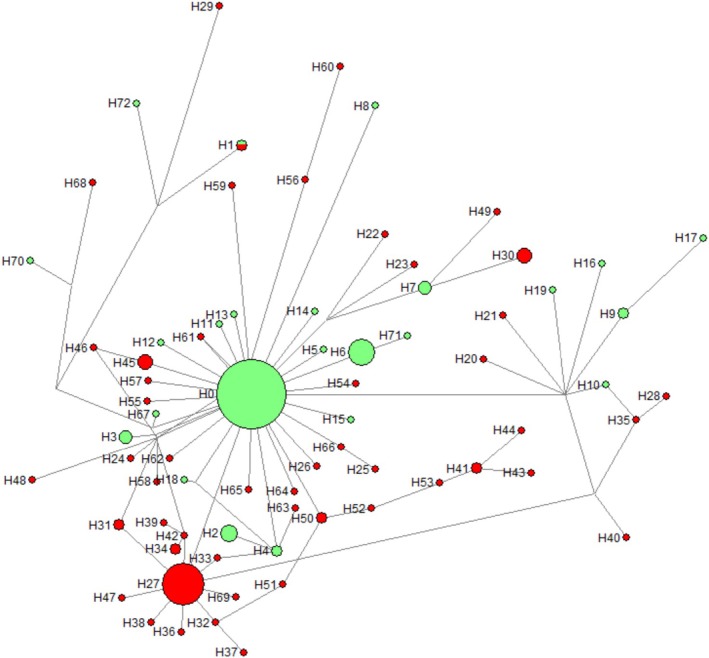 FIGURE 1
FIGURE 1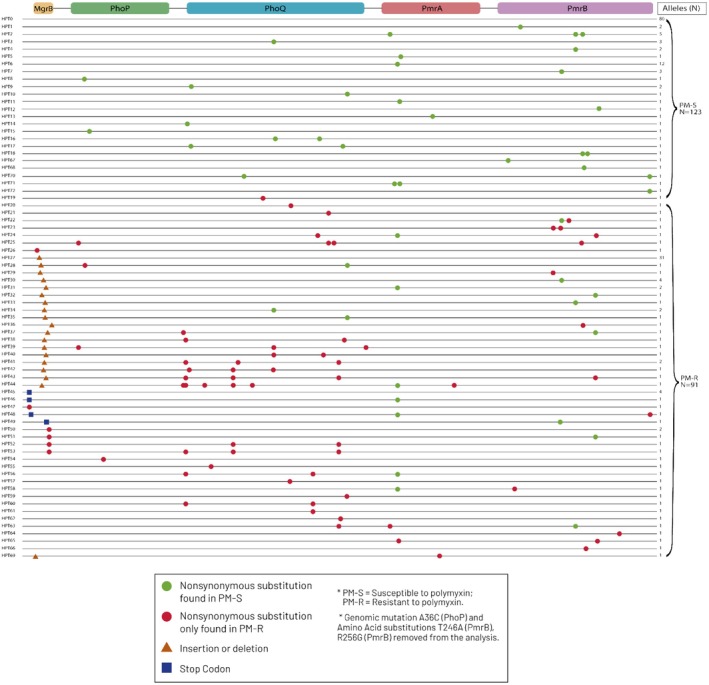 FIGURE 2
FIGURE 2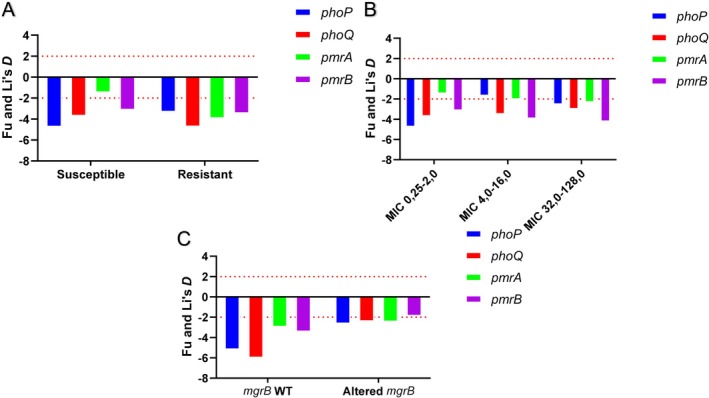 FIGURE 3
FIGURE 3| Strain | MIC | Phenotype | Susceptibility test | Source and reference | Country | Polymyxin | Year of collection |
|---|---|---|---|---|---|---|---|
| 129 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 153 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 154 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 155 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 201 | 8 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 219 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 249 | 0.5 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 257 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 417 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 446 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 7559 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| 7585 | 1 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| CCBH22128 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22137 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22143 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22206 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22237 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22240 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22382 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22391 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22397 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22399 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22404 | 4 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22408 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22462 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22466 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22481 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22491 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22609 | 64 | Resistant | Broth microdilution (Brcast) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22625 | 64 | Resistant | Broth microdilution (Brcast) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22653 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22675 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22740 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22997 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH22999 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23000 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23001 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23024 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23031 | 64 | Resistant | Broth microdilution (Brcast) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23043 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23048 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23050 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23064 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23097 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23167 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23171 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23220 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23247 | 32 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23296 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23323 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23368 | 4 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23454 | 16 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23615 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23650 | 64 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23661 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23663 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH23741 | 128 | Resistant | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH33272 | 2 | Susceptible | Broth microdilution (Brcast) | Sanger (This Work) | Brazil | B | 2023 |
| CCBH72480 | 2 | Susceptible | Broth microdilution (BrVCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH73566 | 2 | Susceptible | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| CCBH73965 | 2 | Susceptible | Broth microdilution (BrCAST) | Sanger (Conceição‐Neto et al. | Brazil | B | 2016 |
| 12_BR_13 | 64 | Resistant | Broth microdilution (CLSI) | SAMN04868732 (Pitt et al. | Brazil | B | 2013 |
| 7610 I | 1 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| CBAS 537 | 0.5 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| CBAS 540 | 2 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| CBAS 541 | 1 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| JC 22 | 1 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| JC 23 | 2 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| JC 31 | 1 | Susceptible | Broth microdilution (BrCAST) | Sanger (This Work) | Brazil | B | 2023 |
| Kp37 | 2 | Susceptible | Broth microdilution (BrCAST) | SAMEA4773886 | Brazil | B | 2016 |
| kp71 | 2 | Susceptible | Broth microdilution (BrCAST) | Sanger (Lourenço | Brazil | B | 2016 |
| 10_GR_13 | 64 | Resistant | Broth microdilution (CLSI) | SAMN04868730 (Pitt et al. | Greece | B | 2013 |
| 13_GR_14 | 16 | Resistant | Broth microdilution (CLSI) | SAMN04868733 (Pitt et al. | Greece | B | 2014 |
| 18_GR_14 | 64 | Resistant | Broth microdilution (CLSI) | SAMN04868738 (Pitt et al. | Greece | B | 2014 |
| 19_GR_14 | 64 | Resistant | Broth microdilution (CLSI) | SAMN04868739 (Pitt et al. | Greece | B | 2014 |
| 23_GR_12 | 8 | Resistant | Broth microdilution (CLSI) | SAMN04868743 (Pitt et al. | Greece | B | 2012 |
| 4_GR_12 | 32 | Resistant | Broth microdilution (CLSI) | SAMN04868724 (Pitt et al. | Greece | B | 2012 |
| 7_GR_13 | 64 | Resistant | Broth microdilution (CLSI) | SAMN04868727 (Pitt et al. | Greece | B | 2013 |
| 8_GR_13 | 64 | Resistant | Broth microdilution (CLSI) | SAMN04868728 (Pitt et al. | Greece | B | 2013 |
| 9_GR_12 | 16 | Resistant | Broth microdilution (CLSI) | SAMN04868729 (Pitt et al. | Greece | B | 2012 |
| ATH10 | 32 | Resistant | Broth microdilution (CLSI) | SAMN12228971 (Zhu et al. | Greece | B | 2016 |
| ATH10 | 64 | Resistant | Broth microdilution (CLSI) | SAMN12228971 (Zhu et al. | Greece | B | 2016 |
| ATH15 | 0,125 | Susceptible | Broth microdilution (CLSI) | SAMN12228972 (Zhu et al. | Greece | E | 2016 |
| ATH16 | 128 | Resistant | Broth microdilution (CLSI) | SAMN12228973 (Zhu et al. | Greece | B | 2016 |
| ATH17 | 0.25 | Susceptible | Broth microdilution (CLSI) | SAMN12228974 (Zhu et al. | Greece | B | 2016 |
| ATH18 | 128 | Resistant | Broth microdilution (CLSI) | SAMN12228975 (Zhu et al. | Greece | B | 2016 |
| ATH21 | 0.5 | Susceptible | Broth microdilution (CLSI) | SAMN12228976 (Zhu et al. | Greece | B | 2016 |
| ATH22 | 32 | Resistant | Broth microdilution (CLSI) | SAMN12228977 (Zhu et al. | Greece | B | 2016 |
| ATH23 | 0.5 | Susceptible | Broth microdilution (CLSI) | SAMN12228978 (Zhu et al. | Greece | B | 2016 |
| ATH24 | 64 | Resistant | Broth microdilution (CLSI) | SAMN12228979 (Zhu et al. | Greece | B | 2016 |
| ATH25 | 0.5 | Susceptible | Broth microdilution (CLSI) | SAMN12228980 (Zhu et al. | Greece | B | 2016 |
| ATH26 | 64 | Resistant | Broth microdilution (CLSI) | SAMN12228981 (Zhu et al. | Greece | B | 2016 |
| ATH30 | 32 | Resistant | Broth microdilution (CLSI) | SAMN12228982 (Zhu et al. | Greece | B | 2016 |
| ATH8 | 128 | Resistant | Broth microdilution (CLSI) | SAMN12228969 (Zhu et al. | Greece | B | 2016 |
| ATH9 | 0.5 | Susceptible | Broth microdilution (CLSI) | SAMN12228970 (Zhu et al. | Greece | B | 2016 |
| AR_0125 | 4 | Resistant | Broth microdilution (EUCAST) | SAMN04014966 (FDA/CDC) | Not mentioned | B | Not mentioned |
| kp19 | 16 | Resistant | E‐test (BioMérieux) | SAMN06917612 (Rimoldi et al. | Italy | E | 2012 |
| kp34 | 16 | Resistant | E‐test (BioMérieux) | SAMN06917627 (Rimoldi et al. | Italy | E | 2013 |
| kp36 | 0.5 | Susceptible | E‐test (BioMérieux) | SAMN06917629 (Rimoldi et al. | Italy | E | 2013 |
| kp37 | 16 | Resistant | E‐test (BioMérieux) | SAMN06917630 (Rimoldi et al. | Italy | E | 2013 |
| kp38 | 0.5 | Susceptible | E‐test (BioMérieux) | SAMN06917631 (Rimoldi et al. | Italy | E | 2013 |
| kp46 | 0.5 | Susceptible | E‐test (BioMérieux) | SAMN06917639 (Rimoldi et al. | Italy | E | 2013 |
| kp49 | 0.5 | Susceptible | E‐test (BioMérieux) | SAMN06917642 (Rimoldi et al. | Italy | E | 2013 |
| AR_0363 | 0.5 | Susceptible | Not mentioned | SAMN07291506 (FDA/CDC) | Not mentioned | B | Not mentioned |
| AR_0454 | 8 | Resistant | Broth microdilution (CLSI) | SAMN07291547 (FDA/CDC) | Not mentioned | B | Not mentioned |
| Kp1001 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371303 (Elias et al. | Portugal | E | 2004 |
| Kp1003 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371304 (Elias et al. | Portugal | E | 2004 |
| Kp1019 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371306 (Elias et al. | Portugal | E | 2004 |
| Kp1031 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371322 (Elias et al. | Portugal | E | 2005 |
| Kp1032 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371323 (Elias et al. | Portugal | E | 2005 |
| Kp1036 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371324 (Elias et al. | Portugal | E | 2005 |
| Kp1122 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371327 (Elias et al. | Portugal | E | 2005 |
| Kp1144 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371346 (Elias et al. | Portugal | E | 2006 |
| Kp1209 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371347 (Elias et al. | Portugal | E | 2006 |
| Kp1363 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA6829111 (Elias et al. | Portugal | E | 2005 |
| Kp1495 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371363 (Elias et al. | Portugal | E | 2007 |
| Kp1495 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371363 (Elias et al. | Portugal | E | 2007 |
| Kp1507 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371364 (Elias et al. | Portugal | E | 2007 |
| Kp1528 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371385 (Elias et al. | Portugal | E | 2008 |
| Kp1675 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA6829114 (Elias et al. | Portugal | E | 2008 |
| Kp1924 | 2 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371699 (Elias et al. | Portugal | E | 2018 |
| Kp1938 | 1 | Susceptible | Broth microdilution (Eucast) | SAMEA110371700 (Elias et al. | Portugal | E | 2018 |
| Kp1990 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371386 (Elias et al. | Portugal | E | 2008 |
| Kp2162 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371387 (Elias et al. | Portugal | E | 2008 |
| Kp2200 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371389 (Elias et al. | Portugal | E | 2008 |
| Kp2209 | 0.25 | Susceptible | Broth microdilution (EUCASTst) | SAMEA6829115 (Elias et al. | Portugal | E | 2008 |
| Kp2224 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371702 (Elias et al. | Portugal | E | 2008 |
| Kp2287 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371702 (Elias et al. | Portugal | E | 2018 |
| Kp2334 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371391 (Elias et al. | Portugal | E | 2008 |
| Kp2447 | 1 | Susceptible | Broth microdilution (Eucast) | SAMEA110371392 (Elias et al. | Portugal | E | 2008 |
| Kp2454 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371393 (Elias et al. | Portugal | E | 2008 |
| Kp2463 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371427 (Elias et al. | Portugal | E | 2009 |
| Kp2476 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371428 (Elias et al. | Portugal | E | 2009 |
| Kp2497 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371429 (Elias et al. | Portugal | E | 2009 |
| Kp2564 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA6829116 (Elias et al. | Portugal | E | 2009 |
| Kp2568 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371430 (Elias et al. | Portugal | E | 2009 |
| Kp2587 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371431 (Elias et al. | Portugal | E | 2009 |
| Kp2605 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371432 (Elias et al. | Portugal | E | 2009 |
| Kp2606 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371433 (Elias et al. | Portugal | E | 2009 |
| Kp2645 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371434 (Elias et al. | Portugal | E | 2009 |
| Kp2786 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371438 (Elias et al. | Portugal | E | 2009 |
| Kp2864 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371439 (Elias et al. | Portugal | E | 2009 |
| Kp2895 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371440 (Elias et al. | Portugal | E | 2009 |
| Kp2948 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371461 (Elias et al. | Portugal | E | 2010 |
| Kp2958 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA6829117 (Elias et al. | Portugal | E | 2010 |
| Kp3000 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371462 (Elias et al. | Portugal | E | 2010 |
| Kp3046 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371463 (Elias et al. | Portugal | E | 2010 |
| Kp3185 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371464 (Elias et al. | Portugal | E | 2010 |
| Kp3270 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371480 (Elias et al. | Portugal | E | 2011 |
| Kp3323 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371482 (Elias et al. | Portugal | E | 2011 |
| Kp3509 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371484 (Elias et al. | Portugal | E | 2011 |
| Kp3660 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371486 (Elias et al. | Portugal | E | 2011 |
| Kp3725 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371488 (Elias et al. | Portugal | E | 2011 |
| Kp3851 | 2 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371510 (Elias et al. | Portugal | E | 2012 |
| Kp3860 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA6829118 (Elias et al. | Portugal | E | 2013 |
| Kp4164 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371233 (Elias et al. | Portugal | E | 1980 |
| Kp4194 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371261 (Elias et al. | Portugal | E | 1982 |
| Kp4195 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371262 (Elias et al. | Portugal | E | 1982 |
| Kp4197 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371263 (Elias et al. | Portugal | E | 1982 |
| Kp4246 | 0.5 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371236 (Elias et al. | Portugal | E | 1980 |
| Kp4256 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371240 (Elias et al. | Portugal | E | 1980 |
| Kp4265 | 0.5 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371247 (Elias et al. | Portugal | E | 1981 |
| Kp4279 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371264 (Elias et al. | Portugal | E | 1982 |
| Kp4287 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371265 (Elias et al. | Portugal | E | 1991 |
| Kp4292 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371266 (Elias et al. | Portugal | E | 1995 |
| Kp4333 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371269 (Elias et al. | Portugal | E | 1995 |
| Kp4367 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371270 (Elias et al. | Portugal | E | 1995 |
| Kp4387 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371272 (Elias et al. | Portugal | E | 1995 |
| Kp4408 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371274 (Elias et al. | Portugal | E | 1996 |
| Kp4855 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371528 (Elias et al. | Portugal | E | 2013 |
| Kp4856 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371537 (Elias et al. | Portugal | E | 2014 |
| Kp4857 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371538 (Elias et al. | Portugal | E | 2014 |
| Kp4860 | 2 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371541 (Elias et al. | Portugal | E | 2014 |
| Kp4861 | 2 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371542 (Elias et al. | Portugal | E | 2014 |
| Kp4862 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371543 (Elias et al. | Portugal | E | 2014 |
| Kp4864 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371561 (Elias et al. | Portugal | E | 2015 |
| Kp4865 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371562 (Elias et al. | Portugal | E | 2015 |
| Kp4869 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371490 (Elias et al. | Portugal | E | 2011 |
| Kp4871 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371566 (Elias et al. | Portugal | E | 2015 |
| Kp4878 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371573 (Elias et al. | Portugal | E | 2015 |
| Kp4882 | 2 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371582 (Elias et al. | Portugal | E | 2016 |
| Kp4886 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371585 (Elias et al. | Portugal | E | 2016 |
| Kp4887 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371586 (Elias et al. | Portugal | E | 2016 |
| Kp4889 | 16 | Resistant | Broth microdilution (EUCAST) | SAMD00060934 (Elias et al. | Portugal | E | 2016 |
| Kp5505 | 16 | Resistant | Broth microdilution (EUCAST) | SAMD00060934 (Elias et al. | Portugal | E | 2016 |
| Kp5506 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371705 (Elias et al. | Portugal | E | 2019 |
| Kp5508 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371706 (Elias et al. | Portugal | E | 2019 |
| Kp5509 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371707 (Elias et al. | Portugal | E | 2019 |
| Kp5510 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371703 (Elias et al. | Portugal | E | 2018 |
| Kp5511 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371708 (Elias et al. | Portugal | E | 2019 |
| Kp5513 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371709 (Elias et al. | Portugal | E | 2019 |
| Kp5514 | 16 | Resistant | Broth microdilution (EUCASTcast) | SAMEA110371710 (Elias et al. | Portugal | E | 2019 |
| Kp5516 | 16 | Resistant | Broth microdilution (EUCASTast) | SAMEA110371711 (Elias et al. | Portugal | E | 2019 |
| Kp5520 | 16 | Resistant | Broth microdilution (EUCAST) | SAMEA110371713 (Elias et al. | Portugal | E | 2019 |
| Kp684 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371275 (Elias et al. | Portugal | E | 1999 |
| Kp689 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371276 (Elias et al. | Portugal | E | 1999 |
| Kp748 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371282 (Elias et al. | Portugal | E | 2001 |
| Kp776 | 0.25 | Susceptible | Broth microdilution (Eucast) | SAMEA110371283 (Elias et al. | Portugal | E | 2001 |
| Kp804 | 0.25 | Susceptible | Broth microdilution (Eucast) | SAMEA110371285 (Elias et al. | Portugal | E | 2002 |
| Kp828 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371287 (Elias et al. | Portugal | E | 2002 |
| Kp829 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371293 (Elias et al. | Portugal | E | 2003 |
| Kp840 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371294 (Elias et al. | Portugal | E | 2003 |
| Kp850 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371295 (Elias et al. | Portugal | E | 2003 |
| Kp874 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371296 (Elias et al. | Portugal | E | 2003 |
| Kp875 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371297 (Elias et al. | Portugal | E | 2003 |
| Kp888 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371298 (Elias et al. | Portugal | E | 2003 |
| Kp898 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371308 (Elias et al. | Portugal | E | 2004 |
| Kp910 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371309 (Elias et al. | Portugal | E | 2004 |
| Kp918 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371311 (Elias et al. | Portugal | E | 2004 |
| Kp919 | 1 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371312 (Elias et al. | Portugal | E | 2004 |
| Kp972 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371313 (Elias et al. | Portugal | E | 2004 |
| Kp986 | 0.25 | Susceptible | Broth microdilution (Eucast) | SAMEA110371314 (Elias et al. | Portugal | E | 2004 |
| Kp997 | 0.25 | Susceptible | Broth microdilution (EUCAST) | SAMEA110371315 (Elias et al. | Portugal | E | 2004 |
| Haplotype | MgrB | PhoP | PhoQ | PmrA | PmrB | Allele copies | Phenotype |
|---|---|---|---|---|---|---|---|
| 0 | 80 | S | |||||
| 1 | T140P | 2 | S/R | ||||
| 2 | A41Y | T240M, L213M | 5 | S/R | |||
| 3 | N253T | 3 | S | ||||
| 4 | L213M | 2 | S | ||||
| 5 | M66I | 1 | S | ||||
| 6 | E57G | 12 | S | ||||
| 7 | M175V | 3 | S | ||||
| 8 | A110S | 1 | S | ||||
| 9 | S56R | 2 | S | ||||
| 10 | V446G | 1 | S | ||||
| 11 | G53S | 1 | S | ||||
| 12 | A282S | 1 | S | ||||
| 13 | I127V | 1 | S | ||||
| 14 | L28F | 1 | S | ||||
| 15 | D135N | 1 | S | ||||
| 16 | N255I, S350Y | 1 | S | ||||
| 17 | S56R, I422S | 1 | S | ||||
| 18 | T240M, V257A | 1 | S | ||||
| 19 | L239P | 1 | S | ||||
| 20 | T281M | 1 | R | ||||
| 21 | G385C | 1 | R | ||||
| 22 | M175V, G207D | 1 | R | ||||
| 23 | T157P, M175V | 1 | R | ||||
| 24 | H339D | E57G | 1 | R | |||
| 25 | S72L | S409R. H410Y | V280L | 1 | R | ||
| 26 | W20R | 1 | R | ||||
| 27 | Insertion | 31 | R | ||||
| 28 | Insertion | A95S | V446G | 1 | R | ||
| 29 | Insertion | P158R | 1 | R | |||
| 30 | Insertion | M175V | 4 | R | |||
| 31 | Insertion | E57G | 2 | R | |||
| 32 | Insertion | A282R | 1 | R | |||
| 33 | Insertion | L213M | 1 | R | |||
| 34 | Insertion | N253P | 2 | R | |||
| 35 | Insertion | V446G | 1 | R | |||
| 36 | Insertion | L222A | 1 | R | |||
| 37 | Insertion | R16C | A282R | 1 | R | ||
| 38 | Insertion | V27H, F398K | 1 | R | |||
| 39 | Insertion | P74L | N253T | 1 | R | ||
| 40 | Insertion | N253T, T439P | 1 | R | |||
| 41 | Insertion | V27H, P103W, C395A | 2 | R | |||
| 42 | Insertion | G39S, A225T, N253T | 1 | R | |||
| 43 | Insertion | V27H, P103W, C395A | A282R | 1 | R | ||
| 44 | Insertion | R16A, V27H, D73I, P103W, S188T | R160S, E57G | 1 | R | ||
| 45 | K3Stop | 4 | R | ||||
| 46 | K3Stop | E57G | 1 | R | |||
| 47 | K3T | 1 | R | ||||
| 48 | L8Stop | E57G | G345R | 1 | R | ||
| 49 | Q30Stop | M175V | 1 | R | |||
| 50 | S36R | 2 | R | ||||
| 51 | S36R | A282R | 1 | R | |||
| 52 | S36R | P103W, C395A | 1 | R | |||
| 53 | S36R | V27H, P103W, C395A | 1 | R | |||
| 54 | G121A | 1 | R | ||||
| 55 | I88N | 1 | R | ||||
| 56 | V27H, Y265C | E57G | 1 | R | |||
| 57 | T246C | 1 | R | ||||
| 58 | E57G | H61Q | 1 | R | |||
| 59 | Q405A | 1 | R | ||||
| 60 | V27H, Y265T | 1 | R | ||||
| 61 | Y265C | 1 | R | ||||
| 62 | E397G | 1 | R | ||||
| 63 | C395V | A41T | L213M | 1 | R | ||
| 64 | M285L | 1 | R | ||||
| 65 | L63H | 1 | R | ||||
| 66 | V280L | 1 | R | ||||
| 67 | E7Q | 1 | S | ||||
| 68 | T240M | 1 | S | ||||
| 69 | I | L132M | 1 | R | |||
| 70 | D150G | P344L | 1 | S | |||
| 71 | E35A, M66I | 1 | S | ||||
| 72 | P346Q | 1 | S |
| Locus | Subpopulation | Alleles ( | Mutations ( | Haplotypes ( | HD |
| Tajima | Fu and Li's |
|---|---|---|---|---|---|---|---|---|
|
| PM‐S | 123 | 0 | 1 | 0 | 0 | — | — |
|
| 6 | 4 | 0.049 | 0.00015 | −198621 | −4.78393 | ||
|
| 14 | 10 | 0.173 | 0.00037 | −21106 | −3.75183 | ||
|
| 7 | 7 | 0.28 | 0.00266 | −161927 | −1.51506 | ||
|
| 21 | 12 | 0.259 | 0.0028 | −2.28478 | −3.17757 | ||
|
| 103 | 5 | 6 | 0.490 | 0.00116 | −0.83934 | −1.09903 | |
|
| PM‐R | 91 | 3 | 4 | 0.464 | 0.00387 | −0.10052 | −0.56404 |
|
| 3 | 4 | 0.065 | 0.0001 | −0.10052 | −0.56404 | ||
|
| 38 | 22 | 0.49 | 0.00136 | −1.62177 | −3.36191 | ||
|
| 8 | 7 | 0.242 | 0.00047 | −2.18456 | −4.77326 | ||
|
| 22 | 14 | 0.406 | 0.00086 | −1.83389 | −3.97162 | ||
|
| 78 | 2 | 3 | 0.393 | 0.00082 | −2.32146 | −3.50909 | |
|
| MIC 0.25–2.0 | 122 | 0 | 1 | 0 | 0 | — | — |
|
| 6 | 4 | 0.049 | 0.00015 | −1.98621 | −4.78393 | ||
|
| 14 | 10 | 0.173 | 0.00037 | −2.1106 | −3.75183 | ||
|
| 7 | 7 | 0.28 | 0.00056 | −1.61927 | −1.51506 | ||
|
| 21 | 12 | 0.259 | 0.0007 | −2.28478 | −3.17757 | ||
|
| MIC 4.0–16.0 | 35 | 3 | 4 | 0.499 | 0.00401 | −0.46554 | −1.55209 |
|
| 1 | 2 | 0.057 | 0.00009 | −1.13552 | −1.73221 | ||
|
| 16 | 8 | 0.365 | 0.00073 | −2.38341 | −3.54379 | ||
|
| 5 | 5 | 0.393 | 0.00071 | −1.25028 | −2.08249 | ||
|
| 21 | 12 | 0.745 | 0.00108 | −2.05115 | −3.62414 | ||
|
| MIC 32.0–128.0 | 56 | 2 | 3 | 0.447 | 0.00382 | 0.45975 | 0.736 |
|
| 2 | 3 | 0.071 | 0.00011 | −1.45172 | −2.58453 | ||
|
| 29 | 16 | 0.563 | 0.00173 | −1.84334 | −3.03588 | ||
|
| 4 | 4 | 0.138 | 0.00031 | −1.67941 | −2.36802 | ||
|
| 13 | 7 | 0.266 | 0.00055 | −2.3188 | −4.26335 | ||
|
|
| 145 | 0 | 1 | 0 | 0 | — | — |
|
| 7 | 5 | 0.057 | 0.00015 | −2.05404 | −5.22767 | ||
|
| 29 | 21 | 0.29 | 0.00055 | −2.44693 | −6.03302 | ||
|
| 9 | 9 | 0.316 | 0.00062 | −1.64261 | −2.99296 | ||
|
| 25 | 16 | 0.313 | 0.0008 | −2.31222 | −3.46716 | ||
|
|
| 69 | 3 | 4 | 0.19 | 0.00243 | −0.84579 | −0.49716 |
|
| 2 | 3 | 0.058 | 0.00009 | −1.42652 | −2.67576 | ||
|
| 26 | 11 | 0.387 | 0.00133 | −1.89522 | −2.45506 | ||
|
| 4 | 4 | 0.165 | 0.00033 | −1.55338 | −2.48398 | ||
|
| 15 | 8 | 0.36 | 0.00084 | −2.0511 | −1.91352 | ||
|
| PM‐S and PM‐R | 181 | 5 | 7 | 0,447 | 0.00101 | −0.80752 | −0.14645 |
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico10.13039/501100003593
- —Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro10.13039/501100004586
- —CAPES10.13039/501100002322
- —Fundação Oswaldo Cruz, Fiocruz10.13039/501100006507
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Evolution and Genetic Dynamics · Antibiotic Use and Resistance
Introduction
1
Antimicrobial resistance (AMR) is a naturally occurring phenomenon characterised by the ability of microorganisms to withstand the effects of antimicrobial drugs. First noted by Fleming following the discovery of penicillin (Fleming 1945), its medical significance escalated with the emergence of penicillin resistance, a trend exacerbated by the widespread use of antibiotics during World War II (Podolsky 2015). Despite continuous efforts by the pharmaceutical industry to develop new antimicrobial agents, the evolution of resistance has persistently outpaced drug innovation (Ventola 2015). Today, AMR is a global crisis, with environmental dissemination facilitated by multiple vectors, including agricultural runoff, hospital effluents, domestic wastewater and aquaculture systems, underscoring its status as a One Health challenge (Hernando‐Amado et al. 2019).
Projections indicate that by 2050, AMR‐related deaths could rival annual cancer mortality rates, signalling an ongoing ‘silent pandemic’ (O'Neill 2016). In response, the World Health Organization (WHO) established a Global Priority Pathogens List in 2017, updated in 2024, to prioritise research and surveillance of critical threats (WHO 2024). Among these, carbapenem‐resistant Enterobacterales (CRE) are classified as urgent, with carbapenem‐resistant Klebsiella pneumoniae (CRKP) posing a paramount risk (WHO 2024). Carbapenems, a type of β‐lactam antibiotics reserved as last‐line therapies for nosocomial infections, are increasingly rendered ineffective by CRKP (Nordmann et al. 2011).
K. pneumoniae , a Gram‐negative bacterium, emerged as a clinically significant pathogen in the 1970s due to its propensity for multidrug resistance (MDR) and role in hospital‐acquired infections (Podschun and Ullmann 1998). By the 1980s, extended‐spectrum β‐lactamase (ESBL)‐producing strains resistant to cephalosporins were documented (Paterson and Bonomo 2005). The early 2000s marked the rise of carbapenemase KPC producers, which remain a global healthcare burden (Munoz‐Price et al. 2013).
CRKP's resilience stems from its ability to acquire and transmit mobile genetic elements (MGEs), enabling horizontal gene transfer (HGT) within and across species boundaries (Partridge et al. 2018). This genetic plasticity, coupled with resistance to last‐line β‐lactams and third‐generation cephalosporins, has necessitated alternative therapies, such as polymyxins (Nation et al. 2014). Polymyxins, cationic peptides, target the anionic lipopolysaccharide (LPS) of K. pneumoniae , disrupting membrane integrity and inducing cell lysis (Velkov et al. 2013). However, escalating polymyxin use has driven the emergence and dissemination of resistant strains, mediated by mutations in genes such as mgrB, phoPQ, pmrAB and MGE‐borne mcr variants (Moffatt et al. 2010). These genetic alterations modify lipid A via phosphoethanolamine or L‐arabinose additions, reducing polymyxin binding affinity (Olaitan et al. 2014).
While prior studies have focused on resistance‐associated mutations and clonal lineages (Poirel, Jayol, and Nordmann 2015), emerging evidence suggests polymyxin resistance is multifactorial, influenced by environmental conditions and infection site dynamics (El‐Sayed Ahmed et al. 2020). Notably, genetic diversity in susceptible strains and the evolutionary trajectories of resistance genes remain underexplored, with limited application of population genetics to elucidate allele frequency shifts, spatiotemporal evolution or selective pressures in K. pneumoniae populations (Wyres and Holt 2018).
This study aims to delineate the evolutionary forces shaping five key polymyxin resistance determinants (mgrB, phoP, phoQ, pmrA, pmrB) in K. pneumoniae . By analysing genetic diversity in both susceptible and resistant strains under varying polymyxin concentrations, we seek to unravel how antimicrobial pressure drives selection in different K. pneumoniae subpopulations.
Materials and Methods
2
Study Design
2.1
The study sample consisted of isolates for which there were polymyxin susceptibility test results and sequence information for the five genes of interest (pmrAB, phoPQ and mgrB), allowing analysis of genetic diversity in relation to the susceptibility phenotype. Isolates were sampled from three sources. Twenty‐one isolates were processed and analysed for the current study (SisGen registration numbers AD6C6D8, AF42316 and A5F86E8). One hundred and forty‐five isolates for which sequence and polymyxin susceptibility profiles were available in Genbank. Finally, forty‐eight isolates previously described (Conceição Neto et al. 2022), yielding a total of 214 samples (Table 1). Given the importance of following gold standards protocols, isolates included in the study had their polymyxin B or E (colistin) Minimum Inhibitory Concentration (MIC) values determined using broth microdilution assays (207 isolates), adhering to standardised protocols (BrCAST 2023; CLSI 2023; EUCAST 2023). The MIC of six isolates was determined by E‐tests, and for one isolate there was no description of the method used. For population and evolutionary analyses, isolates were stratified into two groups: polymyxin‐susceptible and polymyxin‐resistant strains. The primary criteria for classification was the MIC, with values exceeding 2 mg/L indicating a resistant phenotype, consistent with international breakpoint guidelines (CLSI 2023; EUCAST 2023).
Cultivation, DNA Purification, PCR Amplification and Sanger Sequencing
2.2
Twenty‐one isolates were grown on MacConkey agar plates. Pure colonies were subcultured in brain heart infusion (BHI) broth and incubated at 37°C under agitation (180 rpm) for 16 h. Cryopreservation was performed using a 15% glycerol stock (270 μL 99% glycerol + 1530 μL overnight culture) stored at −80°C. For species reconfirmation, isolates were streaked onto nutrient agar, and pure colonies were transferred to a target plate for matrix‐assisted laser desorption/ionisation time‐of‐flight (MALDI‐TOF) mass spectrometry analysis at the National Institute of Infectious Diseases (INI) platform, using the MALDI TOF‐MS Biotyper system (Bruker Daltonics, v4.1.3). Genomic DNA was purified using the QIAamp DNA Mini Kit (Qiagen, v2023) and eluted in 30 μL of nuclease‐free water. Primers targeting mgrB, phoP, phoQ, pmrA and pmrB and other molecular biology protocols are detailed in Data S1. Amplification was performed using GoTaq Green Master Mix 2× (Promega, v2.0) under optimised thermocycling conditions (Data S1). PCR products were purified with the QIAquick PCR Purification Kit (Qiagen, v2023) and eluted in 30 μL. DNA concentration and purity were assessed using a NanoDrop One spectrophotometer (Thermo Fisher Scientific, v1.12) with A260/A280 and A260/A230 ratios. Sanger sequencing of amplicons was conducted at the FIOCRUZ Genomics Platform using a 3730xl DNA Analyser (Applied Biosystems, v5.0) with BigDye Terminator v3.1 chemistry (Thermo Fisher Scientific, v3.1).
Acquisition of
mgrB , phoP , phoQ , pmrA , pmrB and rpoB Sequences
2.3
Whole Genome Shotgun (WGS) sequence data of 145 isolates were retrieved from the National Center for Biotechnology Information (NCBI) via BioProjects PRJNA307517 (Pitt et al. 2018), PRJEB38289 (Elias et al. 2022), PRJNA385863 (Rimoldi et al. 2017) and PRJNA316321 (FDA/CDC Antimicrobial Resistant Isolate Bank) (NCBI Resource Coordinators 2018). For isolates with RefSeq assemblies, coding sequences (mgrB, phoP, phoQ, pmrA, pmrB, rpoB) were directly extracted from annotated GenBank files. For isolates with raw Sequence Read Archive (SRA) data, FASTQ files were generated using fastq‐dump (SRA Toolkit v3.0.0) (Leinonen et al. 2011). Low‐quality reads and adapters were trimmed using fastp (v0.23.4) (Chen et al. 2018). For FASTq reads from the SRA database, gene coverage was calculated based on the single‐copy gene rpoB as a reference for 98 genomes using the bowtie2 mapper and samtools. For 40 assemblies available in Refseq and Genbank, the genomic coverage values reported by the submitters were used directly. The estimated genomic coverage for SRA and Assembly (Genbank/Refseq) data was similar, 91.2× and 102.2×, respectively. The standard deviation of SRA data from Illumina sequences was lower (about 22.6) when compared to that of assemblies (of 82.0) (Table S1). The reference genome K. pneumoniae MGH78578 (GCF_000016305.1) was downloaded from NCBI GenBank. Functional annotation of raw reads was performed using Prokka (v1.14.6) (Seemann 2014), generating GFF annotation files. Filtered reads were aligned to the reference genome using BWA‐MEM (v0.7.17) (Li 2013) and resulting BAM files were sorted and indexed with Samtools (v1.17) (Danecek et al. 2021). Coding regions were extracted using BEDTools getfasta (v2.31.0) (Quinlan and Hall 2010), enabling gene‐specific sequence recovery from metagenomic alignments. For the twenty‐one isolates processed in LabSUR, mgrB, phoP, phoQ, pmrA and pmrB sequence data was obtained by Sanger sequencing. Forward and reverse reads (including internal primers for phoQ) were aligned to the K. pneumoniae MGH78578 reference genes using Geneious Prime (v2023.1.1) (Kearse et al. 2012), with consensus sequences generated via built‐in error correction algorithms. For neutral marker analysis, the rpoB fragment used in Multi Locus Sequence Type (MLST) analysis of K. pneumoniae isolates was chosen (position 1657 to 2157; Diancourt et al. 2005) (Table S2). For thirty‐six isolates, rpoB allele information was inferred based on the sequence type (ST) reported in the original reference (Conceição Neto et al. 2022). The corresponding rpoB alleles were subsequently retrieved from the Klebsiella pneumoniae MLST database hosted at the Institut Pasteur.
Detection of Nonsynonymous Substitutions
2.4
Multi‐FASTA files were generated for mgrB, phoP, phoQ, pmrA and pmrB, encompassing all 214 isolates. Multiple sequence alignment (MSA) was performed using MEGA11 (v11.0.13) (Kumar et al. 2021) with the ClustalW algorithm under default parameters. Alignments were conducted for both nucleotide sequences and their translated amino acid counterparts. The amino acid MSA was specifically analysed to identify nonsynonymous substitutions relative to the wild‐type sequences of the K. pneumoniae MGH78578 reference genome (GCF_000016305.1) (NCBI Resource Coordinators 2018). Positions exhibiting amino acid deviations from the reference were annotated as putative nonsynonymous mutations, with manual validation to exclude alignment artefacts.
Allelic Screening, Mapping and Haplotype Network Construction
2.5
Nonsynonymous substitutions were consolidated into a unified table integrating data from all five genes (mgrB, phoP, phoQ, pmrA, pmrB) for each isolate, with haplotypes and their allele frequencies annotated. A custom Python script concatenated the multiple sequence alignments of these genes into a single contiguous DNA sequence. For isolates harbouring an insertion in mgrB, a guanine residue was manually added at position 124 to maintain reading frame consistency. The concatenated alignment was analysed in DnaSP v6.12.03 (Rozas et al. 2017) using the ‘Generate Haplotype Data’ function, which excluded invariant sites and generated a Roehl Data File (.rdf) for dow nstream analysis. The median‐joining algorithm (Bandelt et al. 1999) was implemented in Network v10.2 (Fluxus Technology Ltd. 2023) to construct the haplotype network, optimising branch connections based on mutational steps and genetic distances.
Subpopulation Definition and Neutrality Tests
2.6
The 214 isolates were stratified into seven subpopulations based on polymyxin susceptibility phenotypes and mgrB genetic profiles: (1) polymyxin‐susceptible (PM‐S), (2) polymyxin‐resistant (PM‐R), (3) isolates with MICs of 0.25–2.0 mg/L, (4) MICs of 4.0–16.0 mg/L, (5) MICs of 32.0–128.0 mg/L, (6) mgrB wild‐type and (7) mgrB‐altered (e.g., mutations, insertions, truncations, deletions). Neutrality tests were conducted in DnaSP v6.12.03 (Rozas et al. 2017) to assess evolutionary pressures acting on the five resistance genes and of a neutral marker (rpoB MLST fragment; Diancourt et al. 2005). Tajima's D (Tajima 1989) and Fu & Li's D (Fu and Li 1993) were calculated for each subpopulation using segregating sites (polymorphic positions). Negative values (D < 0) indicated potential positive selection or population expansion, while positive values (D > 0) suggested balancing selection or population subdivision.
Results
3
Isolates´ Distribution
3.1
The 214 isolates originated from four countries: Brazil (32.71%, n = 70), Greece (11.68%, n = 25), Italy (3.27%, n = 7) and Portugal (50.93%, n = 109) (NCBI Resource Coordinators 2018; BioProject accessions: PRJNA307517, PRJEB38289, PRJNA385863, PRJNA316321) (Table 1). Whole‐genome sequencing (WGS) had been performed on 145 isolates (67.76%), all sourced from European countries (Greece, Italy, Portugal), whereas sequences were obtained by Sanger sequencing in the case of the 69 Brazilian isolates (32.24%) (Table 1). The overrepresentation of isolates from Portugal (50.9%) and Brazil (32.7%) may skew the analysis and impact the generalisation of the results. To address this point, we looked into the ST distribution of the isolates in the dataset, as this is used to establish clonal relations and phylogeny of K. pneumoniae. We found that the five most frequent STs (ST15, 16.9%; ST258, 14.6%; ST11, 6.7%; ST147, 6.8%; ST437, 6.2%) accounted for 51.1% of the 178 isolates for which we could access rpoB fragment sequences. ST15, ST258, ST11 and ST147 are recognised as high‐risk clones with global distribution, often associated with multidrug resistance and hospital outbreaks in different continents (Wyres et al. 2020; Lam et al. 2021 in this reference see Figures S5 and S6; Arcari and Carattoli 2022; Wu et al. 2023; Szijártó et al. 2016; Rodrigues et al. 2022). ST437 is a single‐locus variant of the pandemic clone ST258, and though its distribution is global (Sahoo et al. 2023), its frequency increase has been documented in Europe (Budia‐Silva et al. 2024). In summary, the sampling of isolates from four countries broadly matches the profile of STs with global dissemination, though the representation of some STs may be skewed (as is the case for ST437).
Susceptibility Profile
3.2
The study population comprised 123 alleles (57.48%) associated with polymyxin susceptibility, and 91 alleles (42.52%) linked to resistance (CLSI 2023) (Table S3). Within the susceptible cohort (n = 123), 77 alleles (62.6%) exhibited no nonsynonymous substitutions, while the remaining 46 alleles (37.4%) harboured ≤ 1 mutation. In contrast, 90 resistant alleles (98.9%) carried ≥ 1 genetic alteration, with single (n = 65, 71.4%) and double (n = 28, 30.8%) mutations predominating (Moffatt et al. 2010). Among mgrB wild‐type isolates (n = 145), 84.83% (n = 123) were susceptible, whereas 15.17% (n = 22) displayed resistance despite intact mgrB. All mgrB‐altered isolates (n = 69) were resistant, with 92.75% (n = 64) exhibiting high‐level resistance (MIC ≥ 16.0 mg/L). MIC‐based stratification revealed three subpopulations: susceptible (0.25–2.0 mg/L, n = 122), intermediate resistance (4.0–16.0 mg/L, n = 35) and high resistance (32.0–128.0 mg/L, n = 56) (Table S3).
Genetic Diversity of Haplotypes
3.3
A total of 73 haplotypes were identified among the 214 analysed, with 23 associated with susceptibility and 49 linked to resistance (Figure 1; Table 2). Haplotypes 1 and 2 harboured alleles exhibiting both phenotypes. Resistance‐associated haplotypes displayed 49 unique variations, while susceptible haplotypes contained 25. Eighty alleles (37.4%) were identical to the reference genome (haplotype 0), indicating conserved sequences (Figure 1; Table 2; Table S3). Haplotype 27, the second most frequent (31 alleles), was exclusively resistant and characterised solely by mgrB insertions. Notably, 59 haplotypes (79.7%) were singletons, reflecting rare alleles (27.57% of the total) (Figure 2).
Haplotype network of all alleles. Green colours related to susceptible phenotype and red ones resistant. Size of circumference is proportional to N.
Diagram for visualisation of changes in MgrB, PmrAB and PhoPQ according to haplotype. The number of isolates in which each haplotype was identified is shown on the right.
Neutrality Tests Across Subpopulations
3.4
Analysis of Tajima's D, Fu & Li's D and haplotype diversity (HD) revealed distinct evolutionary patterns among subpopulations (Table 3). In the polymyxin‐susceptible (PM‐S) group (Figure 3), phoP, phoQ and pmrB exhibited significantly negative Tajima's D values (range: −1.98621 to −2.48068; p < 0.05 to p < 0.01), with even lower Fu & Li's D values (−3.17819 to −4.78393) (Tajima 1989; Fu and Li 1993). No substitutions were detected in mgrB within PM‐S, precluding neutrality calculations for this gene. The D values for pmrA did not show significant deviations from neutrality, and HD remained low (0–0.28). The polymyxin‐resistant (PM‐R) subpopulation (Figure 3) displayed similar trends, except for phoQ, which showed a more pronounced Fu & Li's D (−4.77326 vs. −3.75183 in PM‐S). Notably, PM‐R exhibited higher haplotype diversity (HD = 0.406–0.844). mgrB D values did not deviate significantly from neutrality. Subpopulations stratified by MIC intervals (Figure 3) mirrored PM‐S and PM‐R patterns. Low‐MIC isolates (0.25–2.0 mg/L) mirrored PM‐S in D and HD values. However, intermediate‐ (4.0–16.0 mg/L) and high‐MIC (32.0–128.0 mg/L) groups revealed divergent trends: phoQ Tajima's D increased with MIC (p < 0.05), while pmrB values decreased (p < 0.01). HD peaked in intermediate‐MIC isolates before slightly declining in high‐MIC groups. Comparative analysis of mgrB wild type versus altered subpopulations (Figure 3) highlighted stark contrasts. Wild‐type isolates showed markedly lower Fu & Li's D values (phoP: −5.22767 vs. −2.67576; phoQ: −6.03302 vs. −2.45506) alongside elevated HD (Table 3). Analysis of a neutral marker, the rpoB fragment used in MLST of K. pneumoniae, yielded Tajima's D value of −0.80752 and Fu and Li's D value of −0.14645 (Table 3). Albeit these values are negative, the P values are above 0.05, which is coherent with this sequence not deviating neutrality.
(A) Fu and Li's D value for polymyxin susceptible (PM‐S) and resistant (PM‐R) populations. (B) Fu and Li's D value for MIC intervals populations. (C) Fu and Li's D value for mgrB wild type and altered mgrB populations. Bar colours are related to the gene value.
Discussion
4
Polymyxin Resistance Is Linked to Polygenic Adaptation
4.1
This study revealed high genetic diversity in mgrB, phoP, phoQ, pmrA and pmrB under varying polymyxin concentrations, attributed to the direct role of these five genes in mediating the addition of PEtN and L‐Ara4N to the lipid A moiety of LPS. Naturally negatively charged, the incorporation of these molecules alters LPS charge, hindering polymyxin binding to the cell wall (Jayol et al. 2014). mgrB plays the most prominent role in resistance (Poirel, Jayol, Bontron, et al. 2015), consistent with our findings, where all variations in this gene were identified in the PM‐R subpopulation. Following in relevance, phoQ and pmrB contribute significantly, while phoP and pmrA also influence resistance, albeit with lesser impact (Olaitan et al. 2014). The high diversity of mutations, insertions and deletions was quantified using Tajima's D and Fu & Li's statistics. Notably, all genes exhibited significantly negative values in both polymyxin‐resistant and susceptible populations. More negative D values indicate a higher proportion of segregating sites (Tajima) and rare alleles (Fu & Li), with many alleles showing substitutions at multiple loci (Ferretti et al. 2017). These findings suggest resistance‐related selective pressures across distinct loci, particularly as mgrB, phoPQ and pmrAB are genomically distant in K. pneumoniae . This process strongly parallels polygenic adaptation (Pritchard et al. 2010).
The concept of polygenic adaptation emerged over the 20th century. Ronald Fisher established the mathematical foundation for polygenic traits, proposing that continuous phenotypes like height are governed by multiple loci with small additive effects (Fisher 1918). In 1983, Russel Lande developed models integrating complex trait genetics with molecular evolution, demonstrating how selection acts on traits controlled by multiple loci and formalising additive genetic variance and covariance between loci shape species adaptation. The explicit term ‘polygenic adaptation’ was first introduced in Pritchard et al.'s 2010 review, which posited that this evolutionary process involves concurrent selection of genetic variants distributed across multiple genomic loci. Polygenic adaptation could also arise from new mutations at diverse loci, particularly if a newly favoured phenotype was previously strongly disadvantageous.
The latter scenario may explain the high frequency of substitutions and insertions observed in MgrB, PhoPQ and PmrAB. These proteins naturally modulate LPS charge to confer defence under low/high pH or metal ion scarcity. However, amino acid structural changes may lead to their hyperactivation—via inactivation of MgrB (a negative regulator of PhoQ) or enhanced phosphorylation sites in PhoQ and PmrB (Olaitan et al. 2014). Such alterations render the phenotype disadvantageous in environments lacking polymyxin pressure or with normal pH and Ca^2+^ levels (Janssen et al. 2020). The phenotypic preference observed in our study highlights a clear association between resistance and the abundance of variations in MgrB, PhoPQ and PmrAB, indicating polygenic adaptation in resistant populations. Other genes, such as those within the arn operon or pmrCAB, are also linked to polymyxin resistance. However, these were not analysed here, necessitating future investigation to confirm whether mutations in these loci synergistically alter resistance phenotypes alongside the studied genes.
The Studied Population Is Undergoing Recent Expansion
4.2
The results demonstrated very low Tajima's D values in polymyxin‐susceptible subpopulations. Two lines of evidence may explain these low values. First, there may be a bias due to the high frequency of alleles associated with haplotype 0 (N = 80). Haplotype 0 matches the reference genome used in this study and can be considered the ancestral haplotype of the entire population (Bandelt et al. 1999). Low D values are linked to a disproportion between haplotype frequencies and an excess of rare alleles in the subpopulation, with 17 out of 23 PM‐S haplotypes (73.91%) having N = 1. These data suggest a population expansion in PM‐S, as novel alleles are emerging from the ancestral haplotype, even at low frequencies.
Unlike Tajima's D, Fu & Li's statistics exhibit greater sensitivity to recent population expansion events. This is because Fu & Li's mathematical model weights segregating sites by singletons, which the authors proposed are linked to recent expansions, as such alleles have not yet had sufficient time to increase in frequency. The high number of singletons in PM‐S reflects a rapid population expansion of alleles carrying mutations in polymyxin resistance‐associated genes. Notably, polymyxins were officially reintroduced for treating hospital‐acquired infections around 2010, when they were recommended as a last‐resort option for infections caused by carbapenem‐resistant bacteria (Landman et al. 2008). Given that most strains analysed here date to the 2010s, the emergence of singletons may coincide with this period, reflecting a transitional spatiotemporal stage where these alleles have neither become fixed nor purged. A second supporting observation is that even minimal inhibitory concentrations (MICs) ≤ 2 μg/mL—associated with susceptibility—appear sufficient to promote adaptive emergence of new alleles, even if they are not directly linked to resistance. However, robust evidence for active selection in PM‐S is lacking, as the ancestral haplotype remains dominant and unrelated to polymyxin adaptation.
A similar pattern is observed in PM‐R, with low D values and the prevalent haplotype 27 (N = 31) followed by 41 rare alleles. Intriguingly, two primary expansion events underline resistant allele origins. The first stems from the ancestral haplotype, giving rise to multiple rare resistant haplotypes (e.g., haplotypes 45, 54 and 65). The second originates from haplotype 27 itself, which diverges from the ancestral lineage and spawns additional rare haplotypes (e.g., 32, 33, 47, 69) (Figure 1). Thus, two distinct expansion events occurred in the overall population: one in PM‐S arising from the ancestral haplotype and a second in PM‐R originating from haplotype 27. The latter event exclusively produced PM‐R alleles, highlighting divergent evolutionary trajectories under selective pressure.
Alterations in
mgrB Drive Positive Selection via Selective Sweeps
4.3
The analysed subpopulations exhibit Fu & Li's D values below −3.0, which is indicative of extreme population expansion. Intriguingly, the mgrB‐altered subpopulation showed D values ranging from −1.91352 to −2.67576 (Fu & Li) and −1.42652 to −2.0511 (Tajima) for significant comparisons. While these values are less extreme, they remain notably elevated, suggesting potential positive selection acting on mgrB. Allelic frequency analysis revealed a decline in rare alleles with substitutions in phoP, phoQ, pmrA and pmrB when compared to haplotype 27 (N = 31/68), which dominates nearly half of the subpopulation. Haplotype 27 carries insertions such as ISKpn26, which inactivate mgrB as a negative regulator of histidine kinases (HKs), leading to LPS charge modification via hyperactivation (Jayol et al. 2014). This haplotype lacks mutations in phoP, phoQ, pmrA or pmrB, potentially explaining its higher D value. The high frequency of haplotype 27 implies it is sweeping the population through selective sweep dynamics, a hallmark of positive selection (Pritchard et al. 2010; Booker et al. 2017).
Selective sweeps occur when an allele confers a significant adaptive advantage, increasing its frequency across generations while reducing genetic diversity. These are categorised as ‘hard’ (complete or incomplete) or ‘soft’ sweeps. Hard sweeps involve a single advantageous mutation rising to fixation, dragging linked genomic regions along due to reduced recombination (classic model). Incomplete hard sweeps reflect ongoing fixation processes. Soft sweeps arise when preexisting neutral alleles become advantageous under new selective pressures or when multiple independent mutations confer similar benefits, preserving genetic diversity (Booker et al. 2017). Polygenic adaptation, as previously discussed, can also drive partial sweeps, where selection acts gradually across loci, maintaining diversity.
Our findings indicate that haplotype 27 is sweeping the PM‐R subpopulation via horizontal gene transfer (HGT)‐mediated insertion in mgrB rather than point mutation. HGT‐driven sweeps can propagate adaptive alleles faster than clonal reproduction (Wilson et al. 2016), potentially explaining haplotype 27's rapid dominance. Conversely, clonally derived haplotypes with mutations reflect partial sweeps linked to polygenic adaptation, sustaining genetic diversity.
The observed diversity may also stem from compensatory mutations unrelated to direct resistance mechanisms. These mutations mitigate fitness costs imposed by primary adaptations, such as LPS modifications, enabling resistant strains to remain competitive in non‐selective environments (Levin et al. 2000). Additionally, genetic hitchhiking—where neutral or deleterious mutations rise in frequency due to linkage with advantageous loci—may influence diversity in mgrB, phoPQ and pmrAB. This phenomenon, first described by Maynard Smith and Haigh (1974), is exemplified in Mycobacterium tuberculosis , where non‐resistance mutations fixed sequentially alongside adaptive alleles during antibiotic treatment (Eldholm et al. 2014).
Collectively, these findings suggest PM‐R is undergoing positive selection via mgrB‐targeted sweeps. Whether this sweep progresses toward fixation or remains incomplete hinges on polygenic adaptation or compensatory evolution, which could sustain genetic diversity through persistent low‐frequency alleles. Such dynamics underscore the complex interplay between selection, genetic linkage and adaptive trade‐offs in shaping resistance evolution.
Impact of Findings on Healthcare and Hospital Environments
4.4
Recent epidemiological data indicate that polymyxin resistance in K. pneumoniae has a global prevalence of approximately 11.64%, with regional variations: 10.17% in Asia, 16.16% in Europe and 18.67% in the Americas (Sameni et al. 2022). In Brazil, we previously reported that 29% of carbapenem‐resistant K. pneumoniae (CRKP) isolates from diverse regions exhibited resistance to polymyxin B (Conceição‐Neto et al. 2022). As CRKP remains a top priority for surveillance and research (WHO 2024), polymyxin use is likely to escalate in coming decades unless alternative therapies are integrated into clinical protocols.
Our study highlights a rapid population expansion of resistant strains following the reintroduction of polymyxins in hospital settings. This expansion originates from both the ancestral haplotype (haplotype 0) and the mgrB‐altered haplotype 27. In the case of haplotype 27, selective sweeps are driving its overrepresentation among resistant populations. If these sweeps proceed to completion, this haplotype is expected to dominate hospital environments, with rare alleles failing to establish.
However, mechanisms such as compensatory evolution may impede complete sweeps. Outside high‐polymyxin environments, the fitness cost of a positively charged LPS reduces strain competitiveness (Janssen et al. 2020). Rare alleles derived from haplotype 27 (e.g., haplotypes 28–44) encode amino acid substitutions in phoP, phoQ, pmrA and pmrB, potentially attenuating histidine kinase (HK) activity in PhoQ and PmrB despite their overexpression. These mutations may downregulate arn operon transcription via PhoP and PmrA, reducing LPS positive charge. Such intermediate charge could enhance fitness in non‐selective environments, enabling strains to compete with other microbes or evade host immunity. Susceptibility might also re‐emerge if mutations in phoP, phoQ, pmrA or pmrB prove deleterious.
Cross‐sensitivity represents another critical factor. While the isolates studied exhibited defined MICs for polymyxins, clinical treatment histories—unavailable here—may involve β‐lactams or other antibiotics. Thus, these strains could retain susceptibility to alternative agents, underscoring the need for comprehensive antimicrobial stewardship to delay resistance fixation. Collectively, these dynamics emphasise the complex interplay between adaptive evolution, compensatory mechanisms and therapeutic practices in shaping resistance trajectories within healthcare ecosystems.
Final Considerations and Study Limitations
4.5
This study elucidates the evolutionary mechanisms underpinning polymyxin resistance in K. pneumoniae , emphasising the interplay of positive selection, polygenic adaptation, population expansion and selective sweeps. The dominance of mgrB insertions in resistant populations underscores their central role in resistance via LPS modification. However, mutations in phoPQ and pmrAB also contribute to the phenotype, illustrating a polygenic architecture where multiple loci collectively drive adaptation. The prevalence of rare alleles and significantly negative Tajima's D and Fu & Li's values in both PM‐S and PM‐R suggest recent expansions temporally aligned with the resurgence of polymyxin use post‐2010. In PM‐S, the ancestral haplotype persists alongside emerging haplotypes, while in PM‐R, haplotype 27 is overrepresented. Haplotypes derived from haplotype 27 exhibit mutations in other loci, implying compensatory evolution to mitigate fitness costs, which could sustain genetic diversity despite strong selection.
The rapid clonal spread of haplotype 27, facilitated by horizontal gene transfer, exemplifies a classic hard sweep. However, incomplete fixation and persistent diversity point to soft sweeps or polygenic adaptation, where multiple mutations sustain resistance without eliminating variation. This complexity complicates containment strategies, as resistance may arise through diverse pathways.
These findings carry profound implications for global health. Without novel therapies, reliance on polymyxins will fuel further resistance, particularly in endemic regions. Compensatory mutations, while stabilising bacterial fitness, may indirectly prolong resistance persistence even in low‐drug‐pressure environments. Several study limitations warrant acknowledgment. The analysed population is historical rather than natural, with overrepresentation from Brazil and Portugal and limited European allele diversity. Additionally, mcr genes—though globally rare—were not assessed but may exist in the strains used in the study.
Future studies should expand genomic analyses to include arn and pmrC operons, given their influence on resistance. Experimental validation via gene‐edited mutants is essential to confirm compensatory mutations or hitchhiking effects in phoPQ and pmrAB. Addressing these gaps will refine our understanding of resistance dynamics and inform strategies to mitigate its evolution in clinical settings.
Author Contributions
This study was designed by Daniel Miceli Serwy, Fabio Faria da Mota, Teca Calcagno Galvao and Viviane Zahner. Daniel Miceli Serwy conducted the experimental work. Data analysis was performed by Daniel Miceli Serwy, Ana Luiza Carneiro Alencar, Roberto Leonan Morim Novaes and Josué da Costa Lima‐Junior. Ana Paula Carvalho‐Assef and Maria Eduarda Rocha Conde designed and carried out MIC testing, respectively. The manuscript was written by Daniel Miceli Serwy, Teca Calcagno Galvao and Viviane Zahner.
Funding
Daniel Serwy's Masters program scholarship was funded by CAPES (Foundation for the Coordination of Improvement of Higher Education Personnel, Education Ministry/MEC). The project was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq grant 421136/2023‐5), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ grants E‐26/210.228/2018 and 11E‐26/210.982/2021) and Fiocruz.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Data S1: mec70234‐sup‐0001‐DataS1.docx.
Table S1: mec70234‐sup‐0002‐TableS1.docx.
Table S2: mec70234‐sup‐0003‐TableS2.docx.
Table S3: mec70234‐sup‐0004‐TableS3.docx.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arcari, G. , and A. Carattoli . 2022. “Global Spread and Evolutionary Convergence of Multidrug‐Resistant and Hypervirulent Klebsiella pneumoniae High‐Risk Clones.” Pathogens and Global Health 117, no. 4: 328–341.36089853 10.1080/20477724.2022.2121362 PMC 10177687 · doi ↗ · pubmed ↗
- 2Bandelt, H. J. , P. Forster , and A. Röhl . 1999. “Median‐Joining Networks for Inferring Intraspecific Phylogenies.” Molecular Biology and Evolution 16, no. 1: 37–48.10331250 10.1093/oxfordjournals.molbev.a 026036 · doi ↗ · pubmed ↗
- 3Booker, T. R. , B. C. Jackson , and P. D. Keightley . 2017. “Detecting Positive Selection in the Genome.” BMC Biology 15, no. 1: 98.29084517 10.1186/s 12915-017-0434-y PMC 5662103 · doi ↗ · pubmed ↗
- 4Br CAST (Brazilian Committee on Antimicrobial Susceptibility Testing) . 2023. “Recommendations for Antimicrobial Susceptibility Testing (AST).” https://brcast.org.br/documentos.
- 5Budia‐Silva, M. , T. Kostyanev , S. Ayala‐Montaño , et al. 2024. “International and Regional Spread of Carbapenem‐Resistant Klebsiella pneumoniae in Europe.” Nature Communications 15: 5092.10.1038/s 41467-024-49349-z PMC 1117887838877000 · doi ↗ · pubmed ↗
- 6Chen, S. , Y. Zhou , Y. Chen , and J. Gu . 2018. “Fastp: An Ultra‐Fast All‐In‐One FASTQ Preprocessor.” Bioinformatics 34, no. 17: i 884–i 890.30423086 10.1093/bioinformatics/bty 560PMC 6129281 · doi ↗ · pubmed ↗
- 7CLSI (Clinical and Laboratory Standards Institute) . 2023. Performance Standards for Antimicrobial Susceptibility Testing. 33rd ed. CLSI Supplement M 100.
- 8Conceição‐Neto, O. C. , B. S. da Costa , L. D. S. Pontes , et al. 2022. “Polymyxin Resistance in Clinical Isolates of K. pneumoniae in Brazil: Update on Molecular Mechanisms, Clonal Dissemination and Relationship With KPC‐Producing Strains.” Frontiers in Cellular and Infection Microbiology 12: 898125.35909953 10.3389/fcimb.2022.898125 PMC 9334684 · doi ↗ · pubmed ↗