Chloracidobacterium validum sp. nov., a thermophilic chlorophotoheterotrophic bacterium of the phylum Acidobacteriota from an alkaline hot spring microbial mat, represents Chloracidobacterium gen. nov., Chloracidobacteriaceae fam. nov. and Chloracidobacteriales ord. nov
Mohit Kumar Saini, Steven B. Kuzyk, Cristian Villena-Alemany, Sarah Kirstein, Jacqueline Wolf, Meina Neumann-Schaal, Shin Haruta, Satoshi Hanada, Michal Koblížek, Vera Thiel, Marcus Tank, Donald A. Bryant†

TL;DR
Scientists discovered a new thermophilic bacterium from a hot spring microbial mat and classified it as a new species, genus, family, and order.
Contribution
The paper introduces a new species, genus, family, and order of thermophilic chlorophotoheterotrophic bacteria based on phenotypic, phylogenetic, and genomic analyses.
Findings
Strain BV2-CT is a novel thermophilic chlorophotoheterotrophic bacterium requiring branched-chain amino acids for growth.
Genomic analysis revealed two circular chromosomes and a 59.9 mol% G+C content for strain BV2-CT.
The new genus, family, and order are proposed within the class Blastocatellia based on phylogenomic data.
Abstract
A novel chlorophototrophic bacterium, strain BV2-CT, was isolated from a microbial mat (~40 °C) at Rupite hot springs, Bulgaria. Cells were rod-shaped, Gram-negative, with numerous fimbriae/pili, but non-motile. Optimal growth occurred under micro-oxic conditions in the light (20–50 µmol photons m−2 s−1) at 45 °C (range 35–50 °C) and pH 7.2 (range pH 5.5–9.0). Bacteriochlorophyll c, bacteriochlorophyll aP, echinenone and γ-carotene were the major pigments in cells capable of producing chlorosomes and type-1 homodimeric reaction centres. Dense liquid culture appeared greenish-brown, with cells forming small clumps and aggregates. Strain BV2-CT grows photoheterotrophically using organic compounds as carbon source (i.e. a mixture of 20 proteinogenic amino acids or peptone) and thioglycolate as reduced sulphur source. Strain BV2-CT obligately required branched-chain amino acids…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
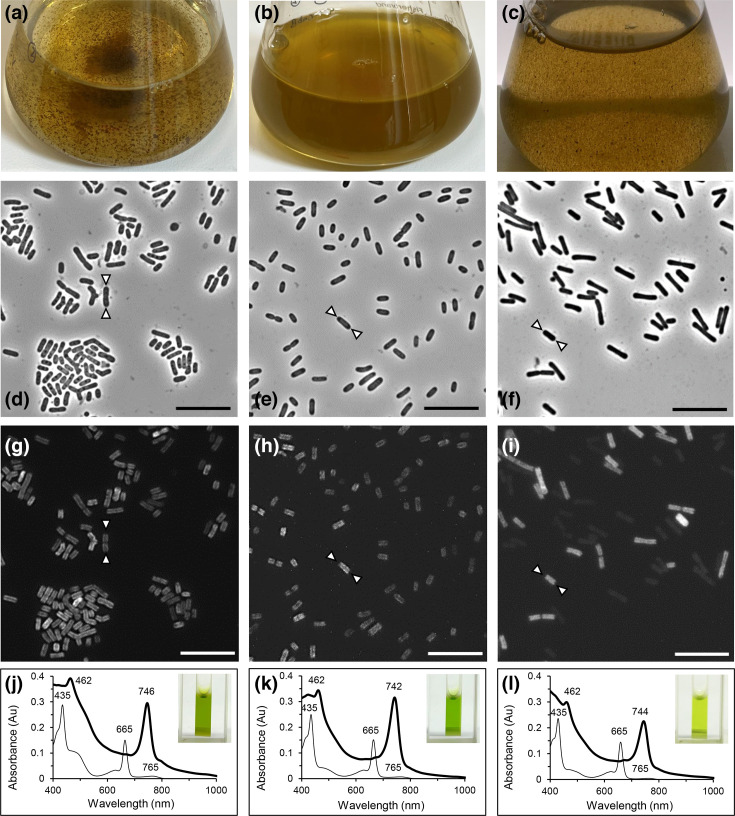 Fig. 1
Fig. 1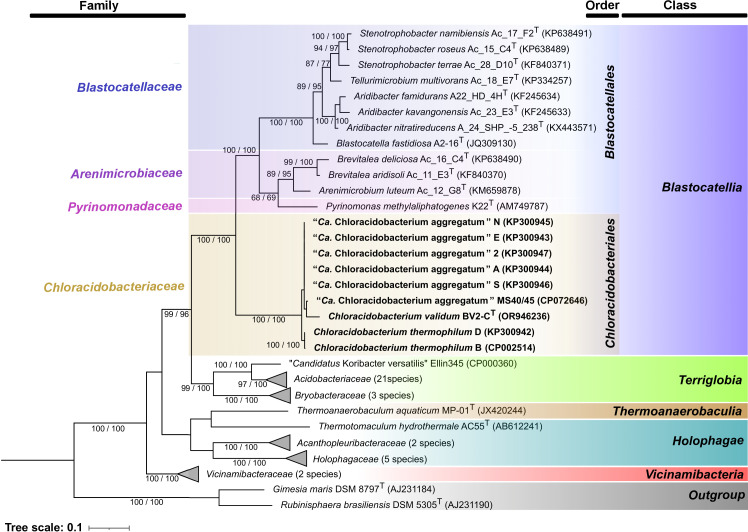 Fig. 2
Fig. 2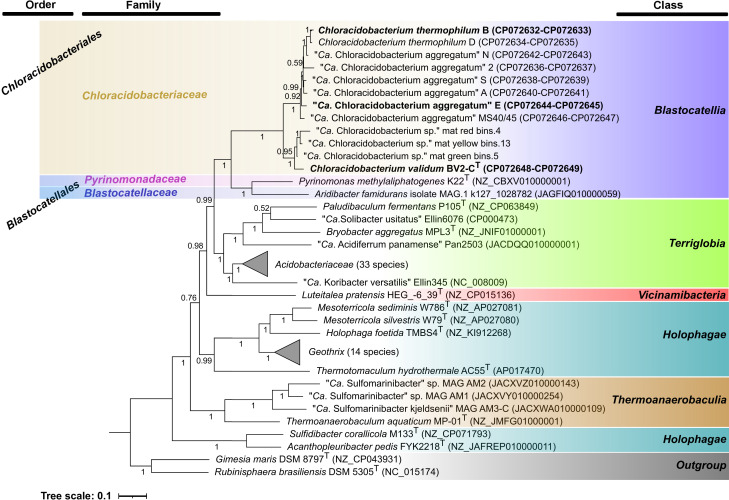 Fig. 3
Fig. 3| Characteristic | Order |
| |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Family |
|
|
|
| |||||||
| Genus |
|
|
|
|
|
|
| ||||
| Strain | BV2-CT | B* | E† | ||||||||
|
| Rod | Rod | Rod | Rod | Ovoid to rod | Rod | Sphere to rod | Rod | Rod | Rod | |
|
| 2–4×0.8–1.0 | ~2.5×0.8–1.0 | 2–4×~0.8 | 1.0–4.0×0.3–0.6 | 1.0–2.3×0.7–1.0 | 0.9–2.2×0.6–0.9 | 0.8–12.0×0.8–0.9 | 1.5–3.0×0.6–0.9 | 0.8–2.3×0.3–0.7 | 0.8–2.5×0.3–0.7 | |
|
| Binary fission | Binary fission | Binary fission | Binary fission | Binary fission | Binary fission | Binary fission/budding | Binary fission | Binary fission | Binary fission | |
|
| No | No | No | No | No | No | Yes | No | No | No | |
|
| 45 (35–50) | ~52 (44–61) | ~50 (39–58) | 65 (50–69) | 28–44.6 (8–44.6) | 35–45 (11–52.5) | 30–35 (14–40) | 24–44 (8–47) | 34–37 (14–43) | 29–41 (16–41) | |
|
| 7.2 (5.5–9.0) | 7 (5.5–9.5) | ~7.0 (5.5–9) | 6.5 (4.1–7.8) | 5.5–7.0 (3.5–9.5) | 5.4–7.0 (4.7–9.0) | 5.0–7.5 (4.0–10.0) | 5.5–9.0 (3.5–10.0) | 5.6–6.5 (4.7–8.4) | 5.5–8.0 (4.5–10.0) | |
|
| Micro | Microaerophilic | Microaerophilic | Aerobic | Aerobic | Aerobic | Aerobic | Facultatively anaerobic to aerobic | Aerobic | Aerobic | |
|
| Greenish-brown | Greenish-brown | Greenish-brown | White-semitransparent | Orange-yellow | White | Orange-pink | White/yellow/pink | White (bright pinkish hue) | Pink | |
|
| Aggregates | Homogenous | Aggregates | Aggregates | Aggregates | Aggregates | No aggregates | Aggregates/no aggregates | Small aggregates | No aggregates | |
|
| Anoxygenic | Anoxygenic | Anoxygenic | Chemoorgano | Chemoorgano | Chemoorgano | Chemoorgano | Chemoorgano | Chemoorgano | Chemoorgano | |
|
| BChl | BChl | BChl | NP | NP | NP | NP | NP | NP | NP | |
|
| Echinenone, canthaxanthin, | Echinenone, canthaxanthin, | Echinenone, canthaxanthin, |
|
|
|
|
|
|
| |
| No | Yes | (Yes)§ |
|
|
|
|
|
|
| ||
|
| Yes | Yes | Yes |
|
|
|
|
|
|
| |
|
| Yes | Yes | Yes |
|
|
|
|
|
|
| |
|
| MetH; no MetE | MetH; no MetE | MetH; no MetE |
| – | – | – | – | – | – | |
|
| – | – | – | – | – | – | |||||
| NrdAB (Class Ia) | Present | Present | Present |
| – | – | – | – | – | – | |
| NrdJ (Class II) | Absent | Present | Absent |
| – | – | – | – | – | – | |
| NrdD-like paralog (Class III) | Present (no NrdG) | Present (no NrdG) | Present (no NrdG) |
| – | – | – | – | – | – | |
|
| BchE (O2-independent) | BchE (O2-independent) | BchE (O2-independent) |
| – | – | – | – | – | – | |
| 3.66 | 3.75 | 3.77 | ~3.80 | – | – | – | ~3.80–4.40¶ | – | – | ||
| Number of chromosomes | 2 | 2 | 2 |
| – | – | – | – | – | – | |
| Chromosome sizes (Mb) | 2.66+1.0 | 2.70+1.05 | 2.67+1.09 |
| – | – | – | – | – | – | |
|
| 59.5 | 61.31 | 62.18 | 59.36 | 66.9 | 54.7–55.9 | 46.5 | 46.5–53.2 | 59.4 | 56.8–58.5 | |
| Order | Family | Genus (strains) | Polar lipids | Major fatty acids | Major quinone | |
|---|---|---|---|---|---|---|
|
|
|
| (BV2-CT) | DGTA, lyso DGTA, PE, PME | iso-C15:0, C16:0, iso-C16:0, C16:0, iso-dibolic acid | MK-8(H2) |
| (B) | DGTA, lyso DGTA, PE, PME | iso-C15:0, C16:0, iso-C16:0, C16:0, iso-dibolic acid | MK-8(H2) | |||
| (E) | DGTA, lyso DGTA, PE, PME | iso-C15:0, C16:0, iso-C16:0, C16:0, iso-dibolic acid | MK-8(H2) | |||
|
|
|
| PE, PC | iso-C15:0, iso-C17:0, iso-C19:0, iso-C21:0 | MK-8 | |
|
|
| DPG, PE, PC, PI | iso-C15:0, iso-C17:0, anteiso-C17:0, iso-C15:1 H/C13:0 3-OH | MK-8 | ||
|
| DPG, PE, PC, PI, Unid GPL | iso-C15:0, iso-C17:0, anteiso-C17:0, iso-C15:1 H/C13:0 3-OH | MK-8, MK-7 | |||
|
|
|
| iso-C16:1 | MK-8 | ||
|
| DPC, PC, PE, PG | iso-C15:0, C13:0 3-OH/iso-C15:1 H, C16:1 | MK-8 | |||
|
| DPG, PE, PC, PG, Unid GPL | C18:1 | MK-8 | |||
|
| DPG, PE, PC, PG | iso-C15:0, C16:1 | MK-8 |
| Organisms | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 16S (%)* | dDDH (%)† | ANI (%)‡,§ | AAI (%)‖ | 16S (%) | dDDH (%) | ANI (%) | AAI (%) | 16S (%) | dDDH (%) | ANI (%) | AAI (%) | |
| 100 | 100 | 100 | 100 | 96.83 | 19.10 | 75.22 | 80.20 | 97.41 | 19.20 | 75.38 | 80.30 | |
| 96.83 | 19.10 | 75.22 | 80.20 | 100 | 100 | 100 | 100 | 98.95 | 53.50 | 93.64 | 95.10 | |
| 97.41 | 19.20 | 75.38 | 80.30 | 98.95 | 53.50 | 93.64 | 95.10 | 100 | 100 | 100 | 100 | |
| 85.75 | 23.10 | 66.40 | 52.50 | 86.29 | 18.90 | 66.40 | 52.90 | 85.81 | 21.60 | 66.81 | 52.60 | |
| 83.63 | (21.8) | (65.6) | (49.9) | 83.02 | (19.7) | (65.98) | (50.1) | 83.17 | (22.0) | (65.5) | (50.1) | |
| 84.91 |
|
|
| 85.13 |
|
|
| 85.22 |
|
|
| |
| 85.58 |
|
|
| 85.13 |
|
|
| 84.68 |
|
|
| |
| 83.35 |
|
|
| 83.50 |
|
|
| 83.37 |
|
|
| |
| 84.13 |
|
|
| 84.01 |
|
|
| 83.88 |
|
|
| |
| 84.49 |
|
|
| 84.83 |
|
|
| 84.78 |
|
|
| |
- —OP JAK program of the Czech Ministry of Education
- —http://dx.doi.org/10.13039/501100019563 Mikrobiologický Ústav, Akademie Věd České Republiky
- —Tokyo Human Resources Fund for City Diplomacy scholarship
- —Photosynthetic Systems Program, Division of Chemical Sciences, Geosciences, and Biosciences (CSGB), Office of Basic Energy Sciences of the U. S. Department of Energy
- —NASA Exobiology program
- —Institute of Fermentation, Osaka (IFO), Japan.
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Photosynthetic Processes and Mechanisms · Algal biology and biofuel production
Introduction
Members of the phylum Acidobacteriota Thrash and Coates 2021, with its first isolate Acidobacterium capsulatum strain 161 obtained from acidic mineral environments [1], are ubiquitously distributed in diverse habitats, including non-acidic soils [24], the rhizosphere [56], sponges [7], deep-sea plankton or sediments [89] and caves [10]. They are also found in extreme habitats, including hot springs [1113], acidic mining lakes [14] and uranium-contaminated soils [15]. Collectively, these bacteria have a phylogenetic diversity nearly as broad as the Pseudomonadota [16]. The phylum Acidobacteriota Thrash and Coates 2021 [1719] encompasses 26 subdivisions clustered in 15 classes [20], of which only 5 are validly named under the International Code of Nomenclature of Prokaryotes (ICNP) and contain cultured representatives (Blastocatellia Pascual et al. 2016 [2122], Terriglobia Thrash and Coates 2022 [2324], Vicinamibacteria Dedysh and Yilmaz 2018 [2025], Thermoanaerobaculia Dedysh and Yilmaz 2018 [2025] and Holophagae Fukunaga et al. 2008 [2627]).
Bioinformatic analyses of metagenomic sequences provided initial evidence for the occurrence of a novel chlorophototroph with homodimeric type-1 reaction centres and chlorosomes in microbial mats associated with the alkaline, siliceous hot springs, Octopus and Mushroom Spring, in Yellowstone National Park (YNP), WY, USA [28]. Initially established as an enrichment culture, Chloracidobacterium thermophilum strain B was shown to belong to the acidobacteria with the pro-validly published name ‘Candidatus C. thermophilum’ [2829]. Leveraging information gleaned from a consensus (non-clonal) genome sequence [30] and information from diel metatranscriptomic analyses of the organism in situ [31], Tank and Bryant described strain B as an axenic isolate with defined growth medium in 2015 [3233]. C. thermophilum strain B is an oligotroph with auxotrophies for branched-chain amino acids (l-leucine, l-isoleucine and l-valine), l-lysine and vitamin B_12_, further requiring reduced sulphur compounds as a sulphur source, with its growth stimulated by the addition of bicarbonate [3234]. Although C. thermophilum is unable to synthesize branched-chain amino acids, it nonetheless can degrade them as well as other amino acids and use them as a carbon, nitrogen and sulphur source [30,3234]. C. thermophilum is additionally a thermophilic microaerophile that obligately requires traces of oxygen to synthesize chlorophylls (Chls) and the amino acid tyrosine from phenylalanine, but is unable to tolerate atmospheric levels of oxygen [30,3234].
Besides Octopus and Mushroom Springs in YNP, organisms similar to Chloracidobacterium have been found in microbial mats associated with other circumneutral and alkaline hot springs in YNP [35] and the Ojo Caliente hot springs in New Mexico, USA [36]. Related bacteria have further been detected during surveys of hot spring microbial mats around the world, for instance, in Tibet [3738], Thailand [39], Japan [40] and Bulgaria [41]. Consequently, different species and/or ecotypes of Chloracidobacterium are suggested to be associated with the microbial mats of alkaline and circumneutral hot springs worldwide [3442]. Based on the characterization of nine representing isolates, Chloracidobacterium is presently the sole genus of the phylum Acidobacteriota whose members perform Chl-dependent phototrophy (i.e. chlorophototrophy) [20323441]. While affiliated with the class Blastocatellia (formerly subdivision 4), this genus is only distantly related to the other genera within the class [203241]. Although it has been assumed that these bacteria potentially define a novel order and family [20], the previously limited number of effectively published names within this genus inhibited the description of a corresponding order and family.
The development of a defined growth medium for C. thermophilum [3233] allowed for several additional strains of Chloracidobacterium species to be isolated from alkaline hot spring environments. Saini et al. [41] recently compared the genomes of nine axenic strains of Chloracidobacterium, of which eight strains were isolated from hot springs in YNP and one strain from Rupite hot spring in Bulgaria. Comparative bioinformatics suggested that these strains represent three species, C. thermophilum, C. aggregatum and C. validum [41]. Currently, the names ‘Chloracidobacterium’ and ‘C. thermophilum’ are only effectively and not validly published due to restrictions on the availability of the type strain B [32]. This prevented a valid publication of novel higher taxa (fam. nov. and ord. nov.). We here propose the names Chloracidobacterium validum sp. nov. (with strain BV2-C^T^ as type strain) as well as Chloracidobacterium gen. nov. (with C. validum as type species), Chloracidobacteriaceae fam. nov. and Chloracidobacteriales ord. nov., within the class Blastocatellia Pascual et al. 2016, phylum Acidobacteriota Thrash and Coates 2021 [2543]. We further propose the name ‘Candidatus Chloracidobacterium aggregatum’ for pro-valid publication with the genome sequence of strain E as nomenclatural type and emend the description of the pro-validly published species name of ‘Ca. C thermophilum’ Bryant et al. 2007 [2829].
Sampling site, isolation and purification
Strain BV2-C^T^ was isolated from a Rupite hot spring microbial mat, located in the Sandanski-Petrich basin, Blagoevgrad Province, SW, Bulgaria (41° 21′ N 23° 14′ E). Situated in the Baba Vanga sanctuary, this spring feeds a cascade of six thermal water pools (BV1–BV6) connected by narrow channels (BV C1–5) (Fig. S1, available in the online Supplementary Material; also see [44]). A microbial mat sample was collected in April 2015 from pool BV2 at a site with a temperature of ~40 °C and pH 7.5–8.0 (Fig. S1). For isolation and purification of strain BV2-C^T^, mat sample material was incubated in agar plates containing C. thermophilum midnight (CTM) medium (Text S1) in the same way as previously described in detail [3341]. The isolation and purification of strain BV2-C^T^ was performed under micro-oxic conditions (2–10% oxygen) with continuous illumination provided by incandescent lamps (60 W, 20–50 µmol photons m^−2^ s^−1^) at 45 °C. Axenicity was initially assessed by epifluorescence phase-contrast microscopy (Nikon Eclipse E600, Nikon, Japan) as described [33] and later verified by genome analysis (see below). Once axenic, colonies were transferred into freshly prepared liquid CTM medium to obtain larger amounts of bacterial biomass for further characterization. C. thermophilum strains B and D, as well as C. aggregatum strains E, N, S, A, 2 and MS40/45, were isolated from samples of hot spring associated microbial mats, recovered from Mushroom and Octopus Springs in YNP, WY, USA. Samples were taken under National Park Service (NPS Scientific Research and Collecting Permit number #YELL-SCI-0129 [3341].
Phenotypic and biochemical characterization
Liquid cultures of strain BV2-C^T^ were similar in colour to strain B, i.e. greenish-brown (Fig. 1a, b). Strain BV2-C^T^ formed visible cellular aggregates in liquid culture (Fig. 1a), similar to cultures of C. aggregatum strain E (Fig. 1c), and was in contrast to the homogenous cell suspensions of C. thermophilum strain B (Fig. 1b). Gram-negative cells of strain BV2-C^T^ were rod-shaped, ~2–4 µm long and 0.8–1.0 µm wide (Fig. 1d), similar to both strains B and E (Fig. 1e, f). Gram-negative cellular membranes with inner peptidoglycan wall were confirmed via transmission electron microscopy (TEM) of thin sections (Fig. S3), prepared as described [45]. TEM of negatively stained cells was performed as described previously [46] and revealed numerous pili or fimbriae, which were larger in diameter and possibly more rigid in strain BV2-C^T^ than those observed on C. thermophilum strain B (Fig. S2). Since flagella were not detected and twitching motility was not observed, these fimbriae are suggested to be involved in attachment, as shown by the clumping of some cultures. Epifluorescence microscopy using a Nikon Eclipse E600 microscope equipped with monochromatic CMOS camera (Orcaflash 4.0, Hamamatsu) and XPS-100 xenon lamp further revealed bacteriochlorophyll (BChl) autofluorescence of all three Chloracidobacterium spp. using 445 nm (+/− 30 nm) excitation filter and 700 nm long pass emission filter (Semrock, Rochester, NY, USA) (Fig. 1g–i). Cells of strain BV2-C^T^ exhibited autofluorescence from BChl c, which was not distributed evenly throughout the cell but was more intense near the lateral cell walls but not at the poles of the cells (Fig. 1g). This distribution has previously been described for C. thermophilum strain B, attributed to chlorosomes placement only along the lateral cell walls [323347]. Taken together, the pattern of autofluorescence and the presence of genes encoding both BChls a and c biosynthesis as well as chlorosome envelope proteins [4147], this provides strong presumptive evidence for the existence of chlorosomes in strain BV2-C^T^.
Dense greenish-brown liquid cultures of (a) aggregating C. validum strain BV2-CT, (b) homogenous C. thermophilum strain B and (c) aggregating ‘Ca. C. aggregatum’ strain E. Phase-contrast microscopic images, showing the rod-shaped cell morphology of (d) strain BV2-CT, (e) strain B and (f) strain E, are complemented by epifluorescence microscopy revealing BChl autofluorescence, but not at the end of the cells (indicated by white arrows) (g)–(i) of the three respective strains. Scale bars are 10 µm. Whole cell (bold line) and extracted pigment (thin line) absorbance spectra of (j) strain BV2-CT, (k) strain B and (l) strain E detailing maxima with green pigment extracts shown as insets.
Pigment analyses
The in vivo (photosynthetic whole cells) and in vitro (organic solvent-extracted pigments) spectra of Chloracidobacterium spp. strains were measured via a PerkinElmer Lambda 365+ (Rodgau, Germany) within the range of 400–1,000 nm. Sample preparation for measurement of the in vivo and in vitro spectra was performed as previously described in 70% glycerol solution and acetone:methanol (7:2), respectively [48]. The in vivo absorption spectrum of strain BV2-C^T^ showed peaks at 746 and 462 nm, suggesting the predominance of BChl c bound in chlorosomes, which was similar to the absorption spectra of strain B and strain E (Fig. 1j–l). Moreover, all Chloracidobacterium strains contain highly similar in vivo spectra with peaks between 742 and 746 nm (Fig. S4). These small deviations in the maxima may suggest minor conformational changes for each photosynthetic complex. In comparison, the in vitro spectrum of strain BV2-C^T^ was identical to the other strains and revealed predominant maxima at 665 and 435 nm (Fig. 1j–l), which represent the Q_y_ band and Soret band of BChl c, respectively. A minor peak at 765 nm further matches trace levels of BChl a in all strains. Complemented by previous work [41], strain BV2-C^T^ and other Chloracidobacterium spp. contain echinenone, canthaxanthin and γ-carotene as predominant carotenoids, and the major BChls are BChls c and aP (Table 1). Further, minor (B)Chls, Chl aPD and Zn-BChl aP′, are also present in functional type-1 reaction centres as in C. thermophilum strain B [4149].
Physiological and biochemical analyses
Unless otherwise specifically stated, all physiological and biochemical experiments involved strain BV2-C^T^ grown using CTM medium in 100 ml Erlenmeyer flasks filled with 80 ml of medium at 45 °C and pH 7.2 under continuous illumination from a 60 W tungsten bulb (∼20–50 µmol photons m^−2^ s^−1^). Due to its oligotrophy, cultures were repetitively fed with aliquots of a 20 proteinogenic amino acid mixture (300–500 mg l^−1^ combined final concentration) at intervals of 3–4 days to obtain higher bacterial biomass yields per culture. The detailed description of CTM medium composition [33] and its preparation is described in Text S1. Strain BV2-C^T^ was able to grow photoheterotrophically in CTM medium under micro-oxic conditions utilizing organic compounds (i.e. a mixture of the 20 proteinogenic amino acids and/or peptone) as carbon source and thioglycolate as reduced sulphur source, similar to all known Chloracidobacterium isolates but unlike any member of Blastocatellales (Table 1). Amino acids could be utilized not only as a carbon source but also served as a supply of nitrogen and reduced sulphur (l-methionine and l-cysteine). Photoautotrophic growth was not observed when incubated with bicarbonate as the only carbon source, which was supported by an absence of carbon fixation genes in all nine sequenced strains [41]. Further, all of the analysed genomes lacked nitrogen fixation-related genes, indicating the strains depend on other nitrogen sources than dinitrogen as mentioned above. Strain BV2-C^T^ did not replicate under dark conditions and thus is an obligate chlorophototroph. It neither grew under a fully anoxic environment nor under atmospheric oxygen levels on solid or in liquid medium. When inoculated into agar deeps, strain BV2-C^T^ grew in the micro-oxic zone (2–10% oxygen) similar to C. thermophilum strain B (Fig. S5). Thus, strain BV2-C^T^ is a microaerophile with a similar oxygen tolerance to C. thermophilum strain B. Growth at different pH values was tested in CTM medium buffered with 20 mM MES (pH 5–6.5), HEPES (pH 7–7.5), Tricine (pH 8–8.5) or CHES (pH 9–9.5). The optimum pH for growth was pH 7.2–7.5 (range pH 5.5–9.0), similar to the optimum value observed for C. thermophilum strain B [3233] or C. aggregatum strain E (Table 1). Strain BV2-C^T^ grew optimally at 45 °C (range 35–50 °C), characterizing it as a moderately thermophilic bacterium. This optimum was lower than that of C. thermophilum strain B (Table 1), yet both are in accordance with the hot spring temperatures from which each was isolated.
Vitamin B_12_ auxotrophy was shown for C. thermophilum strain B in previous studies [3233]. The vitamin requirements for strain BV2-C^T^ (with C. thermophilum strain B as control) were tested by growing both strains in CTM medium (no peptone version) amended either (1) with vitamin solution A (a mixture of 13 vitamins; see Text S1), (2) vitamin solution B (vitamin B_12_), (3) with both vitamin solutions A and B or (4) without any vitamin source. In contrast to C. thermophilum strain B, growth of strain BV2-C^T^ occurred independently of added vitamin B_12_. Growth occurred when cells were provided either with vitamin solution A (mixture of 13 vitamins) or vitamin solution B (vitamin B_12_), whereas strain BV2-C^T^ was inhibited without a vitamin source (Fig. S6). Genomic sequence analysis indicates that strain BVC-2^T^ lacks a complete set of genes for adenosylcobalamin biosynthesis, but a salvage pathway is present that should allow cells to utilize vitamin B_12_ derivatives obtained from the environment [3041]. Vitamin B_12_ is usually used by cells as a cofactor for several important enzymes: cobalamin-dependent methionine synthase (MetH) [50], vitamin B_12_-dependent ribonucleotide reductase (NrdJ) [5152], vitamin B_12_-dependent methyl-malonyl CoA mutase [53] and oxygen-independent magnesium-protoporphyrin IX monomethyl ester oxidative cyclase (BchE) [54]. Differences in the distribution of the genes encoding these enzymes among the Chloracidobacterium spp. could explain the variation in phenotypes (Table 1). For example, genomes of all three species contain Class-Ia ribonucleotide reductase genes (nrdAB), while only C. thermophilum strain B and one C. aggregatum strain (MS 40/45) have Class-II cobalamin-dependent ribonucleotide reductase genes (nrdJ), which could function under anoxic conditions when vitamin B_12_ is present in the environment. Concerning magnesium-protoporphyrin IX monomethyl ester oxidative cyclase: all strains have cobalamin-dependent bchE [54], yet most C. aggregatum strains lack acsF (strain MS40/45 is an exception) (Table 1).
Like C. thermophilum strain B [30], the genome of strain BV2-C^T^ lacks the genes required to synthesize branched-chain amino acids (l-isoleucine, l-leucine and l-valine) and l-lysine but contains genes needed to produce the enzymes capable of degrading such amino acids [41]. To confirm this metabolic inference, strain BV2-C^T^ and C. thermophilum strain B were grown in CTM medium with specific mixtures of amino acids (Fig. S7). Strain BV2-C^T^ and strain B neither grew when all 20 amino acids and/or peptone were omitted from the growth medium nor when supplied a mixture of 16 amino acids (all except branched-chain amino acids and l-lysine). However, both strains could grow when cultures were provided with all proteinogenic branched-chain amino acids and l-lysine; l-lysine alone did not support growth of either strain (Fig. S7). These results indicate that, like C. thermophilum strain B, strain BV2-C^T^ obligatorily requires branched-chain amino acids as well as l-lysine (Table 1). Peptone can serve as a source of these necessary (and other) amino acids, but the biomass yield was lower when only peptone was provided (Fig. S7).
Chemotaxonomy analyses
Cellular respiratory quinones, fatty acids and polar lipids of all strains of Chloracidobacterium were quantified as described previously [55]. MK-8(H_2_) was their major menaquinone, whereas MK-8(H_4_) made up a small fraction in each (Table S1). While minor strain-to-strain deviations in relative abundances in fatty acids were found between every representative (Table S1), clear patterns emerged (Table 2). The predominant fatty acid of Chloracidobacterium is iso-C_15:0_, similar to all other members of the class Blastocatellia, followed by C_16:0_, iso-C_16:0_, C_18:0_ and iso-diabolic acid, the last of which is also common among Acidobacteriota phyla members [56]. The polar lipids diacylglyceryl-hydroxymethyl-N,N,N-trimethyl-beta-alanine (DGTA) and lyso-DGTA were unique to Chloracidobacterium, while phosphatidylethanolamine (PE), phosphatidyl-N-monomethylethanolamine (PME) and phosphatidylcholine (PC) can be found in many other taxa of the class Blastocatellia (Tables 2 and S2). Although phosphatidylglycerol (PG) was previously reported in C. thermophilum strain B [3234], it was not detected in our recent experiments (Table S2). However, this polar lipid was found at trace levels in a few Chloracidobacterium strains.
Phylogenetic and phylogenomic analyses
The methods for DNA isolation, genome sequencing, assembly of circularized chromosomes, annotation, single-value whole-genome or 16S rRNA phylogenetic sequence comparisons were performed as previously described [41]. Fig. 2 shows an inferred maximum-likelihood phylogenetic tree based upon 16S rRNA gene sequences of selected members of Acidobacteriota, including representative strains from all five currently recognized classes of the phylum: Blastocatellia, Terriglobia, Vicinamibacteria, Thermanaerobaculia and Holophagae (Table S3). This phylogenetic tree revealed all Chloracidobacterium spp. form a clade that is most closely related to, but clearly separate and distinct from, other members of the order Blastocatellales within the class Blastocatellia. Although Blastocatellia presently includes two orders, Blastocatellales and ‘Ca. Frugalibacterales’ [2057], where the latter is defined by uncultured organisms ‘GAL08’ found in neutral hot springs [57], Chloracidobacterium spp. clearly separate from both orders [57]. Moreover, Chloracidobacterium spp. strains are not closely related to any member of the three families Pyrimonadaceae, Blastocatellaceae and Arenimicrobiaceae of Blastocatellales (Fig. 2, Tables 3 and S3) [202158]. The most similar members of the class Blastocatellia are merely ~85% identical to the 16S rRNA gene sequence of strain BV2-C^T^ (Fig. 2), which is above the current threshold (78.5%) for class separation as well as for order separation (82%), but below the threshold for family delineation (86.5%) [59]. However, as the threshold for order-level separation (82%) is on the margin, a combined consideration of 16S rRNA, whole-genome phylogenomics (discussed below), as well as single-value whole-genome comparisons suggests Chloracidobacterium should be placed in a separate order. Thus, we propose that the collective Chloracidobacterium spp. represent a new order Chloracidobacteriales, which comprises a single family, Chloracidobacteriaceae (Fig. 2, Table 1).
Inferred maximum-likelihood phylogenetic tree based upon 16S rRNA gene sequences of selected members of the Acidobacteriota. The branches are scaled in terms of the expected number of substitutions per site. The numbers above the branches indicate support values when larger than 60% from maximum-likelihood (left) and maximum parsimony (right) analysis based upon 1,000 bootstrapping replicates. This tree provides strong support for the creation of a new family, Chloracidobacteriaceae (orange-yellow shading), and new order, Chloracidobacteriales, within the class Blastocatellia (blue shading) and within the phylum Acidobacteriota. The five presently recognized classes, Blastocatellia (blue shading), Terriglobia (green shading), Thermoanaerobaculia (brown shading), Holophagae (cyan shading) and Vicinamibacteria (red shading), are shown at the right. The three currently recognized families of the order Blastocatellales are shown by different shading: Blatocatelliaceae (blue), Arenimicrobiaceae (orchid) and Pyrinomonadaceae (magenta). Two strains from the phylum Planctomycetota were used as outgroup. Accession numbers and taxonomic assignments for the organisms included in this tree are provided in Table S3.
The assembled genome of strain BV2-C^T^ was previously deposited in NCBI (GCF_018304825.1) and annotated using the National Center for Biotechnology Information (NCBI) Prokaryotic Genome Annotation Pipeline (https://www.ncbi.nlm.nih.gov/refseq/annotation_prok/). The genome encodes 3,018 proteins and 53 RNAs, including 47 tRNAs, 3 identical rRNA operons (5S, 16S, 23S) and 3 ncRNAs. Due to differences in the annotation software used, this NCBI annotation differs slightly from the previously published annotation done in rapid annotation using subsystem technology [41]. The complete genome of strain BV2-C^T^ consists of two circular chromosomes with lengths of 2,659,040 and 1,000,103 bp, respectively, and shows a total length of 3,659,143 bp with a G+C content of 59.9 mol% (calculated from the sequence), similar to all other strains of the genus (Table 1). These values are comparable to the most similar members of Blastocatellaceae, Pyrimonas methylaliphatogenes K22^T^ and Aridibacter famidurans A22_HD_4H^T^ (genomic information taken from the MAG.1 k127_1028782), which have ~4.0 Mbp genomes with G+C content of 59–66 mol% (Table 1). The calculated average nucleotide identity (ANI) value between strain BV2-C^T^ and C. thermophilum strain B was 75.22%, while digital DNA–DNA hybridization (dDDH) relatedness was 19.10% (Table 3). In addition, ANI values between strain BV2-C^T^ and C. aggregatum strains were 75.3–75.5%, while the dDDH relatedness was 19.1–19.3%. Combined, these ANI and dDDH values fall far below the threshold values for species-level relatedness (95–96% ANI, 70% dDDH) [6061] and together strongly support the proposal of strain BV2-C^T^ as a new species.
Phylogenomic analysis of Acidobacteriota genomes was performed with PhyloPhlAN 3.0 [6263]. Two genomes from the bacterial phylum Planctomycetota were used as an outgroup (Table S4). PhyloPhlAN uses USEARCH [64] to detect 400 universally conserved and ubiquitous proteins, and it aligns them using muscle [65], concatenates the sequences and generates a maximum-likelihood tree with RAxML [66]. Taxonomic assignments and accession numbers for the organisms included in the analysis in Fig. 3 are provided in Table S4. Three metagenome assembled genomes (MAGs) were included in the phylogenomic analysis, which improved the resolution concerning the Chloracidobacterium species. Mat_green_bins.5 (JBDNCY000000000), mat_red_bins.4 (JBDNDF000000000) and mat_yellow_bins.13 (JBDNCT000000000) were reconstructed from samples taken from Rupite hot spring (41° 27′ 29″ N 23° 15′ 43″ E) metagenomes (red SAMN38396676, yellow SAMN38396664 and green SAMN38396669), which have previously been deposited under Bioproject PRJNA1041075. The phylum Acidobacteriota presently comprises five classes and a probable sixth class, ‘Ca. Polarisedimenticolia’ [2067]. A seventh potential class, ‘Ca. Aminicenantia’, might possibly belong to the phylum Acidobacteriota, but these uncultured organisms could also form a candidate phylum; thus, both candidate taxa were not considered in this analysis [206768].
Inferred rooted maximum-likelihood phylogenomic tree of selected members of the phylum Acidobacteriota. The orange-yellow shading indicates the strains for the newly proposed order Chloracidobacteriales and newly proposed family Chloracidobacteriaceae. The Chloracidobacterium spp. strains are assigned to three species, C. validum, ‘Ca. C. aggregatum’ and C. thermophilum, as indicated by the strain names (also see Fig. 2 and text; also see [41]). The blue shading indicates two strains from the order Blastocatellales in the class Blastocatellia (blue shading at right). The other coloured bars at the right indicate strains belonging to the five presently characterized classes within the phylum Acidobacteriota. Two strains from the phylum Planctomycetota were used as outgroup. Accession numbers and taxonomic assignments for the organisms included in this tree are provided in Table S4, and a heat map depiction of the data is shown in Fig. S8
Based upon whole-genome analyses (Fig. 3), the nine isolates of the genus Chloracidobacterium belong to the class Blastocatellia (formerly subdivision 4 [20]). Blastocatellia presently includes the orders Blastocatellales and ‘Ca. Frugalibacterales’ [2058], but like the 16S rRNA analyses described above (Fig. 2), the data in Fig. 3 support Chloracidobacterium spp. to represent a separate monophyletic order and family. The phylogenetic analysis further strongly supports the conclusions based on ANI, dDDH, tetranucleotide frequency and genome synteny [41] that the nine characterized strains define at least three species: C. validum strain BV2-C^T^, C. thermophilum strains D and B and C. aggregatum strains E, N, S, A, 2 and MS40/45. In addition, the phylogenomic tree (Fig. 3) and heat map analyses of the average amino acid identity (AAI) comparisons (Fig. S8) show three MAGs from Rupite hot spring forming a group with C. validum BV2-C^T^. Such additional data support the speciation of C. validum and attribute it as a steady member of the hot spring microbial mats at Rupite, Bulgaria [41]. Although strain MS40/45 was tentatively assigned to C. aggregatum [41], its species assignment remains ambiguous. This strain has a physiology similar to that of C. thermophilum strain B but maintains 16S rRNA sequence identity closer to C. aggregatum strain (Fig. 2, Table 3). The phylogenomic analysis shown in Fig. 3 indicates that this strain is the earliest diverging member of the C. aggregatum and C. thermophilum strains. Collectively, all data suggest strain MS40/45 potentially to be an ecotype or subspecies of C. aggregatum.
Conclusion
We propose that the nine characterized isolates of the genus Chloracidobacterium spp. define a new order, Chloracidobacteriales, and family, Chloracidobacteriaceae, in the class Blastocatellia of the phylum Acidobacteriota. As previously suggested [41], these nine characterized strains can be assigned to three species: ‘Ca. C. thermophilum’, ‘Ca. C. aggregatum’ and C. validum. C. validum strain BV2-C^T^ is further proposed as the new type species and strain of the genus Chloracidobacterium due to the circumstance that the current type species C. thermophilum is only effectively and not validly published [32]. Chloracidobacterium is proposed as the type genus to designate the family Chloracidobacteriaceae and the order Chloracidobacteriales.
Description of Chloracidobacterium gen. nov.
Chlor.a.ci.do.bac.te’ri.um. Gr. masc. adj. chlôros, greenish-yellow, pale green; N.L. neut. n. Acidobacterium, a bacterial genus name; N.L. neut. n. Chloracidobacterium, a green Acidobacterium.
The description is based on that of ‘Chloracidobacterium’ Tank and Bryant 2015 [32]. Microaerophilic, oligotrophic, moderately thermophilic, anoxygenic, chlorophotoheterotrophic eubacterium. Cells stained Gram-negative were non-motile and rods that divide by binary fission. Contain chlorosomes with BChl c as light-harvesting organelles combined with a BChl a-binding Fenna–Matthews–Olson (FMO) protein complex for light energy transfer to homodimeric type-1 reaction centres.
BChl a is found in the reaction centres as BChl aP, Chl aPD and Zn-BChl a P′ as minor Chls [49]. Echinenone, canthaxanthin and γ-carotene are the predominant carotenoids. Cells are unable to reduce sulphate and cannot synthesize branched-chain amino acids but can degrade all three. MK-8(H_2_) and MK-8(H_4_) are their major and minor menaquinones, with iso-C_15:0_, C_16:0_, iso-C_16:0_, C_18:0_ and iso-diabolic acid as primary fatty acids. All genus members have DGTA, lyso-DGTA, PE and PME as their predominant polar lipids. Minor amounts of phosphatidyl-N,N-dimethylethanolamine were also detected, while PG was only present in a few members. The genome comprises two circular chromosomes, and essential genes are found on both. The type species is C. validum.
Description of Chloracidobacterium validum sp. nov.
va’li.dum. L. neut. adj. validum, valid, worthy, referring to Chloracidobacterium validum as the first validly published species name of the genus Chloracidobacterium.
Colonies on solid medium are greenish/brownish and lenticular in shape. Cells in liquid medium predominantly occur as greenish-brown aggregates. Optimal irradiance is ~20–50 µmol photons m^−2^ s^−1^ from a tungsten bulb. The DNA G+C content is 59.5 mol% (by sequence). Growth temperature range is 35–50 °C (T_opt_=45 °C) and the pH range is 5.5–9.5 (pH_opt_=7.2) under micro-oxic conditions. Cells cannot reduce sulphate and require supplementation of organic carbon sources. A mix of the common 20 proteinogenic amino acids can serve as carbon, nitrogen and sulphur sources. Growth is possible with all combinations of amino acids or peptone as long as l-leucine, l-isoleucine, l-valine, l-lysine and a reduced sulphur source (e.g. thioglycolate, methionine or cysteine) are provided. Vitamins (i.e. either vitamin B_12_ or a mixture of other vitamins) are essential for growth.
The type strain BV2-C^T^=DSM 113832^T^=JCM 39534^T^ was isolated from a microbial mat collected in Rupite hot springs, Bulgaria (41° 27′ 28″ N 23° 15′ 43″ E [41]; see Fig. S1 and Strunecký et al. [44] for a more detailed description of Rupite hot spring). The DNA G+C content of strain BV2-C^T^ is 59.5 mol%. The 16S rRNA and whole-genome sequences of strain BV2-C^T^ are available in GenBank/EMBL/DDBJ under OR946236, CP072648 and CP072649.
Description of ‘Candidatus Chloracidobacterium aggregatum’
ag.gre.ga’tum. L. neut. part. adj. aggregatum, aggregated, clustered, as this species forms cell aggregates when grown in liquid medium.
‘Ca. C. aggregatum’ has all general properties of the genus Chloracidobacterium (see above). Colonies on solid medium are greenish-brown in colour and lenticular in shape. Cells in liquid medium predominantly occur as greenish-brown aggregates. Temperature range for growth, ~40–60 °C (optimum ~45–50 °C). Optimal irradiance is ~20–50 µmol photons m^−2^ s^−1^ from a tungsten bulb. Class-Ia ribonucleotide reductase genes (nrdAB) are present, but Class-II, cobalamin-dependent ribonucleotide reductase genes are absent (except strain MS 40/45).
The proposed nomenclatural type is the genome sequence of strain E, represented by GenBank/EMBL/DDBJ accession numbers CP072644 (Chromosome 1) and CP072645 (Chromosome 2). Strain E was isolated from a microbial mat associated with Mushroom Spring in the Lower Geyser Basin of YNP, WY, USA (NPS Scientific Research and Collecting Permit number #YELL-SCI-0129 [3341]). Due to the restriction on its availability and the need for a material transfer agreement (MTA) by the U.S. National Park Service, strain E is considered not suitable for valid publication, nor for pro-valid publication (IJSEM Nomenclature Guidelines, https://www.microbiologyresearch.org/content/journal/ijsem?page=about-journal#3). The DNA G+C content is ~62.1 mol%. The 16S rRNA sequence of strain E is available in GenBank/EMBL/DDBJ under KP300943.
Description of Chloracidobacteriaceae fam. nov.
Chlor.a.ci.do.bac.te.ri.a’ce.ae. N.L. neut. n. Chloracidobacterium, type genus of the family; - aceae, ending to denote family; N.L. fem. pl. n. Chloracidobacteriaceae, the family of the genus Chloracidobacterium.
The family Chloracidobacteriaceae was identified by comparative phylogenetic analysis of 16S rRNA gene sequences and by genomic analyses, which indicated that members of the genus Chloracidobacterium form a separate clade (this study; also see [20323441]). Members of this family are chlorophototrophic, microaerophilic, oligotrophic, moderately thermophilic, Gram-stain-negative, non-motile rods that divide by binary fission. Cells employ chlorosomes, the BChl a-binding FMO protein for harvesting light energy, and homodimeric type-1 reaction centres to carry out conversion of light energy into chemical energy. Cells synthesize BChl c, BChl a and Chl a; major carotenoids are echinenone and γ-carotene, and minor carotenoids include canthaxanthin, deoxyflexixanthin, lycopene and β-carotene. Cells are unable to synthesize vitamin B_12_, l-lysine and branched-chain amino acids (l-valine, l-leucine and l-isoleucine) but can degrade branched-chain amino acids. The type genus is Chloracidobacterium.
Description of Chloracidobacteriales ord. nov.
Chlor.a.ci.do.bac.te.ri.a’les. N.L. neut. n. Chloracidobacterium, type genus of the order; -ales, ending to denote an order; N.L. fem. pl. n. Chloracidobacteriales, the order of the genus Chloracidobacterium.
The order Chloracidobacteriales was identified by comparative phylogenetic analysis of 16S rRNA gene sequences, phylogenomic analyses of whole genomic sequences and physiologically distinct traits, i.e. chlorophototrophy, which indicated that the family forms a separate, deeply divergent clade within the class Blastocatellia (formerly subdivision 4) in the phylum Acidobacteriota. The order Chloracidobacteriales is proposed to include a single family, Chloracidobacteriaceae, with a single genus, Chloracidobacterium, containing three species (‘Ca. C. thermophilum’, ‘Ca. C. aggregatum’ and C. validum). The type genus is Chloracidobacterium.
Emended description of ‘Candidatus Chloracidobacterium thermophilum’ Bryant et al. 2007
The description is based on that of Bryant et al. [28] and Tank and Bryant [32] with the following modifications. Bicarbonate stimulates growth but is not essential. Some strains produce PC. The proposed nomenclatural type is the genome sequence of strain B, represented by GenBank/EMBL/DDBJ accession numbers CP072632 (Chromosome 1) and CP072633 (Chromosome 2). Strain B [32] was isolated from a microbial mat associated with Octopus Spring in the Lower Geyser Basin of YNP, WY, USA (NPS Scientific Research and Collecting Permit number #YELL-SCI-0129 [33, 41]). Due to the restriction on its availability and the need for a material transfer agreement (MTA) by the U.S.National Park Service, strain B is considered not suitable for valid publication, nor for pro-validpublication (IJSEM NomenclatureGuidelines, https://www.microbiologyresearch.org/content/journal/ijsem?page=about-journal# 3). The DNA G+C content is~61.3 mol%.
Strain availability
C. validum strain BV2-C^T^ has been deposited in the open culture collections German Collection for Microorganisms and Cell Cultures and the Japan Collection of Microorganisms (JCM) as DSM 113832 and JCM 39534, respectively. C. thermophilum strain B is available from JCM (JCM 30199) as well as the American Type Culture Collection (ATCC), as ATCC BAA-2647. For JCM 30199, an MTA with YNP must be obtained prior to strain acquisition; in the case of ATCC BAA-2647, a ‘Material Transfer Agreement Addendum for National Park Service Products’ is necessary. C. thermophilum strain D and C. aggregatum strains E, N, S, A, 2 and MS40/45 are available from Dr Vera Thiel upon request under a research MTA from YNP Service, USA.
Supplementary material
10.1099/ijsem.0.007003Uncited Fig. S1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kishimoto N Kosako Y Tano T Acidobacterium capsulatum gen. nov., sp. nov.: an acidophilic chemoorganotrophic bacterium containing menaquinone from acidic mineral environment Curr Microbiol 1991221710.1007/BF 0210620523835745 · doi ↗ · pubmed ↗
- 2Barns SM Takala SL Kuske CR Wide distribution and diversity of members of the bacterial kingdom Acidobacterium in the environment Appl Environ Microbiol 1999651731173710.1128/AEM.65.4.1731-1737.199910103274 PMC 91244 · doi ↗ · pubmed ↗
- 3Zhang Y Cong J Lu H Li G Qu Y et al Community structure and elevational diversity patterns of soil Acidobacteria J Environ Sci 2014261717172410.1016/j.jes.2014.06.01225108728 · doi ↗ · pubmed ↗
- 4Navarrete AA Kuramae EE de Hollander M Pijl AS van Veen JA et al Acidobacterial community responses to agricultural management of soybean in Amazon forest soils FEMS Microbiol Ecol 20138360762110.1111/1574-6941.1201823013447 · doi ↗ · pubmed ↗
- 5Nunes da Rocha U Plugge CM George I van Elsas JD van Overbeek LS The rhizosphere selects for particular groups of Acidobacteria and verrucomicrobia P Lo S One 20138 e 8244310.1371/journal.pone.008244324349285 PMC 3862674 · doi ↗ · pubmed ↗
- 6Lee SH Ka JO Cho JC Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil FEMS Microbiol Lett 200828526326910.1111/j.1574-6968.2008.01232.x 18557943 · doi ↗ · pubmed ↗
- 7O’Connor-Sánchez A Rivera-Domínguez AJ Santos-Briones C de los López-Aguiar LK Peña-Ramírez YJ et al Acidobacteria appear to dominate the microbiome of two sympatric Caribbean Sponges and one Zoanthid Biol Res 2014476710.1186/0717-6287-47-6725723107 PMC 4335776 · doi ↗ · pubmed ↗
- 8Quaiser A López-García P Zivanovic Y Henn MR Rodriguez-Valera F et al Comparative analysis of genome fragments of Acidobacteria from deep Mediterranean plankton Environ Microbiol 2008102704271710.1111/j.1462-2920.2008.01691.x 18627413 · doi ↗ · pubmed ↗