Septic Shock Caused by Coinfection of Shewanella algae Bloodstream Infection and Epstein‐Barr Virus: Clinical Characteristics and Genomic Analysis
Jianmei Chen, Dong Ling, Feng Wang, Liping Liu, Yucheng Ren, Chengying Chen, Na Su

TL;DR
A case of septic shock in inland China is caused by a coinfection of Shewanella algae and Epstein-Barr virus, with genomic analysis revealing freshwater adaptation and immune system impact.
Contribution
The study reveals freshwater adaptation of Shewanella algae and the synergistic pathogenic role of EBV coinfection in causing septic shock.
Findings
The S. algae strain showed freshwater adaptation through unique genes for transporters and carbohydrate enzymes.
EBV coinfection accelerated septic shock via immunosuppression and inflammation.
The patient succumbed to multi-organ failure from the coinfection.
Abstract
Shewanella algae, a marine‐origin opportunistic pathogen, has shown a significant increase in non‐coastal infections, yet its environmental adaptability and synergistic pathogenic mechanisms with Epstein‐Barr virus (EBV) coinfection remain unclear. This study reports a clinical case of S. algae bloodstream infection complicated by EBV reactivation leading to septic shock in Sichuan Province, China, and elucidates the molecular mechanisms through genomic analysis. Pathogen identification was performed via blood culture, antibiotic susceptibility testing, and genomic annotation. The strain harbored resistance genes (acrB, tolC, tet(35), golS) and virulence factors (bplL/bplF, clpC/clpP, tonB). Phylogenetic analysis indicated the highest genetic affinity to freshwater‐derived Shewanella chilikensis, while pan‐genome analysis identified 1412 unique genes, including transmembrane…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
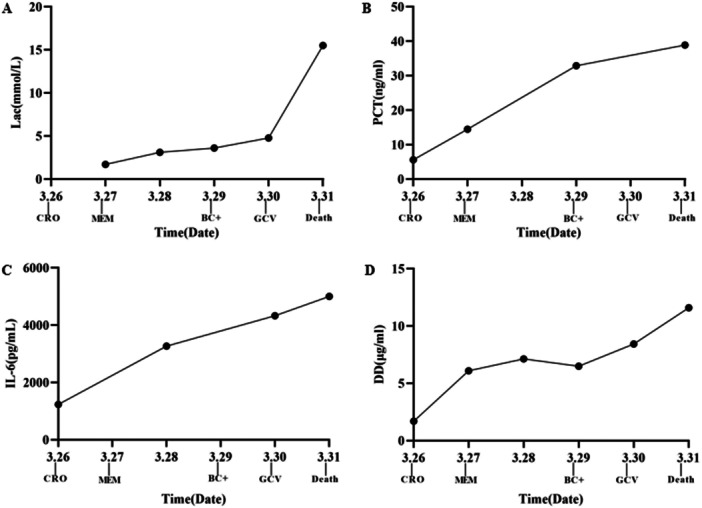 Figure 1
Figure 1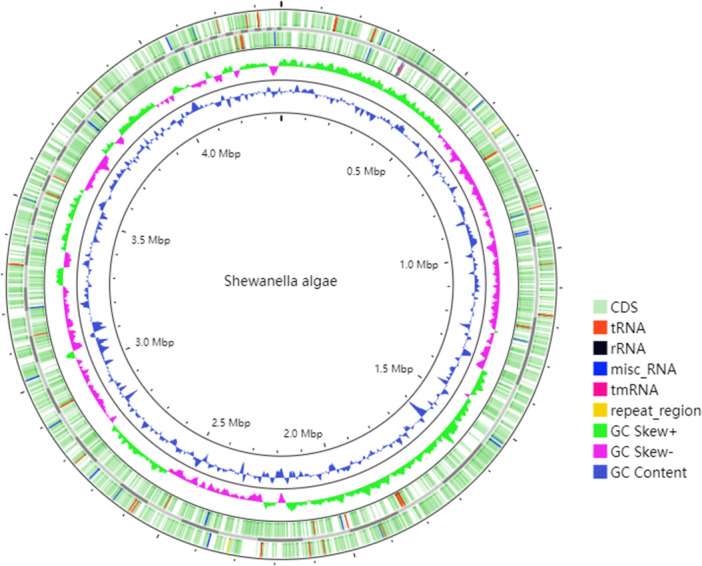 Figure 2
Figure 2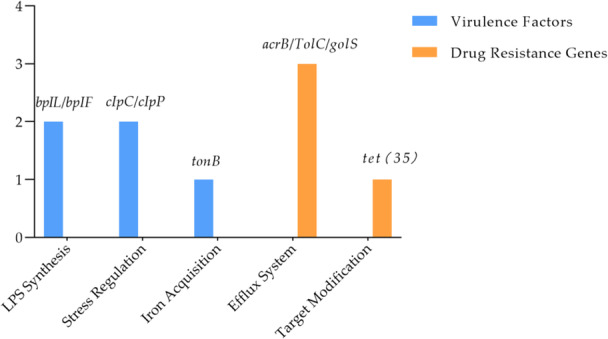 Figure 3
Figure 3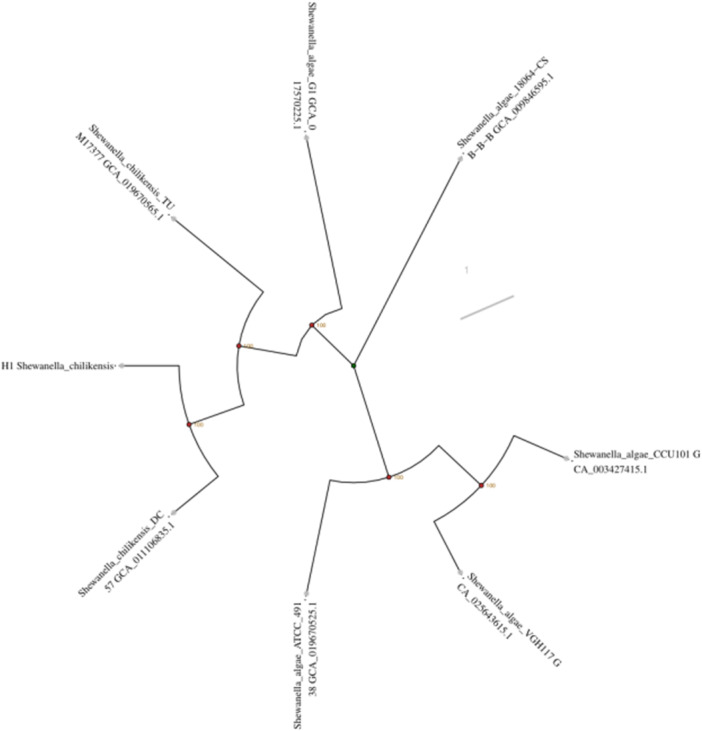 Figure 4
Figure 4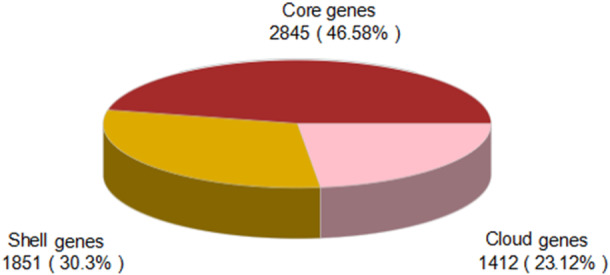 Figure 5
Figure 5| Antibiotic class | Antibiotic name | MIC (μg/mL) | Interpretation (S/R) |
|---|---|---|---|
| β‐lactams | Ceftriaxone | 64 | R |
| β‐lactam/β‐lactamase inhibitor | Ampicillin/Sulbactam | 2 | S |
| Cefoperazone/Sulbactam | 4 | S | |
| Quinolones | Ciprofloxacin | 4 | R |
| Levofloxacin | 8 | R | |
| Sulfonamides + Trimethoprim | Trimethoprim/Sulfamethoxazole | 0.5 | S |
| Aminoglycosides | Amikacin | 64 | R |
| Carbapenems | Meropenem | 0.25 | S |
| Imipenem | 0.25 | S | |
| Monobactams | Aztreonam | 32 | R |
| Tetracyclines | Minocycline | 0.25 | S |
- —Chengdu Medical Research Project
- —Youth Innovation Project of Sichuan Medical Association
- —Youth Innovation Medical Research Project of Chongzhou People's Hospital
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Marine Bivalve and Aquaculture Studies · Bacteriophages and microbial interactions
Introduction
1
Shewanella algae is a halophilic gram‐negative opportunistic pathogen traditionally associated with marine environments. Human infections are commonly linked to seawater exposure or immunocompromised states (Masmoudi et al. 2023; Ibrahim et al. 2021; Symanzik et al. 2022). In recent years, however, cases of infection in non‐coastal regions have been increasingly reported (Fernandes et al. 2023; Rodriguez‐Vargas et al. 2022; Weiss et al. 2021), suggesting its potential environmental adaptive evolution to occupy new ecological niches. Existing literature has documented mixed infections of S. algae with pathogens such as Morganella morganii, yet clinical studies on its coinfection with viruses, particularly the synergistic mechanisms in sepsis progression, remain unexplored (Huang et al. 2021; Martins Sousa et al. 2022). Notably, S. algae bloodstream infections carry a high mortality rate (35%–40%) (Ainoda et al. 2022) and often exhibit multidrug resistance (Wang et al. 2024; Huang et al. 2022), posing significant challenges to clinical management.
Epstein‐Barr virus (EBV), a ubiquitous human herpesvirus, can reactivate in immunocompromised patients (Houen and Trier 2020; Murata et al. 2021; Wei et al. 2023) and exacerbate inflammatory cascades by modulating host immune responses (Verbist and Nichols 2024; Xu et al. 2021; Ureshino et al. 2018). Studies indicate that EBV suppresses hepcidin expression (Mei et al. 2023), increasing serum free iron levels, while S. algae relies on iron acquisition systems (e.g., tonB gene) to hijack host iron resources (Liu et al. 2022; Fang et al. 2022). These interactions may form a metabolic symbiosis, further compromising host defenses. However, the molecular mechanisms underlying such cross‐species interactions in sepsis remain poorly understood.
This study presents the first clinical case of S. algae bloodstream infection co‐occurring with EBV reactivation in Sichuan Province, China. By integrating whole‐genome sequencing to dissect the strain's resistance profile, virulence factors, and evolutionary traits, we aim to provide empirical evidence for early diagnosis and treatment of inland S. algae infections. Our findings also highlight freshwater contamination as a potential novel transmission route, urging heightened vigilance in non‐coastal regions.
Materials and Methods
2
Case Report
2.1
An 80‐year‐old male patient was admitted on March 26, 2025, with complaints of “recurrent dizziness, palpitations, and dyspnea for 1 year, aggravated for 3 days.” His medical history included poorly controlled type 2 diabetes mellitus, megaloblastic anemia, and chronic cardiac insufficiency (preserved ejection fraction heart failure, cardiac function class III). The patient resided in a rural area and consumed unfiltered well water (prone to sewage contamination during rainy seasons), with no history of seawater or seafood exposure. Physical examination revealed high fever (39.4°C), anemic pallor (hemoglobin 37 g/L), and bilateral pitting edema. Laboratory tests showed elevated inflammatory markers (procalcitonin PCT 5.6 ng/mL, interleukin‐6 IL‐6 1232.1 pg/mL) and abnormal coagulation (D‐dimer 1.69 μg/mL). Chest CT indicated right pleural effusion with pulmonary atelectasis.
Blood culture turned positive after 26 h post‐admission, and the pathogen was identified as S. algae (strain H1, where H denotes host origin and 1 represents the strain number) using the VITEK 2 Compact system. Broth microdilution antimicrobial susceptibility testing revealed sensitivity to meropenem but resistance to ceftriaxone and ciprofloxacin (Table 1). Metagenomic next‐generation sequencing (mNGS) detected a high EBV DNA load (1.2 × 104 copies/mL; reference range: < 5 × 102 copies/mL). Serological tests confirmed EBV reactivation (EBV VCA‐IgM +, EA‐IgG ++ [Masmoudi et al. 2023]). Initial treatment included ceftriaxone (4 g QD), but the patient progressed to septic shock with multiple organ failure (acute liver failure, coagulopathy) within 72 h. Therapy was escalated to meropenem (1 g Q8H) combined with ganciclovir, alongside mechanical ventilation and closed thoracic drainage (800 mL bloody fluid evacuated).
The patient's condition deteriorated rapidly (Figure 1): PCT surged from 5.6 ng/mL (March 26) to 38.9 ng/mL (March 31), IL‐6 spiked to 5000 pg/mL, and organ failure progressed (peak ALT 842 U/L, creatinine 214.5 μmol/L, D‐dimer 11.6 μg/mL, hemoglobin 26 g/L). On March 31, the patient experienced cardiac arrest. Cardiopulmonary resuscitation restored transient spontaneous circulation, but irreversible multi‐organ failure led to clinical death at 18:26 after family withdrawal of life support.
Dynamic changes in key laboratory parameters (March 26–31) and clinical interventions. (A) Lactate (Lac, mmol/L). (B) Procalcitonin (PCT, ng/mL). (C) Interleukin‐6 (IL‐6, pg/mL). (D) D‐dimer (DD, μg/mL). Labels on the X‐axis indicate the timing of key clinical events: CRO (ceftriaxone), BC+ (blood culture positive), MEM (meropenem), GCV (ganciclovir), and death.
Bacterial Pathogen Detection
2.2
Peripheral blood specimens were collected at admission and inoculated into BactAlert 3D aerobic and anaerobic blood culture bottles. Positive cultures were isolated and purified, followed by identification of the pathogen as S. algae using the VITEK 2 Compact system. For both the identification procedure and the antimicrobial susceptibility testing, Pseudomonas aeruginosa ATCC 27853 was used as the quality control strain. AST was performed using the broth microdilution method in accordance with the Clinical and Laboratory Standards Institute (CLSI) guidelines (M100, 2024). The minimum inhibitory concentration (MIC) values were determined for the antibiotics listed in Table 1. Due to the absence of specific CLSI breakpoints for S. algae, the MIC values were interpreted based on the criteria established for P. aeruginosa for the majority of antibiotics. However, for ceftriaxone, ampicillin/sulbactam, trimethoprim/sulfamethoxazole, and minocycline, for which P. aeruginosa breakpoints are not provided by CLSI, the interpretive criteria for Escherichia coli were applied.
Viral Pathogen Detection
2.3
Two milliliters of EDTA‐anticoagulated blood were collected from the patient. Plasma cell‐free DNA was extracted using the Plasma Free DNA Extraction Kit (TIANGEN DP338). After DNA library construction, sequencing was performed on the Illumina NextSeq. 550 platform. Raw sequencing data underwent quality control using FastQC (version 0.11.9) to remove low‐quality reads and adapter contamination. Clean reads were aligned to the NCBI NT/NR database (2025 version) for pathogen identification. EBV viral load was quantified by normalizing the number of mapped reads to copies per milliliter (copies/mL) using a standardized bioinformatics pipeline.
Genomic Sequencing and Bioinformatics Analysis
2.4
Following bacterial pure culture, genomic DNA was extracted using the TIANGEN DP338 DNA Extraction Kit. DNA concentration was quantified using Qubit 4.0 (≧20 ng/μL), and quality was assessed via agarose gel electrophoresis. Paired‐end sequencing (150 base pairs bp read length, total data volume 1.69 × 109 bp) was performed on the Illumina NextSeq. 550 platform. Raw sequencing data were processed with Trimmomatic v0.36 for quality control, including removal of low‐quality sequences and adapter contamination using a sliding window approach (Q20≧97.5%).
Genome assembly was conducted using SPAdes v3.5.0 to generate contigs. Gene annotation, including protein‐coding genes, rRNA, and tRNA, was performed using Prokka v1.10. Antibiotic resistance genes were annotated against the Comprehensive Antibiotic Resistance Database (CARD, https://card.mcmaster.ca/), while virulence factors were screened using the Virulence Factor Database (VFDB, http://www.mgc.ac.cn/VFs/).
Pan‐Genome and Functional Evolutionary Analysis
2.5
Pan‐genome analysis of the isolated strain (H1) and 30 closely related Shewanella strains from the NCBI RefSeq database was performed using Roary v3.13.0. Gene clusters were categorized as follows: core genes (shared by all strains), accessory genes (shared by≧2 strains), and unique genes (exclusive to H1). Metabolic functions of unique genes were annotated by aligning Prokka‐predicted protein sequences against the CAZy database (Carbohydrate‐Active Enzymes, http://www.cazy.org/) and the TCDB database (Transporter Classification Database, http://www.tcdb.org/) using HMMER3 v3.1b1. Key annotations included AA2 manganese peroxidases and ion transporters.
Phylogenetic analysis was conducted by aligning core single‐copy genes with MAFFT, followed by tree construction via FastTree using the neighbor‐joining method (Bootstrap values = 1000). Functional annotations were integrated to elucidate the freshwater adaptation mechanisms of the strain.
Results
3
Pathogen Detection Results
3.1
Blood culture turned positive after 26 h of incubation, with the pathogen identified as S. algae. mNGS detected a high EBV DNA load of 1.2 × 104 copies/mL (reference value: < 5 × 102 copies/mL). Serological tests confirmed EBV reactivation, with positive EBV VCA‐IgM (+) and elevated EA‐IgG (++).
Genomic Characteristics
3.2
The genome of the S. algae isolate (H1) had a total length of 4,445,496 base pairs (bp) with a GC content of 52.03%. The assembly comprised 90 contigs, with an N50 of 123,101 bp and a maximum contig length of 326,936 bp (Figure 2). Gene prediction revealed that coding regions accounted for 87.12% of the genome, encompassing 3948 protein‐coding genes, 6 rRNA genes, and 92 tRNA genes. Non‐coding regions were enriched with regulatory sequences and transposable elements.
Circular genome map of Shewanella algae (H1). (Outer to inner rings: Distribution of coding regions and non‐coding RNAs, repetitive sequences, GC Skew [± values indicating the direction of replication strands], and dynamic GC content [10‐kb sliding window].)
Analysis of Resistance Gene Profile
3.3
Annotation via the CARD database revealed four resistance‐associated genes in the S. algae (Figure 3):Efflux pump system genes: acrB(antibiotic efflux) and TolC(efflux channel protein), suggesting potential resistance to quinolones through active efflux mechanisms. Tetracycline resistance gene: tet(35)(ribosomal protection protein), likely mediating tetracycline resistance via target modification. Metal ion tolerance gene: golS, potentially involved in heavy metal resistance. AST demonstrated sensitivity to meropenem but resistance to quinolones (ciprofloxacin, levofloxacin) and ceftriaxone.
Antibiotic resistance genes (acrB, tolC, tet(35), golS) and virulence factors (bplL/bplF, clpC/clpP, tonB) identified in Shewanella algae strain H1 via CARD and VFDB. Bar heights reflect relative gene abundance; colors distinguish resistance and virulence categories.
Virulence Factor Characteristics
3.4
Annotation via the VFDB database revealed that the strain harbored key virulence genes, including: Lipopolysaccharide synthesis‐related genes bplL and bplF, which may enhance bacterial endotoxin activity to promote host inflammatory responses. Stress response regulatory genes clpC/clpP (ATP‐dependent protease), involved in bacterial tolerance and survival within the host intracellular environment. Iron acquisition system gene tonB (siderophore transporter protein), hypothesized to exacerbate immunosuppression by competitively hijacking host iron resources (Figure 3).
Phylogenetic and Evolutionary Analysis
3.5
Phylogenetic analysis based on 16S rRNA and core single‐copy genes revealed that the strain clustered with the freshwater‐derived Shewanella chilikensis (GCA_011106835.1) with the highest genetic affinity (Bootstrap value = 1000) (Figure 4). Pan‐genome analysis (including 30 closely related Shewanella strains) indicated that the H1 strain harbored 2845 core genes (46.58%) and 1412 unique genes (23.12%). Functional annotation of the unique genes revealed enrichment in transmembrane transporters (10.61% of TCDB‐annotated genes) and carbohydrate‐active enzymes (0.94% of CAZy‐annotated genes) (Figure 5), suggesting freshwater adaptive evolution through metabolic features such as transmembrane transporter gene enrichment.
Phylogenetic tree constructed using core single‐copy genes (aligned with MAFFT and built via the neighbor‐joining method in FastTree, Bootstrap value = 1000). The H1 strain (red label) exhibits the closest genetic affinity to the freshwater‐derived Shewanella chilikensis (GCA_011106835.1).
Pan‐genome composition analysis of Shewanella algae (H1).
Discussion
4
In this case, the patient had no history of seawater exposure, and the isolated strain (H1) exhibited the closest phylogenetic affinity to the freshwater‐derived S. chilikensis. Combined with the patient's long‐term consumption of contaminated well water, these findings suggest that S. algae may have colonized inland water systems through genomic adaptive evolution (Martín‐Rodríguez et al. 2022; Fu et al. 2024; Chen et al. 2020). Pan‐genome analysis revealed that the H1 strain harbored 1412 unique genes (23.12%) (Kim et al. 2020; Rosconi et al. 2022), enriched in transmembrane transporter genes (e.g.,tonB) and carbohydrate‐active enzyme (CAZy) genes. The tonB gene, encoding a siderophore transporter protein, may enhance bacterial tolerance to low‐salinity environments (Akinbosede et al. 2022; Pollet et al. 2021; Yong et al. 2023), while CAZy genes (e.g., AA2 manganese peroxidases) likely contribute to the degradation of complex organic matter in freshwater habitats (Nguyen et al. 2025; Vela Gurovic et al. 2021; Sun et al. 2023). These molecular insights provide evidence for the potential transmission of S. algae via agriculturally polluted inland water sources (Blanco and Díaz de Tuesta 2021).
Although AST indicated sensitivity to meropenem, the patient's treatment ultimately failed, suggesting that phenotypic results may underestimate the risk of cryptic resistance mechanisms (Mehrotra et al. 2023; Zhou et al. 2022). Genomic analysis revealed that efflux pump genes (acrB and tolC) mediate resistance to quinolones (e.g., ciprofloxacin) through active drug efflux (Dhara and Tripathi 2024; Albarri et al. 2022; Khoshnood et al. 2021). Notably, such efflux systems may exhibit low‐level activity against carbapenems (e.g., meropenem) (Lian et al. 2024; Tian et al. 2021), reducing intracellular drug concentrations and compromising clinical efficacy. Ceftriaxone resistance may arise from synergistic effects between penicillin‐binding protein (PBP) mutations and efflux pumps (Fischer et al. 2020; Sethuvel et al. 2023). Additionally, the metal ion tolerance gene golS might indirectly influence antibiotic susceptibility by regulating heavy metal resistance (Hasegawa et al. 2024). Severe immunosuppression caused by concurrent EBV reactivation likely further hindered bacterial clearance by antibiotics (Blot et al. 2022). These multifactorial interactions emphasize the need for genomic‐guided optimization of treatment regimens, such as administering high‐dose meropenem (1 g every 8 h) combined with efflux pump inhibitors (e.g., cilastatin).
The virulence genes carried by the strain exacerbate host pathological damage through multiple mechanisms. The lipopolysaccharide synthesis genes bplL and bplF significantly enhance bacterial endotoxin activity, activating the host TLR4/NF‐κB pathway and triggering a surge in inflammatory cytokines such as IL‐6 (Dong et al. 2020; Guo et al. 2024; Ali et al. 2024), directly leading to microcirculatory dysfunction. The stress response regulatory genes clpC and clpP, encoding an ATP‐dependent protease system, confer bacterial resistance to host oxidative stress, enabling survival within macrophage phagosomes and persistent release of toxic factors (Wen et al. 2025; Biswas et al. 2021). The iron acquisition gene tonB exacerbates anemia and suppresses neutrophil extracellular trap formation by competitively hijacking host free iron through the secretion of high‐affinity siderophores (Kumar et al. 2024). The dynamic elevation of lactate levels further corroborates the severity of microcirculatory impairment and tissue hypoxia (Yang et al. 2023, 2022). These metabolic disturbances synergize with bacterial endotoxins and EBV‐induced inflammatory storms, accelerating multi‐organ failure.
EBV reactivation reshapes the host immune response through multiple pathways. The EBV‐encoded latent membrane protein LMP1 downregulates the expression of host MHC class I molecules, impairing CD8 + T cell recognition efficiency of antigen presentation (Choi et al. 2021; Dudaniec et al. 2021), while simultaneously inhibiting the IFN‐γ signaling pathway, leading to exhaustion of specific immune responses. The EBV latent protein LMP2A activates the non‐canonical NF‐κB pathway, synergistically amplifying the secretion of pro‐inflammatory cytokines such as IL‐6 and TNF‐α with bacterial endotoxins, thereby forming an “inflammatory storm” that accelerates multi‐organ damage (Madayag et al. 2022; Yifei et al. 2024). Notably, the EBV‐encoded EBNA‐3C protein suppresses hepcidin expression, increasing serum free iron levels (Cortese et al. 2024). This establishes a metabolic mutualism network with the bacterial tonB‐mediated iron acquisition system, exacerbating anemia and immunosuppression (Li et al. 2021; Kloepfer and Kennedy 2023; Chevallereau et al. 2022).
Clinical Implications: Inland patients with unexplained sepsis, particularly those with comorbidities such as diabetes or anemia, should be vigilant against S. algae infection (Song et al. 2021). Early combination of blood culture, mNGS, and resistance gene detection is recommended (Sun et al. 2022). For treatment, carbapenems should be administered at full doses, and efflux pump inhibitors (e.g., cilastatin) may be added if necessary (Zack et al. 2024). In patients with EBV reactivation, combination therapy with ganciclovir and IL‐6 antagonists (e.g., tocilizumab) may be considered to block the inflammatory cascade.
Conclusions
5
Shewanella algae can cause infections in non‐coastal regions through freshwater adaptive evolution (e.g., enrichment of transmembrane transporters and CAZy genes). EBV coinfection accelerates septic shock progression via immune‐metabolic interactions. Clinically, pathogen surveillance in inland areas should be strengthened, and treatment strategies should be optimized through early integration of genomics and mNGS.
Author Contributions
Jianmei Chen: conceptualization, methodology, validation, formal analysis, resources, data curation, writing – original draft, writing – review and editing, visualization, funding acquisition. Dong Ling: conceptualization, methodology, resources, writing – original draft. Feng Wang: conceptualization, supervision. Liping Liu: conceptualization, supervision. Yucheng Ren: methodology, project administration. Chengying Chen: methodology, resources, writing – review and editing, project administration. Na Su: validation, resources, funding acquisition.
Ethics Statement
The authors have nothing to report.
Consent
Written informed consent for publication was obtained from the patient's family.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ainoda, Y. , E. Tanaka , T. Wajima , et al. 2022. “A Case of Shewanella algae‐Induced Bacteremia in Japan: Case Report and Literature Review.” Journal of Infection and Chemotherapy 28: 1430–1432. 10.1016/j.jiac.2022.06.015.35777628 · doi ↗ · pubmed ↗
- 2Akinbosede, D. , R. Chizea , and S. A. Hare . 2022. “Pirates of the Haemoglobin.” Microbial Cell 9: 84–102. 10.15698/mic 2022.04.775.35434122 PMC 8977872 · doi ↗ · pubmed ↗
- 3Albarri, O. , M. Al Matar , M. M. Öcal , and F. Köksal . 2022. “Overexpression of Efflux Pumps Acr AB and Oqx AB Contributes to Ciprofloxacin Resistance in Clinical Isolates of K. pneumonia .” Current Protein & Peptide Science 23: 356–368. 10.2174/1389203723666220630162920.35786184 · doi ↗ · pubmed ↗
- 4Ali, W. , K. Choe , J. S. Park , et al. 2024. “Kojic Acid Reverses LPS‐Induced Neuroinflammation and Cognitive Impairment by Regulating the TLR 4/NF‐κB Signaling Pathway.” Frontiers in Pharmacology 15: 1443552. 10.3389/fphar.2024.1443552.39185307 PMC 11341365 · doi ↗ · pubmed ↗
- 5Biswas, S. , H. P. S. Dhaked , A. Keightley , and I. Biswas . 2021. “Involvement of Clp E AT Pase in Physiology of Streptococcus mutans .” Microbiology Spectrum 9: e 0163021. 10.1128/Spectrum.01630-21.34851151 PMC 8635124 · doi ↗ · pubmed ↗
- 6Blanco, G. , and J. A. Díaz de Tuesta . 2021. “Seasonal and Spatial Occurrence of Zoonotic Salmonella Serotypes in Griffon Vultures at Farmland Environments: Implications in Pathogen Pollution and Ecosystem Services and Disservices.” Science of the Total Environment 758: 143681. 10.1016/j.scitotenv.2020.143681.33250252 · doi ↗ · pubmed ↗
- 7Blot, S. , E. Ruppé , S. Harbarth , et al. 2022. “Healthcare‐Associated Infections in Adult Intensive Care Unit Patients: Changes in Epidemiology, Diagnosis, Prevention and Contributions of New Technologies.” Intensive and Critical Care Nursing 70: 103227. 10.1016/j.iccn.2022.103227.35249794 PMC 8892223 · doi ↗ · pubmed ↗
- 8Chen, Y. J. , G. C. He , J. F. Cheng , et al. 2020. “Comparative Genomics Reveals Insights Into Characterization and Distribution of Quorum Sensing‐Related Genes in Shewanella algae From Marine Environment and Clinical Sources.” Comparative Immunology, Microbiology and Infectious Diseases 73: 101545. 10.1016/j.cimid.2020.101545.32927298 · doi ↗ · pubmed ↗