Impact of virulence factors overexpression on Listeria monocytogenes F2365 epidemic strain fitness and the limitations of surrogate species in UHT and raw milk
Alba Espí-Malillos, Inmaculada López-Almela, Pilar Ruiz-García, María Carmen López-Mendoza, Nerea Carrón, Pedro González-Torres, Jazmin Meza-Torres, Javier Pizarro-Cerdá, Juan J. Quereda

TL;DR
The study shows that common Listeria surrogates do not accurately model the behavior of a hypervirulent Listeria monocytogenes strain in milk products.
Contribution
The research evaluates the suitability of Listeria surrogates and the impact of virulence factor overexpression in dairy environments.
Findings
PrfA overexpression reduced L. monocytogenes fitness in UHT and raw milk.
Surrogate Listeria species failed to replicate wild-type growth patterns.
Raw milk microbiota was unaffected by Listeria presence.
Abstract
Identifying reliable surrogates for Listeria monocytogenes is critical to safely model its behavior in dairy environments. Many dairy-related listeriosis outbreaks are linked to unpasteurized milk products enriched in hypervirulent L. monocytogenes from lineage I. We used an outbreak-associated lineage I strain to evaluate Listeria innocua, Listeria valentina, and Listeria ivanovii as potential surrogates in UHT and raw milk at 4 °C. We also assessed how overexpression of key virulence factors, including LIPI-3 and the PrfA regulon, influenced growth. While LIPI-3 overexpression had no significant impact, PrfA overexpression reduced fitness in both matrices. None of the other Listeria species tested accurately replicated the L. monocytogenes wild type strain growth patterns. Additionally, the native raw milk microbiota remained largely unaffected by the presence of any Listeria strain.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
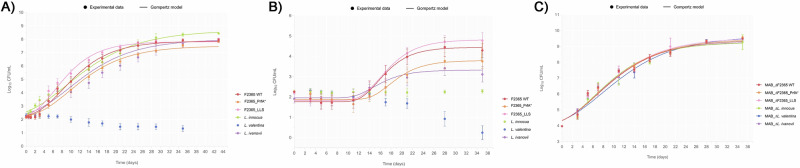 Figure 1
Figure 1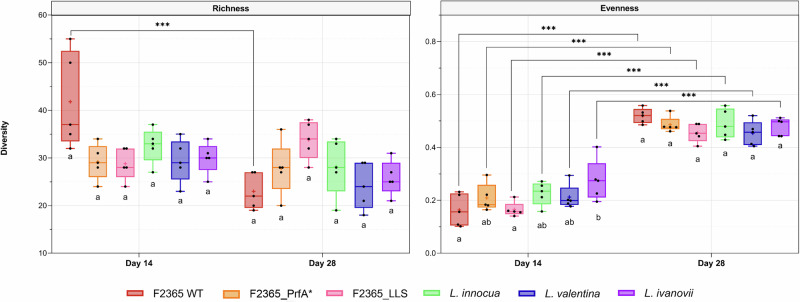 Figure 2
Figure 2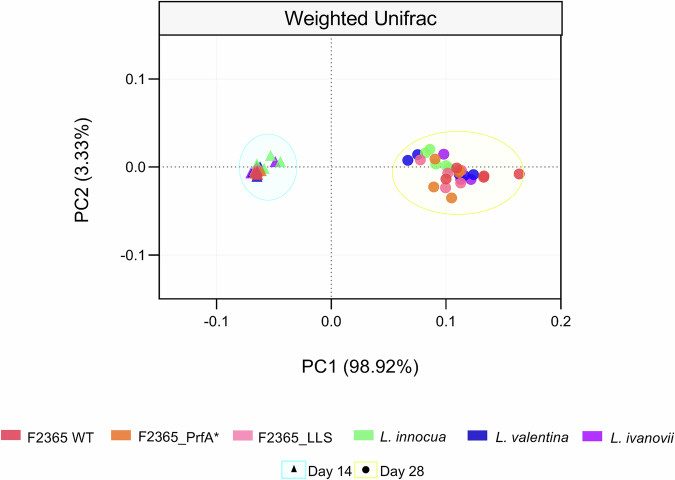 Figure 3
Figure 3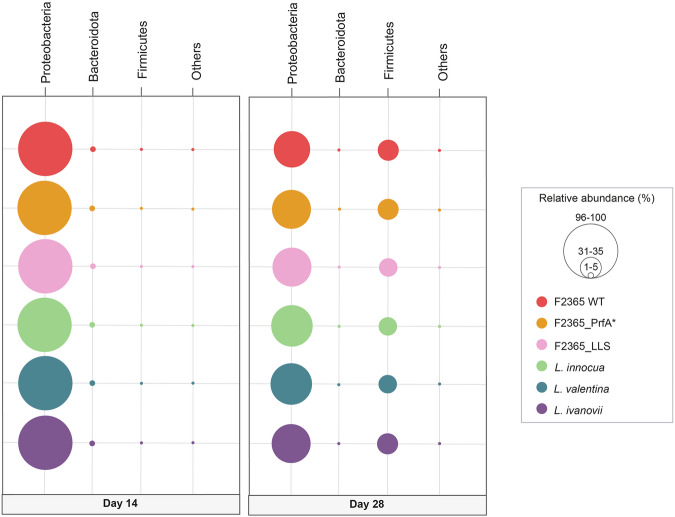 Figure 4
Figure 4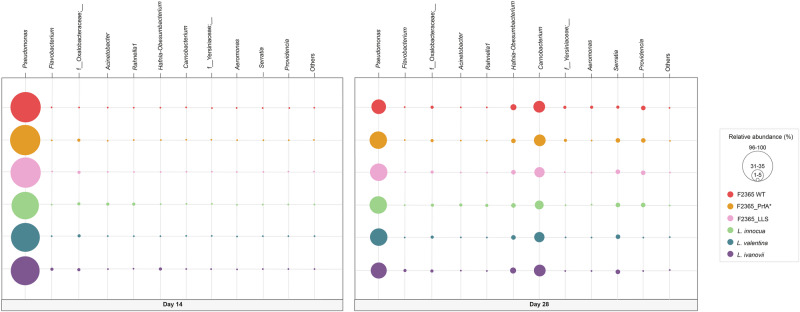 Figure 5
Figure 5- —https://doi.org/10.13039/501100003359Generalitat Valenciana
- —https://doi.org/10.13039/100014440Ministerio de Ciencia, Innovación y Universidades
- —https://doi.org/10.13039/100031289Universidad CEU Cardenal Herrera
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsListeria monocytogenes in Food Safety · Salmonella and Campylobacter epidemiology · Mycotoxins in Agriculture and Food
Introduction
The genus Listeria is composed of 29 species^1^. Only two Listeria spp. are considered pathogenic within the genus: L. monocytogenes and L. ivanovii^2^. L. ivanovii affects ruminants and rarely causes infections in humans^3^, whilst L. monocytogenes causes illness in humans and other animals, mainly domestic ruminants^4^. Listeriosis accounted for the highest case fatality rate (19.7%) among all reported foodborne diseases^5^.
The most studied species is L. monocytogenes, a ubiquitous, saprophytic, and psychrophilic bacterium that can survive and grow in different environments, including water, soil, sewage, vegetation, animal feeds, farms, food products, and food-processing settings^4^. It is also a facultative intracellular pathogen that causes foodborne listeriosis^6^. After the ingestion of contaminated food, L. monocytogenes can cross the intestinal barrier, disseminate via the bloodstream, and reach the central nervous system and fetus, causing septicemia, meningoencephalitis, and abortion^6,7^.
L. monocytogenes can be divided into four evolutionary lineages and more than 170 clonal complexes (CCs)^8^. While lineage I isolates, in particular CC1, CC4, and CC6, are hypervirulent and associated with human and animal clinical cases, lineage II, in particular CC9 and CC121, is mainly associated with food and food processing environments^9^. Importantly, epidemiological studies have shown that hypovirulent isolates of CC9 and CC121 are strongly associated with meat products, whereas hypervirulent CC1, CC4, and CC6 are associated with dairy products^10^. Moreover, dairy products are involved with nearly half of the registered listeriosis outbreaks in the United States and Europe^11–13^.
L. monocytogenes virulence depends mainly on Listeria pathogenicity islands (LIPIs) LIPI-1, LIPI-3, LIPI-4, and the internalin A and B loci^8^. PrfA is the major virulence regulator in L. monocytogenes and controls the internalin A and B loci for attachment to and invasion of the intestinal epithelium and LIPI-1, which is required for intracellular growth and transmission from cell to cell^4,8^. Although not common, there are atypical L. monocytogenes strains that display a PrfA* mutation leading to constitutive production of LIPI-1 virulence genes (e.g. the lineage II EGD strain)^14,15^. The internalin A and B locus and LIPI-1 genes are present in all L. monocytogenes strains^8^. However, LIPI-3 (which encodes the bacteriocin listeriolysin S (LLS) required for intestinal colonization) and LIPI-4 (involved in brain and placental invasion) are only present in some hypervirulent lineage I clonal complexes^9,16^.
The saprophytic/extracellular and the infective/intracellular facets of L. monocytogenes are complementary and have frequently been studied independently. However, there are cases such as ActA, a virulence factor encoded in LIPI-1 and regulated by PrfA, that plays a critical role during infection and the extracellular lifestyle of L. monocytogenes^17^. During the intracellular cycle, ActA contributes to L. monocytogenes actin-based motility, cell-to-cell spread, and dissemination within host tissues^17^. ActA also acts extracellularly, mediating L. monocytogenes aggregation and biofilm formation^17^. The consequences of the contamination of a dairy product with a hypervirulent strain of L. monocytogenes with high PrfA or LIPI-3 activity are currently unknown. This lack of knowledge led us to investigate the role that PrfA-controlled genes and LIPI-3 could play in food contamination and growth.
Understanding how L. monocytogenes grows and survives in food is essential for proposing practical strategies that prevent its transmission. Food industries try to identify possible scenarios that represent good indicators of conditions in which L. monocytogenes can thrive. The identification of bacteria belonging to the genus Listeria has been used as an indicator to detect conditions that allow for the presence, growth, and persistence of L. monocytogenes^18^. Due to L. monocytogenes pathogenicity, modeling and understanding its behavior in foods and food processing environments could require using a surrogate organism. L. innocua is a closely related, generally non-pathogenic species of the sensu stricto group traditionally used as a surrogate organism to better understand the behavior of L. monocytogenes in different environments^19^. The use of L. innocua to predict the behavior of L. monocytogenes in agricultural and food processing settings has historically been justified by the two species’ ecological cohabitation, genomic closeness, and physiological resemblance^20^. L. monocytogenes, L. innocua, and L. ivanovii belong to the Listeria sensu stricto clade, and therefore share phylogenetic proximity and ecological traits^21,22^. These three species are the most well characterized among the Listeria genus^4,22^. Despite the genetic closeness of these three species, the use of L. ivanovii and L. innocua as surrogates in dairy products lacks extensive experimental evidence*. L. valentina* belongs to the Listeria sensu lato clade and was recently described in a dairy farm^23^. However, the use of L. valentina as a surrogate has never been tested before.
Here, using the lineage I strain F2365 (responsible for the 1985 California outbreak related to Mexican fresh cheese, one of the deadliest bacterial foodborne outbreaks ever reported in the United States^11^ we first analyzed the reliability of L. innocua, L. valentina, or L. ivanovii as surrogates for the pathogenic L. monocytogenes to monitor growth in Ultra-High-Temperature (UHT) and raw milk at refrigeration temperatures. Secondly, we determined the extent to which virulence factors such as the bacteriocin LLS encoded in LIPI-3 or those virulence factors controlled by PrfA contribute to L. monocytogenes growth in UHT and raw milk at refrigeration temperatures. Finally, we analyzed the impact of milk contamination with L. innocua, L. valentina, L. ivanovii, L. monocytogenes F2365 or its isogenic mutants overexpressing PrfA and LIPI-3 on the growth and composition of the native raw milk microbiota by using growth kinetics studies and metagenomic analysis.
Results
The growth of L. monocytogenes strains F2365, F2365_PrfA*, and F2365_LLS and L. innocua, L. ivanovii, and L. valentina strains was investigated at 4 °C at an inoculum level of 200 CFU/mL in UHT milk for 43 days and raw milk for 35 days. Additionally, the pH of the milk in the inoculated tubes was measured throughout the storage period.
Growth of non-pathogenic Listeria spp., L. ivanovii, and epidemic L. monocytogenes overexpressing virulence determinants in UHT milk at refrigerated temperature
All L. monocytogenes isolates, L. innocua, and L. ivanovii grew in UHT milk at 4 °C after a storage period of 1-2 days and reached a maximal population density between 7.50 and 8.00 Log_10_ CFU/mL after 28 days (Fig. 1A). In contrast, L. valentina did not grow under the tested conditions and showed a progressive decline in CFU/mL after day 7 (Fig. 1A). When L. monocytogenes isolates, L. ivanovii, and L. innocua were individually inoculated into UHT milk, statistically significant differences in lag phase (λ), maximal growth rate (µ_max_), and maximal population density were observed among the strains during the 43 days of the study (P < 0.05; Table 1). L. innocua exhibited a significantly shorter λ in UHT milk compared to both L. monocytogenes F2365 and F2365_PrfA* (P < 0.05) (Fig. 1A, Table 1). Additionally, the F2365_PrfA*, L. innocua and L. ivanovii showed significantly lower μ_max_ than F2365_LLS and L. monocytogenes F2365 (P < 0.05) (Table 1). L. innocua reached the highest maximal population density in UHT milk at 4 °C, which was comparable among all strains, except for L. monocytogenes F2365_PrfA*, which reached a significantly lower maximal population density (Fig. 1A, Table 1).Fig. 1. Growth dynamics of Listeria species and mesophilic aerobic bacteria in milk at 4 °C.Growth of Listeria spp. was monitored in UHT milk (A) and raw milk (B), and mesophilic aerobic bacteria (MAB) was assessed in raw milk (C) at 4 °C. Listeria spp. counts were enumerated at 0, 0.5, 1, 2, 3, 5, 7, 10, 14, 17, 21, 25, 29, 35, and 43 days in UHT milk. Listeria spp., and MAB counts were enumerated at 0, 3, 5, 7, 11, 14, 17, 21, 28, and 35 days in raw milk. The error bars showing standard deviations were performed based on four biological replicates for each strain and type of milk.Table 1. Estimated kinetic parameters of Listeria spp. grown in milk at 4 °C obtained from the modified Gompertz modelKinetic parameterMilk typeListeria spp. strainLm F2365Lm F2365_PrfALm* F2365_LLS*L. innocuaL. valentina**L. ivanovii*λ (days)UHT milk2.29 ± 0.62a2.42 ± 0.33a1.42 ± 0.18 ab0.00 ± 0.00b- -1.47 ± 0.60 abraw milk12.21 ± 1.45ab14.17 ± 1.13a12.25 ± 0.76 ab----11.17 ± 0.12 bµ_max_ [Log_10_(CFU/mL)/day]UHT milk0.35 ± 0.03 a0.28 ± 0.01b0.41 ± 0.02a0.27 ± 0.04b--0.26 ± 0.04 braw milk0.30 ± 0.03 a0.21 ± 0.03b0.30 ± 0.03a- -- -0.15 ± 0.04bLog_10_Nf_max_ [Log_10_(CFU/mL)]UHT milk7.86 ± 0.04ab7.50 ± 0.11 a7.81 ± 0.09ab8.66 ± 0.04b- -8.00 ± 0.22 abraw milk4.44 ± 0.50ab3.79 ± 0.38ab4.81 ± 0.43a----3.32 ± 0.69bRMSEUHT milk0.2590.2630.3150.275-0.263raw milk0.1570.1840.135--0.198R^2^adj.UHT milk0.9850.9820.9790.981-0.983raw milk0.9800.9530.989--0.860Data represents estimated parameter ± standard deviation. Values in the same row followed by the same lowercase letter are not significantly different (P > 0.05). Parameters are estimated from the growth curves and each point of the curves was based on four replicate samples in UHT milk and raw milk.
At the beginning of the experiment, UHT milk had a typical pH of 6.82, and no significant changes in pH were observed throughout the monitored period (Table S1). No correlation was found between bacterial growth (Log_10_CFU/mL) and pH values for any of the strains tested (Table S1).
Altogether, these results indicate that the L. innocua, L. valentina and L. ivanovii isolates tested are non-reliable surrogates for monitoring Listeria monocytogenes growth in UHT milk stored at 4 °C. Furthermore, the reduced growth rate of the PrfA* mutant shows a fitness cost compared to the parental strain in UHT milk at 4 °C (Fig. 1A, Table 1).
Growth of non-pathogenic Listeria spp., L. ivanovii, and epidemic L. monocytogenes overexpressing virulence determinants in raw milk at refrigerated temperature
All L. monocytogenes isolates and L. ivanovii grew in raw milk at 4 °C after a storage period ranging from 11 to 14 days and reached a maximal population density between 3.32 and 4.81 Log₁₀ CFU/mL after 28 days (Fig. 1B, Table 1). L. innocua did not grow, but its population levels remained stable throughout the 35-day study (Fig. 1B). In contrast, L. valentina failed to grow and showed a progressive decline in CFU/mL counts after day 14 (Fig. 1B). When L. monocytogenes isolates and L. ivanovii were individually inoculated into raw milk, statistically significant differences in lag phase (λ), maximal growth rate (µ_max_), and maximal population density were observed among the strains during the 35 days of the study (P < 0.05) (Table 1). Among all strains tested L. monocytogenes F2365_PrfA* displayed the longest adaptation period, with a significantly longer λ than L. ivanovii, which adapted most rapidly to raw milk at 4 °C (P < 0.05) (Fig. 1B, Table 1). Both L. monocytogenes F2365_PrfA* and L. ivanovii showed impaired growth rates in raw milk, with significantly lower μ_max_ values compared to L. monocytogenes F2365 and F2365_LLS strains (P < 0.05) (Table 1). Regarding L. monocytogenes and L. ivanovii isolates, L. monocytogenes F2365_LLS and L. ivanovii reached the maximal and minimal population density, respectively (P < 0.05) (Table 1 and Fig. 1B). Although not statistically significant, L. monocytogenes F2365_PrfA* showed a lower maximal population density than L. monocytogenes F2365.
At the beginning of the experiment, raw milk had a typical pH of 6.86, and no significant changes in pH were observed throughout the monitored period (Table S2). No correlation was found between bacterial growth (Log_10_CFU/mL) and pH values for any of the strains tested (Table S2).
Altogether, these results indicate that L. ivanovii, L. innocua, and L. valentina are not suitable surrogates for the F2365 L. monocytogenes epidemic strain in monitoring growth in raw milk at refrigeration temperatures. PrfA* overexpression resulted in a significant fitness cost; in contrast, LLS constitutive expression did not significantly affect growth parameters compared to the wild type strain during competition with the native microbiota of raw milk (Fig. 1B, Table 1).
Growth of mesophilic aerobic bacteria in raw milk at refrigerated temperature
The initial mesophilic aerobic bacteria (MAB) population density on raw milk samples at the beginning of the study were 3.97 Log_10_CFU/mL (Fig. 1C, Table S3). MAB increased between 9.12 and 9.65 Log_10_CFU/mL after 35 days of storage at 4 °C in all inoculated samples (Fig. 1C, Table S3). No statistically significant differences were detected for λ, μ_max_, and the maximal final concentration of MAB in raw milk contaminated with the different Listeria spp. isolates tested (P > 0.05) (Table S3).
Altogether, these results showed that the pathogenic properties of the Listeria spp. inoculated do not influence the kinetic parameters of MAB populations of raw milk.
Effect of contamination of non-pathogenic Listeria spp., L. ivanovii, and epidemic L. monocytogenes overexpressing virulence determinants on the bacterial microbiome of raw milk at 4 °C
The Richness of microbial communities on raw milk samples inoculated with L. monocytogenes F2365 was significantly reduced at day 28 when compared to day 14 (P ≤ 0.001) (Fig. 2). The Richness of microbial communities on raw milk samples inoculated with the rest of Listeria spp. was not significantly reduced at day 28 when compared to day 14 (P > 0.05) (Fig. 2).Fig. 2. Alpha diversity of raw milk samples.Boxplots showing the differences in alpha diversity measured as Richness, meaning the number of observed ASVs (amplicon sequence variants), and Evenness, regarding the abundance of these ASVs, according to L. monocytogenes F2365 wild type (WT), its constitutively active PrfA mutant (PrfA*), and the listeriolysin S mutant (LLS), together with the non-pathogenic species L. innocua, L. valentina, and L. ivanovii. Differences between groups were calculated by ANOVA based on estimated marginal means. *P < 0.05; **P > 0.01; ***P < 0.001.
Independently of the Listeria spp. inoculated into the raw milk, Evenness at day 28 was significantly higher than at day 14 (P ≤ 0.001) (Fig. 2). Although raw milk samples inoculated with L. ivanovii showed significantly higher Evenness than those inoculated with L. monocytogenes F2365 WT and F2365_LLS at day 14 (P ≤ 0.001), those differences were not observed at day 28 (P > 0.05) (Fig. 2).
The PCoA based on Weighted UniFrac analysis revealed that the majority of the differences in microbial composition were attributed to time-related alterations in the native microbiota, as indicated by the separation along PC1, which accounted for 98.92% of the variance (P ≤ 0.05) (Fig. 3). The PCoA findings indicated that samples collected on day 14 demonstrated higher similarity with limited variation. In contrast, samples from day 28 displayed more significant variability (Fig. 3). Comparisons within day 14 revealed differences between raw milk samples inoculated with L. monocytogenes F2365 and L. innocua, L. monocytogenes F2365 and L. ivanovii, F2365_LLS and L. ivanovii, and F2365_PrfA* and L. ivanovii (P ≤ 0.05) (Fig. 3). At day 28, significant differences were also detected between raw milk samples inoculated with L. monocytogenes F2365 and L. innocua (P ≤ 0.05) (Fig. 3). Altogether, these findings indicate that temporal progression was the dominant factor shaping the microbial communities in raw milk.Fig. 3. Beta diversity of raw milk samples.Principal coordinates analysis (PCoA) based on Weighted UniFrac distances shows the structure of raw milk bacterial communities inoculated with Listeria monocytogenes F2365 wild type (WT), its constitutively active PrfA mutant (PrfA*), and the listeriolysin S mutant (LLS), together with the non-pathogenic species L. innocua, L. valentina, and L. ivanovii. Bacterial communities are denoted with distinct symbols for day 14 (triangles), and for day 28 (circles). Each point corresponds to one of the five biological replicates for each raw milk sample.
On both days 14 and 28 of the study, the microbial communities in raw milk were primarily composed of the Proteobacteria phylum (>97% in all samples at day 14), regardless of the specific Listeria species present (Fig. 4). On day 14, the second most prevalent phylum was Bacteroidota (Fig. 4). Regardless of the inoculated Listeria species, Firmicutes exhibited a significant increase in relative abundance by day 28, becoming the second most prevalent phylum in the microbial community (Fig. 4).Fig. 4. Phyla of raw milk samples.Bubble plots showing the composition of the raw milk microbiota population inoculated with L. monocytogenes F2365 wild type (WT), its constitutively active PrfA mutant (PrfA*), and the listeriolysin S mutant (LLS), together with the non-pathogenic species L. innocua, L. valentina, and L. ivanovii. Phyla with a relative abundance of less than 1% have been grouped as “Other”.
On days 14 and 28 of the study, the microbial communities in raw milk were primarily dominated by the Pseudomonas genus (more than 90% relative abundance in all groups on day 14, regardless of the inoculated Listeria spp. (Fig. 5)). On day 14, genera belonging to f_Oxalobacteraceae were the second most prevalent (Fig. 5). In contrast, on day 28, Carnobacterium increased its relative abundance and became the second most prevalent genus, irrespective of the Listeria spp. present in the raw milk (Fig. 5). Moreover, there were significant increases in the relative abundances of Hafnia-Obesumbacterium and Serratia in samples inoculated with all the Listeria strains used in this study (Fig. 5). A significant increase in the relative abundance of Providencia was also observed at day 28 in samples inoculated with L. monocytogenes, L. monocytogenes_LLS, L. monocytogenes_PrfA*, and L. innocua (Fig. 5).Fig. 5. Genus of raw milk samples.Bubble plots showing the composition of the raw milk microbiota population inoculated with L. monocytogenes F2365 wild type (WT), its constitutively active PrfA mutant (PrfA*), and the listeriolysin S mutant (LLS), together with the non-pathogenic species L. innocua, L. valentina, and L. ivanovii. Genera with a relative abundance of less than 1% have been grouped as “Other”.
Overall, these data indicate that the overexpression of virulence traits of the contaminating L. monocytogenes F2365 or the specific characteristics of the non-pathogenic Listeria species did not influence the dynamics of microbial communities in raw milk.
Discussion
Comprehensive approaches must be implemented when modeling and predicting the growth of L. monocytogenes in dairy products made of milk under refrigeration conditions. To our knowledge, apart from the present study, only two studies have previously evaluated the growth potential of the low number of Listeria spp. cells frequently counted in raw milk^24,25^. In the present study, lower CFU counts were observed for Listeria spp. in raw milk compared to UHT milk (Fig. 1A, B), showing that native microbiota can limit the growth of Listeria spp. This inhibitory effect aligns with previous studies of our group where raw milk also restricted L. monocytogenes growth under cold storage^25^. Notably, this growth limitation contrasts with the homogenous growth of MAB CFU across the study (Fig. 1C).
In food safety studies, L. innocua is commonly used as a non-pathogenic surrogate for L. monocytogenes^26–32^. However, our findings indicate that L. innocua growth in raw milk microbiota at refrigeration temperatures is restricted. This result suggests that L. innocua may not always be a reliable model for L. monocytogenes survival or growth, particularly in complex microbial environments such as refrigerated dairy products made of raw milk. Previous studies have shown that certain L. monocytogenes strains are more resistant to cold stress than L. innocua^33,34^, and variations in L. innocua’s thermal resistance^26^. further complicate its use as a universal indicator. Our findings indicate that L. innocua or L. ivanovii would not be adequate surrogates in milk at refrigeration temperatures. Finally, L. valentina is not a reliable surrogate for L. monocytogenes in UHT or raw milk at refrigeration temperatures, as it did not grow under these conditions. The present results highlight the need for a more precise selection of surrogate organisms tailored to specific food matrices and environmental conditions, ensuring an accurate risk assessment. Moreover, different surrogates may be required depending on the L. monocytogenes lineage to be evaluated, such as host-adapted lineage I (hypervirulent) or food-adapted lineage II (hypovirulent)^9,20^.
Most dairy-related outbreaks have been linked to products made of unpasteurized milk^13^. Research across various food categories revealed a link between hypervirulent L. monocytogenes clones (CC1, CC4, and CC6) and dairy products produced from raw milk^10^. However, no studies evaluated the growth behavior of outbreak-related L. monocytogenes and its isogenic derivatives overexpressing key virulence factors in UHT and raw milk. The most important virulence factors of L. monocytogenes strains are encoded in the inlA-inlB loci and the pathogenicity islands LIPI-1, LIPI-3, and LIPI-4^8^. LIPI-1 and the inlA-inlB locus are regulated by the transcriptional activator PrfA and form part of the core genome, whilst LIPI-3 is part of the accessory genome^4^. Genetic analysis of 6,633 food and clinical isolates of L. monocytogenes, in conjunction with epidemiological and clinical data, identified full-length InlA and LIPI-3 as being strongly associated with infectious potential^9^. Moreover, listeriosis clinical cases have been associated with lineage I strains^9^. The role of virulence determinants in food contamination has been somewhat overlooked, as most studies primarily focus on their role in microbial pathogenesis. Consequently, whether PrfA-controlled genes and LIPI-3 affect the performance of hypervirulent lineage I L. monocytogenes in UHT and raw milk remained experimentally unconfirmed. Here, we analysed the fitness consequences of overexpressing virulence traits (LIPI-3 and the PrfA regulon) using a CC1 lineage I L. monocytogenes strain related to a cheese outbreak^11^. Using LIPI-3 and PrfA-controlled genes to be constitutively activated, we demonstrate that LIPI-3 overexpression, compared to PrfA overexpression, enhances the listerial growth rate in UHT and raw milk at refrigeration temperature. This fitness advantage conferred by LIPI-3 could impose a beneficial cost outside the host and provide new insights into understanding why LIPI-3 is frequently found in CC1, CC4, and CC6 isolates, which are also overrepresented in dairy products^10^. On the contrary, PrfA overexpression imposed a significant cost to L. monocytogenes in UHT and raw milk, which is in agreement with Vasanthakrishnan et al. (2015), who previously demonstrated experimentally that PrfA overexpression lowers L. monocytogenes fitness in BHI medium and soil^35^. Similar to our results related to PrfA constitutive activation, a study on Salmonella associated with the cost of virulence factors in in vitro culture showed that expression of the type III secretion system (TTSS)-1 was related to growth retardation^36^. Contamination with non-pathogenic Listeria spp., L. ivanovii, or virulence-overexpressing L. monocytogenes did not alter the raw milk microbiome, maintaining a stable community throughout the study, which was dominated by psychrotrophic genera such as Pseudomonas and Carnobacterium. Indeed, Pseudomonas has been consistently reported as a dominant taxon in refrigerated milk^37^ and identified as a member of the core raw milk microbiota in a recent large-scale survey^38^. The occurrence of Carnobacterium in raw milk across different breeds, together with functional traits such as bacteriocin production, has been previously reported^39,40^. Therefore, the observed stability of the raw milk microbiota suggests that these psychrotrophic genera play a central role in shaping the community dynamics of this type of food matrix.
In conclusion, our study shows that at 4 °C: (i) the PrfA* mutant exhibits a fitness cost in milk, reflected in its reduced growth rate compared to the epidemic L. monocytogenes F2365 wild type strain; (ii) constitutive expression of LLS does not significantly impact growth parameters compared to the epidemic L. monocytogenes F2365 wild type strain; (iii) L. ivanovii, L. innocua, and L. valentina are not suitable surrogates for the epidemic L. monocytogenes F2365 strain in UHT and raw milk; and (iv) neither the overexpression of virulence traits in epidemic L. monocytogenes F2365 strain nor the characteristics of non-pathogenic Listeria species altered the dynamics of microbial communities in raw milk.
Our results regarding the use of Listeria spp. as index or indicator organisms will help food-processing facilities identify conditions that increase the risk of L. monocytogenes presence, growth, and/or contamination. This knowledge will help to reduce the number of human listeriosis cases caused by food contamination with L. monocytogenes.
Methods
Bacterial strains and inoculum preparation
Six Listeria spp. isolates were used in this study, comprising both pathogenic (L. monocytogenes (n = 3) and L. ivanovii) and non-pathogenic strains (L. innocua and L. valentina). Detailed strain information is provided in Table S4. The L. monocytogenes F2365 wild type (WT) strain was selected for its epidemiological significance, as it was responsible for the 1985 listeriosis outbreak linked to Mexican-style cheese, which resulted in 142 human cases and 48 deaths^11^. To achieve constitutive activation of PrfA, we used the L. monocytogenes F2365_PrfA* strain, which carries a Gly145Ser substitution in this transcriptional activator, leading to the continuous expression of LIPI-1 virulence factors^14,41^. Additionally, to assess the role of LLS in growth within UHT and raw milk, we employed L. monocytogenes F2365_LLS (previously denominated pHELP:llsA, a synthetic strain designed to constitutively express LLS by placing the LIPI-3 genes under the control of the strong constitutive pHELP promoter^16^.
All isolates were stored at −80 °C in glycerol stocks. Before use, strains were streaked onto Brain Heart Infusion (BHI) agar and incubated at 37 °C for 48 h. A single colony was aseptically transferred to 2 mL of BHI broth and grown at 37 °C for 16 h with shaking at 250 rpm. Cultures were then resuspended in phosphate-buffered saline (PBS) to achieve a final inoculum of approximately 2.3 Log₁₀ CFU/mL ( ~ 200 CFU/mL).
Milk sample collection and microbiological assessment
To assess Listeria spp. growth in UHT milk, four 1 L containers of the same commercial brand were purchased locally. Raw bovine milk was collected under refrigeration 24 h post-milking from the Milk Analysis Laboratory (Polytechnic University of Valencia) and processed immediately upon arrival. Listeria spp. detection and MAB counts were performed as previously described^25,42^ following the ISO 11290-1^43^, and ISO 11290-2^44^ for Listeria spp., and according to ISO 4833-2^45^ for quantification of MAB. The initial MAB count at 4 °C was 9,400 CFU/mL (3.97 Log₁₀CFU/mL).
Assessment and modelling of Listeria growth in milk matrices
Bacterial growth was monitored at 4 °C for up to 43 days in UHT milk and 35 days in raw milk. Listeria spp. was enumerated on BHI agar (for UHT milk) or ALOA agar (for raw milk), while MAB were quantified on Plate Count Agar (PCA). Samples with ≤100 CFU/mL were directly plated; otherwise, serial dilutions in PBS were performed. Plates were incubated at 37 °C for 24 h (Listeria spp.) and 30 °C for 72 h (MAB). Each experiment included four biological replicates per isolate and matrix, with three technical replicates per time point. Log₁₀CFU/mL values and pH (measured with a sensION™+ pH meter and XS SEMI-MICRO electrode) were recorded at each sampling day.
The growth of Listeria spp. over time was modeled using the modified Gompertz equation^46^, a widely used primary model that describes bacterial population dynamics as a function of time. As an empirical sigmoidal function, it enables reliable estimation of kinetic parameters such as maximum growth rate (μ_max_), lag time (λ), and asymptotic population size (C). Although the Baranyi model^47^ offers a mechanistic approach better suited to fluctuating environmental conditions, the Gompertz equation has been widely validated for constant environments^48,49^ and frequently applied to model L. monocytogenes growth in refrigerated dairy products^50–52^. Given that the present experiments were conducted under static conditions, the Gompertz model was considered the most appropriate choice.
The equation used was:
\documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${\mathrm{Log}}_{10}{\rm{N}}({\rm{t}})={\mathrm{Log}}_{10}{\mathrm{N}}_{0}+\begin{array}{c}{\mathrm{C}}* \end{array}\exp \left(-\exp \left(\left(\frac{\begin{array}{c}2.718* \end{array}{{\rm{\mu }}}_{\max }}{\mathrm{C}}\right)* \left(\lambda -\mathrm{t}\right)+1\right)\right)$$\end{document}Where N(t) represents Listeria spp. concentration (CFU/mL) at a particular time; N0 the initial Listeria spp. concentrations (CFU/mL); C the difference between the curve asymptotes (Log_10_(CFU/mL)), corresponding to the difference between the maximum and the minimum Log_10_ CFU/mL reached; μ_max_ the maximal growth rate (Log_10_(CFU/mL)/day); λ the lag phase (days); and t the particular time (days).
Initial estimates for N₀ of each biological replicate were derived by plotting experimental Log_10_ CFU/mL data against time. Kinetic parameters were fitted using Statgraphics Centurion XVII software. The goodness of fit was evaluated by calculating the Root Mean Square Error (RMSE), which reflects the deviation of model predictions from observed values, and the adjusted coefficient of determination (R²adj), which indicates the proportion of variance explained by the model. The goodness-of-fit indicators obtained in our study demonstrated that the model reliably described the data: all the RMSE values were <0.259 and <0.157 for UHT and raw milk, respectively, and the adjusted R²_adj_ values > 0.981 and >0.860 for UHT and raw milk, respectively, confirming its accuracy in capturing the growth kinetics of Listeria spp. in UHT and raw milk (Table 1).
DNA extraction, library preparation, and 16S rRNA sequencing of raw milk samples
Metagenomic analysis was conducted on raw milk samples collected at days 14 and 28, corresponding to the exponential and stationary phases of Listeria spp., to assess microbiota dynamics. DNA was extracted using the DNeasy PowerLyzer PowerSoil Kit (Qiagen) using Tissue lyser II (Qiagen) at 30 Hz, 10 min, 4 °C. Amplification targeted the V3–V4 region of the 16S rRNA gene using specific primers. PCR products were purified with AMPure XP beads and dual-indexed using Nextera XT v2 adapters in a second PCR. Libraries were normalized (SequalPrep kit (ThermoFisher Scientific)), quantified by qPCR, pooled, and sequenced on an Illumina MiSeq platform (2 × 300 bp, v3 chemistry, 10 pM loading, 15% PhiX spike-in). To ensure data integrity, negative controls were analyzed. Amplicon data were processed in QIIME2 (v2019.4) using DADA2 for quality filtering, denoising, and amplicon sequence variant calling (ASV)^53,54^. Reads were truncated at Q20 (299 nt forward, 243 nt reverse); average sample size was 83,230 reads with 8,428 ASVs detected. ASVs were aligned using the qiime alignment mafft method^55^. The alignment was used to create a tree and to calculate phylogenetic relations between ASVs using qiime2 phylogeny fasttree method^56^. Diversity metrics (richness, Pielou’s evenness) and Weighted UniFrac distances were calculated after rarefaction to 49,400 reads. The smallest sample size was chosen for subsampling^57^. Taxonomy was assigned using a Bayesian Classifier trained with Silva database (i.e., 99% ASVs database) using the qiime feature-classifier classify-sklearn method^58^. Phylotypes were filtered to discard contaminant Eukariota DNA-derived amplicons using Blast against the mentioned database with a 90% identity cutoff. Mock community and negative controls were processed the same way as samples.
Statistical analysis
Statistically significant differences in Listeria spp. and MAB growth, and taxon abundance were assessed using ANOVA or Kruskal–Wallis tests. Alpha diversity was analyzed using generalized mixed models: negative binomial for Richness and beta regression for Evenness. Beta diversity (Weighted UniFrac analysis) was compared using PERMANOVA, ANOSIM, and PERMDISP. Taxon differential abundance was evaluated with Kruskal–Wallis’s test. For all statistical analyses, the significance level was set at 0.05. Figures were generated with GraphPad Prism (Version 8.0) and Adobe Illustrator (Version 27.0).
Supplementary information
Supplementary information
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brown, P. et al. Listeria tempestatis sp. nov. and Listeria rocourtiae subsp. hofi subsp. nov. Int. J. Syst. Evol. Microbiol.75, 006774 (2025).10.1099/ijsem.0.006774 PMC 1237842940358009 · doi ↗ · pubmed ↗
- 2Vázquez-Boland, J. A. et al. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev.14, 584–640 (2001).10.1128/CMR.14.3.584-640.2001 PMC 8899111432815 · doi ↗ · pubmed ↗
- 3Guillet, C. et al. Human listeriosis caused by Listeria ivanovii. Emerg. Infect. Dis.16, 136–138 (2010).10.3201/eid 1601.091155 PMC 287437820031061 · doi ↗ · pubmed ↗
- 4Quereda, J. J. et al. Pathogenicity and virulence of Listeria monocytogenes: A trip from environmental to medical microbiology. Virulence 12, 2509–2545 (2021).10.1080/21505594.2021.1975526 PMC 849654334612177 · doi ↗ · pubmed ↗
- 5European Food Safety Authority (EFSA) & European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2023 Zoonoses report. EFSA J 22, e 9106 (2024).10.2903/j.efsa.2024.9106 PMC 1162902839659847 · doi ↗ · pubmed ↗
- 6Disson, O., Moura, A. & Lecuit, M. Making sense of the biodiversity and virulence of Listeria monocytogenes. Trends Microbiol.29, 811–822 (2021).10.1016/j.tim.2021.01.00833583696 · doi ↗ · pubmed ↗
- 7Maury, M. M. et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet.48, 308–313 (2016).10.1038/ng.3501 PMC 476834826829754 · doi ↗ · pubmed ↗
- 8Maury, M. M. et al. Hypervirulent Listeria monocytogenes clones’ adaption to mammalian gut accounts for their association with dairy products. Nat. Commun.10, 2488 (2019).10.1038/s 41467-019-10380-0PMC 655440031171794 · doi ↗ · pubmed ↗