Cloning and functional analysis of the lobed-leaf gene BjA10.LL in Brassica juncea L
Jiajia Liu, Yunyun Ma, Yuxuan He, Xiaohui Cui, Shaojie Ma, Zijin Liu, Saiqi Yang, Yuan Guo, Mingxun Chen

TL;DR
This study identifies a gene responsible for leaf margin lobation in Brassica juncea and shows how regulatory variations affect leaf shape, aiding in breeding programs.
Contribution
The study identifies BjA10.LL as a gene controlling leaf lobation and demonstrates the role of cis-regulatory variations in this trait.
Findings
BjA10.LL, an HD-ZIP I gene, is responsible for leaf margin lobation in Brassica juncea.
Cis-regulatory variations in the promoter region of BjA10.LL influence its expression and leaf morphology.
A co-dominant molecular marker for BjA10.LL indels co-segregates with the lobed-leaf phenotype in an F₂ population.
Abstract
Leaf shape plays a crucial role in plant growth and development. Among various leaf traits, marginal lobation serves as an ideal morphological marker for breeding programs. However, the genetic mechanism underlying leaf margin lobation in Brassica juncea L. remains unclear. Through RNA sequencing and map-based cloning, we identified an incompletely dominant gene, BjA10.LL, which encodes an HD-ZIP I protein and is responsible for the formation of leaf margin lobation in B. juncea. Sequence analysis of parental alleles revealed no critical variations in the coding region but identified substantial variations in regulatory regions. Heterologous expression of BjA10.LL in Arabidopsis thaliana confirmed its sufficiency to induce lobed leaves. To functionally link the regulatory variations to the phenotype, we analyzed promoter activity and developed a co-dominant molecular marker targeting…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
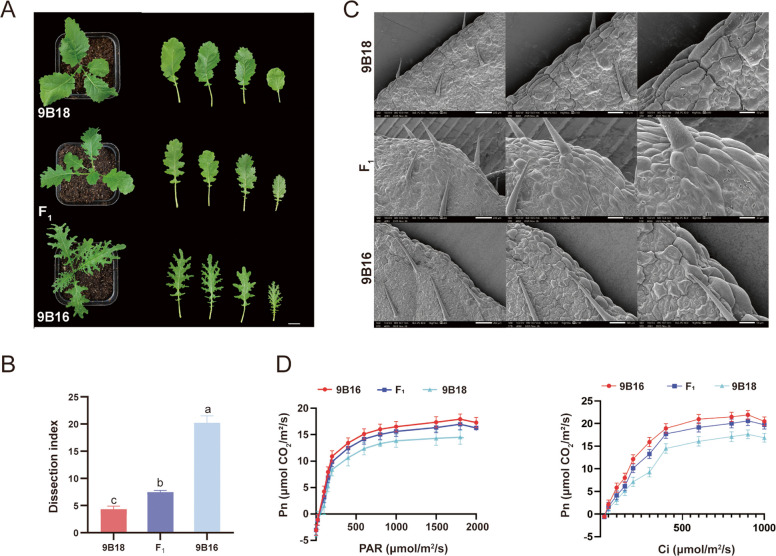 Figure 1
Figure 1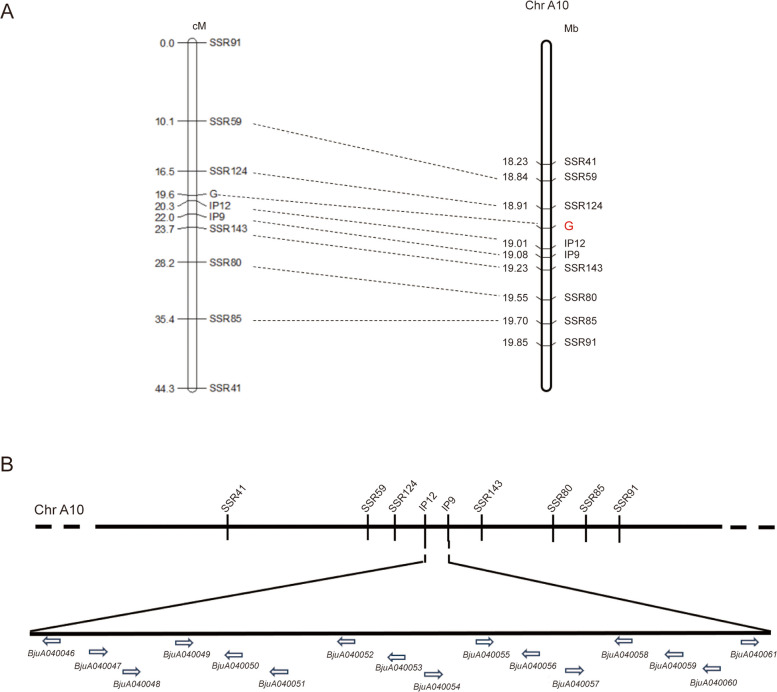 Figure 2
Figure 2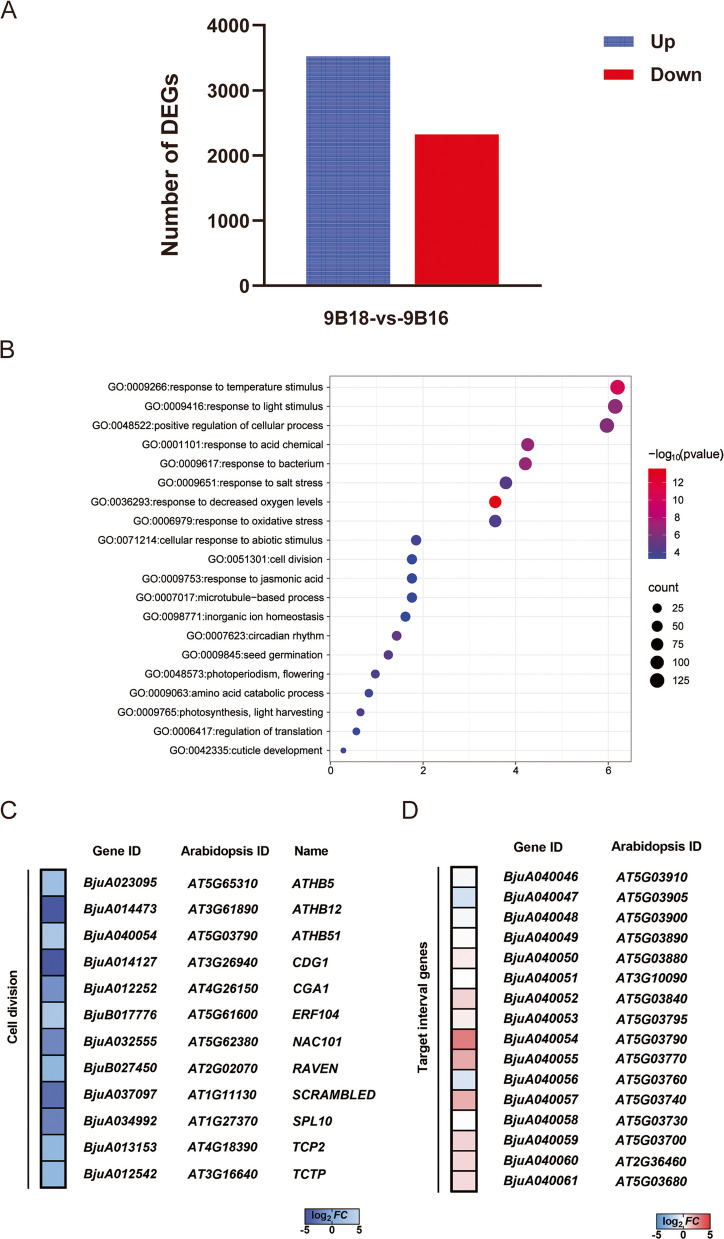 Figure 3
Figure 3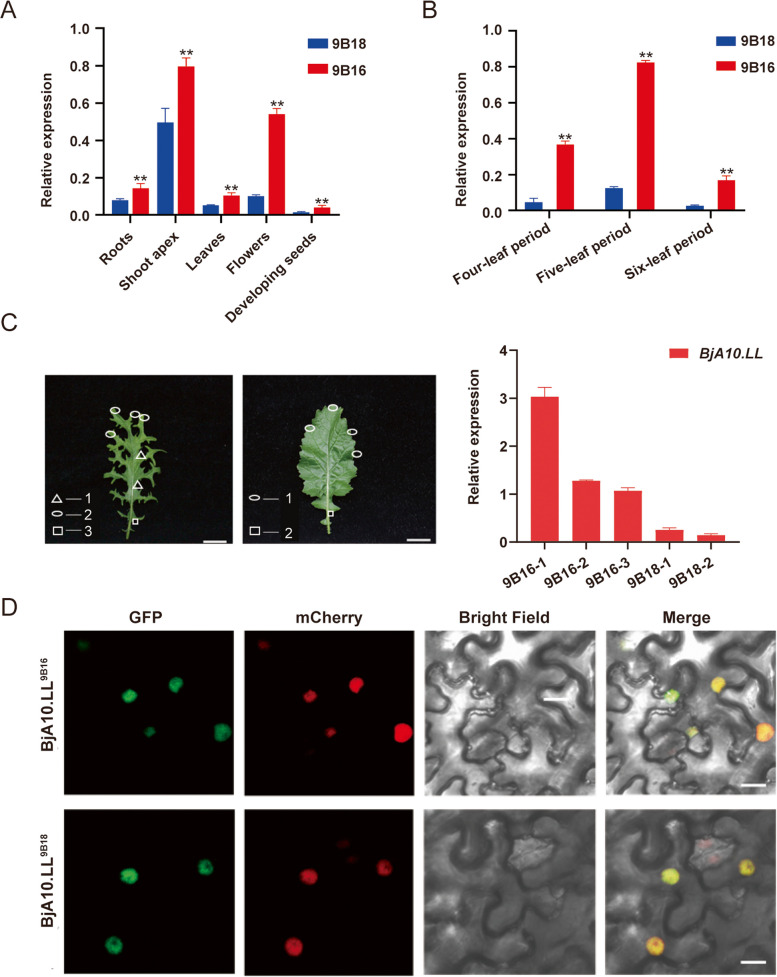 Figure 4
Figure 4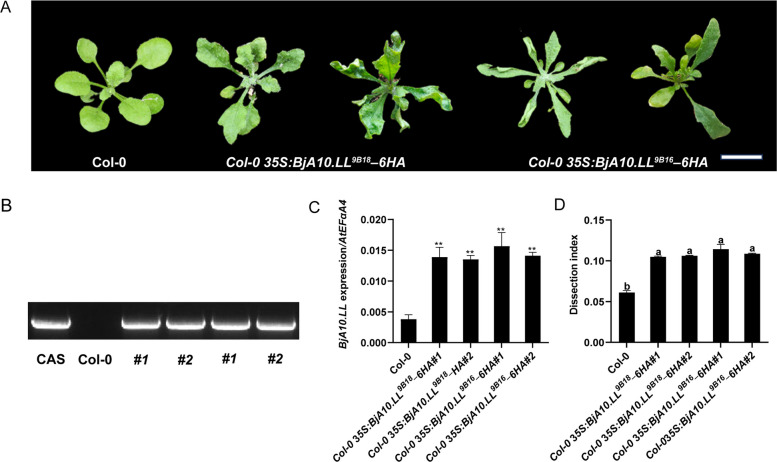 Figure 5
Figure 5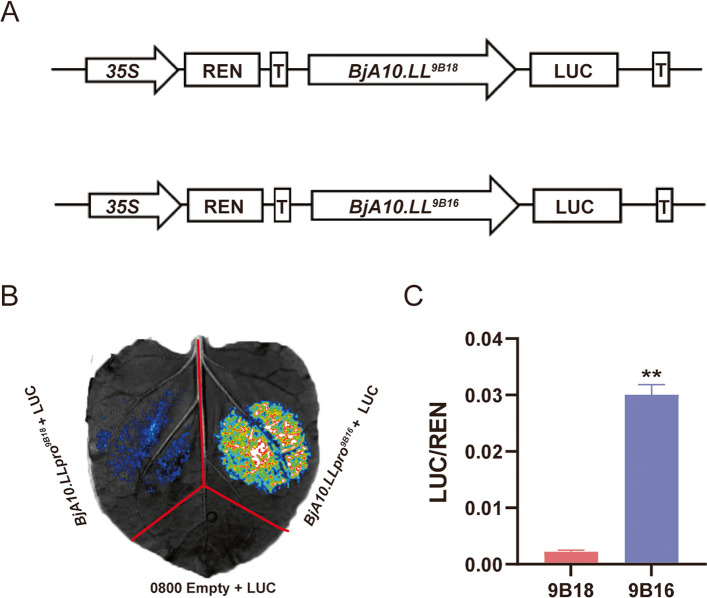 Figure 6
Figure 6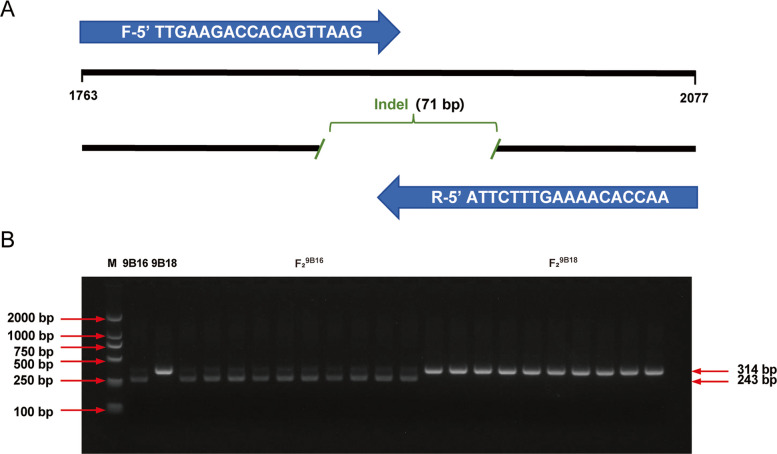 Figure 7
Figure 7- —National Key Research and Development Program
- —http://dx.doi.org/10.13039/501100015401Key Research and Development Projects of Shaanxi Province
- —Scientific and Technological Innovation Team of Shaanxi Province
- —the Science and Technology Program Project of Yangling Demonstration Zone
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant nutrient uptake and metabolism · Plant Reproductive Biology
Introduction
Rapeseed (Brassica juncea L., AABB, 2n = 36) exhibits distinct morphological characteristics and extensive genetic diversity. It is mainly cultivated in India, Pakistan, Africa, central Russia, Canada, Eastern Europe and the southwest and northwest regions of China. This species possesses multiple valuable traits, including drought resistance, poor soil tolerance, diseases and pests resistance, cold resistance, lodging resistance and pod shattering resistance (Kang et al. 2021).
Leaves are the primary organs of B. juncea, essential for photosynthesis, gas exchange, and water transport (Li et al. 2007). One of the main drivers of leaf morphological diversification is leaf margin variation (Tsukaya 2006). Leaf margin lobation is a leaf shape variation formed by plants to adapt to their living environments, which has important biological significance and is a key focus in the study of leaf morphology in Brassicaceae crops. Based on the depth of leaf margin lobation, leaves can be classified into several types, such as entire leaves, serrated leaves, and lobed leaves. Lobed leaves exhibit greater spatial extensibility, can regulate leaf surface temperature, and reduce water loss, thereby accumulating more photosynthetic products to adapt to harsh environments such as high temperature, cold, and drought (Tsukaya 2006; Vogel 2009). Therefore, identifying the genes controlling leaf margin lobation and elucidating their regulatory mechanisms will lay a solid foundation for the utilization of leaf shape traits in the genetic improvement of B. juncea.
In Arabidopsis thaliana, it was first demonstrated that the Homeobox-Leucine Zipper (HD-ZIP) transcription factor encoded by the LATE MERISTEM IDENTITY 1 (LMI1) gene plays an important role in the formation of serrated leaves (Saddic et al. 2006). Through forward genetic screening of leaf shape in Cardamine hirsuta, the REDUCED COMPLEXITY (RCO) gene was successfully identified (Vlad et al. 2014). This gene is thought to have evolved via tandem duplication of the LATE MERISTEM IDENTITY 1 (LMI1) gene and has no homolog in A. thaliana. The increased expression of RCO in A. thaliana increases leaf complexity, indicating that the loss of RCO during the process of speciation is the key to the simple leaf phenotype of this species (Vlad et al. 2014). However, unlike LMI1, RCO is specifically expressed at the base of leaflets. Through time-lapse imaging and growth analysis at the cellular level, it was found that RCO inhibits local growth at the base of leaflets (Vlad et al. 2014; Vuolo et al. 2016; Nikolov et al. 2019). There are three homologous copies of LMI1-like genes in C. hirsuta, namely LMI1-like1, LMI1-like2 (RCO), and LMI1-like3. RCO promotes the formation of leaflets by inhibiting the growth of the leaf margin. In addition, the LMI1-like transcription factor plays an important role in the regulation of leaf shape in cotton. Silencing of GhOKRA results in wider cotton leaves, while its overexpression in A. thaliana enhances leaf lobation depth (Chang et al. 2016; Andres et al. 2017). RCO directly regulates genes involved in cytokinin (CK) homeostasis, and enhancing CK signaling within the RCO expression domain in A. thaliana increases leaf complexity (Hajheidari et al. 2019). In Brassicaceae, RCO does not affect auxin transport but cooperates with the CUC-auxin regulatory module to inhibit cell growth on both sides of the lobation, thereby modulating lobation depth (Vlad et al. 2014). More recently, BnA10.RCO was shown to positively regulates the formation of leaf margin lobation and overall leaf complexity in Brassica napus (Hu et al. 2020). However, the regulatory mechanism of the RCO gene underlying leaf shape formation in B. juncea remains unclear. Here, we employed two phenotypically distinct advanced inbred lines of B. juncea (the entire-leaf line 9B18 and the deeply lobed-leaf line 9B16), which exhibit stable and significant leaf morphological differences after multiple generations of selfing, making them ideal genetic resources for investigating leaf morphogenesis. We demonstrate that the BjA10.LL gene positively regulates leaf margin lobation in B. juncea. This finding not only provides novel insights into the molecular regulation of leaf shape but also offers a valuable candidate gene for molecular design breeding in rapeseed.
Results
Phenotypic and physiological characteristics of entire-leaf margins and lobed-leaf margins
In this study, two advanced inbred lines, 9B18 and 9B16, which exhibit significant differences in leaf margin morphology, were used as parental materials. An F_1_ population was generated by crossing these two lines. The results showed that 9B16 displayed deeply lobed leaves, while the hybrid F_1_ progeny exhibited serrated leaves, consistent with an intermediate phenotype of incomplete dominance (Fig. S1).
Further analysis of the phenotypic and physiological characteristics of leaf margins in two advanced inbred parental lines (9B18 and 9B16) and their F_1_ hybrid revealed that the lobed-leaf margin trait could be observed from the first true leaf stage throughout the entire growth period (Fig. 1A). Significant differences in leaf anatomical indices were observed between the entire-leaf parent and the lobed-leaf parent (Fig. 1B). The formation of lobed-leaf margins is attributed to the activity of meristematic tissues in specific regions of the leaf margin, which prolongs the undifferentiated development of the leaf and delays its maturation. To elucidate the relationship between leaf margin cell characteristics and leaf shape, we conducted detailed observations of the microscopic structures of leaf margins in entire-leaf 9B18, lobed-leaf 9B16 and F_1_ using scanning electron microscopy. The results showed that, compared to the typical elongated rod-shaped cells and larger protrusions at the leaf margins of 9B18, the leaf margins of 9B16 exhibited characteristics of meristematic cells, including smaller, rounder, and densely arranged cells (Fig. 1C). These findings suggest that cell division and growth processes resembling meristematic activity ultimately lead to the formation of the lobed-leaf margin trait in 9B16.Fig. 1. Phenotypic and physiological characteristics of the leaf margins in the two advanced-generation inbred lines 9B18, 9B16 and their F_1_ hybrid. A The lobed-leaf margin phenotypes of the leaves of 9B18, 9B16 and F_1_. 9B18 represents entire-leaf margins, 9B16 represents lobed-leaf margins. Scale bar = 2 cm. B Leaf dissection index of 9B18, 9B16 and F_1_ plants. Values are means ± SD (n = 10). Bars labeled with different lowercase letters indicate significant differences (P ≤ 0.01, one-way ANOVA followed by Tukey's HSD test). C Scanning electron microscopy observation of leaf margin cells in seedling leaves of 9B18, 9B16 and F_1_. Scale bars = 200 μm, 100 μm, and 50 μm. D Photosynthetically active radiation intensity and intercellular CO_2_ concentration between the leaves of 9B18, 9B16 and F_1_. Values are means ± SD (n = 10)
Additionally, we analyzed the impact of the lobed-leaf margin trait on photosynthetic efficiency. By measuring the photosynthetically active radiation intensity (PAR) and intercellular CO₂ concentration (Ci) in both parental lines (9B18 and 9B16) and their F_1_ hybrid, we found that 9B16 exhibited significantly higher PAR intensity and Ci compared to 9B18, with F_1_ progeny showing intermediate values (Fig. 1D). These results indicate that the lobed-leaf material demonstrated greater photosynthetic utilization efficiency than the entire-leaf material in this study.
Inheritance of the lobed-leaf margin trait
To study the genetic pattern of the lobed-leaf margin trait, we constructed F_1_, F_2_ and BC_1_ segregating populations by crossing 9B18 (entire-leaf margin) and 9B16 (lobed-leaf margin). Based on two consecutive years of field observations in 2021 and 2022, the segregation of the target traits occurred in both the F_2_ and BC_1_ populations, which were manifested as entire-leaf margins and lobed-leaf margins respectively. Chi-square test showed that the segregation ratio of entire-leaf margin to lobed-leaf margin individuals in the F_2_ population fitted a 1:3 ratio, while the ratio in the BC_1_ population fitted 1:1. These results indicate that the lobed-leaf margin phenotypic trait of 9B16 is controlled by a single nuclear gene, without cytoplasmic genetic effects, and the lobed-leaf margin is an incompletely dominant trait (Table 1). Table 1. Genetic analysis of the lobed-leaf margin traitYearsPopulationPlants testedEntire-leafLobed-leafMendelian expectationsχ^2^P20219B18 (P_1_)1001000———9B16 (P_2_)1000100———F_1_1000100———F_2_181346513481:30.4060.52420229B18 (P_1_)1001000———9B16 (P_2_)1000100———F_1_1000100———BC_1_7874033841:10.4590.498F_2_4691273421:31.0810.299
Fine mapping of the lobed-leaf margins gene
To rapidly localize the target gene region, this study employed the high-density Brassica 50 K Illumina Infinium™ SNP array, combined with bulked segregant analysis (BSA) and collinearity analysis, to screen for SNP markers and preliminarily map the leaf margin lobation gene in B. juncea. SNP array analysis identified four polymorphic SNP loci between 9B18 and 9B16 from genome-wide distributed SNP markers, all located on the A10 chromosome of B. napus (Table S2). Through collinearity comparison, the 17.07 ~ 17.38 Mb region on the A10 chromosome of B. napus was mapped to the 18.16 ~ 19.32 Mb region on the A10 chromosome and the 0.51 ~ 0.53 Mb region on the B02 chromosome of B. juncea (Table S3).
Within the target interval, 179 pairs of simple sequence repeat (SSR) primers and 17 pairs of intron polymorphism (IP) primers were designed. Among these, seven SSR primers (SSR41, SSR59, SSR80, SSR85, SSR91, SSR124, and SSR143) and two IP primers (IP9 and IP12) exhibited stable polymorphism in a small population (Fig. S2). These nine stable molecular markers were used to genotype 839 individuals from the F_2_ population, and the genetic distances between the markers and the target gene were calculated based on the number of recombinant individuals detected. Ultimately, the target gene was confined to a 3.8 cM region. The closest flanking markers, IP12 and SSR124, were located at genetic distances of 19,010,522 bp and 18,911,075 bp from the target gene, respectively, with a physical distance of 995 kb between them (Fig. 2A). The localization analysis revealed that the target interval encompasses a total of 16 genes (Fig. 2B). Functional annotation of these 16 genes was performed by integrating the B. juncea and A. thaliana databases (Table S4). Among these genes, only BjuA040054 encodes an HD-ZIP I protein, which has been shown to play a significant role in the formation of leaf margin lobation (Hu et al. 2020).Fig. 2. Fine mapping of the BjA10.LL genes. A Genetic map and physical map of BjA10.LL genes. B The distribution of 16 genes in the target interval
Sequence alignment and transcriptome analysis identified BjA10.LL as a candidate gene
The amino acid sequences of the candidate gene BjuA040054 and the RCO gene in B. napus were aligned using the DNAMEAN software (Fig S3). The results showed that the protein encoded by BjuA040054 shares high sequence identity with RCO protein (BnaA10g26320D) from B. napus. This indicates that the proteins encoded by the BjuA040054 gene in B. juncea and the RCO gene in B. napus may have similar biological functions.
To further validate the candidate gene and explore the molecular mechanism of the lobed-leaf margin formation in B. juncea, we used entire-leaf plants (9B18) as the control group and lobed-leaf plants (9B16) as the treatment group, and performed transcriptome sequencing on the leaves at the five-leaf stage. For differential expression analysis, |Log_2_ (Fold Change)|≥ 2 and FDR < 0.05 were set as the screening criteria. The analysis results showed that a total of 5846 differentially expressed genes (DEGs) were identified, among which 3523 genes were up-regulated and 2323 genes were down-regulated (Fig. 3A). To explore the biological functions of the DEGs related to the formation of lobed-leaf margins in this study, we used the BLAST software for sequence alignment to obtain the homologous genes of each DEGs in A. thaliana, and then conducted GO enrichment analysis. The biological processes that were significantly and commonly enriched include cell division, photosynthesis, photoperiodism, positive regulation of cellular process, etc.(Fig. 3B). Meanwhile, based on the GO annotation information, we further analyzed the DEGs related to cell division. Among them, genes such as ATHB5, ATHB51, TCP2, and TCTP were upregulated, while genes such as CDG1, SPL12, and NAC101 were downregulated (Fig. 3C). To verify the authenticity of the RNA sequencing (RNA-seq) data, we randomly selected nine DEGs for RT-qPCR verification. The results showed that the expression trends of these genes in 9B18 and 9B16 were consistent with the results of transcriptome sequencing (Fig. S4), confirming the reliability of the transcriptome sequencing data.Fig. 3. Transcriptome analysis of BjA10.LL regulation. A Statistics of differentially expressed genes. B Differential expression genes GO enrichment. C Gene expression analysis related to cell differentiation. D Expression analysis of candidate genes within the target interval. The numerical values in the legend represent the log_2_ fold change value of the expression level of this gene in 9B16 compared to that in 9B18
By integrating transcriptome and gene mapping results, it was found that among the 16 genes in the mapped candidate interval, only BjuA040054 was differentially expressed. In conclusion, we speculate that BjuA040054 (designated as BjA10.LL) is the candidate gene controlling the lobed-leaf margin trait in B. juncea.
BjA10.LL expression pattern and subcellular localization
To study the function of BjA10.LL, we analyzed its spatiotemporal expression patterns in 9B18 and 9B16 using RT-qPCR. The results showed that BjA10.LL is widely expressed in various tissues, with particularly high expression in the shoot apex (Fig. 4A). Additionally, its expression level was highest at the five-leaf stage (Fig. 4B). Furthermore, we investigated the detailed expression patterns of BjA10.LL in different leaf parts at the five-leaf stage in 9B18 and 9B16. The results revealed that in 9B18, BjA10.LL expression was relatively low in all leaf parts, whereas in 9B16, it was expressed in nearly all leaf parts at relatively high levels (Fig. 4C).Fig. 4. Analysis of the BjA10.LL expression pattern. A RT-qPCR analysis of the expression of BjA10.LL in different tissues of B. juncea. Scale bars: 1 cm. B RT-qPCR analysis of BjA10.LL in the leaves of 9B18 and 9B16 seedlings at the four-leaf period, five-leaf period and six-leaf period. C RT-qPCR analysis of 9B18 and 9B16 in different leaf segments at the five-leaf period. D Subcellular localization of BjA10.LL^9B18^ and BjA10.LL^9B16^ proteins fused with green fluorescent protein (GFP) (35S:BjA10.LL.^XX^–GFP) in tobacco leaves. mCherry is a nuclear localized protein fused with a red fluorescent protein; merge, merge of mCherry, GFP, and bright field images. Scale bars = 50 μm. Results were normalized against the expression of BjuUBQ9 as an internal control. Values are means ± SD (n = 3). Double asterisks indicate that there are highly significant differences in the expression levels between 9B18 and 9B16 (two-tailed paired Student's t-test, P ≤ 0.01). The ‘XX’ of BjA10.LL means 9B18 and 9B16
To determine whether BjA10.LL functions as a transcription factor, we observed the GFP fluorescence signal of the fusion protein BjA10.LL^XX^–GFP expressed in tobacco leaves. The results showed that both 35S:BjA10.LL^9B18^–GFP and 35S:BjA10.LL^9B16^–GFP were specifically localized in the nucleus (Fig. 4D). These results suggest that BjA10.LL may act as a transcription factor to regulate leaf development in B. juncea.
Functional analysis of BjA10.LL in regulating the formation of leaf margin lobes in A. thaliana
To investigate the role of the BjA10.LL gene in the formation of lobed-leaf margins, we amplified the DNA fragments of BjA10.LL from the 9B18 and 9B16 lines, respectively. Comparative analysis revealed three nonsynonymous SNP differences between the two parental lines (Fig. S5A). Further amplification of the cDNA fragments from 9B18 and 9B16 showed that, at the 35th bp position of the first exon, the nucleotide sequence changed from ACG to ATG, resulting in the substitution of threonine (Thr) with methionine (Met) at the 12th amino acid position (Fig. S5B).
In order to further explore whether the changes in amino acids affect the formation of leaf margin lobation, we transformed the overexpression constructs of 35S:BjA10.LL^9B18^–6HA and 35S:BjA10.LL^9B16^–6HA into A. thaliana Col-0 plants, respectively. Through hygromycin selection, 10 independent T_1_ transgenic lines were obtained. The expression level of BjA10.LL in the representative overexpression transgenic lines of the T_3_ generation was measured by RT-qPCR. The results showed that, compared with the wild type, the expression of BjA10.LL in the transgenic lines was significantly up-regulated. All the transgenic lines produced deeply incised leaves that had never been observed in the wild type (Fig. 5A-D). These results indicate that BjA10.LL positively regulates the formation of leaf margin lobation in A. thaliana leaves.Fig. 5. Functional analysis of BjA10.LL in the formation of leaf lobes in A. thaliana. A Leaf phenotypes of the 35S:BjA10.LL^XX^–6HA overexpression transgenic and the wild type A. thaliana plants (Col-0). Scale bar: 1 cm. B PCR-based DNA genotyping of 35S:BjA10.LL^XX^ transgenic plants. Cas, cassette. C The expression levels of BjA10.LL in the fifth true leaves of the 35S:BjA10.LL^XX^–6HA overexpression transgenic and the wild type A. thaliana plants (Col-0). Values are means ± SD (n = 3). Double asterisks indicate that there are highly significant differences in the expression levels of the fifth true leaves between the 35S:BjA10.LL^XX^–6HA overexpression transgenic and the wild type A. thaliana plants (two-tailed paired Student's t-test, P ≤ 0.01). D The leaf dissection index of the fifth true leaves of the 35S:BjA10.LL.^XX^–6HA overexpression transgenic and the wild type A. thaliana plants (Col-0). The ‘XX’ of BjA10.LL means 9B18 and 9B16
Promoter activity assays of BjA10.LL
To gain deeper insights into the function of BjA10.LL and investigate whether sequence variations in its promoter region are associated with the formation of lobed-leaf margins, we cloned ~ 3 kb promoter fragments of BjA10.LL from both parental lines. Sequence alignment revealed significant differences in this region between 9B18 and 9B16 (Fig. S6).
To explore whether the sequence variations in the BjA10.LL promoter are responsible for the formation of leaf margin lobes, the promoter activities of BjA10.LL in 9B18 and 9B16 were tested via transient transcriptional activity assays. As shown, the promoter activity of BjA10.LL^9B16^ was significantly higher than that in BjA10.LL^9B18^ (Fig. 6).Fig. 6. Transient dual luciferase (dual-LUC) assays of BjA10.LL^9B18^ and BjA10.LL^9B16^ in tobacco leaves. A Sequence alignments of the ~ 3.0 kb region upstream of the start codon of BjA10.LL in 9B18 and 9B16. B Activity assays of the promoters of BjA10.LL^9B18^ and BjA10.LL.^9B16^ based on the dual luciferase system. C Quantification of the dual-luciferase assay. The relative LUC/REN activity ratio confirms that the lobed-leaf haplotype (9B16) possesses significantly stronger enhancer activity than the entire-leaf haplotype (9B18). Data are presented as mean ± SD (n = 5) (two-tailed paired Student's t-test, P ≤ 0.01)
To further validate that the sequence variation within this enhancer region is the causal polymorphism underlying natural variation in leaf morphology, we developed a co-dominant molecular marker. Primers were designed to flank two major indels within the ~ 500 bp enhancer of BjA10.LL, which effectively distinguished the parental haplotypes. Genotyping of the segregating F_2_ population revealed a perfect co-segregation between the marker haplotype and the leaf margin phenotype (Fig. 7). All plants carrying the lobed-leaf haplotype exhibited lobed leaves, whereas those for the entire-leaf haplotype displayed entire leaves.Fig. 7. Genotyping with the co-dominant molecular marker developed in the ~ 3.0 kb region upstream of the BjA10.LL start codon. A Schematic diagram of molecular marker-specific primers and corresponding sequence structure. B Genotyping results revealed by PCR amplification in the parental lines (9B16 and 9B18) and a subset of F_2_ plants displaying distinct leaf phenotypes. The PCR products were separated on a 2% agarose gel. M: Marker
This tight genetic linkage, together with the differential transcriptional activity demonstrated by the dual-luciferase assay, provides compelling evidence that the structural variation in the BjA10.LL enhancer region is responsible for the differential gene expression and, consequently, the divergence in leaf shape between the two parental lines.
Discussion
HD-ZIP I proteins have been demonstrated to play a crucial role in leaf development. For instance, transgenic plants overexpressing AtHB23, AtHB3, AtHB13, and AtHB20 exhibit significant changes during leaf growth and development (Wang et al. 2003; Henriksson et al. 2005; Kim et al. 2007). In B. napus, previous studies have identified that the HD-ZIP I transcription factors BnA10.RCO and BnA10.LMI1 are involved in regulating the formation of leaf margin lobation. In this study, we performed amino acid sequence alignment of the candidate gene BjA10.LL with these genes, revealing that only BnA10.RCO shares high homology with BjA10.LL. Functional annotation of the A. thaliana homologous gene further confirmed that BjA10.LL encodes an HD-ZIP I protein, consistent with previous findings.
Previous studies have shown that the RCO gene (but not LMI1) plays a pivotal role in leaf morphological development in Arabidopsis lyrata, C. hirsuta, and B. napus (Sicard et al. 2014; Vlad et al. 2014; Heng et al. 2020; Hu et al. 2020). In this study, we identified significant cis-regulatory differences between the parental alleles of BjA10.LL. Consistent with this finding, the promoter activity of BjA10.LL^9B16^ was significantly enhanced in the incised-leaf parent compared to the entire-leaf parent. Additionally, we observed variations in the enhancer region of the lobed-leaf parent, including six SNPs, one insertion, and two deletions. To genetically link these sequence variations to the phenotype, we developed a co-dominant molecular marker targeting two major indels within a core ~ 500 bp enhancer region. This marker exhibited perfect co-segregation with the leaf margin phenotype in our segregating F_2_ population, providing strong genetic evidence that these enhancer variations are indeed the causal polymorphisms responsible for differential gene expression and the consequent leaf shape determination. mVISTA alignment analysis of the RCO gene (BjA10.LL) in B. juncea with those in B. napus and C. hirsuta revealed high sequence conservation, including a conserved B region (Fig. S7). These results suggest that the function of the RCO gene in B. juncea may be similar to that in C. hirsuta. To validate this hypothesis, future studies should truncate the RCO gene and construct promoter deletion fragments of varying lengths combined with a reporter gene system to investigate the regulatory function of the B region on RCO gene expression. This research will help elucidate the expression regulation mechanism of the RCO gene in B. juncea and provide important insights into its molecular function in leaf morphogenesis.
Cytokinins, as one of the five major classes of plant hormones, play a crucial regulatory role in the growth and development of higher plants. Research has demonstrated that cytokinins are not only involved in regulating cell division and differentiation but also play key roles in various physiological processes, including organogenesis, apical dominance maintenance, and leaf senescence delay (Miyawaki et al. 2004). Particularly in the complex process of leaf margin development, cytokinins exert significant influence on leaf morphogenesis by maintaining the activity of meristematic tissues.
From a molecular perspective, the biosynthesis and metabolism of cytokinins are precisely regulated by multiple key genes. Among these, IPT7 is a critical gene in the cytokinin biosynthesis pathway. Studies in tomato (Solanum lycopersicum) have shown that overexpression of the A. thaliana AtIPT7 gene significantly increases cytokinin levels, thereby promoting leaf morphological complexity and leading to more pronounced serrated leaf margins (Dello Ioio et al. 2012). This finding has been further validated by reverse genetics experiments: when the cytokinin degradation gene CKX3 is heterologously expressed, cytokinin levels are significantly reduced, resulting in the disappearance of leaf lobation (Shani et al. 2010; Bar et al. 2017). In C. hirsuta, enhanced cytokinin activity in the RCO gene region has been shown to significantly promote the formation of leaf complexity (Hajheidari et al. 2019). These results collectively confirm the positive regulatory role of cytokinins in the formation of leaf margin lobation. Notably, the regulation of leaf margin morphology by cytokinins may involve a complex signaling network. Early research by Rupp et al. (1999) demonstrated that cytokinins significantly promote the expression of the KNOX family gene STM, which has been shown to enhance the development of complex leaf margins. In this study, we conducted transcriptome sequencing analysis on the entire-leaf margin parent 9B18 and the lobed-leaf parent 9B16, followed by GO functional enrichment analysis of differentially expressed genes. The results revealed significant enrichment of genes related to cytokinin biosynthesis, metabolism, and signal transduction. This finding further supports the potential key role of cytokinins in regulating leaf margin lobing morphogenesis.
Based on these results, future research could further explore the molecular mechanisms of cytokinins in leaf lobe formation, particularly their interactions with the regulatory network of the RCO gene. This will provide new theoretical insights and research directions for understanding the genetic basis of plant leaf morphological diversity.
Conclusion
This study revealed through genetic analysis that the lobed-leaf margin phenotype in B juncea is controlled by a single gene and exhibits incomplete dominance. Using map-based cloning techniques, we identified BjA10.LL as the key gene regulating the formation of lobed-leaf margins in B. juncea. Promoter activity assays demonstrated its positive regulatory role in this trait. Corresponding to the gene’s core enhancer variations, a co-dominant molecular marker targeting these indels was developed, which perfectly co-segregates with the phenotype and thus facilitates marker-assisted breeding of B. juncea.
Materials and methods
Plant materials
For the gene mapping of the lobed-leaf margin gene BjA10.LL in B. juncea, a F_1_, F_2_, BC_2_ segregation population was constructed using parental lines of B. juncea with entire-leaf margins (9B18) and lobed-leaf margins (9B16). These materials were planted at the experimental farm of Northwest A&F University (Yangling, Shaanxi, China) in the middle and late September each year from 2021 to 2024. Field management followed standard agricultural practices. At the five-leaf stage, the leaf types of both parental materials and the mapping population were recorded. Leaves were collected from individual plants, flash-frozen in liquid nitrogen, and transported to the laboratory for DNA extraction. The extracted DNA was used for subsequent gene mapping and cloning analyses. The parental lines (9B18 and 9B16) and their F_1_ hybrids were grown in a growth chamber at 22 °C under a long day duration (16 h light/8 h dark). Phenotypic observations were conducted at the five-leaf stage, and leaves from 20-day-old seedlings were sampled for scanning electron microscopy (SEM) analysis. A. thaliana (Col-0) was used as the wild type control in this study. All A. thaliana plants were grown in a growth chamber at 22 °C under long-day conditions. Tobacco plants were incubated in a growth chamber at 22 °C with a 16 h light/8 h dark photoperiod for 72 h.
Trait measurement
In order to quantify the degree and complexity of leaf lobing, the leaf was calculated according to (perimeter-squared)/(4π × area) (Hu et al. 2018).
To clarify the relationship between leaf margin cell characteristics and leaf shape, scanning electron microscopy was used to observe the leaf margin cells. Leaves smaller than 0.2 mm were collected from 20-day-old seedlings and fixed in FAA fixative solution for over 12 h. The samples were then dehydrated 1 to 2 times using a series of graded ethanol solutions, with each solution volume being at least three times that of the samples. After dehydration, the materials were dried using the critical point drying method. Subsequently, the dried samples were mounted onto copper stages with conductive adhesive, sputter-coated with gold, and observed under a scanning electron microscope (TM4000 Plus).
To evaluate the impact of the lobed-leaf margin trait on photosynthetic characteristics, PAR and Ci of 9B16 and 9B18 were measured in the field. The fifth leaves of 9B18 and 9B16 under consistent growth conditions were analyzed using a portable photosynthesis system (LI-6400XT; Li-COR Inc., Lincoln, NE, USA). Measurements were conducted between 9:00 and 11:00 in the morning (Sun et al. 2016). Three biological replicates were used, with each replicate consisting of 10 plants.
DNA extraction and SNP genotyping
For single-nucleotide polymorphism (SNP) genotyping (Brassica 50 K Illumina Infinium™ SNP), total genomic DNA was extracted from fresh leaves of the two parents (10 individual plants each for 9B18 and 9B16) and the F₂ population plants, including 30 plants with entire-leaf margins and 30 plants with lobed-leaf margins, using a plant genomic DNA extraction kit (Tiangen Biotech, Beijing, China) following the manufacturer’s protocol. Each bulk DNA sample was constructed with equivalent amounts of DNA from ten individuals of each parent and 30 individuals from the F₂ population. Finally, four bulk DNA samples, including the two parents, F₂ entire-leaf margin plants, and F_2_ lobed-leaf margin plants, were used for SNP genotyping with a Brassica 50 K Illumina Infinium™ SNP array, according to the method described by Xiao et al. (2021). Meanwhile, the gene sequences were subjected to BLAST alignment, and the resulting data were analyzed for synteny using TBtools software.
Molecular marker development, linkage analysis and locus mapping
Based on the genome assemblies of the Brassica database (BRAD: http://brassicadb.cn/#/) (Wang et al. 2011; Chen et al. 2022), molecular markers including SSR and IP in the BjA10.LL locus region, were developed. The SSR primers were designed using the web-based SSR finder tool (http://www.geboc.org/index/) Meanwhile, the IP primers were designed using Primer 5.0 software (http://www.premierbiosoft.com/primerdesign/). For functional characterization of BjA10.LL, specific primers were designed for promoter and CDS cloning. The sequence information of candidate genes is provided by the BRAD (http://brassicadb.org/brad/) and TAIR (http://www.arabidopsis.org) databases. Primers were designed based on standard criteria using the Primer-BLAST suite on the NCBI platform (https://www.ncbi.nlm.nih.gov/tools/primer-blast/). The resulting primer sequences were commercially synthesized by Sangon Biotech (Shanghai, China). The primers used are detailed in Supplemental Table S1.
Candidate gene analysis
Extract the gene annotation information within the 995 kb interval of BjA10.LL from the Brassica database and the TAIR database. We further screened the candidate genes by analyzing the transcriptome data between the two parents. Using the Brassica database (http://brassicadb.org/brad/) as the reference genome, the following bioinformatics analysis pipeline was employed, including sequencing data quality assessment, reference sequence alignment analysis, gene expression analysis, screening of DEGs, and Gene Ontology (GO) enrichment analysis of the DEGs. During the screening of differentially expressed genes, the criteria were set as |Log_2_ (Fold Change)|≥ 2 and FDR < 0.05. The screened DEGs were subsequently submitted to the Bioinformatics Online Tool (https://www.bioinformatics.com.cn/) for GO enrichment analysis, and significantly enriched GO terms were identified. Finally, we predicted the candidate genes by combining sequence alignment analysis, transcriptome analysis and the functional annotations of the Arabidopsis homologs of the candidate genes.
Sequence alignment analysis of BjA10.LL
The amino acid sequences of candidate genes and their homologous genes were retrieved from the B. juncea database (BRAD: http://brassicadb.cn/#/) and the B. napus database (BnIR: http://yanglab.hzau.edu.cn/), respectively. Sequence alignment was then performed using DNAMAN software.
RNA extraction and quantitative real‑time PCR (RT‑qPCR) analysis
Total RNA samples were isolated from various vegetative tissues of plants, leaves at different stages, and different segments of the fifth leaf of parents during the rosette stage using the MiniBEST Plant RNA Extraction Kit (TaKaRa Bio, Dalian, China). Subsequently, the RNA was reverse-transcribed using PrimeScript RT Mix (TaKaRa Bio, Dalian, China). Quantitative real-time PCR was performed with three biological replicates using SYBR Green Master Mix (Cofitt, Hong Kong, China). BjuUBQ9 and AtEF1aA4, ubiquitously expressed genes, were used as internal controls. All measurements were conducted with three independent biological replicates and two technical replicates. The primers used for gene expression analysis are listed in Table S1.
Subcellular localization analysis of BjA10.LL
The 35S:BjA10.LL^9B18^–GFP and 35S:BjA10.LL^9B16^–GFP constructs were separately transformed into Agrobacterium tumefaciens strain GV3101 and transiently expressed in the young leaves of the transgenic tobacco with nuclear localization signal as previously described (Yang et al. 2010).
Plant transformation
The constructs of 35S:BjA10.LL^9B18^–6HA and 35S:BjA10.LL^9B16^–6HA were transformed into the A. thaliana wild type (Col-0) separately using the floral dip method mediated by A. tumefaciens strain GV3101 to generate transgenic A. thaliana plants (Clough and Bent 2008).
Promoter activity analysis of BjA10.LL
To analyze the promoter activity of BjA10.LL, specific amplification primers were designed. Approximately 3 kb promoter sequences of BjA10.LL were cloned from the genomic DNA of 9B18 and 9B16. The amplified products were subsequently inserted into the pGreenII 0800–LUC vector, generating the recombinant constructs. These constructs were then transferred into A. tumefaciens strain GV3101 with the pSoup-P19 (Weidi Biotechnology, Shanghai, China). The Agrobacterium culture was mixed with the recombinant vectors at a volume ratio of 1:1 and co-infiltrated into the young leaves of 4-week-old tobacco plants. After 72 h of cultivation, the infiltrated leaves were collected, and LUC signals were detected using a low-light cooled charge-coupled device imaging apparatus (Tanon 4600, Shanghai, China). Firefly LUC and Renilla luciferase (REN) activities were examined using the dual-luciferase reporter assay kit (YEASEN, Shanghai, China) according to the instructions. Five independent biological samples were assayed. The experiment was performed with five independent biological replicates, each consisting of three technical replicates.
Supplementary Information
Additional file 1: Fig. S1 The characteristics of the leaf lobes of entire-leaf blades (9B18), lobed-leaf blades (9B16) and their F_1_ hybrid leaves. Fig. S2 Verification results of molecular markers in the parental plants and individual plants with entire and lobed leaves in the F_2_ generation. Fig. S3 Alignment of amino acid sequences of candidate genes. Fig. S4 RT-qPCR validation of some differentially expressed genes in the transcriptome. Fig. S5 Sequence analysis of BjA10.LL. Fig. S6 Sequence alignments of the ~ 3.0 kb upstream of the start codon of BjA10.LL in 9B18 and 9B16. Fig. S7 mVISTA graph of the alignment of the upstream sequence of the BjA10.LL gene in B. juncea with the RCO genes in B. napus and of C. hirsuta.Additional file 2: Table S1. Sequences of primers. Table S2. Polymorphism SNP_S_ between the entire-leaf samples and the lobed-leaf samples obtained by Brassica 50K Illumina Infnium™. Table S3. Collinearity analysis of chromosomal locations. Table S4. The information of 16 genes in the target interval.