Integrative taxonomic revision of three Camellia species from section Tuberculata (Camellia, Theaceae) by morphological, anatomical, palynological, and molecular evidence
Weihao Gu, Mingtai An, Chao Yan, Xu Xiao, Zhaohui Ran, Zhi Li

TL;DR
This study clarifies the taxonomy of three Camellia species using morphological, anatomical, and molecular evidence.
Contribution
The study provides a framework for species delimitation in morphologically complex plant groups using integrative taxonomic methods.
Findings
Camellia zengii is confirmed as a synonym of C. lipingensis.
Camellia rhytidocarpa is recognized as a distinct species.
Molecular and morphological data consistently support the revised taxonomy.
Abstract
The section Tuberculata (Camellia L.) comprises 18 species, forming a monophyletic group with unique “tuberculate-wrinkled fruit pericarp” morphological characteristics. However, the interspecific relationships within this section remain poorly resolved. A notable taxonomic controversy involves Camellia lipingensis, C. zengii, and C. rhytidocarpa, which were previously considered conspecific. On the basis of extensive population surveys conducted in their type localities, we identified significant morphological disparities among these three taxa. To comprehensively clarify their taxonomic status and relationships, we conducted an integrated study incorporating morphology, micromorphology (leaf epidermis and pollen), and molecular systematics (cpDNA and nrDNA ITS). Evidence from morphology, anatomy, palynology, and molecular systematics consistently supports the treatment of Camellia…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 10
Figure 10 Figure 11
Figure 11 Figure 1
Figure 1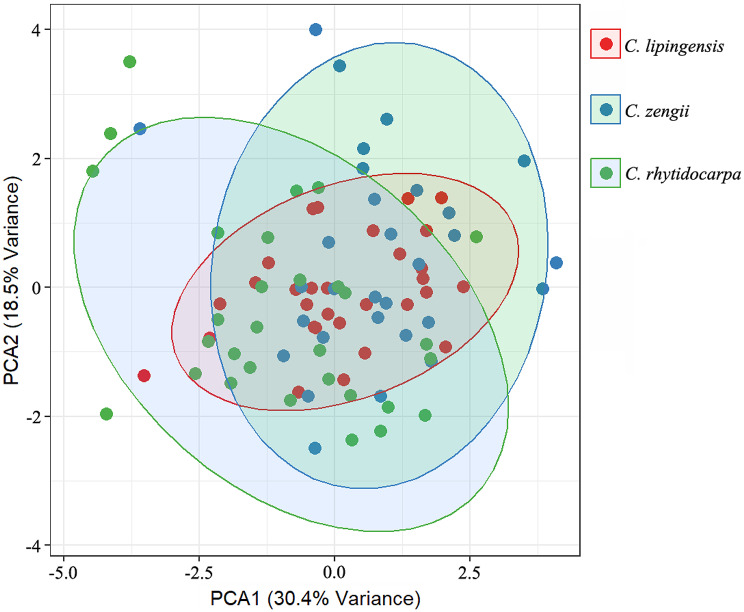 Figure 2
Figure 2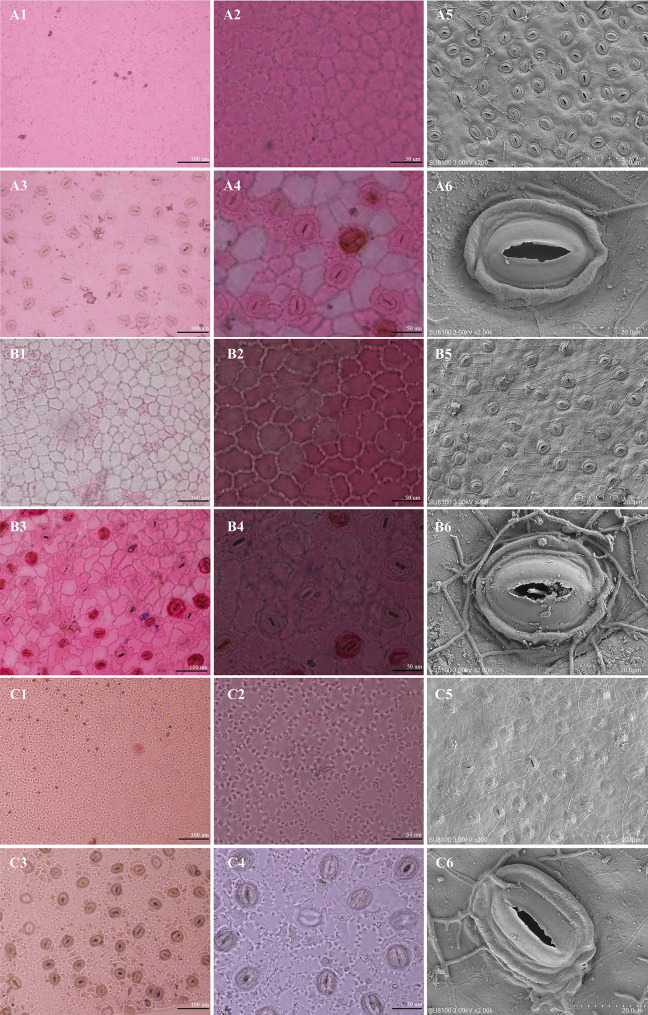 Figure 3
Figure 3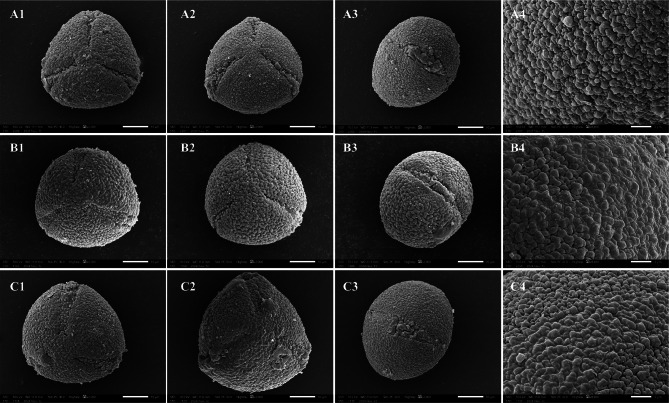 Figure 4
Figure 4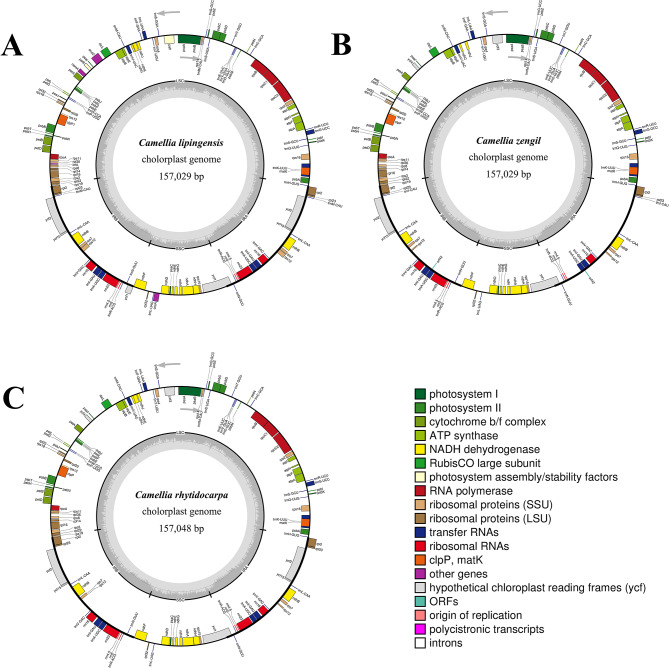 Figure 5
Figure 5 Figure 6
Figure 6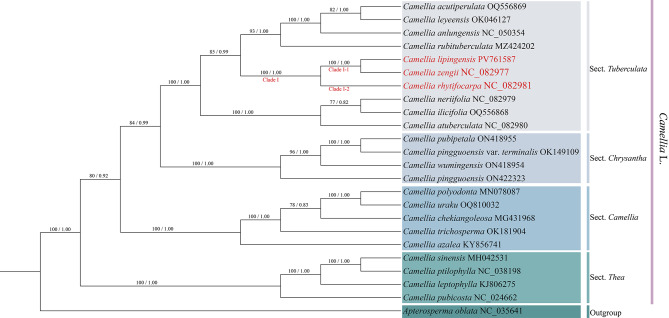 Figure 7
Figure 7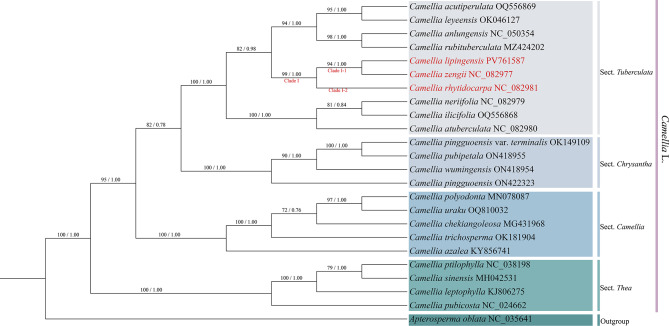 Figure 8
Figure 8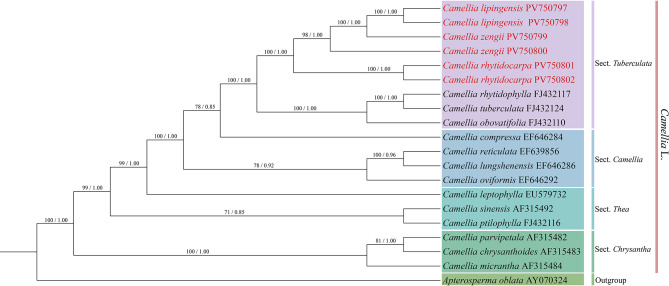 Figure 9
Figure 9- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
- —the 2024 Guizhou Science and Technology Innovation Talent Team Construction Project: Wildlife Innovation Team of the Forestry college of Guizhou University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and Fungal Species Descriptions · Plant Diversity and Evolution · Plant and animal studies
Introduction
Section Tuberculata of the Camellia genus is distinguished from other Camellia species by its unique “tuberculate-wrinkled fruit pericarp” (Chang and Ren 1991; Min and Zhong 1993; Chang 1998). In the early 20th century, the renowned botanist, Prof. Chien (1939), discovered a taxon with tuberculate ovaries and pericarps during fieldwork in Jiading district, Sichuan province, China, and named it Camellia tuberculata S.S. Chien. Sealy (1958) later placed it in section Heterogenea on the basis of a literature review. It was not until 1981 that Hung-Ta Chang established the sect. Tuberculata, which is based on the diagnostic “tuberculate-wrinkled fruit pericarp” that initially included 6 species, with 12 additional species reported over the next decade. In 1991, Chang subdivided the section into two subsections (subsect. Tuberculata Chang and subsect. Nudicarpa Chang) on the basis of ovary pubescence. Tianlu Min (1993), a famous Camellia taxonomist, revised this classification process and divided 18 species into 6 species, 6 varieties, and 1 form (Min and Zhong 1993). Notably, the taxonomic treatments proposed by these two researchers were developed under the methodological constraints of their time, relying principally on herbarium specimens with limited field validation and minimal integration of multidisciplinary taxonomic evidence. As a result, the infraspecific classification within sect. Tuberculata has remained taxonomically controversial.
The complex species comprises closely related taxa that share morphological, genetic, or ecological similarities and often have ambiguous taxonomic boundaries (He et al. 2022). These complexes typically include subspecies or varieties or may involve hybridization, incomplete lineage sorting, or cryptic diversity—factors that complicate species delimitation using traditional morphology. Camellia lipingensis, C. zengii, and C. rhytidocarpa were first described by Hung-Ta Chang in 1984 (Chang and Ren 1996). The types of C. lipingensis and C. zengii are confined to Wulong Mountain in Liping county, Guizhou province, China, where they grow in the natural habitats of mixed forests or bamboo groves. C. lipingensis is distinguished by narrow lanceolate, thick, leathery leaves, glabrous styles, and 5 cm-diameter flowers. C. zengii features villous styles, stamens 1.8–2.4 cm long, 10 segments, and sepals 1–4 cm long. The type specimen of C. rhytidocarpa, from Tianping Mountain in Longsheng county, Guangxi province, China, grows on valley areas in the understory of forests. In addition, the straight-line distance between the origins of the three varieties exceeds 200 km. C. rhytidocarpa has lanceolate to oblong leaves (8–12 cm long) with 6–7 pairs of unsunken lateral veins. A previous taxonomic study merged these three species into C. rhytidocarpa (Min et al., 1993), citing shared traits such as narrowly lanceolate leaves with caudate or acuminate apices, ovate sepals and ovaries, and sparsely pubescent trilobed styles. However, this review relied exclusively on herbarium specimens, with minimal field observation. Moreover, supporting evidence from disciplines such as molecular phylogenetics, palynology, or anatomical morphology is lacking. In recent decades, Camellia taxonomy has increasingly incorporated pollen micromorphology, cpDNA, and leaf epidermal features for these species (Jiang et al. 2010; Yan et al. 2024; Ran et al. 2024c).
In recent years, our extensive field investigations at the type localities of these three species revealed consistent disparities in key taxonomic traits of flowers and fruits, indicating that they should not be treated as conspecific. These initial findings prompted a comprehensive, multidisciplinary study integrating detailed morphological (including leaf epidermis and pollen morphology), anatomical, and molecular phylogenetic (cpDNA and ITS) analyses. The congruent evidence from these approaches consistently supports the recognition of three distinct species. Herein, we present a detailed taxonomic revision based on this integrative evidence.
Materials and methods
Observation and inspection of type specimens
Type specimen examination was systematically conducted to clarify species boundaries and verify historical taxonomic foundations, utilizing digitized resources from the Herbarium of the Institute of Botany, Chinese Academy of Sciences (PE; http://pe.ibcas.aCamelliacn/peweb/), Sun Yat-sen University Herbarium (SYS; https://eco.sysu.edu.cn/platforms/museum), and the Chinese Virtual Herbarium (CVH; https://www.cvh.ac.cn/index.php). Morphological parameters were rigorously measured using standardized techniques (Ran et al. 2024a), with type specimens of Camellia lipingensis, C. zengii, and C. rhytidocarpa sourced through an original literature review and authoritative digital platforms, including CVH and the National Specimen Resource Sharing Platform (NSII; http://www.nsii.org.cn/2017/home.php) (Wu et al. 2022; Seo et al. 2025). Comprehensive morphological observations and comparative analyses were performed on all the examined samples.
Material collection
The field materials used for experimental and morphological data measurements in this study were collected through systematic field surveys between 2022 and 2024. Fresh leaves and floral organs collected in the field were preserved via dry ice flash-freezing to ensure sample integrity (Xiao et al. 2024). Voucher specimens were processed following international herbarium standards and deposited in the Herbarium of the Forestry College, Guizhou University (Camellia lipingensis: LZ-20231126-7; Camellia zengii: LZ-20231127-3; Camellia rhytidocarpa: LZ-20240114-4).
Morphology study
In this study, plant morphological characteristics were systematically investigated through integrated herbarium specimen examinations and field observations, and natural habitats and overall plant morphology were documented before standardized measurements of qualitative and quantitative traits from ≥ 10 voucher specimens per species were conducted. The comprehensive measurements included stem/branch bark coloration and texture, leaf characteristics, floral features, and fruit attributes, with all the parameters measured in triplicate and averaged (Ran et al. 2024b). The standardized dataset (Table S1) was processed using Excel 2025 and subsequently analyzed through principal component analysis (PCA) of 35 morphological variables in R v4.4.2 (R Core Team 2020).
Leaf epidermis and pollen micromorphology
Fresh leaves and floral buds of Camellia lipingensis, C. zengii, and C. rhytidocarpa were collected for micromorphological analysis. The leaf samples were processed as follows: (1) The samples were fixed in FAA (70% ethanol: acetic acid: formaldehyde = 90:5:5, v/v), rinsed, dissected along the midvein (0.5 cm²), treated with sodium hypochlorite at 30 °C, and sectioned. Epidermal sections were stained with acetate fuchsin and observed under a light microscope to examine the periclinal walls and stomatal apparatus (Jiang et al. 2022; Situngu et al., 2022). (2) The leaf tissue was rinsed with PBS, fixed in EM fixative (2 h, RT), and stored at 4 °C. The tissue blocks were subsequently washed three times in 0.1 M PB (pH 7.4, 15 min each), postfixed in 1% OsO₄ (1–2 h, RT), subsequently washed, dehydrated in an ethanol series, critical-point dried, mounted on aluminum stubs, sputter-coated with gold (30 s), and imaged by SEM (Zhang et al. 2021; Neilands et al. 2023).
Pollen grains (10 per species) from FAA-fixed anthers were acetolyzed, released via microdissection, coated with gold, and examined by SEM (JSM-6490LV). Morphology size (polar/equatorial axes), colpi, exine ornamentation, and wall sculpturing were quantitatively analyzed using ImageTool software (Ocaña et al., 2022; Pan et al. 2022).
Total DNA extraction, sequencing, assembly, and annotation
Fresh leaves of the three species collected during field surveys were subjected to total DNA extraction using the cetyltrimethylammonium bromide (CTAB) method (Chai et al. 2023; Xing et al. 2023). Following Illumina DNA library preparation standards, paired-end sequencing libraries were constructed with 350-bp insert fragments (Li et al. 2021). DNA quality and concentration were assessed by agarose gel electrophoresis. Qualified fragments were sequenced on the Illumina platform after random fragmentation, end repair, and adapter ligation (Zong et al. 2023).
Raw reads from base-calling (CASAVA) were assembled using SPAdes v.3.15.2 and annotated with CPGAVAS and ORFFinder, with validation via BLASTN/BLASTP alignments. tRNA genes were annotated with ARWEN v1.2 (Zheng et al., 2023; Gu et al. 2024). The chloroplast genome was finalized through online annotation, BLAST alignment, and manual curation using C. rubituberculata Chang & Yu (MZ424202) as a reference genome (Xiao et al. 2025). Genome maps were generated with OGDRAW (Yan et al. 2023; Ran et al. 2024c). The sequences were deposited in NCBI GenBank to obtain accession numbers.
IR boundary expansion and contraction
The chloroplast genome sequences of Camellia lipingensis, C. zengii, and C. rhytidocarpa were screened, and three complete chloroplast genomes were obtained from the samples. The sequences of Theaceae species were downloaded from the NCBI for Biotechnology Information database. IR boundary expansion/contraction analysis was performed, and comparative maps were generated using IRscope (https://irscope.shinyapps.io/irapp/) (Amiryousefi et al. 2018; Xiao et al. 2024).
Analysis of adaptive nucleotide substitution rates in three species
To assess the selective pressure on the protein-coding genes in the chloroplast genomes of the three species, we calculated their nonsynonymous (Ka) and synonymous (Ks) substitution rates, along with the Ka/Ks ratio, using KaKs_calculator v3.0 (Gutiérrez et al. 2023). The analysis was performed under the MYN model (Zhou et al. 2023). A Ka/Ks ratio > 1 indicates positive selection, 1 suggests neutral evolution, and < 1 reflects purifying selection (Zhang et al. 2006).
Phylogenetic analysis
The chloroplast genomes of 24 Camellia species were retrieved from NCBI, with Apterosperma oblata (NC_035641) used as the outgroup (Ran et al. 2024a). Sequence alignment was performed using MAFFT v7, followed by maximum likelihood (ML) tree construction in IQ-TREE v1.6.12. Manual refinement in MEGA X resulted in the selection of optimal substitution models (Smith et al. 2021). MrModeltest v2.3 identified (TVM + F + I) as the best-fit model, and Bayesian inference (BI) trees were reconstructed using MrBayes v3.2.7 (Catalano et al. 2025). Phylogenies based on protein-coding genes and ITS were congruent with those of whole chloroplast genomes. The final trees were visualized with iTOL v5 (Letunicet al. 2021).
Results
The morphological characteristics of the three species
Systematic morphological comparisons revealed significant phenotypic variations in morphology among the three species (Fig. 1; Table 1). Camellia lipingensis and C. zengii shared brownish mottled bark exfoliating to green/grey-green surfaces, red-brown quadrangular branchlets, and leathery lanceolate leaves (vs. occasionally oblong in C. rhytidocarpa) with acute apices and sparse serrations. The leaf dimensions (6.66–12.48 × 2.16–3.93 cm vs. 5.81–11.60 × 1.17–4.10 cm) and petiole lengths (0.12–1.10 cm vs. 0.30–0.91 cm) overlapped. Floral traits included nearly identical petal dimensions (3.44–5.20 × 1.30–2.00 cm vs. 3.47–4.71 × 1.34–2.21 cm), numbers (6–9 vs. 7–8), ovate sepals, and overlapping ovary/style pubescence. Fruit diameter (2.24–3.15 cm vs. 2.26–3.18 cm) and shape (subglobose) were consistent, with only minor pericarp thickness variation (0.45–0.60 cm vs. 0.43–0.52 cm) falling within intraspecific geographic variation.
Camellia rhytidocarpa and C. lipingensis exhibited morphological differences. A comparison of the dimensional measurements of its occasional oblong leaves and acuminate apices revealed distinct differences in leaf dimensions (6.42–12.50 × 2.16–4.21 cm vs. 6.66–12.48 × 2.16–3.93 cm). The relatively dense serration density of C. rhytidocarpa leaves may reflect a local adaptation. Floral traits partially overlapped: Petal dimensions (3.20–4.28 × 1.25–1.90 cm vs. 3.44–5.20 × 1.30–2.00 cm), and obtuse sepal apices exhibited intraspecific polymorphism. Fruit diameter (2.24–3.09 cm vs. 2.24–3.15 cm) and shape (subglobose/oblate) were similar, although pericarp thickness was reduced (0.30–0.49 cm), suggesting developmental plasticity. The three species exhibit distinct morphological differences with continuous variation, with C. rhytidocarpa being the most abundant, exceeding both C. lipingensis and C. zengii.
Fig. 1. The main morphological characteristics of the three species (A: C. lipingensis; B: C. zengii; C: C. rhytidocarpa. 1: Wild-type habitat, 2: Flower, 3: Fruit, 4: Petals, 5: Calyx, 6: Mature fruit, 7: Seeds, 8: Filaments, 9: One-year branches, 10: Front and back of the leaf)
Table 1. Comparison of the main morphological characteristics of the three species of sect. TuberculataSpecies C. lipingensis
C. zengii
C. rhytidocarpa TrunkBrownish with mottled exfoliation, revealing glaucous green trunks after bark sheddingMottled brown with inconspicuous and exfoliating rugose barkRusset bark with conspicuous exfoliation, revealing glaucous green trunksBranchesAnnual branchlets tetragonally ribbedAnnual branchlets tetragonally ribbedAnnual branchlets tetragonally ribbedLeaf typeLeathery, lanceolate, leaf tail acuteLeathery, lanceolate, leaf tail acuteLeathery, lanceolate or long elliptic, acuminate leaf tailLeaf length × width (cm^2^)6.66–12.48 × 2.16–3.935.81–11.60 × 1.17–4.106.42–12.50 × 2.16–4.45Petiole length (cm)0.12–1.100.30–0.910.43–1.30Number of petals6–97–86–8Number of calyxes6–97–86–8SepalOvate, apex roundedOvate, few rounded, apex roundedRounded, apex roundedFruit diameter (cm)2.24–3.152.26–3.182.24–3.09Fruit shapeSubglobularSubglobularOblateShell thickness (cm)0.45–0.600.43–0.520.30–0.49
To assess the taxonomic value of the morphological traits across the three species, we performed a principal component analysis (PCA). The results clearly separated all the samples into three clusters. While C. lipingensis, C. zengii, and C. rhytidocarpa exhibited substantial morphological overlap, with measured traits reflecting a nested relationship among them, C. rhytidocarpa displayed the greatest variation (Fig. 2).
Fig. 2. Principal component analysis of the morphological characteristics of the three species. Confidence ellipses indicate the 95% normal distribution range for each group
Leaf epidermal micromorphological characteristics
Microscopy revealed (Fig. 3; Table 2) highly conserved yet continuously variable leaf micromorphology across the three species. Camellia lipingensis, C. zengii, and C. rhytidocarpa shared irregular epidermal cells with sinuous anticlinal walls on adaxial surfaces. Crucially, oil glands were absent in C. lipingensis and C. zengii but were sparse in C. rhytidocarpa.
Abaxially, Camellia lipingensis and C. zengii exhibited sinuous anticlinal walls versus undulate walls in C. rhytidocarpa, although all the species presented a tight cell arrangement and anomocytic stomata with reniform guard cells and 3–5 similarly configured subsidiary cells.
Fig. 3. Micromorphological characteristics of the leaf epidermis of three species (A: C. lipingensis; B: C. zengii; C: C. rhytidocarpa. 1, 2: Adaxial surface; 3, 4: Hypodermis; 1, 3: 20×; 2, 4: 40×; 5, 6: stomatal apparatus)
Table 2. Morphological characteristics of the leaf epidermis of the three speciesSpeciesShape of stomataSize of stomata (length/µm × width/µm)Length / WidthInner margin of outer rimStomatal density (stomata/mm²)Type of stomatal apparatus C. lipingensis Long ellipse40.8–40.9 × 32.5–36.51.12–1.25Shallow waveform68–86Oval C. zengii Long ellipse39.9–40.8 × 31.9–36.71.10–1.26Shallow waveform65–85Oval C. rhytidocarpa Long ellipse40.1–41.5 × 31.4–36.71.14–1.22Shallow waveform62–78Oval
Camellia lipingensis and C. zengii shared elliptical stomata with fully overlapping dimensions: Length (40.8–40.9 μm vs. 39.9–40.8 μm), width (32.5–36.5 μm vs. 31.9–36.7 μm), length-to-width ratio (1.12–1.25 vs. 1.10–1.26), and density (68–86 vs. 65–85 stomata /mm²). Both exhibit sinuous inner periclinal walls.
The morphology of Camellia rhytidocarpa and C. lipingensis showed morphological continuity: The stomatal length (40.1–41.5 μm vs. 40.8–40.9 μm) differs slightly, but width (31.4–36.7 μm vs. 32.5–36.5 μm) and L/W ratio (1.14–1.23 vs. 1.12–1.25). Differences in density were detected (62–78 vs. 68–86 stomata/mm²) (Table S2).
The microscopic morphological characteristics of the pollen of the three species
Palynological evidence (Table 3; Fig. 4) revealed nearly identical pollen characteristics in Camellia lipingensis and C. zengii. Both species produce subspherical pollen with partially overlapping polar axis (36.7–36.8 μm vs. 36.8 μm) and equatorial axis lengths (40.3 μm vs. 41.3 μm). The P/E ratio (0.90 vs. 0.9) showed complete convergence, whereas the colpus dimensions exhibited minimal divergence: Length (26.3–26.4 μm vs. 26.3 μm), width (7.3 μm vs. 7.4 μm), and a mean L/W ratio of 3.6. Identical granular exine ornamentation and muri-granular complexes further support morphological continuity.
Compared with C. lipingensis and C. zengii, Camellia rhytidocarpa exhibits continuous clinal variation in pollen morphology but maintains substantial overlap. Compared with that of C. zengii (36.8 μm), the oblate-spheroidal pollen of C. rhytidocarpa has a slightly longer polar axis (37.2–37.8 μm). In contrast, its equatorial axis (41.3–41.4 μm) and P/E ratio (0.7 vs. 0.9) demonstrate inconsistent variation relative to those of the other taxa. While the granular exine ornamentation is semblable, differences in cell wall thickness (0.5 μm in C. rhytidocarpa versus 0.5–0.6 μm in C. zengii) further indicate ultrastructural differentiation among these species.
Table 3. Pollen morphological characteristics of the three speciesSpeciesPollen shapePolar axis (P)/µmEquatorial axis (E)/µmPolar axis/equatorial axisGermination groove/µmLengthWidthLength/Width (average data) C. lipingensis Near-spherical shape36.7–36.840.30.926.3–26.47.33.6 C. zengii Near-spherical shape36.841.30.926.37.43.6 C. rhytidocarpa Near-spherical shape37.2–37.841.3–41.40.727.7–27.77.43.3
Fig. 4. Electron microscopy images of the three species. (A: C. lipingensis; B: C. zengii; C: C. rhytidocarpa)
Characteristics of the chloroplast genome
Integrated molecular analyses (Table 4; Fig. 5) revealed high chloroplast genome conservation among Camellia lipingensis, C. zengii, and C. rhytidocarpa. The genome sizes were 157,029 bp (C. lipingensis/C. zengii) and 157,048 bp (C. rhytidocarpa), with identical GC contents (37.3%). LSC/SSC/IR lengths were conserved in C. lipingensis and C. zengii (LSC 86,630 bp, SSC 18,281 bp, IR 52,118 bp), whereas C. rhytidocarpa showed minimal LSC expansion (86,648 bp) and stable SSC (18,282 bp).
The regional GC content exhibited negligible variation: The LSC ranged from 35.31 to 35.32%, the SSC ranged from 30.61%, and the IR ranged from 42.94 to 42.97%. The gene inventories were identical: 132 total genes (87 protein-coding, 37 tRNA, and 8 rRNA). Slight divergence occurred only in the third-codon position GC content (29.40-29.47%). IR boundary analysis confirmed conserved gene positioning (rpl22, rpl2, psbA). The expansion of the LSC in C. rhytidocarpa may represent potential ecological adaptation.
Table 4. Genome-wide characteristics of chloroplasts in three speciesSpecies C. lipingensis
C. zengii
C. rhytidocarpa Genome size (bp)157,029157,029157,048GC (%)37.337.337.3LSC size (bp)86,63086,63086,648SSC size (bp)18,28118,28118,282IR size (bp)52,11852,11852,118GC in LSC (%)35.3235.3235.31GC in SSC (%)30.6130.6130.61GC in IR (%)42.9442.9442.97GC in CDS (%)37.6137.6137.611st position GC (%)45.2145.2145.372nd position GC (%)37.937.938.043rd position GC (%)29.4729.4529.40Length of CDS79,10079,10079,100Number of genes132132132Number of CDS878787Number of tRNA373737Number of rRNA888
Fig. 5. Chloroplast genome maps of the three species. (A: C. lipingensis; B: C. zengii; C: C. rhytidocarpa)
IR boundary contraction/expansion analysis (Fig. 6) confirmed the conserved positioning and lengths of the rpl22, rpl2, and psbA genes in Camellia lipingensis, C. zengii, and C. rhytidocarpa. Minimal interspecific divergence occurred, with slight variations in the length of the LSC. Comparative assessment with C. rubituberculata and C. anlungensis revealed distinct IR boundary initiation patterns: The focal species initiated at the trnH gene, whereas C. rubituberculata and C. anlungensis initiated at tRNA genes. All twenty-four species exhibited similar contraction/expansion trends. Despite the conserved gene architecture, subtle interspecific differences indicate lineage-specific adaptations. These structural variations likely reflect ecological selection pressures, influencing genomic organization and functional expression.
Fig. 6. Analysis of the contraction and expansion of IR boundaries in twenty-four species
Selection of stress analysis results
A total of 93 protein-coding genes from the chloroplast genomes of Camellia lipingensis, C. zengii, and C. rhytidocarpa were analyzed to estimate nucleotide substitution rates. Using the MYN model in KaKs_calculator v3.0, we calculated non-synonymous (Ka) and synonymous (Ks) substitution rates and their ratio (Ka/Ks). Among the genes compared, eight yielded calculable results: accD, atpA, ndhC, ndhD, psaI, psaJ, psbA, and psbB. Two of these genes (psbA and psaJ) presented a Ka/Ks > 1, indicating positive selection (Table 5). These findings suggest limited evidence for ecological adaptation in these species in response to environmental pressures.
Table 5. Ka/Ks values for protein-coding gene alignments among the three speciesSpecies 1Species 2geneKaKsKa/Ks C. rhytidocarpa
C. zengii
accD 00.0030 C. lipingensis
C. rhytidocarpa
atpA 00.0030 C. rhytidocarpa
C. zengii
atpA 00.0030 C. rhytidocarpa
C. zengii
atpA 00.0030 C. rhytidocarpa
C. zengii
ndhC 00.0020 C. lipingensis
C. rhytidocarpa
ndhD 00.0020 C. lipingensis
C. rhytidocarpa
psaI 0.0010.0020.743 C. lipingensis
C. zengii
psaI 0.0010.0020.744 C. rhytidocarpa
C. zengii
psaJ 0.0200.0201.034 C. lipingensis
C. rhytidocarpa
psbA 0.0200.0201.034 C. lipingensis
C. zengii
psbA 0.0230.0230.988 C. lipingensis
C. zengii
psbA 0.0200.0201.034 C. lipingensis
C. rhytidocarpa
psbB 00.0190 C. lipingensis
C. zengii
psbB 00.0190
Phylogenetic analysis
Phylogenomic analysis (Fig. 7) revealed that the support rates of Camellia lipingensis, C. zengii, and C. rhytidocarpa were high for Clade I (ML = 100; BI = 1.00). Specifically, C. lipingensis and C. zengii formed a monophyletic clade in clade I-1 (ML = 100, BI = 1.00), whereas C. rhytidocarpa occupied clade I-2. Congruently, two phylogenetic trees based on protein-coding genes (Fig. 8) and ITS sequences (Fig. 9) consistently resolved all three species within sect. Tuberculata, clustering them on a shared branch with high statistical support.
Fig. 7. Construction of a phylogenetic tree based on the chloroplast genome (maximum likelihood (ML) and Bayesian (BI) trees; BS ≥ 50% and PP ≥ 0.95 are indicated above the branches as BS/PP)
Fig. 8. Construction of a phylogenetic tree based on the PCGs (maximum likelihood (ML) and Bayesian (BI) trees; BS ≥ 50% and PP ≥ 0.95 are indicated above the branches as BS/PP)
Fig. 9. Construction of a phylogenetic tree based on the ITS (maximum likelihood (ML) and Bayesian (BI) trees; BS ≥ 50% and PP ≥ 0.95 are indicated above the branches as BS/PP)
Taxonomic treatment
The comprehensive evidence from morphology, anatomy, palynology, and molecular phylogenetics all support the synonymization of Camellia zengii under C. lipingensis, with the latter name being retained in accordance with the principle of priority in botanical nomenclature. In contrast, C. rhytidocarpa should be considered a distinct species. (Fig. 10).
Fig. 10. Type specimen information of the three species. (A: C. lipingensis; B: C. zengii; C: C. rhytidocarpa. Figure B was obtained from the National Specimen Platform (http://www.nsii.org.cn/2017/home.php) and was found to be a type of specimen after verification
Camellia lipingensis Hung T. Chang, Acta Sci. Nat. Univ. Sunyatseni 23(2): 78. 1984.
Type: CHINA. Guizhou Province: Liping County, Wulong Mountain, s. Camellia 808, 81,064 (holotype SYS; isotype GZFI).
= Camellia zengii Hung T. Chang, Acta Sci. Nat. Univ. Sunyatseni 23(2): 77. 1984. syn. nov.
Type: CHINA. Guizhou Province: Liping County, Wulong Mountain, s. Camellia 8017, 8018 (holotype GZFI; isotypes SYS).
Botanical description
Small macrophanerophytes with glabrous, lustrous young branches. Leaves thickly coriaceous, lanceolate, 6–12 cm long × 2–4 cm wide; apex caudate-acuminate, base broadly cuneate to rounded; adaxially greenish and slightly lustrous when dry, abaxially fulvous and glabrous except for sparse villous hairs along midvein; lateral veins 5–10 pairs prominently raised on both surfaces, reticulation obscure; margin sharply serrate; petiole 5–11 mm. Flowers white, terminal, subsessile, to 5 cm diameter. Outer bracts 4, scarious, apex apiculate and pubescent. Inner bracts ovate, 1–1.4 cm long, apex acute, margin scarious and densely pubescent. Petals 10, oblong to ovate, 3.5 cm long. Stamens 3 cm long, outer filaments nearly free. Ovary 3-loculed, densely pubescent; styles 3, 3.5 cm long, pubescent. Capsule oblate, tuberculate, 2.5–3 cm diameter, 3-loculed; seeds 1 per locule, densely tomentose. Flowering from August to September.
Distribution and habitat
C. lipingensis and C. zengii are endemic to Liping county, Guizhou province. Their typical habitats include Wulong Mountain (Qiutuan village), Cenba village, and Huangtianbang, where they thrive in mountain ecosystems at 1000–1200 m elevation, predominantly on slope gradients of 15–45 °C (Fig. 11).
Fig. 11. Map of collection loci of C. lipingensis, C. zengii and C. rhytidocarpa
Camellia rhytidocarpa Hung T. Chang & S. Y. Liang in Chang, Tax. Gen. Camellia: 49. 1981. syn. nov.
Type: CHINA. Guangxi Zhuang Autonomous Region: Longsheng County, Tianpingshan Village, s. Camellia 700, 908 (holotype SCBI; isotype PE).
Botanical description
A perennial shrub or small tree with glabrous young branches. Leaves thickly coriaceous, oblong or lanceolate, 6–12 cm long and 2.5–4.5 cm wide; apex acuminate, base obtuse to subrounded; adaxial surface olive-green when dried, glossy, abaxial surface yellowish-green, glabrous; lateral veins 6–7 pairs, slightly impressed adaxially and prominent abaxially; margin sharply serrulate, with teeth spaced 2–3 mm apart; petiole 8–12 mm, glabrous. Flowers terminal, white, sessile; bracts and sepals in 10 segments, gradually increasing in size, the outermost 3 semilunar, the inner 7 ovate, 1–1.4 cm long, sericeous abaxially; petals 6–8, 3.2–4.3 cm long, basally connate for approximately 2/5 of their length, free portions obovate, the outermost 1 or 2 petals slightly shorter and pubescent abaxially; stamens 2–2.2 cm long, outer whorl with filaments fused up to 1.3 cm at base, both free parts and filament tube glabrous, filament tube adnate to petals at base; ovary pubescent; styles 3–4, 2 cm long, glabrous. Capsule globose, tuberculate, 2–2.5 cm in diameter, 1-loculed with 1 seed, or occasionally bilobed and 2-loculed; seeds spherical, pubescent. Flowering occurs from November to December.
Distribution and habitat
Camellia rhytidocarpa is restricted to Tianping Mountain village, Longsheng County, Guangxi Zhuang Autonomous Region. This species occupies gentle valleys, forest understories, and riparian zones within the same altitudinal range (1000–1200 m) (Fig. 11).
Discussion
The taxonomic controversy surrounding Camellia lipingensis, C. zengii, and C. rhytidocarpa has long centered on intraspecific variation in floral and leaf morphology (Min et al., 1993; Chang 1996). In this study, morphological, anatomical, palynological, and molecular phylogenetic evidence is integrated with long-term field surveys and type specimen examinations to clarify their delimitation. These findings indicate that morphological traits such as leaf shape, petiole length, and fruit size are consistent with their classification as two distinct species. Such trait stability represents a phenotypic response to heterogeneous habitats in terms of plant taxonomy (Williams 2010). The consistent bark, branchlets, leaves, flowers, and fruits strongly support the merging of C. lipingensis and C. zengii. In contrast, C. rhytidocarpa remains distinct because of its leaf serration, shape, and thinner pericarp.
In terms of leaf epidermal micromorphology, Camellia rhytidocarpa has a sparse distribution of oil glands. Both C. lipingensis and C. zengii exhibit sinuous anticlinal walls, whereas C. rhytidocarpa displays undulate walls. The overlapping stomatal dimensions and shared sinuous wall structure strongly support the merging of C. lipingensis and C. zengii. In contrast, the morphological continuity and gland distribution patterns of C. rhytidocarpa substantiate its status as a distinct species. These micromorphological distinctions align with the recognition of C. rhytidocarpa as separate from the C. lipingensis-C. zengii complex. This conclusion is consistent with the principle that intraspecific variation manifests gradually, whereas interspecific differences remain discernible (Pinedo et al. 2016; Longhi et al. 2024). Additionally, stomatal micromorphology data corroborate Barrio’s (2023) finding that leaf morphological conservatism is significantly correlated with local environmental selection pressure.
Type specimen verification and wild population sampling are essential for species delimitation (Xue et al. 2018; Xu et al. 2023). In this study, when the digital specimen banks (such as CVH, GBIF, and Camellia) and the type of specimens in the collection were systematically verified, some information deficiencies were detected in the existing samples, such as the absence of complete morphological characteristics of flowers and fruits. Such deficiencies have historically led to overreliance on fragmented characteristics, neglecting population-level trait continuity (Bebber et al. 2010). By integrating voucher specimens with field-collected data, we reconstructed morphological profiles and taxonomic status for these taxa within sect. Tuberculata, as corroborated by Bossa-Castro et al.’s (2024) finding that > 30% of global herbarium specimens lack critical diagnostic traits, necessitating field validation.
The diversity of pollen morphology is the result of plants adapting to different environmental and ecological conditions over the course of long-term evolution (Mander et al. 2021). The continuous variation in the pollen polarization ratio and outer wall ornamentation (coarse net-ridge-papilla composite structure) of Camellia lipingensis, C∙ zengii, and C. rhytidocarpa is consistent with intraspecific pollen polymorphism (Gamal 2025), whereas the flattened spherical pollen and significantly reduced papilla density of C. rhytidocarpa point to an independent evolutionary path. This difference may reflect their ability to adapt to different pollination environments, with their distribution in drier, hotter regions potentially driving the restructuring of pollen outer wall structures. Additionally, the regular differences in the distribution of germination pores further reinforce their taxonomic independence (Amstutz et al. 2024).
Molecular phylogenetic evidence provides critical support for taxonomic revision (Wei et al. 2023; Abe et al. 2024; Qin et al. 2024). Chloroplast genome analysis revealed high sequence similarity between Camellia lipingensis and C. zengii, whereas notable differences distinguished them from C. rhytidocarpa. Phylogenetic reconstruction further revealed that C. lipingensis and C. zengii formed a well-supported clade (ML = 100; BI = 1.00), whereas wrinkled-fruit tea (C. rhytidocarpa) constituted a distinct lineage. These results support the recognition of C. lipingensis and C. zengii as conspecifics, with C. rhytidocarpa representing a separate species. Nevertheless, it is inferred that C. rhytidocarpa shares a closer phylogenetic affinity with both C. lipingensis and C. zengii in terms of early evolutionary history. These findings align with the view proposed by Chang et al. (2021) that chloroplast genomes remain largely conserved among closely related species. Despite the sympatric distribution of C. lipingensis and C. zengii, the nuclear ITS phylogeny confirms their genetic homogeneity, validating the reliability of multigene analysis in terms of taxonomy reported by Wang et al. (2025).
Geographic isolation and niche differentiation drive interspecific divergence (Lindelof et al. 2020). Camellia lipingensis and C. zengii cooccur in similar mountainous habitats (1000–1200 m) in Liping county, Guizhou, whereas C. rhytidocarpa is restricted to Longsheng county, Guangxi. The differences in leaf distribution points indirectly demonstrate the characteristic species differentiation within distinct environmental zones. Habitat heterogeneity likely drives morphological adaptations through natural selection. The PCA results indicate that while C. rhytidocarpa has slightly larger leaves and longer petioles (0.43–1.30 cm), its smaller corolla size significantly distinguishes it from both C. lipingensis and C. zengii. However, the floral traits, such as petal number and the presence of complete bisexual flowers, did not markedly differ between these two varieties. Environmental variation elicits differential phenotypic responses within species; e.g., acuminate leaf apices may reduce transpiration in drier microhabitats (Liu et al. 2025).
Molecular phylogenetic analyses revealed that the chloroplast genomes of Camellia lipingensis and C. zengii measure 157,029 bp, distinguishing them from that of C. rhytidocarpa (157,048 bp). C. lipingensis and C. zengii share identical LSC, SSC, and IR region lengths (86,630 bp, 18,281 bp, and 52,118 bp, respectively), whereas C. rhytidocarpa has a slightly expanded LSC region (86,648 bp). These findings align with earlier observations by Mondini et al. (2009), who reported that chloroplast genomic divergence among closely related species is generally subtle. In the phylogenetic tree, C. lipingensis and C. zengii formed a single clade, whereas C. rhytidocarpa formed a distinct subclade. This finding is consistent with the asynchrony of morphological-genetic differentiation demonstrated by Zou et al. (2021) in Rhododendron L., which suggests that phenotypic plasticity may play a dominant role in the formation of morphological differences.
Conclusion
This study pioneers an integrated multidisciplinary analysis of Camellia lipingensis, C. zengii, and C. rhytidocarpa, systematically resolving their morphoanatomical traits, palynological patterns, and molecular phylogenetics. This research revealed the synonymization of C. lipingensis and C. zengii while affirming C. rhytidocarpa as a distinct taxon. This revision resolves the long-standing synonymy question of the three species in sect. Tuberculata. The revealed morpho-molecular coevolution further provides a theoretical foundation for assessing intraspecific variation, genetic diversity patterns, and adaptive evolution in Camellia L.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
Supplementary Material 2
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Catalano SA, Escapa I, Pugh KD, Hammond AS, Goloboff P, Almécija S (2025) Place My Fossils: an integrative approach to analyze and visualize the phylogenetic placement of fossils using backbone trees. Systematic Biology Syaf 025. 10.1093/sysbio/syaf 025.10.1093/sysbio/syaf 025PMC 1264008040244059 · doi ↗ · pubmed ↗
- 2Chang HT (1998) Camellia. In: Fl. Reipubl. Popularis Sin. (eds) Flora. Science Press, Beijing. 37–48. https://www.iplant.cn/info/Sect.%20Tuberculata?t=z.
- 3Letunic I, Bork P (2021). Interactive Tree Of Life (i TOL) v 5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Research. 10.1093/nar/gkab 30110.1093/nar/gkab 301PMC 826515733885785 · doi ↗ · pubmed ↗
- 4Min TL, Zhong YC (1993) A revision of genus Camellia sect. Tuberculata. Acta Botanica Yunnanica 15:123–130
- 5Neilands J, Svensäter G, Boisen G, Robertsson C, Wickström C, Davies JR. Formation and analysis of mono-species and polymicrobial oral biofilms in flow-cell models. In: Nordenfelt P, Collin M, editors. Bacterial pathogenesis. Methods in Molecular Biology. Vol. 2674. New York (NY): Humana Press; 2023. 10.1007/978-1-0716-3243-7_2.10.1007/978-1-0716-3243-7_237258958 · doi ↗ · pubmed ↗
- 6Ran Z, Li Z, Xiao X, Tang M (2024 a) Camellia neriifolia and Camellia ilicifolia (Theaceae) as separate species: evidence from morphology, anatomy, palynology, molecular systematics. Botanical Studies 65:23. 10.1186/s 40529-024-00430-2.10.1186/s 40529-024-00430-2PMC 1126632539042341 · doi ↗ · pubmed ↗