Staphylococcus aureus Extracellular Vesicles Enhance PslE‐Mediated Pathogenesis in Pseudomonas aeruginosa
Phawinee Subsomwong, Rojana Sukchawalit, Naoko Watabe, Akio Nakane, Krisana Asano

TL;DR
Staphylococcus aureus extracellular vesicles increase Pseudomonas aeruginosa pathogenicity by enhancing PslE-related processes like biofilm formation and immune evasion.
Contribution
This study identifies PslE as essential for extracellular vesicle-mediated enhancement of Pseudomonas aeruginosa pathogenicity.
Findings
SaEVs upregulate pslE and related genes in P. aeruginosa, increasing LPS production and biofilm formation.
PslE is required for SaEVs to reduce macrophage uptake of P. aeruginosa.
Deletion of pslE abolishes the effects of SaEVs on pathogenicity traits.
Abstract
Coinfection of Pseudomonas aeruginosa (P. aeruginosa) and Staphylococcus aureus (S. aureus) is frequently observed. Our previous study demonstrated that S. aureus‐derived extracellular vesicles (SaEVs) promote P. aeruginosa pathogenicity by increasing lipopolysaccharide (LPS) production, promoting biofilm formation and decreasing the uptake of P. aeruginosa by macrophages. Proteomic analysis revealed that SaEVs enhance the production of PslE, an exopolysaccharide biosynthetic protein in P. aeruginosa, but the role of Psl exopolysaccharide polymerization on SaEV‐mediated P. aeruginosa pathogenicity is unclear. In this study, a pslE‐deletion mutant of P. aeruginosa (PaΔpslE) was constructed, and the effect of SaEVs on the pathogenicity of this mutant was evaluated. Our results showed that SaEVs significantly increased the expression of pslA, E, J, K, and L genes in the psl cluster of P.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
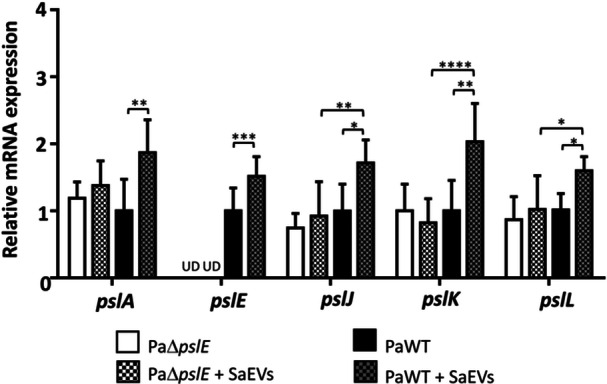 Figure 1
Figure 1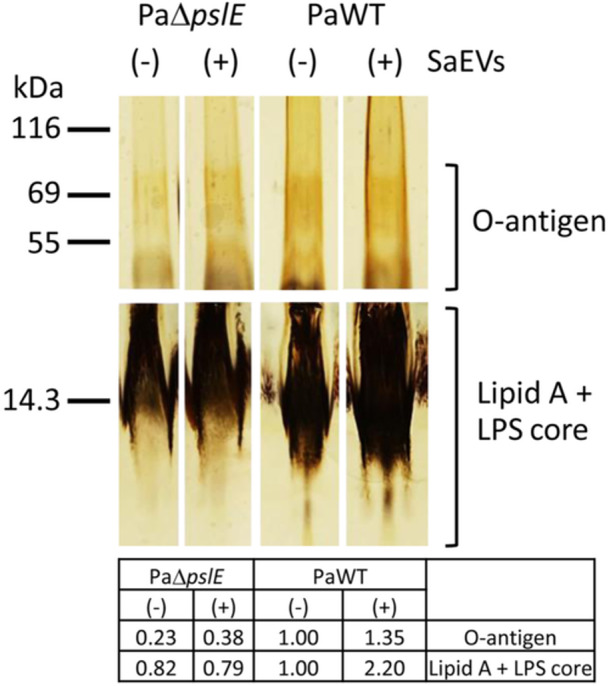 Figure 2
Figure 2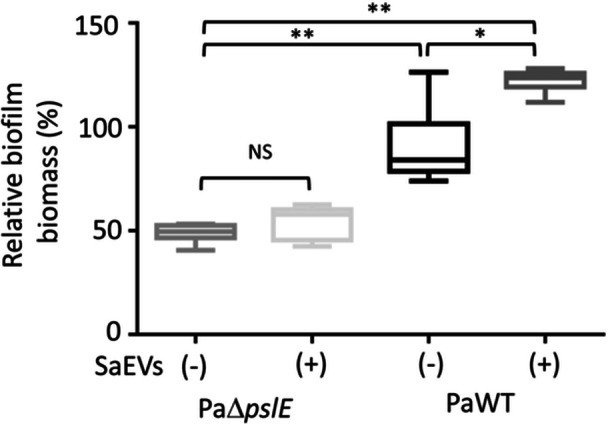 Figure 3
Figure 3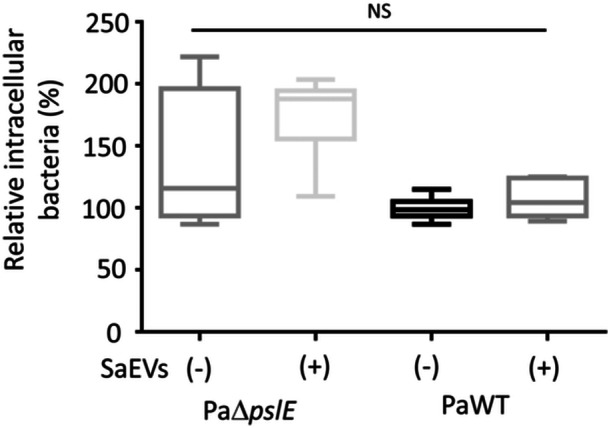 Figure 4
Figure 4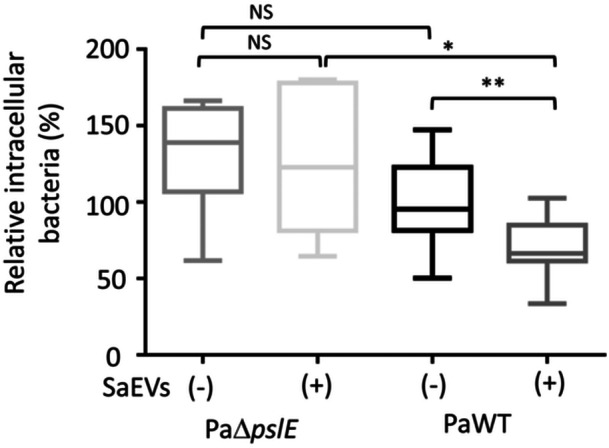 Figure 5
Figure 5- —This study was supported by Japan Society for the Promotion of Science KAKENHI (Grant Numbers 22K15454, 2022–2024, 23K06525, 2023–2025), the Research Center for GLOBAL and LOCAL Infectious Diseases, O
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Infections and Vaccines · Bacterial biofilms and quorum sensing · Neutrophil, Myeloperoxidase and Oxidative Mechanisms
Introduction
1
Pseudomonas aeruginosa (P. aeruginosa) and Staphylococcus aureus (S. aureus) are opportunistic pathogens that often coexist in various conditions, including in chronic wounds or cystic fibrosis (CF) lungs (Bernardy et al. 2022; DeLeon et al. 2014; Fugère et al. 2014). Longitudinal culture positivity for S. aureus and P. aeruginosa in children and adults with CF revealed that coinfection with these pathogens is common and increased from 30.6% to 50.7% over a 10‐year period (Fischer et al. 2021). Many reports have shown that coinfection with S. aureus and P. aeruginosa is more virulent than infection with a single species and worsens the prognosis of patient, especially when both species are multidrug‐resistant strains (Briaud et al. 2019, 2020; Filkins et al. 2015).
Several studies have indicated that bacteria in biofilms are more resistant to antibiotics and host immune defenses than planktonic bacteria (Thöming and Häussler 2022; Moser et al. 2017). P. aeruginosa biofilms are frequently found in permanent bladder catheter tubes, ventilator‐associated pneumonia tubes, lungs of patients with CF, and in chronic wounds (Costerton et al. 1999; Cangui‐Panchi et al. 2022; Werneburg 2022). Elimination of these biofilms is difficult because the extracellular polymers within the biofilms inhibit the penetration of antimicrobial agents. Therefore, repeated exposure to high concentrations of antibiotics is required to eliminate biofilms, which increases the risk of the emergence of antibiotic‐resistant strains (Srinivasan et al. 2021; Pinheiro et al. 2014; Dan et al. 2023; Liu et al. 2022). Biofilms are composed of microbial cells and extracellular polymeric substances, including polysaccharides (Di Martino 2018; Donlan 2002). To form biofilm, P. aeruginosa synthesizes three types of exopolysaccharides: pellicle (Pel), polysaccharide synthesis locus (Psl), and alginate. Overproduction of alginate promotes P. aeruginosa to form persistent infections in the CF lungs, while the rugose colonies overproducing Psl and Pel exopolysaccharides show an enhancement of P. aeruginosa persistence in both CF lungs and chronic wounds (Hentzer et al. 2001; Pestrak et al. 2018). There is evidence that Psl exopolysaccharides protect P. aeruginosa from host defenses during the initial phase of infection in the CF lungs. Therefore, the Psl‐derived exopolysaccharides in biofilm matrix may be important in the early stage of chronic lung infection before the bacteria change to produce an alginate‐based biofilm matrix (Mishra et al. 2012; Jones and Wozniak 2017).
As mentioned, Psl is one of the exopolysaccharides that plays an important role in P. aeruginosa biofilm formation. These saccharides consist of repeating pentasaccharide units of d‐mannose, d‐glucose, and l‐rhamnose. The psl gene cluster contains 15 genes (pslA–pslO), which encode proteins involved in polysaccharide biosynthesis (Byrd et al. 2009). The promoter of psl operon is located upstream of the pslA gene. Five psl genes (pslA, pslE, pslJ, pslK, and pslL) encoded Psl proteins (PslA, PslE, PslJ, PslK, and PslL, respectively) have inner membrane‐spanning domains and make up the Psl polymerization complex (Franklin et al. 2011). PslA plays a role in initiating Psl polysaccharide synthesis, which is crucial for the early stage of biofilm formation, attachment, and immune evasion. Comparative attachment analysis showed that P. aeruginosa PAO1ΔpslA is defective in attachment and that constitutive expression of the psl operon is required for efficient attachment to surfaces via synthesized exopolysaccharides (Overhage et al. 2005). PslE is involved in the export and assembly of the Psl polysaccharide, contributing to the biofilm's structural integrity, stability, and ability to withstand hostile environments (Franklin et al. 2011). The other three Psl proteins (PslJ, PslK, and PslL) are related to the transport, export, modification, and assembly of the Psl polysaccharide in P. aeruginosa. These genes work together to contribute to the biofilm formation and pathogenicity of P. aeruginosa to persist in chronic infections (Franklin et al. 2011).
Our previous study has demonstrated that S. aureus‐derived extracellular vesicles (SaEVs) promoted P. aeruginosa pathogenicity by increasing lipopolysaccharide (LPS) production, biofilm formation, and epithelial cell invasion, and by decreasing the uptake of P. aeruginosa by macrophages (Subsomwong et al. 2024). In addition, differential proteomic analysis revealed a significant increase in PslE production in P. aeruginosa after SaEV treatment (14‐fold increase compared to non‐SaEV treatment) (Subsomwong et al. 2024). However, the role of PslE and Psl exopolysaccharide polymerization on SaEV‐mediated P. aeruginosa pathogenicity is unclear. In this study, we aimed to investigate whether SaEVs influence P. aeruginosa pathogenicity in a PslE‐dependent manner. We constructed a pslE gene‐deletion mutant of P. aeruginosa (PaΔpslE). Using this mutant, the effects of SaEVs on the expression of five genes related to Psl polymerization, biofilm formation, and various aspects of P. aeruginosa pathogenicity were investigated in comparison to the P. aeruginosa wildtype (PaWT).
Materials and Methods
2
Bacterial Strains, Cell Cultures, and Growth Conditions
2.1
P. aeruginosa strain ATCC 15692, a standard reference strain of the PAO1 lineage, was used in this study. It was cultured in Tryptic soy agar (TSA) (BD Bioscience, Sparks, MD) at 37°C for 24 h. A single colony of P. aeruginosa was precultured in Tryptic soy broth (TSB) (BD Bioscience) under aerobic conditions (125 rpm) for 16 h. The P. aeruginosa preculture was inoculated into TSB and cultured at 37°C under aerobic conditions for 4 h. Then, the bacterial cells were collected and washed with 1× phosphate‐buffered saline (PBS) twice. The bacterial number was adjusted by converting optical density at 600 nm (OD_600nm_) measured with a UV‐1900i spectrophotometer (Shimadzu, Kyoto, Japan). The bacterial number at OD_600nm_ of 1.0 was estimated to 1.5 × 10^9^ CFU/mL.
HaCaT human keratinocytes and RAW 264.7 mouse macrophages were cultured at 37°C, 5% CO_2_ in Dulbecco's Eagle's minimum essential medium (DMEM; Nissui Pharmaceutical Co., Tokyo, Japan) supplemented with 10% fetal bovine serum (FBS; JRH Biosciences, Lenexa, KS), 0.075% NaHCO_3_ (Wako Pure Chemical Industries Ltd., Osaka, Japan), 0.03% l‐glutamine (Wako Pure Chemical), and 1× Antibiotic–Antimycotic (Gibco; Thermo Fisher Scientific, Waltham, MA).
Construction of pslE Gene Deletion in P. aeruginosa
2.2
Cre‐loxP system was used to construct a PaΔpslE. Details of the construction process are provided in Appendix 1 (Tables A1, A2 and Figures A1, A2, A3). The absence of pslE gene in the PaΔpslE genome was confirmed by PCR (Figures A4 and A5), DNA sequencing, and reverse transcription (RT)‐qPCR (Figure A6).
Preparation of SaEVs
2.3
SaEVs were prepared as described in our previous study (Subsomwong et al. 2024). Briefly, a standard laboratory strain of S. aureus (ATCC 1718) was cultured in Brain Heart Infusion broth for 8 h under aerobic conditions. The supernatant was collected and centrifuged to precipitate SaEVs at 100,000g, 4°C for 90 min using an ultracentrifuge (Himac CP80WX, HITACHI, Tokyo, Japan). After washing with ice‐cold PBS, the pellet was suspended and subjected to discontinuous iodixanol gradient ultracentrifugation (40%, 20%, 10%, and 5% iodixanol in 0.25 M sucrose/10 mM Tris, pH 7.5). After spinning at 100,000g, 4°C for 16 h using a swing out rotor, six fractions were collected from the top of the gradient. The substances in each fraction were then precipitated by ultracentrifugation at 100,000g for 2 h at 4°C. The resulting pellets were washed and resuspended in an appropriate volume of ice‐cold PBS. The protein concentration of each fraction was determined by Bradford protein assay (Bio‐Rad Laboratories, Richmond, CA).
Expression Analysis of Psl‐Related Genes in P. aeruginosa by RT‐qPCR
2.4
PaWT and PaΔpslE were precultured and the bacterial number was adjusted to OD_600nm_ = 1.0 (estimated to 1.5 × 10^9^ CFU/mL) in TSB. The bacterial cells were incubated at 37°C for 4 h with or without 5 µg/mL SaEVs under static conditions. The bacterial cells were harvested by centrifugation at 5800g, 4°C for 10 min, and washed twice with ice‐cold PBS.
Total RNA was extracted by TRIzol reagent (Thermo Fisher Scientific) as described in the manufacturer's instructions. To degrade the remaining gDNA, DNase I (5 µL of 1 U/µL) (Takara Bio Inc., Shiga, Japan) was added, and the RNA was extracted by TRIzol reagent again. The concentration of RNA was measured using NanoDrop Lite Plus spectrophotometer (Thermo Fisher Scientific). cDNA was synthesized from total RNA, random primer, and M‐MLV reverse transcriptase (Invitrogen). The expression of five genes; pslA, pslE, pslJ, pslK, and pslL, which are associated with polysaccharide polymerization in P. aeruginosa psl operon was determined by RT‐qPCR using SYBR Green Supermix (Bio‐Rad). The expression of rpoD gene, encoding RNA polymerase sigma factor, was used for normalization. The primer sequences and RT‐qPCR conditions used in this experiment are mentioned in our previous study (Subsomwong et al. 2024). The cycle threshold (C_T_) values obtained by the Bio‐Rad CFX96 RT‐PCR system were used to calculate the gene expression level. The C_T_ value of the rpoD gene was used for normalization with the formula [2^−(CT of target gene − CT of rpoD gene)^].
LPS Isolation and Detection
2.5
PaWT and PaΔpslE (OD_600nm_ = 1.0) were incubated in TSB with or without 5 µg/mL SaEVs at 37°C for 24 h under static conditions. The bacterial cells were then harvested by centrifugation at 5800g, 4°C for 10 min, washed with ice‐cold PBS twice, and stored at −80°C until use.
LPS was extracted from the bacterial cells by a hot aqueous‐phenol method as described previously (Jackson et al. 2004) with some modifications. Briefly, the bacterial cells were resuspended in 200 µL of SDS‐lysis buffer (4% β‐mercaptoethanol, 4% SDS, and 20% glycerol in 0.1 M Tris‐HCl, pH 6.8) and boiled for 15 min. DNase I and RNase solution (5 µL of 10 mg/mL each; Roche Diagnostic, Mannheim, Germany) were added and the sample was incubated for 30 min at 37°C to eliminate the DNA and RNA, respectively. Thereafter, proteinase K (10 µL of 10 mg/mL; Roche Diagnostic) was added and the sample was incubated at 59°C overnight. Ice‐cold Tris‐saturated phenol (200 µL; Invitrogen) was then added, the sample was incubated at 65°C for 15 min, cooled down to room temperature, and vortexed for 5–10 s with diethyl ether (1 mL; Nacalai Tesque Inc., Kyoto, Japan). Blue solution in the bottom layer was collected after centrifugation at 20,600g for 10 min. LPS in the sample was re‐extracted with the ice‐cold Tris‐saturated phenol step until the solution in the bottom layer was clear. The purified LPS was observed by SDS‐PAGE and stained using a Silver Stain 2 kit (Wako Pure Chemical) according to manufacturer's instructions. The quantitative intensity of each group was measured using ImageJ 1.54 g software, and the intensity of PaWT without SaEVs was set as 1.00.
Biofilm Formation and Quantification
2.6
PaWT and PaΔpslE were adjusted to OD_600 nm_ = 1.0 in TSB with or without 5 μg/mL SaEVs and were transferred to 96‐well plates (100 μL/well). After incubation at 37°C for 16 h, the planktonic cells in the culture supernatant were then removed. The biofilm was fixed by adding absolute methanol and incubating at room temperature for 15 min. The methanol was removed, the plates were left to dry at room temperature. The biofilm was stained with crystal violet solution [1% (w/v) in distilled water] for 5 min, washed with distilled water twice, and dried. Crystal violet bound to biofilm was dissolved in 96% ethanol and quantified by measuring the OD at 595 nm using a microplate reader (MULTISCAN Sky, Thermo Fisher Scientific). The relative biofilm biomass was calculated based on the biomass of PaWT without SaEVs as 100%.
Invasion of P. aeruginosa Into Human Keratinocytes
2.7
HaCaT cells (5 × 10^6^ cells/well) were seeded in 24‐well plates and incubated at 37°C, 5% CO_2_ for 48 h. Meanwhile, PaWT and PaΔpslE were prepared and adjusted to OD_600 nm_ = 1.0 in TSB. After incubation with or without 5 µg/mL SaEVs at 37°C for 24 h, the bacterial cells were collected, washed with PBS twice, and resuspended in antibiotic‐free DMEM. The HaCaT cells were washed with washing medium (FBS‐ and antibiotic‐free DMEM) and infected with PaWT or PaΔpslE at MOI = 100. After incubation at 37°C, 5% CO_2_ for 90 min, extracellular bacterial cells in the culture supernatant were removed. The cells were then washed twice with washing medium, once with PBS, and incubated in washing medium containing 120 μg/mL of Gm (Wako Pure Chemical) for 1 h. The cells were washed twice with washing medium and once with PBS, and then lysed with 1% 3‐[(3‐cholamidopropyl)‐dimethylammonio]−1‐propanesulfonate (DOJINDO, Kumamoto, Japan) in PBS for 15 min. Intracellular bacterial number was evaluated by plate count assay after incubating the TSA plates at 37°C for 24 h.
P. aeruginosa Uptake by Macrophages
2.8
RAW 264.7 cells (1 × 10^6^ cells/well) were seeded into 24‐well plates and incubated at 37°C, 5% CO_2_ for 48 h. Meanwhile, PaWT and PaΔpslE were adjusted, incubated with or without 5 µg/mL SaEVs for 24 h, and collected as described above. The RAW 264.7 cells were washed with washing medium and infected with PaWT or PaΔpslE at MOI = 10. After incubation at 37°C, 5% CO_2_ for 90 min, extracellular bacterial cells in culture supernatant were eliminated, and intracellular bacterial cells were evaluated by plate count assay.
Statistical Analysis
2.9
Statistical analyses were performed using GraphPad Prism (version 10.0; GraphPad Software Inc., La Jolla, CA). Data were presented as mean ± standard deviation, mode, min, and max. One‐way ANOVA with Tukey's multiple comparison test was used for relative gene expression analysis. Kruskal–Wallis with Dunn's multiple comparison test was applied for biofilm formation, bacterial uptake by macrophages, and bacterial invasion to keratinocytes experiments. The data are considered statistically significant when a p value is < 0.05. Significant levels indicating statistical significance were denoted as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
Results
3
SaEVs Enhance the Expression of Polysaccharide Polymerization‐Related Genes in the P. aeruginosa psl Cluster via pslE Gene
3.1
PaΔpslE was constructed, and successful deletion of the pslE gene was confirmed (Figure A6). By using this mutant and PaWT, we investigated whether SaEVs promote P. aeruginosa pathogenicity via PslE and Psl polysaccharide polymerization. The effect of SaEVs on the expression of pslE and other Psl polysaccharide polymerization‐related genes in psl cluster was evaluated. As shown in Figure 1, SaEVs significantly upregulated the expression of all five polysaccharide polymerization‐related genes in the psl cluster of PaWT. Based on the pslE gene deletion in the PaΔpslE genome, pslE gene expression in this PaΔpslE mutant was undetectable. In addition, the ability of SaEVs to promote the expression of pslA, pslJ, pslK, and pslL genes in the PaΔpslE was abolished (Figure 1). These results suggest that SaEVs enhance the expression of polysaccharide polymerization‐related genes, and the presence of pslE gene is required for SaEVs to promote pslA, pslJ, pslK, and pslL gene expression.
*Effect of SaEVs on the expression of polysaccharide polymerization‐related genes in psl cluster of pslE gene‐deletion mutant of P. aeruginosa (PaΔpslE) and P. aeruginosa wildtype (PaWT). PaΔpslE and PaWT were treated with 0 and 5 µg/mL SaEVs for 4 h. Relative mRNA expression of pslA, pslE, pslJ, pslK, and pslL was assessed by RT‐qPCR. The values are from two independent experiments (n = 6). Relative gene expression was calculated by referring to the expression of each target gene in PaWT as 1.0. Statistical analysis was performed using one‐way ANOVA with Tukey's multiple comparison test for relative gene expression. Asterisks indicate p values (*p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001). UD: undetectable.
SaEVs Enhance LPS Production in P. aeruginosa Partially in a pslE‐Dependent Manner
3.2
Since the function of psl gene cluster is involved in biofilm formation (Jones and Wozniak 2017), and LPS has been implicated in the architecture of P. aeruginosa biofilm (Rocchetta et al. 1999), we speculated that SaEVs may affect LPS production via pslE upregulation. Therefore, we further examined the ability of SaEVs to promote LPS production in PaΔpslE and PaWT. As shown in Figure 2, SaEVs‐treated PaWT had a higher intensity of O‐antigen (1.35‐fold) and lipid A and LPS core (2.20‐fold) compared to untreated PaWT control. LPS profile in PaΔpslE was identical to that in PaWT, although deletion of pslE gene resulted in a reduced intensity of O‐antigen (0.23‐fold) and lipid A and LPS core (0.82‐fold). In the PaΔpslE, the activity of SaEVs to promote lipid A and LPS core was abolished (0.79‐fold). However, SaEVs still promoted O‐antigen intensity (0.38‐fold) in the PaΔpslE (Figure 2). These results suggest that SaEVs may promote the LPS production in P. aeruginosa partially via pslE gene. In particular, SaEVs are involved in lipid A and LPS core but not O‐antigen biosynthesis in the pslE‐dependent manner.
Effect of SaEVs on LPS production in pslE gene deletion mutant of P. aeruginosa (PaΔpslE) and P. aeruginosa wildtype (PaWT). PaΔpslE and PaWT were treated with or without 5 µg/mL SaEVs for 24 h. LPS were isolated, separated by SDS‐PAGE, and stained using Silver Stain kit. All samples were started with the same number of bacterial cells, prepared by the same process, and applied in equal volumes. The table demonstrates the quantitative intensity of each group, setting the intensity of PaWT without SaEVs as 1.00.
pslE Gene is Required for SaEVs to Promote Biofilm Formation in P. aeruginosa
3.3
We further investigated the effect of SaEVs on the biofilm formation in PaΔpslE and PaWT. As expected, SaEVs significantly enhanced the biofilm biomass of PaWT (122.0%) compared to untreated PaWT (Figure 3). In addition, the biofilm biomass of PaΔpslE was significantly reduced by ~50% compared to that of PaWT. There was no significant difference in biofilm biomass between SaEV‐treated and untreated PaΔpslE (Figure 3). These results suggest that pslE is one of the important factors for biofilm formation in P. aeruginosa and SaEVs promote biofilm formation through the pslE gene. In addition, the biofilm biomass in PaΔpslE may be formed by other mechanisms that are not influenced by SaEVs.
*Effect of SaEVs on biofilm formation in pslE gene deletion mutant of P. aeruginosa (PaΔpslE) and P. aeruginosa wildtype (PaWT). PaΔpslE and PaWT were cultured with or without 5 µg/mL SaEVs for 16 h. Biofilm biomass was quantified by crystal violet assay. Relative biofilm biomass was calculated by referring PaWT without SaEVs as 100%. The values are from two independent experiments (n = 6). Statistical analysis was performed using Kruskal–Wallis with Dunn's multiple comparisons. Asterisks indicate p values (*p < 0.05, *p < 0.01), NS: not significant.
SaEVs and pslE Gene Are Not Involved in P. aeruginosa Invasion Into Epithelial Cells
3.4
We have previously demonstrated that SaEVs promote P. aeruginosa invasion into human keratinocyte HaCaT cells (Subsomwong et al. 2024). However, we were unable to replicate this result in the current experiment. As shown in Figure 4, intracellular bacterial numbers of SaEV‐treated and untreated PaWT were comparable. Although the intracellular bacterial number of PaΔpslE was higher than that of PaWT, this difference was not statistically significant in our data. In addition, pretreatment of PaΔpslE with SaEVs did not affect the intracellular bacterial number of PaΔpslE in the HaCaT cells (Figure 4). This result suggests that SaEVs and pslE gene do not contribute to P. aeruginosa invasion into the epithelial cells.
Effect of SaEVs on invasion of pslE gene deletion mutant of P. aeruginosa (PaΔpslE) and P. aeruginosa wildtype (PaWT) into human keratinocytes. PaWT and PaΔpslE were treated with or without 5 µg/mL SaEVs for 24 h before infection. HaCaT cells were infected with PaΔpslE and PaWT at MOI = 100 for 90 min. The extracellular bacterial cells were eliminated by gentamicin treatment for 1 h. Then, the intracellular bacterial number was evaluated by plate count assay. Relative intracellular bacterial number was calculated by referring PaWT without SaEVs as 100%. The values are from two independent experiments (n = 6). Statistical analysis was performed using Kruskal–Wallis with Dunn's multiple comparisons. NS: not significant.
SaEVs Reduce Uptake of P. aeruginosa by Macrophages in a pslE‐Dependent Manner
3.5
Biofilm formation is a key factor in P. aeruginosa resistance to host immune defenses (Thöming and Häussler 2022; Moser et al. 2017). Therefore, we further investigated whether pslE upregulation in P. aeruginosa by SaEVs contributes to evasion of macrophage uptake. Our results demonstrated that the uptake of PaWT by macrophages significantly reduced by SaEV‐treatment (Figure 5). Lack of pslE gene in PaΔpslE resulted in a slight increase of PaΔpslE uptake by macrophages but this increase did not reach statistical significance. In addition, SaEVs did not alter intracellular bacterial number of PaΔpslE in macrophages. These results suggest that SaEVs reduce uptake of P. aeruginosa by macrophages in a pslE‐dependent manner.
*Effect of SaEVs on uptake of pslE gene deletion mutant of P. aeruginosa (PaΔpslE) and P. aeruginosa wildtype (PaWT) by mouse macrophages. PaΔpslE and PaWT were treated with or without 5 µg/mL SaEVs for 24 h before infection. RAW 264.7 cells were infected with PaΔpslE and PaWT at MOI = 10 for 90 min. The extracellular bacterial cells were eliminated by gentamicin treatment for 1 h. Then, the intracellular bacterial number was evaluated by plate count assay. Relative intracellular bacterial number was calculated by referring PaWT without SaEVs as 100%. The values are from two independent experiments (n = 6). Statistical analysis was performed using Kruskal–Wallis with Dunn's multiple comparisons. Asterisks indicate p values (*p < 0.05, *p < 0.01), NS: not significant.
Discussion and Conclusions
4
Coinfections with P. aeruginosa and S. aureus are commonly found in patients with chronic conditions, including CF and chronic wounds (Bernardy et al. 2022; DeLeon et al. 2014; Fugère et al. 2014). P. aeruginosa and S. aureus can form mixed biofilms that protect them from the host immune defenses and significantly reduce the efficiency of antibiotic treatment, resulting in recalcitrant, difficult‐to‐treat chronic infections (Vestweber et al. 2024), leading to increased morbidity and mortality worldwide (Bernardy et al. 2022; Phan et al. 2023; Hubert et al. 2013; Limoli et al. 2016). The interaction between P. aeruginosa and S. aureus has been reported. However, their interaction mediated by EVs and the underlying mechanisms are not well understood. Our previous study has reported that EVs secreted by S. aureus interact P. aeruginosa, promote biofilm formation and contribute to P. aeruginosa pathogenicity (Subsomwong et al. 2024). To understand more detailed mechanisms, differential proteomic analysis between SaEV‐treated and untreated P. aeruginosa was performed. Our data revealed that 22 proteins were upregulated (Subsomwong et al. 2024). Among these proteins, a 74.56 kDa protein which is identical to PslE in P. aeruginosa PAO1 shows the highest fold change with 14.85‐fold (Subsomwong et al. 2024). Since PslE is one of the polysaccharide polymerization proteins in psl cluster (Franklin et al. 2011), we speculated that SaEVs may promote Psl exopolysaccharide production via PslE production, resulting in an enhancement of P. aeruginosa pathogenicity.
The results in this study demonstrated that SaEVs significantly upregulated the expression of pslE gene as well as the other polysaccharide polymerization‐related genes (pslA, pslJ, pslK, and pslL) in psl operon of PaWT. Interestingly, the effect of SaEVs in promoting pslA, pslJ, pslK, and pslL gene expression was abolished in PaΔpslE. This result suggests that SaEVs may not directly regulate the expression pslA, pslJ, pslK, and pslL gene, but rather these genes may be upregulated by SaEVs through pslE gene or PslE protein. Although this mechanism is required to be elucidated, the expression of all five polysaccharide polymerization‐related genes in psl operon promoted by SaEVs may act together to enhance exopolysaccharide polymerization in the PaWT.
The phosphomannomutase enzyme (encoded by the algC gene), which is crucial for alginate exopolysaccharide biosynthesis, was shown to be involved in LPS biosynthesis (Ye et al. 1994). This enzyme helps to convert mannose‐6‐phosphate to mannose‐1‐phosphate, a precursor for both alginate and LPS. Jackson et al. (2004) have demonstrated that an algC mutant of P. aeruginosa PAO1 clearly lacks core oligosaccharides. Although psl cluster is not directly involved in the LPS biosynthesis, we determined if the psl‐dependent effect of SaEVs is LPS related. Our results showed that the intensity of LPS in PaΔpslE was lower than that in the PaWT. However, the LPS profile in both O‐antigen and LPS core in PaΔpslE was identical to that in PaWT. The activity of SaEVs to promote lipid A and LPS core was abolished in PaΔpslE, suggesting that SaEVs may be involved in lipid A and LPS core biosynthesis via pslE gene. However, SaEVs still promoted O‐antigen production in PaΔpslE. These results suggested that the activity of SaEVs in promoting O‐antigen production may be involved in the other mechanism(s) rather than psl‐dependent manner.
Exopolysaccharides are important for the formation and stability of biofilms and the biofilm benefits P. aeruginosa by promoting its survival in stressful environments and allowing its persistence on the surface (Ciofu and Tolker‐Nielsen 2019; Thi et al. 2020). The role of Psl in mucoid biofilm has been reported. The ability of P. aeruginosa in forming biofilms decreased when the Psl‐encoding genes are absent, indicating that Psl is a crucial component for mucoid biofilm formation (Ma et al. 2012). Disruption of the initial gene of the psl cluster (pslA either alone or with pslB) in P. aeruginosa PAO1 affects biofilm formation under both static and continuous flow culture conditions (Overhage et al. 2005; Jackson et al. 2004). Likewise, the mutation of PA2231 (pslA) in the PAO1 strain resulted in the formation of thin, unstructured, abnormal biofilms that differ from those formed by the wildtype strain (Jackson et al. 2004; Matsukawa and Greenberg 2004). Our result showed that PaΔpslE exhibited about 50% reduction in biofilm biomass compared to the PaWT. This suggests that not only pslA and pslB, but also pslE plays a role in biofilm synthesis. In addition, SaEVs upregulate pslE gene expression, resulting in an enhancement of biofilm formation.
Evasion of phagocytosis by host immune cells is one of the pathogenic mechanisms used by P. aeruginosa to persist in the host. Phagocytosis by innate immune cells is a process by which the host cells eradicate the microbes (Hastings et al. 2023; Marzhoseyni et al. 2023). Exopolysaccharides on the surface of P. aeruginosa are not only important for biofilm formation but also have been known to play a significant role in reducing immune cell recognition and uptake (Leid et al. 2005). It has been reported that Psl‐deficient P. aeruginosa were internalized and killed by neutrophils and macrophages more efficiently than PaWT and Psl overexpressing strains (Mishra et al. 2012). Likewise, our results demonstrated that SaEVs reduce P. aeruginosa uptake by macrophages in a pslE‐dependent manner. Regarding epithelial cell invasion, our data suggest that neither SaEVs nor pslE gene contribute to P. aeruginosa invasion into the keratinocytes. We hypothesized that the bacteria produce proteins related to cell invasion, host immune response, and colonization during the acute phase, while proteins associated with biofilm formation, antibiotic resistance, and immune evasion are produce during chronic phase. Fleming et al. (2022) showed that neither the overexpression nor the absence of Pel and Psl polysaccharides significantly alter the ability of P. aeruginosa to establish an infection and survive in a mouse model of wound infection. These data suggest that the production of Psl exopolysaccharides does not affect the pathogenesis of P. aeruginosa in keratinocytes and wound infections.
Overall, our results demonstrated that SaEVs promote gene expression in psl cluster, biofilm formation, and antiphagocytosis of P. aeruginosa in a *pslE‐*dependent manner. However, the detailed mechanism by which SaEVs regulate the pathogenicity of P. aeruginosa via the psl operon remains unknown. We hypothesized that SaEVs may activate the psl promoter and/or directly upregulate the pslE gene to promote the expression of other psl genes and exopolysaccharide production. In addition, the molecules in SaEVs involved in the regulation of psl operon need to be considered.
In conclusion, the present study demonstrated that EVs derived from S. aureus mediate the pathogenicity of P. aeruginosa via a pslE‐dependent mechanism, resulting in increased biofilm formation and antiphagocytosis by macrophages. These findings are important to support further development of therapeutic approaches against S. aureus and P. aeruginosa coinfection, such as inhibiting SaEV interaction with P. aeruginosa or targeting psl polymerization to inhibit biofilm formation. In addition, the knowledge of EV‐mediated interactions and pathogenicity will lead to the development of therapeutics against other coinfections with opportunistic bacteria, especially those capable of forming biofilms.
Author Contributions
Phawinee Subsomwong: conceptualization (supporting), methodology (lead), data curation (lead), investigation (lead), validation (lead), formal analysis (lead), funding acquisition (lead), writing – original draft (lead), writing – review and editing (supporting). Rojana Sukchawalit: methodology (supporting), resources (supporting), validation (supporting), writing – review and editing (supporting). Naoko Watabe: software (supporting), writing – review and editing (supporting). Akio Nakane: funding acquisition (supporting), writing – review and editing (supporting). Krisana Asano: conceptualization (lead), supervision (lead), project administration (lead), funding acquisition (supporting), writing – original draft (supporting), writing – review and editing (lead).
Ethics Statement
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bernardy, E. E. , V. Raghuram , and J. B. Goldberg . 2022. “ Staphylococcus aureus and Pseudomonas aeruginosa Isolates From the Same Cystic Fibrosis Respiratory Sample Coexist in Coculture.” Microbiology Spectrum 10, no. 4: e 0097622. 10.1128/spectrum.00976-22.35867391 PMC 9431432 · doi ↗ · pubmed ↗
- 2Briaud, P. , S. Bastien , L. Camus , et al. 2020. “Impact of Coexistence Phenotype Between Staphylococcus aureus and Pseudomonas aeruginosa Isolates on Clinical Outcomes Among Cystic Fibrosis Patients.” Frontiers in Cellular and Infection Microbiology 10: 266. 10.3389/fcimb.2020.00266.32582568 PMC 7285626 · doi ↗ · pubmed ↗
- 3Briaud, P. , L. Camus , S. Bastien , A. Doléans‐Jordheim , F. Vandenesch , and K. Moreau . 2019. “Coexistence With Pseudomonas aeruginosa Alters Staphylococcus aureus Transcriptome, Antibiotic Resistance and Internalization Into Epithelial Cells.” Scientific Reports 9, no. 1: 16564. 10.1038/s 41598-019-52975-z.31719577 PMC 6851120 · doi ↗ · pubmed ↗
- 4Byrd, M. S. , I. Sadovskaya , E. Vinogradov , et al. 2009. “Genetic and Biochemical Analyses of the Pseudomonas aeruginosa Psl Exopolysaccharide Reveal Overlapping Roles for Polysaccharide Synthesis Enzymes in Psl and LPS Production.” Molecular Microbiology 73, no. 4: 622–638. 10.1111/j.1365-2958.2009.06795.x.19659934 PMC 4409829 · doi ↗ · pubmed ↗
- 5Cangui‐Panchi, S. P. , A. L. Ñacato‐Toapanta , L. J. Enríquez‐Martínez , J. Reyes , D. Garzon‐Chavez , and A. Machado . 2022. “Biofilm‐Forming Microorganisms Causing Hospital‐Acquired Infections From Intravenous Catheter: A Systematic Review.” Current Research in Microbial Sciences 3: 100175. 10.1016/j.crmicr.2022.100175.36518176 PMC 9743049 · doi ↗ · pubmed ↗
- 6Ciofu, O. , and T. Tolker‐Nielsen . 2019. “Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents‐How P. aeruginosa Can Escape Antibiotics.” Frontiers in Microbiology 10: 913. 10.3389/fmicb.2019.00913.31130925 PMC 6509751 · doi ↗ · pubmed ↗
- 7Costerton, J. W. , P. S. Stewart , and E. P. Greenberg . 1999. “Bacterial Biofilms: A Common Cause of Persistent Infections.” Science 284, no. 5418: 1318–1322. 10.1126/science.284.5418.1318.10334980 · doi ↗ · pubmed ↗
- 8Dan, B. , H. Dai , D. Zhou , H. Tong , and M. Zhu . 2023. “Relationship Between Drug Resistance Characteristics and Biofilm Formation in Klebsiella pneumoniae Strains.” Infection and Drug Resistance 16: 985–998. 10.2147/IDR.S 396609.36824066 PMC 9942501 · doi ↗ · pubmed ↗