Effect of gentiopicroside on endogenous formaldehyde homocysteine–pathway related proteins in rats with non-alcoholic steatohepatitis
Jiaxin Chen, Huiling Zuo, Yuhang Jiao, Huanhuan Zhao, Xuan Ma, Yahui Xiao, Xiangqiong Li, Wei Zhao, Anhua Shi, Wenhui Chen

TL;DR
Gentiopicroside (GPS) may help treat non-alcoholic steatohepatitis (NASH) by reducing liver fat and oxidative stress in rats.
Contribution
This study shows GPS improves NASH by modulating the endogenous formaldehyde-homocysteine pathway and reducing oxidative stress in rats.
Findings
GPS reduces liver steatosis and NAFLD activity scores in NASH rats.
GPS increases HDL-C and decreases harmful serum markers like ALT, AST, and LDL-C.
GPS modulates proteins like ALDH2 and MTHFR in the FA-HCY pathway, reducing oxidative stress.
Abstract
Non-alcoholic steatohepatitis (NASH) is a progressive form of non-alcoholic fatty liver disease (NAFLD) characterised by high prevalence, increasing incidence among younger individuals and poor prognosis. NASH pathophysiology is not completely understood, and at present, there are no viable pharmacological treatments for this condition in clinical practice. Gentiopicroside (GPS). Can alleviate NASH by reducing inflammatory responses, inhibiting oxidative stress and influencing blood lipid levels. To examine the preventive and therapeutic effects of gentiopicroside (GPS) on proteins and metabolites related to the endogenous formaldehyde-homocysteine (FA-HCY) pathway as well as on oxidative stress in NASH rats. A high-fat, high-sugar diet was used to create a NASH rat model, and the rats’ liver weight, body weight and liver index levels were measured. Oil red O and hematoxylin and eosin…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
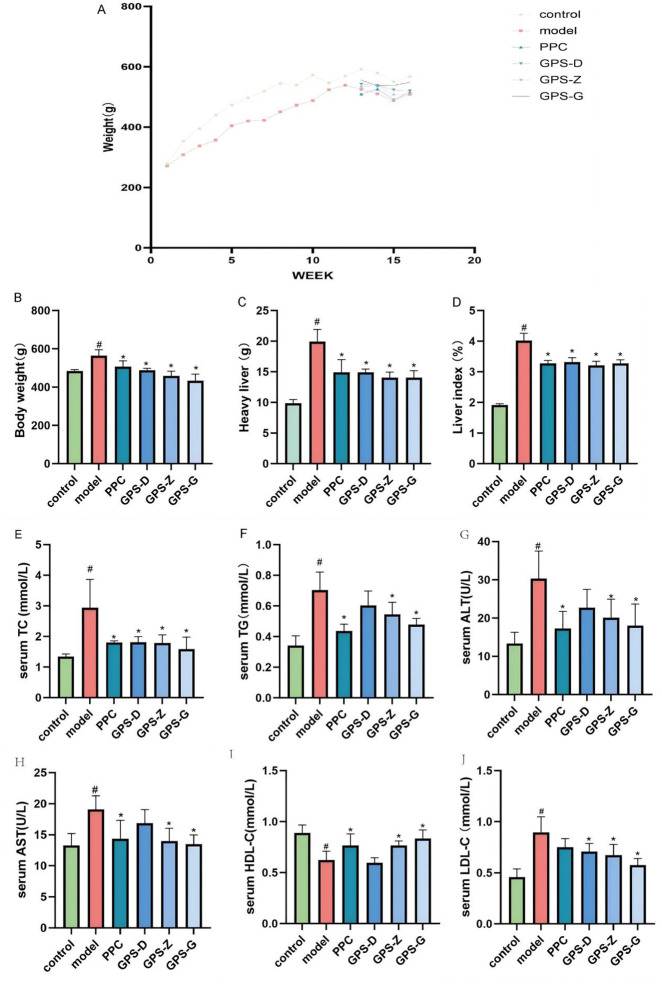 FIGURE 1
FIGURE 1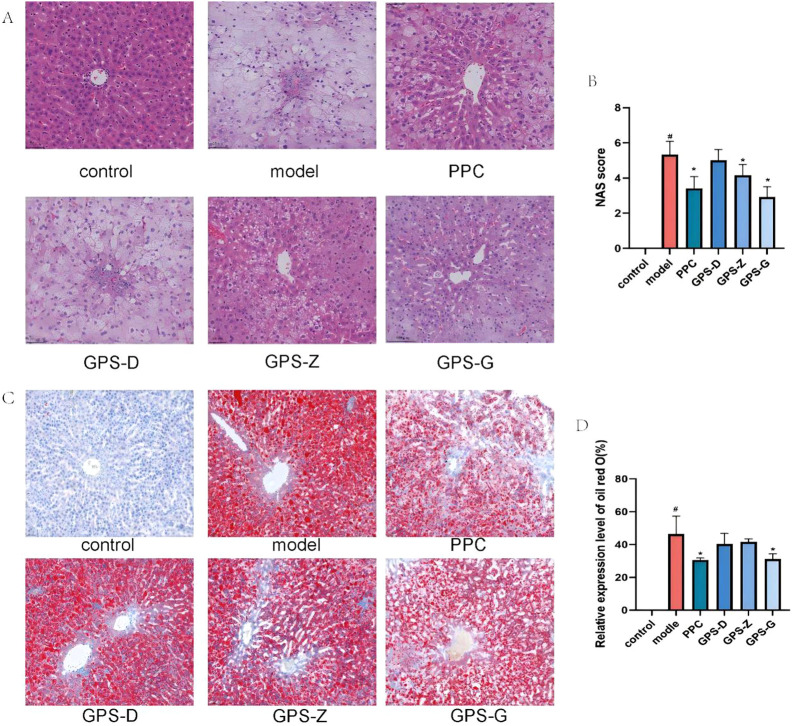 FIGURE 2
FIGURE 2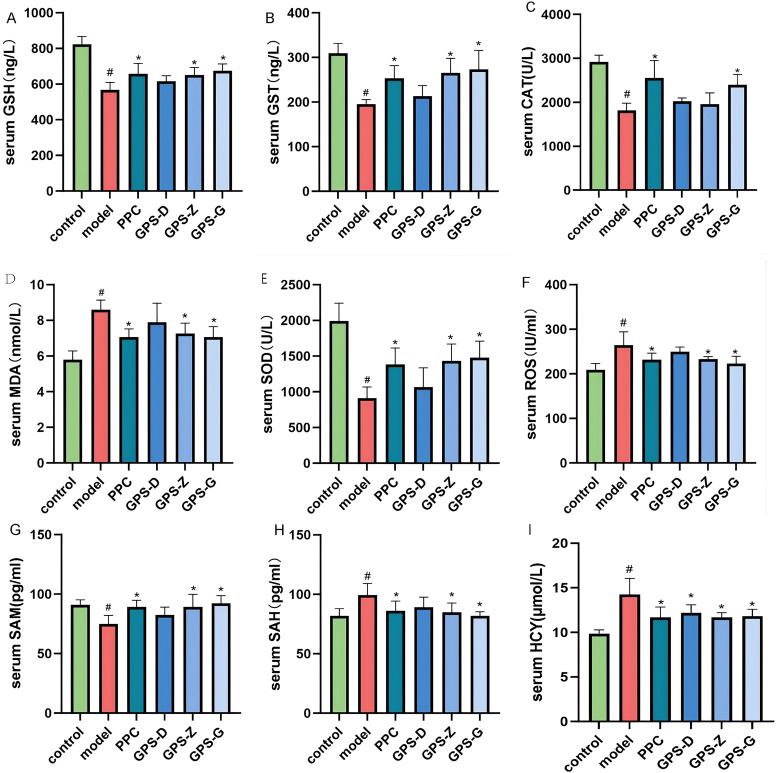 FIGURE 3
FIGURE 3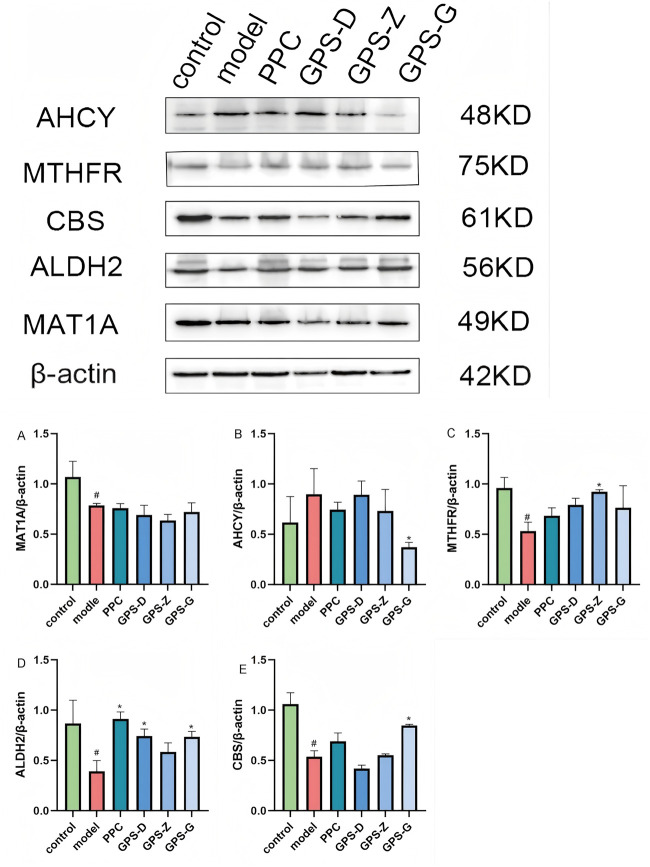 FIGURE 4
FIGURE 4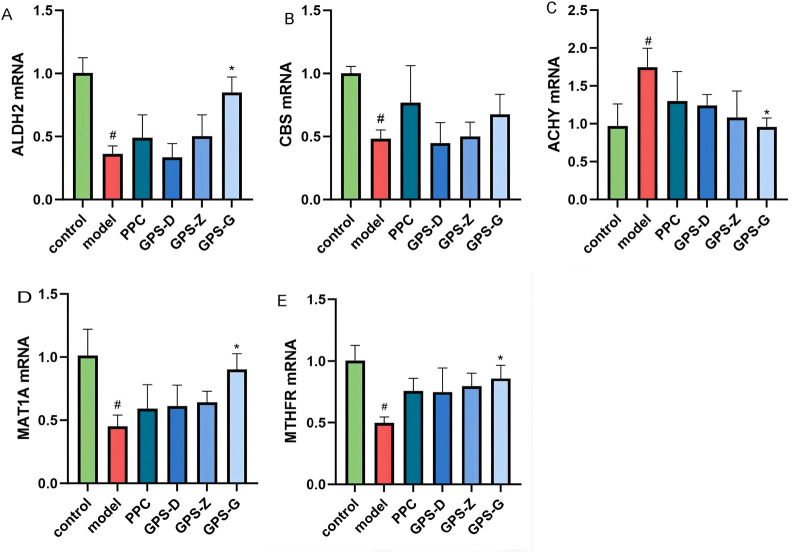 FIGURE 5
FIGURE 5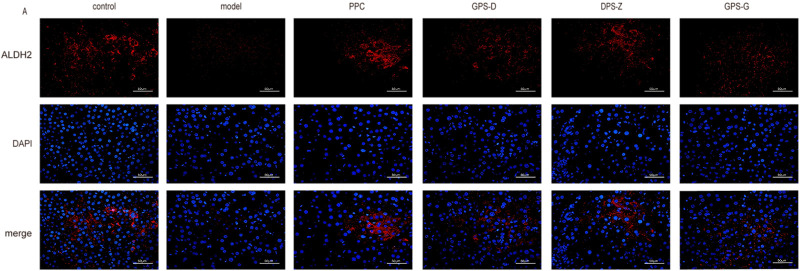 FIGURE 6
FIGURE 6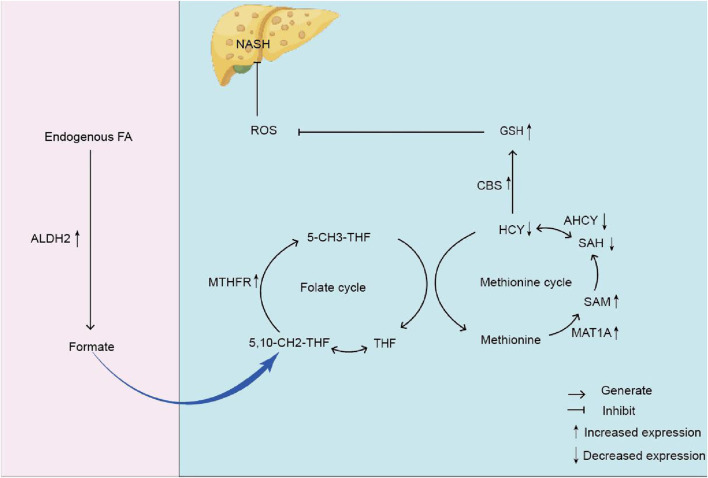 FIGURE 7
FIGURE 7| Project | 0 points | 1 points | 2 points | 3 points |
|---|---|---|---|---|
| Hepatocyte steatosis | <5% | 5%∼<33% | 33%∼<66% | >66% |
| Inflammation within the lobules | No | <2/200x field of view | 2–4/200x field of view | >4/200x field of view |
| Balloon-like transformation of hepatocytes | No | Occasionally seen | Common | |
| Primer name | Primer sequence (5′-3′) |
|---|---|
| ALDH2-F | GGTTGGTGGGCGGTGTTTG |
| ALDH2-R | GCGTGTGCGTGCGTGTG |
| MTHFR-F | GCGTATCAGCCTTGCCTTCAC |
| MTHFR-R | CTCTGGTCCTGCCTCAAC |
| CBS-F | GGGATGGTGACTGGGAAC |
| CBS-R | GGATCGGCTTGAACTGCTTGTAG |
| AHCY-F | ATATCATCCTTGGTCGGCACTTTG |
| AHCY-R | CCTTCTCCACAGCGTTGTCATTG |
| MAT1A-F | TGCCGCTCACCATTGTTCTTG |
| MAT1A-R | AATCAGGTCTCAGCCAGGGAAG |
| β-actin-F | ACTGCCGCATCCTCTTCCTC |
| β-actin-R | GAACCGCTCATTGCCGATAGTG |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLiver Disease Diagnosis and Treatment · Alcohol Consumption and Health Effects · Drug-Induced Hepatotoxicity and Protection
Introduction
1
Non-alcoholic steatohepatitis (NASH), development of non-alcoholic fatty liver disease (NAFLD), is characterised by liver steatosis ≥5%, hepatocyte injury, inflammation and various degrees of liver fibrosis (Chalasani et al., 2018). The global frequency of NASH in the general population is 2%–6% (Kadi et al., 2024). The occurrence and development of this condition involve various factors, including insulin resistance (Branković et al., 2022), disturbances in glucose and lipid metabolism (Wang et al., 2022), dietary imbalances and intestinal microbiota (Yang et al., 2023) and genetic polymorphisms (Lee et al., 2022). The pathogenesis of NASH is not completely understood, and there are currently no effective pharmacological treatments available in clinical practice (Wei et al., 2024). Therefore, identifying pharmacological agents to effectively treat NASH is vital.
Endogenous formaldehyde (FA) serves as a methyl donor and participates in numerous cellular biological processes in vivo, such as one-carbon metabolism, nucleotide synthesis, DNA methylation, epigenetic modification, gene expression regulation and metabolic maintenance (Burgos-Barragan et al., 2017; Ridpath et al., 2007; Yu et al., 2015). Alcohol dehydrogenase 5 (ADH5) (Morellato et al., 2021) or aldehyde dehydrogenase 2 (ALDH2) (Kim et al., 2020; Nattermann et al., 2023) can convert endogenous FA into formate (Umansky et al., 2022).
It can directly engage in one-carbon metabolism and spontaneously condense with tetrahydrofolate (THF) to yield 5,10-CH_2_-THF. This compound then participates in one-carbon metabolism (Morellato et al., 2021), which encompasses the folic acid and methionine cycles. Homocysteine (HCY) is generated from methionine by removing its terminal methyl group and serves as an intermediary in the methionine cycle (Besen et al., 2024). Dysregulation of methionine metabolism can lead to fat buildup and is intimately associated with the pathological alterations in hepatic steatosis (Murthy et al., 2003; Radziejewska et al., 2020). HCY can undergo trans-sulfuration to yield cysteine, which is further metabolised by Cystathionine -β -synthase (CBS) to generate hydrogen sulphide (H_2_S) and GSH (Dahlhoff et al., 2013). The accumulation of HCY, prevention of its remethylation and trans-sulfuration and inadequate production of GSH to mitigate the effects of reactive oxygen species (ROS) contribute to promoting oxidative stress (Mato et al., 2017; Raghubeer and Matsha, 2021).
Moreover, HCY can directly produce ROS through auto-oxidation or lead to ROS accumulation by disturbing the equilibrium between pro-oxidative and anti-oxidant activities (Harb et al., 2020). HCY build up can enhance lipid peroxidation reactions (Fang et al., 2023), trigger oxidative stress, activate inflammatory and fibrotic signalling pathways (Gonzalez et al., 2020), lead to inflammatory infiltration of hepatocytes (Gulsen et al., 2005) and facilitate the progression of steatosis to NASH (Liu et al., 2015).
In conclusion, endogenous FA build up and the mismatch between supply and demand of one-carbon units result in abnormalities in one-carbon metabolism that cause HCY accumulation, further encouraging lipid build up and oxidative stress and leading to NASH. Consequently, dysfunction of the endogenous FA-HCY pathway can facilitate the onset and progression of NASH.
Gentiopicroside (GPS) is a cyclic ether terpene glycoside extracted from plants of genus Gentiana and family Gentianaceae. This molecule exhibits several pharmacological properties, including cholesterol-lowering, anti-inflammatory, anti-oxidant and hepatoprotective properties and plays a crucial role in managing NAFLD by reducing lipid build up (Choubey et al., 2024; Dai et al., 2018; He et al., 2015; Liu et al., 2024; Xiao et al., 2022).
This study replicated the rat model of NASH to observe the preventive and therapeutic effects of GPS on NASH rats, and it preliminarily explored the relationship between the therapeutic effect of GPS and the metabolism-related indicators of the endogenous formaldehyde-homocysteine pathway, as well as oxidative stress.
Materials and methods
2
Experimental materials
2.1
Experimental animals
2.1.1
In total, 50 male Sprague–Dawley rats, weighing 160–180 g, of SPF grade and aged 6 weeks, were acquired from the Laboratory Animal Center of Kunming Medical University (licence no.: SCXK (Dian) 2020–0004). The experimental animals were housed in individual cages (six animals per cage) at the Animal Experiment Center of Yunnan University of Traditional Chinese Medicine. Temperature was maintained at 22 °C–27 °C, with relative humidity of 40%–60%. The rats had free access to food and water, and periods of 12 h of light and 12 h of darkness were alternated to replicate the typical circadian rhythm. The experimental protocols regarding diet, anaesthesia, blood and tissue sample collection and disposal of dead animals complied with the ethical guidelines issued by the Experimental Animal Ethics Committee of Yunnan University of Traditional Chinese Medicine (No. R-062023223).
Experimental reagents
2.1.2
GPS with a purity ≥98% (Baoji Chenguang Biotechnology Co., LTD., HR2475W19); polyene phosphatidylcholine (Sanofi [Beijing] Pharmaceutical Co., LTD., National Drug Approval No. EBJD4198); mouse Afodin solution (G0-R047, Beijing Hanhai Tuoxin Biotechnology Co., LTD.). Total cholesterol (TC), triglycerides (TG), alanine aminotransferase (ALT), aspartate aminotransferase (AST), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C) kits were purchased from the Nanjing Jiancheng Institute of Bioengineering with item numbers A111-1-1, A110-1-1, C009-2-1, C010-2-1, A112-1-1 and A113-1-1, respectively. ROS, GSH, MDA, SOD, SAH, SAM, GST, CAT, GSH, HCY were obtained from Jiangsu Enzyme Immunoassay Biotechnology Co., LTD. Their item numbers are MM-88564O1, MM-20251R1, MM-2037H1, MM-20387R1, MM-50456H1, MM-0248H1, MM-21254R1, MM-20447R1, MM-20251R1 and MM-50456H1. β-actin antibody (Proteintech, 20536-1-AP); ALDH2 antibody (Proteintech, 15310-1-AP); MAT1A antibody (Proteintech, 12395-1-AP); MTHFR antibody (Proteintech, 26591-1-AP); CBS antibody (Proteintech, 14787-1-AP); AHCY antibody (Proteintech, 10757-2-AP); PVDF membrane with a thickness of 0.45 μm (IPVH00010, Millipore); Universal total RNA extraction reagent (U7431, Suzhou Youyi Landi Biotechnology Co., LTD.) Taq SYBR® Green qPCR Premix (Universal) (EG0117M, Jiangsu Bishi Mei Biotechnology Co., LTD.) All-in-One First-Strand Synthesis MasterMix (with dsDNase) was obtained from Jiangsu Bestmate Biotechnology Co., LTD. (item number (EG15133S).
Experimental methods
2.2
Experimental animals
2.2.1
The 50 SPF-grade Sprague–Dawley rats selected for this experiment were acclimatised for 1 week and subsequently randomly assigned to control (n = 8) and modeling (n = 42) groups. The modeling group received a high-fat diet supplemented with 5% brown sugar water for 12 weeks. After the model verification is successful, the Modelin group will be randomly assigned to the model group (n = 8, from the original group), one polyene phosphatidylcholine (PPC) treatment (23.3 mg/kg, n = 8) and three GPS treatment groups at low, medium and high doses. GPS-D (50 mg/kg, n = 8),GPS-Z (100 mg/kg, n = 8),GPS-G (200 mg/kg, n = 8), respectively. Each drug was administered for 4 weeks. The rats were weighed on a weekly basis, and liver and serum samples were obtained at the end of the experiment.
Measurement of indicators
2.3
Serum biochemical indicators
2.3.1
The levels of TC,TG, LDL-C,HDL-C, ASTand ALT in the serum of rats in each group were measured using biochemical kits. and calculated liver index. The liver index is as follows: liver mass/body mass × 100%.
Hematoxylin and eosin (H&E) staining method
2.3.2
Liver tissue samples were preserved in 4% paraformaldehyde solution, embedded in paraffin, sectioned into 5-μm thick slices and stained with H&E reagent per standard protocols. Then, histological alterations were examined using slide scanning image analysis (SQS1000-S, Shengqiang Technology Co., LTD.). Steatosis, lobular inflammation and hepatocyte ballooning were assessed to determine the severity of NAFLD using the NAFLD activity score. The specific evaluation criteria are included in Table 1. NAS <3 excludes NASH, 3–4 may indicate NASH and ≥4 confirms the presence of NASH.
Following comprehensive theoretical instruction, two students from our research group performed a blind evaluation of liver tissue sections. Six pathological specimens were selected from each group, for 36 slice samples. The intensities of hepatic inflammation, necrosis and fibrosis were assessed to determine the NAFLD activity score. The mean scores of the two students were computed, and the data were analysed in SPSS v. 27.0.
Liver oil red O staining
2.3.3
The liver lobules preserved in 4% paraformaldehyde will be pruned and dehydrated. After embedding them with OCT adhesive, the samples were frozen, sectioned, dyed with oil red O and sealed with glycerol gelatin. After drying, they were photographed using a glass slide scanner. In this technique, lipid droplets are coloured red, whereas cell nuclei are dark blue. The oil red O-stained area was quantitatively assessed using ImageJ. Oil Red O expression = (area value of lipid droplets in each group/mean total area) × 100%. The acquired data were analysed in SPSS 27.0 and the results were displayed as ( ).
Enzyme-linked immunosorbent assays (ELISA)
2.3.4
ELISA kits were used to measure the levels of HCY, SAM, SAH, ALDH2, CBS, MDA, SOD, ROS, GSH, GST and CAT expression in rat serum. All procedures were carefully executed by following kit instructions, and preliminary studies were conducted for each assay. Optical density was measured using a multi-functional microplate reader (Synergy LX, BIOTEK, Berten Instruments, USA), and the standard curve equation was calculated based on the concentration of the standard substance. The index concentration was computed, followed by data collection, and analysed using SPSS 27.0. The results were visualised as ( ).
Gas chromatography-mass spectrometry
2.3.5
Gas chromatography-mass spectrometry was used to determine the amounts of endogenous FA in the rats’ livers and serum. Samples were processed by adding 1% formic acid water and subjecting them to ultrasonic extraction for 30 min. Then, 1 mL of the sample solution was added with 100 μL of 2, 4-dinitrophenylhydrazine solution (10 mg/10 mL 1% formic acid) and placed in a water bath at 60 °C for derivatisation for 30 min. After rapid cooling, 1 mL of n-hexane solution was added, and the mixture was shaken for extraction. Then, the upper layer was removed and loaded into the chromatograph for analysis. The FA standard products were derived simultaneously, with standard concentrations of 10, 20, 50, 100 and 200 μg/L. The following conditions were maintained for operating the chromatograph (Agilent 7890B): chromatographic column, DB-5; detector, ECD; injection port, 250 °C; detector, 300 °C; split ratio, 10:1; gradient, maintain at 80 °C for 1 min, increase from 20 °C/min to 200 °C, then from 30 °C/min to 280 °C and maintain for 2 min.
Western blot analysis
2.3.6
Liver tissue samples, stored in a freezer at −80 °C, were homogenised using radioimmunoassay assay (RIPA) buffer (Beyotime Biotechnology, Shanghai, China). Each sample was separated using a 10% SDS-PAGE gel and subsequently transferred to a PVDF membrane. The membrane was blocked with 5% skim milk for 2 h and subsequently incubated overnight at 4 °C with the ALDH2, MAT1A, MTHFR, CBS and AHCY antibodies. Another incubation step with a suitable HRP-conjugated secondary antibody was then carried out at room temperature for 1 h. ImageJ was used to analyse protein levels, which were normalised based on β-actin levels. The data were analysed in SPSS 27.0, and the results were visualised as ( ).
Real-time (RT) PCR
2.3.7
Total RNA was isolated from the frozen rat liver tissues using RNA extraction kits, and RNA purity was subsequently assessed using a micro-spectrophotometer. Next, cDNA was synthesised from 1 μg of high-quality total RNA using a reverse transcription premix kit. RT PCR amplification was conducted at the following conditions: pre-denaturation at 95 °C for 30 s, denaturation at 95 °C for 5 s, annealing at 55 °C for 30 s and extension at 72 °C for 30 s. Amplification was conducted for 40 cycles, using β-actin as the internal reference gene, with the 2^−ΔΔCT^ value representing the relative level of mRNA expression. The data were analysed in SPSS 27.0, and the results were visualised as ( ). The primer sequences are included in Table 2.
Immunofluorescence analysis
2.3.8
Paraffin sections were prepared from the rat liver tissue samples. A dewaxing solution was used for the dewaxing process, and EDTA was employed for repair purposes. After the samples cooled naturally, they were blocked with 3% BSA for 30 min and then supplemented with the primary antibody overnight. The next day, they were washed with PBS three times and incubated with the secondary antibody at room temperature for 50 min. Images were captured via self-fluorescence quenching and then analysed using ImageJ. Mean fluorescence intensity was calculated as the sum of fluorescence intensities within the specified region (IntDen) divided by the area. The data were analysed in SPSS v. 27.0, and the results were visualised as ( ).
Statistical analysis
2.4
All data were analysed in SPSS v. 27.0, and the results were presented as the mean ± standard deviation. Before analysis, tests were conducted to assess the normality of data distribution for each group and the homogeneity of variances. For homogeneous variances, one-way ANOVA was employed for group comparisons, followed by the LSD test, whereas for heterogeneous variances Tamhane’s T2 all-pairs comparison test was conducted instead. Statistically significant differences were defined by a P-value of <0.05. Multiple groups of non-normally distributed data were analysed using the non-parametric Kruskal–Wallis test for independent samples, with statistical significance also set at P-value of <0.05. Graphs depicting statistical results were plotted using GraphPad Prism v. 10.1.2.
Results
3
Effects of GPS on serum lipid metabolites and liver function in NASH rats
3.1
The liver index, defined as the ratio of liver weight to body weight in rats, serves as a notable predictor of hepatic fat build up. The GPS treatment groups exhibited a significant decrease in body and liver weight, and thus in the liver index (P < 0.05). Notably, GPS can mitigate NASH in rats. Compared with the control group, the model group had elevated levels of serum TC, TG, AST, ALT and LDL-C (P < 0.05). Compared with the model group, the PPC treatment group exhibited elevated levels of TC, TG, ALT, AST and HDL-C (P < 0.05) but low levels of LDL-C, although this difference was not statistically significant. All GPS treatment groups (three dosages) exhibited significantly reduced levels of TC, TG, ALT, AST and LDL-C (P < 0.05) but increased levels of HDL-C (P < 0.05) (Figure 1). Therefore, GPS boosts lipid deposition in the liver in NASH rats, improving its function (Figure 2).
GPS enhances body weight, blood lipid metabolites, and liver function parameters in NASH rats. (A) Time-course body weight curves for each group of rats; (B) Body weight levels of rats in each group (n = 6); (C) Liver weight levels of rats in each group (n = 6); (D) Liver index levels of rats in each group (n = 6); (E) Serum TC levels of rats in each group (n = 6); (F) Serum TG levels of rats in each group (n = 6); (G) Serum ALT levels of rats in each group (n = 6); (H) Serum AST levels of rats in each group (n = 6); (I) Serum HDL-C levels of rats in each group (n = 6); (J) Serum LDL-C levels of rats in each group (n = 6). Note: Data are expressed as the mean ± SD. “#” indicates that P < 0.05 compared with the normal group; “” indicates that P < 0.05 compared with the model group.*
GPS mitigates lipid accumulation and the infiltration of inflammatory factors in the livers of NASH rats. (A) H&E staining results of livers in each group of rats (20 × 10); (B) NAS score levels of rats in each group (n = 6); (C) Results of oil red O staining in each group of rats (scale = 500 μm); (D) Expression levels of oil red O area in each group of rats (n = 6). Note: Data are expressed as the mean ± SD. “#” indicates that P < 0.05 compared with the normal group; “” indicates that P < 0.05 compared with the model group.*
Influence of GPS on liver pathological indicators
3.2
The H&E staining results indicated a distinct liver architecture in the control group rats, with hepatocytes arranged in an orderly and compact manner. In contrast, in the model group, the organisation of hepatocytes was aberrant: cells were palely stained and lipid vacuoles along with inflammatory infiltration were present in the cytoplasm. Compared with the model group, the hepatocytes in the GPS-D, GPS-Z, GPS-G and PPC treatment groups exhibited an orderly arrangement, with fewer lipid vacuoles and inflammatory infiltration. The NAFLD activity scores of rats in the model group were significantly higher than those of rats in the control (P < 0.05) and GPS-Z, GPS-G and PPC treatment groups (P < 0.05).
The result of oil red O staining indicated a distinct liver architecture in the control group, with minimal formation of red lipid droplets. A considerable accumulation of red lipid droplets was observed in the livers of rats in the model group. A notable amelioration of hepatic steatosis was detected in the GPS-D, GPS-Z, GPS-G and PPC treatment groups. The relative expression level of oil red O was significantly higher in the model group than in the control (P < 0.05) and GPS-G and PPC treatment groups (P < 0.05). For the GPS-Z and GPS-D groups, the relative expression levels of oil red O were also lower than those in the model group; however, the differences were not statistically significant (Figure 2).
Effects of GPS on serum oxidative stress levels and metabolites related to the endogenous FA-HCY pathway in NASH rats
3.3
The model group exhibited higher levels of MDA and ROS (P < 0.05) but lower levels of SOD, GSH, GST and CAT (P < 0.05) compared with the control and GPS treatment groups. Considerably, in NASH rats, GPS may lower the degree of oxidative stress in the serum (Figure 3).
In NASH rats, GPS lowers serum oxidative stress levels. (A) Serum GSH levels of rats in each group (n = 6); (B) Serum GST levels of rats in each group (n = 6); (C) Serum CAT levels of rats in each group (n = 6); (D) Serum MAD levels of rats in each group (n = 6); (E) Serum SDO levels of rats in each group (n = 6); (F) Serum ROS levels of rats in each group (n = 6); (G) Serum SAM levels of rats in each group (n = 6); (H) Serum SAH levels of rats in each group (n = 6); (I) Serum HCY levels of rats in each group (n = 6). Note: Data are expressed as the mean ± SD. “#” indicates that P < 0.05 compared with the normal group; “” indicates that P < 0.05 compared with the model group.*
The model group showed higher levels of SAH and HCY (P < 0.05) but lower levels of SAM, ALDH2 and CBS (P < 0.05) compared with the control and GPS-G treatment group (Figure 3; Supplementary Figure S1).
Therefore, GPS may influence markers of metabolites associated with the endogenous FA-HCY pathway in rats with NASH induced by a high-fat and high-sugar diet.
Gas chromatography results indicated that, compared with the control group, the model group rats exhibited a higher level of endogenous FA expression in the liver (P < 0.05) but a lower level in the serum, although this difference was not statistically significant. After GPS treatment, the levels of endogenous FA in the serum increased; however, the difference was not statistically significant (Supplementary Figure S1).
Influence of GPS on the expression levels of endogenous FA-HCY pathway-related proteins and mRNA in the livers of NASH rats
3.4
Western blotting was used to detect the expression of proteins related to the endogenous FA-HCY pathway in NASH rats. Notably, the levels of MAT1A, MTHFR, ALDH2 and CBS expression in the liver in the model group were significantly lower than those in the control group (P < 0.05), whereas the level of AHCY expression was higher, although this difference was not statistically significant. Following GPS treatment, the level of AHCY expression decreased (P < 0.05). MAT1A levels also decreased, although this change was not statistically significant. Conversely, the levels of ALDH2 and CBS expression increased significantly after GPS treatment (P < 0.05). Additionally, an increase was observed for MTHFR expression, but it was not statistically significant (Figure 4).
Expression levels of CBS, ALDH2, AHCY, MAT1A and MTHFR proteins in liver tissues of rats in each group. (A) Expression levels of MAT1A protein in liver tissues of rats in each group (n = 3); (B) Expression levels of AHCY protein in liver tissues of rats in each group (n = 3); (C) Expression levels of MYHFR protein in liver tissues of rats in each group (n = 3); (D) Expression levels of ALDH2 protein in liver tissues of rats in each group (n = 3); (E) Expression levels of CBS protein in liver tissues of rats in each group (n = 3). Note: Data are expressed as the mean ± SD. “#” indicates that P < 0.05 compared with the normal group; “” indicates that P < 0.05 compared with the model group.*
The results of RT PCR revealed higher levels of AHCY and related mRNA expression in the liver in the model group than in the control group (P < 0.05), but decreased expression of MTHFR, MAT1A, CBS and ALDH2 and related mRNA (P < 0.05). The GPS treatment groups exhibited a significant reduction in AHCY and related mRNA expression (P < 0.05), along with significant increases in MTHFR, MAT1A and ALDH2 mRNA expressions (P < 0.05). CBS and related mRNA expression also increased, although not significantly (Figure 5).
Expression levels of CBS, ALDH2, AHCY, MAT1A and MTHFRmRNA in liver tissues of rats in each group (A) Expression levels of ALDH2 mRNA in liver tissues of rats in each group (n = 3); (B) Expression levels of MTHFRmRNA in liver tissues of rats in each group (n = 3); (C) Expression levels of CBS mRNA in liver tissues of rats in each group (n = 3); (D) Expression levels of AHCY mRNA in liver tissues of rats in each group (n = 3); (E) Expression levels of MAT1A mRNA in liver tissues of rats in each group (n = 3). Note: “#” indicates that P < 0.05 compared with the normal group; “” indicates that P < 0.05 compared with the model group.*
Effect of GPS on the relative fluorescence intensity of endogenous FA-HCY pathway-related proteins
3.5
In the fluorescence microscopy images, the proteins ALDH2, MAT1A and MTHFR exhibited red fluorescence, whereas cell nuclei displayed blue fluorescence. The observed fluorescence intensity facilitated the localisation and semi-quantitative analysis of the proteins under investigation. The results showed significantly lower (P < 0.05) levels of MTHFR, MAT1A and ALDH2 expression in the liver in the model group than in the control group. Conversely, the expression of these proteins was significantly increased in the GPS treatment groups (P < 0.05) (Figure 6; Supplementary Figure S3).
Fluorescence expression intensity of ALDH2 in liver tissues of rats in each group. Scale = 50 μm.
Discussion
4
Lipid metabolism disorder is a pathological mechanism of NASH (Ji et al., 2023). Abnormal lipid metabolism results in altered serum levels of TG, TC, HDL-C, and LDL-C (Hegazy et al., 2020). HDL-C and LDL-C serve as primary risk factors for lipid deposition, with a notable positive correlation observed between LDL and liver inflammation (Higashi, 2023). ALT and AST levels correlate with liver function and serve as biomarkers for liver cell inflammatory injury in clinical settings (Chinnappan et al., 2023). Furthermore, ALT is significantly associated with the extent of liver fat accumulation, helping predict fibrosis levels in NASH (Xuan et al., 2024; Zeng et al., 2022).
The experimental results indicated that, in comparison to the control group, the model group rats exhibited increased levels of TC, TG, AST, ALT, and LDL-C, while HDL-C levels decreased. In comparison to the model group rats, the GPS treatment group exhibited decreased levels of TC, TG, AST, ALT, and LDL-C, while HDL-C levels increased. Therefore, GPS enhances liver function and ameliorate lipid metabolism disorders in NASH rats, thereby reducing lipid deposition, similar to the existing literature. For instance (Kou et al., 2023), demonstrated that GPS administration significantly reduced liver function indicators (ALT and AST) in H_2_O_2_-induced HepG2 cells, and two other studies on NASH mice reported that GPS lowered the levels of TC, TG, ALT and AST (Yong et al., 2024; Huang et al., 2024) the levels of TC, TG and LDL-C (Huang et al., 2024) in the serum.
Steatosis and hepatocyte ballooning are the hallmarks of steatohepatitis, a condition characterised by enlarged hepatocytes with a thin or wispy cytoplasm. H&E-stained images allow the quantification and spatial localisation of hepatic steatosis (Mairinoja et al., 2023), which visually appears as a ‘spiderweb’ (Lai et al., 2022). Herein, the H&E-stained images of rat liver tissue samples from each GPS treatment group showed well-aligned hepatocytes, diminished fat vacuoles and inflammatory infiltrates and lower NAFLD activity scores relative to the images obtained for the model group. The integration of H&E and oil red O facilitated a more accurate quantification of hepatic steatosis (expressed as a percentage), enhancing the visibility of lipid droplets (Riva et al., 2018). Oil red O staining indicated reduced area of red lipid droplets in the liver along with decreased relative expression of oil red O in the GPS-treated groups. The hepatic index indicates hepatic lipid deposition (Koehler et al., 2013). Our experimental results demonstrated that, compared with the model group, body weight, liver weight and the hepatic index were significantly reduced across all GPS treatment groups. Thus, GPS effectively reduces liver steatosis and lipid deposition induced by a high-fat and high-sugar diet in NASH rats, while attenuating the degree of pathological changes in the liver.
Polyenephosphatidylcholine (PPC) is a commonly used hepatoprotective agent in clinical practice (Xu et al., 2022), demonstrating favorable therapeutic effects for various liver diseases such as drug-induced liver injury (Li et al., 2022), and non-alcoholic fatty liver disease (Lu et al., 2022; Yang, et al., 2023). PPC is the primary active component of essential phospholipids, playing a crucial role in maintaining cellular membrane fluidity and preserving membrane integrity. It exhibits high bioavailability and multiple biological functions, including inhibiting abnormal lipid accumulation, anti-inflammatory effects (Chen X. et al., 2024) antioxidant properties (Lei et al., 2024), and immunomodulation (Yin et al., 2023). It promotes hepatocyte regeneration and maintains hepatocyte membrane stability, playing a significant role in improving NASH (Lei et al., 2024). In 2000, PPC had already garnered attention for its protective effects against liver damage (Lu et al., 2022; Yang, et al., 2023). PPC can improve and repair damaged hepatocyte membranes. Supplementation with PPC reduces liver weight and improves insulin resistance in mice fed a high-fat diet (HFD). Studies have demonstrated that PPC treatment significantly improves hepatic steatosis (Y. Li et al., 2022) and liver function markers (ALT, AST) (Shao et al., 2025). The objective of this experiment is to validate the effects of GPS on proteins associated with the endogenous formaldehyde-homocysteine pathway in NASH rats. Currently, no targeted drugs exist for this pathway; therefore, PPC, which has demonstrated efficacy against NASH, was selected as the positive control drug.
Oxidative stress directly damages lipids, proteins and DNA, while activating inflammatory and fibrotic signalling pathways (Gonzalez et al., 2020), and it is a crucial factor in the progression of steatosis to NASH. This process is primarily characterised by ROS overproduction, direct depletion of anti-oxidant molecules such as GSH and inhibition of superoxide dismutase (SOD) activity (Liu et al., 2015). GSH is predominantly found in the liver and serves as the primary protector against oxidative stress. ROS can be converted into hydrogen peroxide (H_2_O_2_) through SOD, which is subsequently reduced to H_2_O by GSH, thus safeguarding cells from the detrimental effects of free radicals and pro-oxidants (Averill-Bates, 2023; Gao et al., 2022; Vairetti et al., 2021). ROS can interact with biomolecules, including proteins, leading to lipid peroxidation and the production of malondialdehyde (MDA) (Martín-Fernández et al., 2022). MDA, the end product of lipid peroxidation, significantly contributes to the progression of steatosis to NASH (Cordiano et al., 2023). SOD catalyses the conversion of superoxide anion (•O_2_ ^−^) into O_2_ and H_2_O_2_, which are subsequently degraded to H_2_O by CAT for metabolic processes (Huang et al., 2021). Glutathione S-transferases (GSTs) are categorised as phase II detoxification enzymes. GSH can bind with various endogenous and exogenous electrophilic compounds (Prysyazhnyuk et al., 2021). GSTs can employ GSH to transform lipid peroxides and other peroxides into less harmful compounds for detoxification (Csiszár et al., 2016). Reportedly, compared with the model group, the GPS treatment groups exhibited decreased levels of MDA and ROS, along with increased levels of SOD, GSH, GST and CAT. Thus, GPS ameliorates serum oxidative stress levels in NASH rats.
The body metabolism of endogenous FA is mostly regulated by the liver. The body can eliminate endogenous FA by metabolising it in the liver and releasing it into the serum. The experimental results of this study indicated that the level of endogenous FA in the liver was higher in the model group than in the control and all three GPS treatment groups. In contrast, endogenous FA content in the serum was lower in the model group than in the control group; however, this difference was not statistically significant. The endogenous FA concentration in the serum in the GPS-G treatment group was higher than that in the model group. We hypothesize that the observed phenomenon may result from increased endogenous formaldehyde levels in the serum due to hepatic metabolism following medication delivery. Currently, there are no direct methods available for the detection of endogenous formaldehyde. The quantities of various metabolites of endogenous formaldehyde can be ascertained by detection. This effort initially intended to utilize high-performance liquid chromatography for the detection of endogenous formaldehyde, given that formaldehyde can be metabolized via many pathways and that metabolite measurements may contain inaccuracies. Nevertheless, in later practice, the concentration of formaldehyde was insufficient for detection. Therefore, gas chromatography-mass spectrometry was selected for the identification of endogenous formaldehyde. Formaldehyde’s tendency to volatilize outside the body may result in losses during transportation and sample processing. Furthermore, the samples may exhibit issues such as significant variability and limited sample sizes attributable to individual differences among rats. Consequently, the statistical differences lack sufficient significance. The effect of a high-fat diet on endogenous formaldehyde concentrations in the body is now ambiguous, necessitating additional experimental validation.
Various methionine metabolites, including SAM, SAH and HCY, are crucial factors influencing lipid levels in the liver. HCY is produced at a crucial juncture in one-carbon metabolism, and blood HCY levels are positively associated with NASH (Matthews et al., 1998; Tripathi et al., 2022). SAH and HCY levels in the serum are significantly correlated with the extent of hepatic steatosis (Tang et al., 2022). Increased SAH levels in particular are a defining feature of NASH lesions (Pogribny et al., 2017). SAM is a principal methyl donor in numerous transmethylation processes and a multi-functional molecule integral to the regulation of hepatic lipid metabolism (Fernández-Ramos et al., 2022). The decrease of SAM levels is a significant catalyst for the onset of human NAFLD and subsequent progression to NASH (Zubiete-Franco et al., 2016). Supplementation with SAM or increasing cysteine intake can enhance GSH synthesis, restore ROS levels, boost TG metabolism in the liver, avoid endoplasmic reticulum stress, minimise de novo fat creation in hepatic cells and mitigate liver fat build up (Chen G. et al., 2024; Chen et al., 2023; Van der Crabben et al., 2008). Herein, the GPS treatment groups exhibited higher SAM levels than the model group, but lower levels of SAH and HCY (Bacil et al., 2023). revealed that the livers of mice with NASH induced by a diet rich in saturated fats and sugars exhibited reduced SAH levels and increased SAM levels. Additionally, Tang et al. (2022) indicated that the levels of SAM, SAH and HCY in the serum were remarkably increased in patients with NAFLD.
Given that these disparities may stem from varying diets, the concentrations of methionine metabolites among patients on a methionine-choline deficient, high-fat, and/or standard diet could be discrepant. CBS, the rate-limiting enzyme in the trans-sulfuration pathway, is crucial for detoxifying harmful HCY (Robert et al., 2005). The overexpression of CBS reportedly decreases the plasma HCY levels in mice (Wang et al., 2004). CBS deficiency can lead to hepatic steatosis, inflammation and fibrosis in animal models; nevertheless, the correlation between CBS and NASH remains undetermined (Robert et al., 2005). This study demonstrated that GPS treatment elevated CBS protein and mRNA expression levels in the liver of rats in the mod group; however, the difference in protein expression was not statistically significant. The absence of statistical significance in our observations can be ascribed to the regulation of CBS metabolism by SAM. Specifically, SAM can non-covalently interact with the heme groups of CBS and modulate its redox sensitivity. Thus, reduced SAM concentrations can diminish CBS activity (Werge et al., 2021). Nevertheless, limited studies have been conducted on the potential role of CBS and underlying mechanisms in NASH. Additional tests are required to ascertain whether the transcriptional level of CBS is influenced by other variables. ALDH2, a mitochondrial aldehyde dehydrogenase, is mostly expressed in the liver and scavenges aldehydes in vivo, thereby safeguarding cells against oxidative stress damage (Bae et al., 2017; Li et al., 2013; Schug, 2018). By conducting cellular tests with ALDH2 deletion, Bae (Bae et al., 2017) revealed that the absence of aldehyde dehydrogenases may increase the levels of ROS and hazardous aldehydes, further promoting apoptosis (Li et al., 2023). found that NASH worsened in ALDH2 knockout mice, with the lack of this enzyme possibly exacerbating inflammation, facilitating the degradation of the gut flora and inhibiting the FXR pathway. ALDH2 activators such as Alda-1 may mitigate liver damage in mice by improving aldehyde elimination and reversing liver steatosis and apoptosis.
MTHFR is essential for single-carbon metabolism, involving the methionine and folic acid cycles, synthesis of proteins, DNA and RNA, and remethylation of HCY, and is vital for cellular homeostasis. This protein is responsible for regulating the equilibrium of methionine and HCY, averting cellular malfunction and is essential for the methylation of HCY to methionine (Cai et al., 2024; Liew and Gupta, 2015; Raghubeer and Matsha, 2021). Reduced MTHFR and SAM levels and increased SAH levels in the liver in addition to mild micro-alveolar hepatic steatosis reportedly influence the inflammatory and lipid pathways in a murine model (Raghubeer and Matsha, 2021). Our experimental results showed lower levels of MTHFR and mRNA expression in the liver in the model group than in the control and GPS treatment groups. Previous research indicated a potential association between allelic variants of MTHFR (A1298C and C677T) and susceptibility to NAFLD (Hao et al., 2021). Nonetheless, the role of MTHFR polymorphism in the manifestation of NAFLD remains undetermined research across several groups is yet to clarify the influence of MTHFR polymorphism on NAFLD development (An et al., 2017). Liew et al. (Liew and Gupta, 2015) suggest that the MTHFR C677T polymorphism may not be linked to the development of non-alcoholic steatosis into NASH. Consequently, more experiments are required to elucidate these dynamics.
The MAT protein comprises two subtypes, MAT1A and MAT2A, which are predominantly expressed in the adult liver, regulate SAM homeostasis and play a crucial role in lipid metabolism (Sáenz de Urturi et al., 2022). MAT1A can be activated to produce elevated SAM concentrations, establishing a positive feedback loop (Murray et al., 2019). The lack of MAT1A may result in spontaneous steatosis, further progressing to NASH and fibrosis (Fernández-Ramos et al., 2022). Reduced MAT1A expression and diminished SAM levels can induce genomic instability and exacerbate the dysregulation of one-carbon metabolism via many mechanisms during hepatic SAM depletion (Ramani and Lu, 2017). Reportedly, MAT1A^−/−^ mice can spontaneously develop steatosis, exhibiting reduced SOD activity and heightened lipid peroxidation, thereby accelerating the progression of NAFLD to NASH (Robinson et al., 2023). Moreover, mice lacking the MAT1A gene demonstrated reduced SAM/SAH levels and manifested spontaneous NASH at 9–10 months of age (Alarcón-Vila et al., 2023). The experimental results of this study showed lower levels of MAT1A and mRNA expression in the liver in the model group than in the control GPS treatment groups. In contrast, Quinn C et al. (Quinn et al., 2022) reported a substantial upregulation of MAT1A in NASH mice. This discrepancy may be attributed to the body’s compensating mechanisms, along with inherent individual variances among rats. S-adenosylmethionine (AdoMet) can modulate the expression of MAT1A and MAT2A (GarcÍa-Trevijano et al., 2000). The production of AdoMet from various sources may result in fluctuations in the levels of MAT1A. Nevertheless, there is a paucity of studies regarding the potential role of MAT1A and its regulation in NASH. For example, additional tests are required to ascertain whether the transcriptional level of this protein is influenced by other factors.
AHCY is a distinctive enzyme and one of the most conserved proteins across animal groups, crucial for regulating SAM and SAH levels (Krushkal et al., 2016). In mammals, AHCY is the sole enzyme capable of facilitating this process (Vizán et al., 2021). A reduction in AHCY activity will increase SAH levels, hence inhibiting several methylation processes (Matthews et al., 2009). The methylation of AHCY may influence enzyme activity, resulting in abnormalities in HCY metabolism (Li et al., 2020). The suppression of AHCY expression during NASH progression may disrupt trans-sulfuration and transmethylation processes, exacerbating NASH and creating a detrimental feedback loop (Zhang et al., 2020).
The experiment’s results demonstrated that, in comparison to the control group rats, the model group rats had elevated expression levels of AHCY protein and mRNA in the liver. In comparison to the model group of rats, the expression levels of AHCY protein and mRNA in the GPS therapy group were diminished. Quinn C (Quinn et al., 2022). discovered that AHCY expression was diminished in the livers of NASH mice. Our findings align with those of prior research. However, it must be said that the fundamental mechanism associated with AHCY and NASH is still ambiguous and necessitates additional experimental validation (Figure 7).
The mechanism diagram of endogenous formaldehyde participating in one-carbon metabolism leading to homocysteine accumulation and causing NASH. Note: formaldehyde (Endogenous FA); Glutathione (GSH) Mitochondrial aldehyde dehydrogenase 2 (ALDH2); Formate; Tetrahydrofolate (THF); 5, 10-methylenetetrahydrofolate (5, 10-CH2-THF); Methylenetetrahydrofolate reductase (MTHFR); Homocysteine (HCY); S-adenosine homocysteine (SAH); Reactive oxygen species (ROS); Non-alcoholic steatohepatitis (NASH); Methionine Cystathionine -β -synthase (CBS); 5-methyltetrahydrofolate (5-CH3-THF); S-adenosylmethionine (SAM); Methionine adenosyltransferase 1A (MAT1A); S-adenosine homocysteine hydrolase (AHCY).
The translation of mRNA is a crucial step in the expression of protein-coding genes (Kusnadi et al., 2022). This study reveals that mRNA expression levels of target genes exhibit consistent trends with their corresponding protein expression levels. This finding reflects the synergistic regulatory mechanisms at the transcription-translation interface within this pathway. Despite fluctuations in gene mRNA levels, complex coordination exists between temporally and spatially distinct phases of transcription and translation, maintaining stable levels of specific protein subpopulations through multiple mechanisms (Famà et al., 2025). Furthermore, the effects of high-fat diets on the genes encoding these metabolic enzymes remain poorly understood. Disruption of these enzymes would impair one-carbon metabolism, leading to homocysteine accumulation and promoting the development of NASH. The impact of high-fat diets on endogenous formaldehyde levels remains unclear. Consequently, experimental studies may exhibit elevated methionine metabolism enzymes due to the inclusion of normal dietary methionine content.
In conclusion, GPS ameliorates liver steatosis generated by a high-fat and high-sugar diet in a rat model of NAFLD possibly by affecting the endogenous FA-HCY pathway. Consequently, our research offers a viable therapeutic approach for managing hepatic steatosis and related damage.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alarcón-Vila C. Insausti-Urkia N. Torres S. Segalés-Rovira P. de la Rosa L. C. Nuñez S. (2023). Dietary and genetic disruption of hepatic methionine metabolism induce acid sphingomyelinase to promote steatohepatitis. Redox Biol. 59, 102596. 10.1016/j.redox.2022.102596 36610223 PMC 9827379 · doi ↗ · pubmed ↗
- 2An X. Yang Z. An Z. (2017). Mi R-149 compromises the reactions of liver cells to fatty acid via its polymorphism and increases non-alcoholic fatty liver disease (NAFLD) risk by targeting methylene tetrahydrofolate reductase (MTHFR). Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 23, 2299–2307. 10.12659/msm.901377 28507283 PMC 5443364 · doi ↗ · pubmed ↗
- 3Averill-Bates D. A. (2023). The antioxidant glutathione. Vitamins Hormones (Vol. 121, pp. 109–141. 10.1016/bs.vh.2022.09.002 36707132 · doi ↗ · pubmed ↗
- 4Bacil G. P. Romualdo G. R. Piagge P. M. Cardoso D. R. Vinken M. Cogliati B. (2023). Unraveling hepatic metabolomic profiles and morphological outcomes in a hybrid model of NASH in different mouse strains. Antioxidants 12 (2), 290. 10.3390/antiox 12020290 36829849 PMC 9952348 · doi ↗ · pubmed ↗
- 5Bae S. Chon J. Field M. S. Stover P. J. (2017). Alcohol dehydrogenase 5 is a source of formate for de novo purine biosynthesis in Hep G 2 cells. J. Nutr. 147 (4), 499–505. 10.3945/jn.116.244467 28228507 PMC 5368588 · doi ↗ · pubmed ↗
- 6Besen S. Ozkale Y. Ceylaner S. Noyan A. Erol I. (2024). Clinical and laboratory findings and etiologies of genetic homocystinemia: a single-center experience. Acta Neurol. Belg. 124 (1), 213–222. 10.1007/s 13760-023-02356-1 37728847 · doi ↗ · pubmed ↗
- 7BrankovićM. JovanovićI. DukićM. RadonjićT. OprićS. Klašnja S. (2022). Lipotoxicity as the leading cause zof non-alcoholic steatohepatitis. Int. Journal Molecular Sciences 23 (9), 5146. 10.3390/ijms 23095146 35563534 PMC 9105530 · doi ↗ · pubmed ↗
- 8Burgos-Barragan G. Wit N. Meiser J. Dingler F. A. Pietzke M. Mulderrig L. (2017). Mammals divert endogenous genotoxic formaldehyde into one-carbon metabolism. Nature 548 (7669), 549–554. 10.1038/nature 23481 28813411 PMC 5714256 · doi ↗ · pubmed ↗