A revision of the genus Beesia (Ranunculaceae) as informed through integrative taxonomy, with description of a new species from Sichuan (China)
Andrey S. Erst, Elizaveta Yu. Mitrenina, Denis A. Krivenko, Tatyana V. Erst, Yulia V. Cheldysheva, Igor V. Gorbenko, Renata Borosova, Lian Lian, Yuan-Yuan Ling, Huan-Wen Peng, Jun Zhang, Shukherdorj Baasanmunkh, Hyeok Jae Choi, Ivan V. Tatanov, Alexander A. Kuznetsov

TL;DR
This study uses multiple data types to revise the taxonomy of the Beesia genus, discovering a new species from Sichuan, China.
Contribution
The study introduces a new cryptic species, Beesia yangii, using an integrative taxonomic approach in plant systematics.
Findings
Morphological, phylogenetic, and cytogenetic data identified three distinct Beesia species.
A new species, Beesia yangii, was described and is critically endangered.
Integrative taxonomy improved species delimitation and conservation insights.
Abstract
Taxonomic diversity at different levels of organization is determined by different levels of biological diversity, including morphological, phylogenetic (genetic or/and genomic), and cytogenetic differences. However, focused studies utilizing a robust combination of such data are still in their infancy in plant systematics. Here, we integrate multiple forms of data to better understand species delimitation and taxonomy within the eastern Asiatic genus Beesia (Ranunculaceae), traditionally thought to include two species. Specifically, 26 morphological characters were measured, plastome regions were compared to reconstruct evolutionary relationships, and novel karyotype data were generated. Morphological analyses parsed three species of Beesia, with little overlap expressed for most measured characters. Phylogenetic data reconstructed these species into monophyletic groups, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
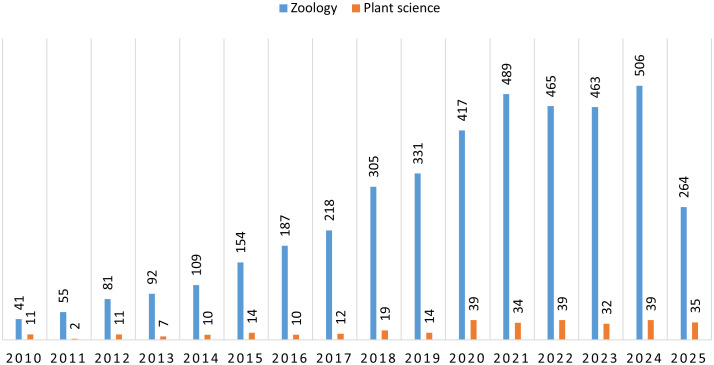 Figure 1
Figure 1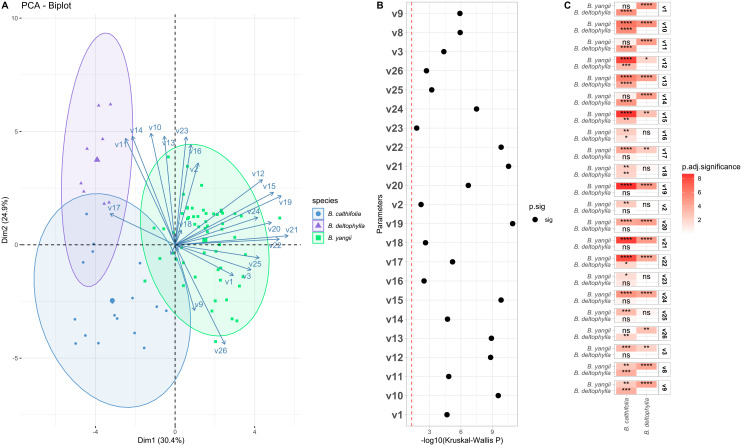 Figure 2
Figure 2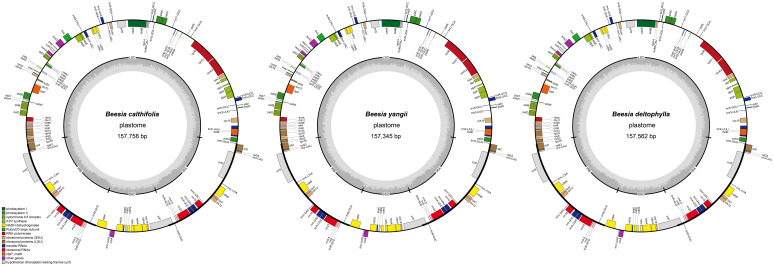 Figure 3
Figure 3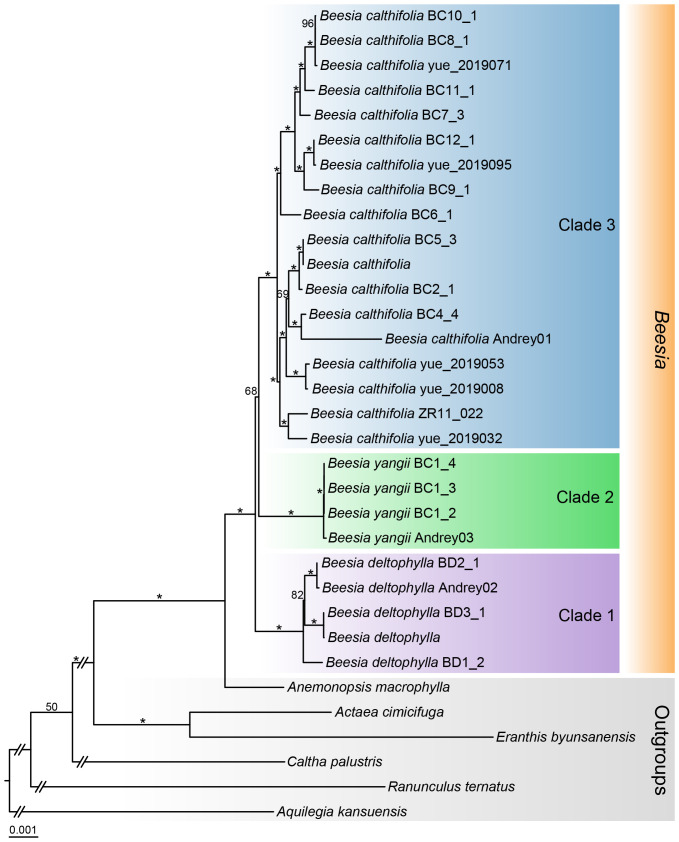 Figure 4
Figure 4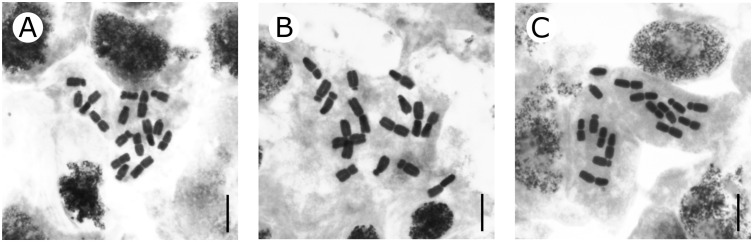 Figure 5
Figure 5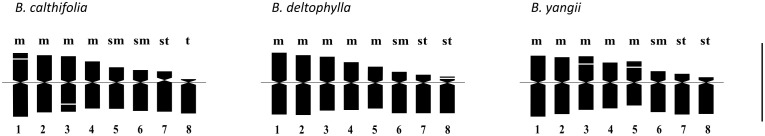 Figure 6
Figure 6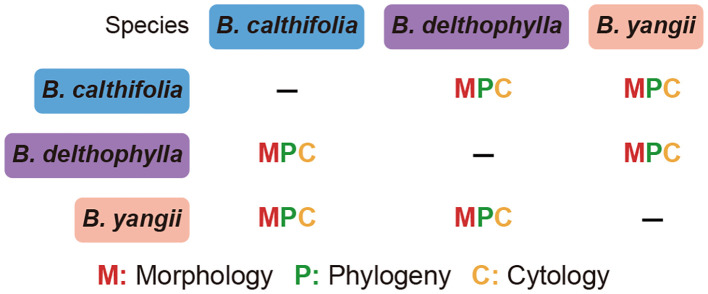 Figure 7
Figure 7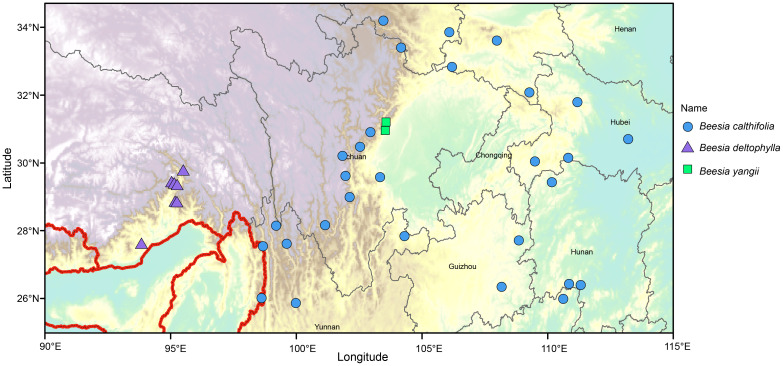 Figure 8
Figure 8 Figure 9
Figure 9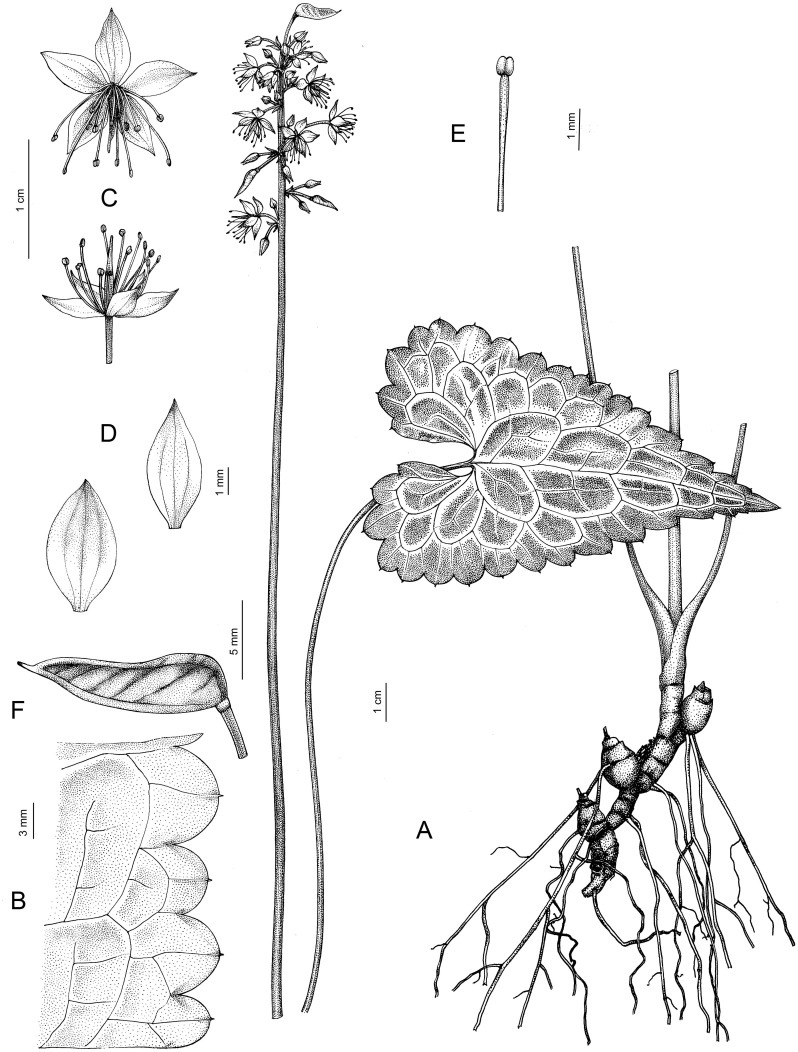 Figure 10
Figure 10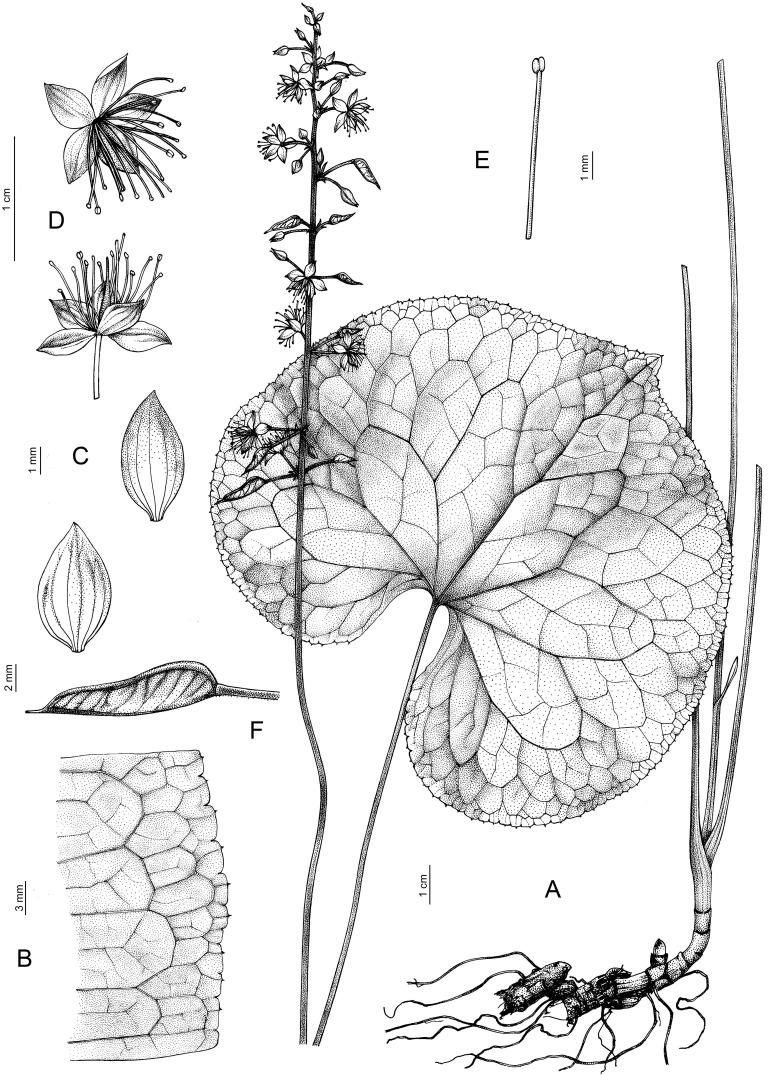 Figure 11
Figure 11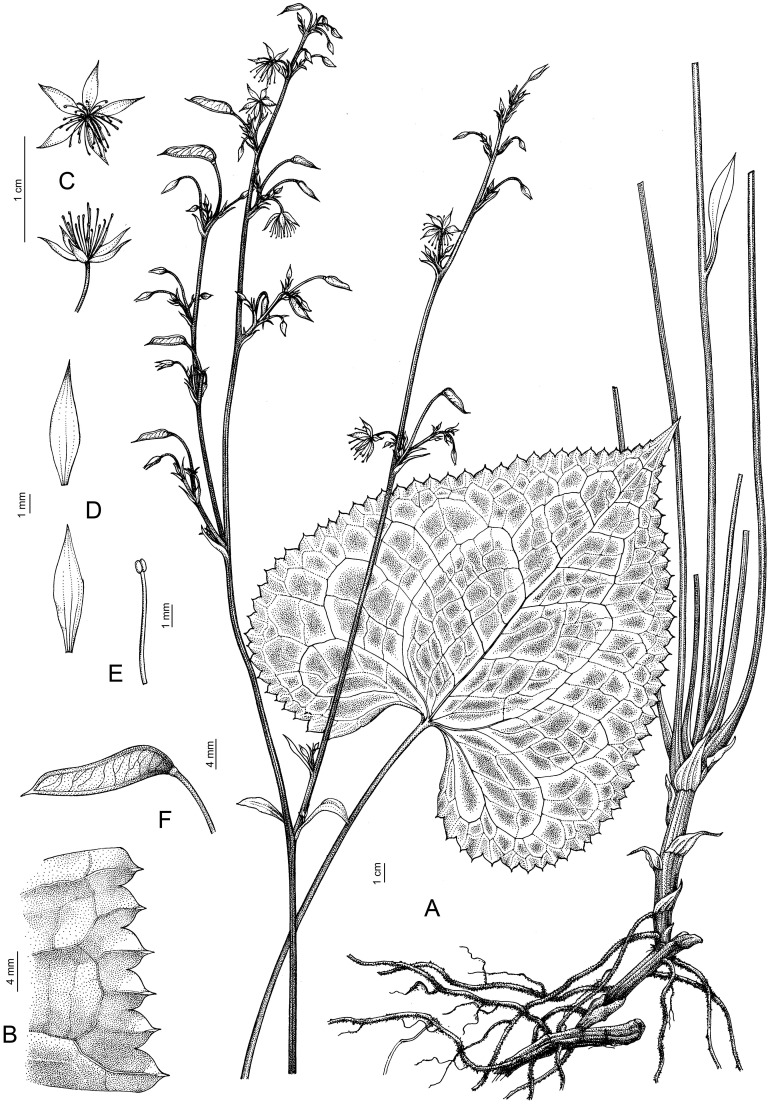 Figure 12
Figure 12| № | Characteristic |
|
|
|
|---|---|---|---|---|
| 1 | Plant height, cm* | 14–58 | 14–87.5 | 40.5–74.7 |
| – | Rhizome color | Purple or pink | Yellow or light brown | Purple or pink |
| 2 | Rhizome length, cm* | ~10 | ~15 | ~10 |
| 3 | Rhizome diameter, cm* | 0.5–0.7 | 0.3–0.9 | 0.4–1.1 |
| – | Scape trichome type | Unicellular lageniform | Unicellular lageniform | Unicellular lageniform |
| 4 | Scape pubescence in upper part (+/–)* | 1 | 1 | 1 |
| 5 | Scape pubescence in lower part (+/–)* | 1 | 1 | 1 |
| – | Leaf blade shape | Cordate-triangular | Reniform | Cordate |
| – | Leaf blade color | Dark green | Light green | Dark green |
| – | Leaf blade vein color | Light veins | Inconspicuous | Light veins |
| – | Leaf blade texture | Glossy | matte | Glossy |
| – | Leaf blade apex | Acuminate | Rounded to acuminate | Acuminate |
| 6 | Leaf petiole length, cm* | 5.5–15.0 | 5.6–26.5 | 8.4–31.15 |
| 7 | Leaf blade length, cm* | 2.1–8.4 | 2.4–9.5 | 1.9–13.5 |
| 8 | Leaf blade width, cm* | 1.9–4.5 | 4.0–13.6 | 3.0–12.0 |
| – | Edge of the leaf blade | Crenulate-mucronate | Crenulate-mucronate | Crenulate-mucronate |
| – | Shape of leaf teeth | Crenately lobed, each lobe with mucronate apex, some lobes doubly lobed | Toothed, widely crenate with long mucro at apex, sometimes off-center | Toothed, narrowly triangular with long sharp mucro |
| 9 | Number of teeth* | 14–51 | 22–179 | 26–120 |
| 10 | Length of the tooth at the base of the leaf, mm* | 1.66–5.62 | 0.83–1.86 | 1.05–2.98 |
| 11 | Width of the tooth at the base of the leaf, mm* | 2.58–8.31 | 1.9–5.84 | 0.98–5.31 |
| 12 | Length of the mucro at the base of the leaf, mm* | 0.48–1.1 | 1.29–0.65 | 0.5–1.0 |
| 13 | Length of the tooth middle part of the leaf blade, mm* | 1.46–4.36 | 0.78–1.67 | 1.23–2.56 |
| 14 | Width of the tooth middle part of the leaf blade, mm* | 3.49–7.97 | 1.87–5.90 | 1.45–6.97 |
| 15 | Length of the mucro middle part of the leaf blade, mm* | 0.43–1.0 | 0.31–0.69 | 0.45–0.94 |
| 16 | Length of the mucro at the apex the leaf blade, mm* | 2.79–11.99 | 2.05–9.72 | 2.54–14.46 |
| 17 | Width of the mucro at the apex the leaf blade, mm* | 2.62–7.11 | 2.13–11.49 | 0.85–6.93 |
| 18 | Length of the mucro at the apex of the leaf, mm* | 0.3–1.2 | 0.31–0.71 | 0.33–1.4 |
| 19 | Flower diameter, cm* | 0.6–0.9 | 0.8–1.1 | 1.5–2.2 |
| 20 | Length of peduncle, cm* | 0.4–1.0 | 0.3–0.8 | 0.5–1.0 |
| Sepal shape | Wide elliptic | Wide elliptic | Narrow elliptic | |
| Sepal color | White or pink | White or pink | White or white-green | |
| Sepals/stamens length ratio | >/= | < | >/= | |
| 21 | Sepal length, cm* | 0.4–0.5 | 0.4–0.6 | 0.5–0.9 |
| 22 | Sepal width, cm* | 0.2–0.25 | 0.15–0.20 | 0.2–0.4 |
| 23 | Stamens length, cm* | 0.4–0.6 | 0.30–0.31 | 0.4–0.8 |
| 24 | Number of flowers in an inflorescence* | 1–10 | 3–20 | 3–26 |
| 25 | Follicle length, cm* | 0.6–1.2 | 0.6–1.3 | 0.7–1.8 |
| 26 | Persistent style length, сm* | 0.1–0.2 | 0.1–0.3 | 0.1–0.2 |
| Species | Pair | CL, µm | r | RL, % | Type | THL | MCA | CVCL | CVCI |
|---|---|---|---|---|---|---|---|---|---|
|
| 1 | 8.57 ± 0.13 | 1.21 ± 0.05 | 8.21 | msat | 52.16 ± 1.58 | 28.18 ± 0.41 | 19.73 ± 0.80 | 36.27 ± 1.37 |
| 2 | 7.95 ± 0.39 | 1.08 ± 0.05 | 7.62 | m | |||||
| 3 | 7.36 ± 0.26 | 1.08 ± 0.04 | 7.06 | msat | |||||
| 4 | 6.48 ± 0.39 | 1.24 ± 0.08 | 6.21 | m | |||||
| 5 | 5.69 ± 0.33 | 1.76 ± 0.09 | 5.45 | sm | |||||
| 6 | 5.68 ± 0.35 | 2.30 ± 0.17 | 5.45 | sm | |||||
| 7 | 5.63 ± 0.31 | 3.37 ± 0.35 | 5.39 | st | |||||
| 8 | 4.81 ± 0.26 | 7.79 ± 0.53 | 4.61 | t | |||||
|
| 1 | 9.47 ± 0.66 | 1.03 ± 0.02 | 8.12 | m | 58.29 ± 2.92 | 26.72 ± 1.12 | 20.64 ± 1.19 | 36.27 ± 1.37 |
| 2 | 9.29 ± 0.49 | 1.14 ± 0.06 | 7.97 | m | |||||
| 3 | 8.16 ± 0.59 | 1.06 ± 0.04 | 7.00 | m | |||||
| 4 | 7.31 ± 0.48 | 1.26 ± 0.05 | 6.27 | m | |||||
| 5 | 6.29 ± 0.41 | 1.52 ± 0.05 | 5.40 | m | |||||
| 6 | 6.39 ± 0.14 | 2.58 ± 0.22 | 5.48 | sm | |||||
| 7 | 5.90 ± 0.38 | 3.67 ± 0.60 | 5.06 | st | |||||
| 8 | 5.48 ± 0.31 | 5.90 ± 0.65 | 4.70 | stsat | |||||
|
| 1 | 7.24 ± 0.23 | 1.20 ± 0.05 | 8.06 | m | 44.92 ± 0.35 | 25.80 ± 1.23 | 17.99 ± 1.60 | 34.86 ± 1.43 |
| 2 | 6.73 ± 0.37 | 1.21 ± 0.05 | 7.49 | m | |||||
| 3 | 6.31 ± 0.19 | 1.06 ± 0.02 | 7.02 | msat | |||||
| 4 | 5.42 ± 0.16 | 1.27 ± 0.04 | 6.03 | m | |||||
| 5 | 5.12 ± 0.08 | 1.10 ± 0.05 | 5.70 | msat | |||||
| 6 | 4.97 ± 0.14 | 2.43 ± 0.15 | 5.53 | sm | |||||
| 7 | 4.87 ± 0.18 | 3.45 ± 0.26 | 5.42 | st | |||||
| 8 | 4.27 ± 0.27 | 6.23 ± 0.61 | 4.75 | st |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Chromatography in Natural Products · Chromosomal and Genetic Variations
Introduction
1
Species delimitation based on morphology can be challenging due to intraspecific variation among populations and limited differentiation between closely related species (Whittall et al., 2004; Lissambou et al., 2019; Wang et al., 2022). Accurate species delimitation and identification play a pivotal role in the assessment, monitoring, and conservation of biological diversity (Agapow et al., 2004; Carstens et al., 2013). Taxonomic study of biodiversity is particularly complicated by the presence of cryptic species, defined as two or more separate species that are incorrectly classified as a single species (Bickford et al., 2007; Fu et al., 2023). They may be barely distinguishable morphologically, albeit along different evolutionary trajectories (Struck et al., 2018; Fu et al., 2023). These hidden or unrecognized species represent a substantial part of undescribed biodiversity, posing challenges for realizing comprehensive taxonomy and robust biodiversity assessments and implementing well-informed conservation plans (Renner, 2020; Fu et al., 2023). To date, there remains no unified species concept relating to the angiosperm clade. This is because a “gray zone” frequently exists, in which boundaries between taxa become murky and may be difficult to define (De Queiroz, 2007; Bock et al., 2023). In contrast to oftentimes more simple cases of biological species resolution in the animal clade, the angiosperm clade in particular commonly exhibits, e.g., widespread apomixis and/or self pollination potentials, rampant hybridization across time and space (sometimes itself resulting in formation of novel lineages with new genetic architecture), phenotypic variation across often relatively large geographic ranges and/or phenotypic plasticity, and myriad forms of clonal reproduction in absence of the sexual process (Hörandl, 2010; Grossenbacher et al., 2017, Kremer and Hipp, 2020, Karbstein et al., 2024; Sharples and Manzitto-Tripp, 2024; Chambers et al., 2025, Erst et al., unpubl. data). These factors and others have led to multiple interpretations and categories of species delimitation in angiosperms, these often taxon-dependent.
Cryptic diversity can arise through different mechanisms (Liu et al., 2022; Li and Wiens, 2023), such as weak morphological differentiation among geographically proximate yet reproductively isolated populations (Sharples, in prep.), rapid radiation (Fior et al., 2013), convergent evolution driven by similar ecological pressures (Yeaman et al., 2016; Kinosian et al., 2020), hybridization (Aköz and Nordborg, 2019; Wróbel et al., 2024), and/or changes in chromosome structure (Erst et al., 2020; Mitrenina et al., 2024). An important approach to species identification and conservation of rare species is standard single-locus Sanger markers (Trivedi et al., 2016), which has also been useful in solving problems in taxonomy (Vargas et al., 2012; Ali et al., 2014). Standard single-locus Sanger markers can help to quickly and accurately identify species and has the potential to lead to the discovery of new species (Erst et al., 2022). However, standard single-locus Sanger markers are not always effective in systematic biology, especially when considering taxa that radiated recently or those that possess otherwise complex evolutionary histories (Percy et al., 2014; Yan et al., 2015). To address these challenges in part, a growing number of studies have involved multiple lines of evidence in species delineation, termed the integrative approach (Erst et al., 2020). This approach is especially effective for delimiting species with previously uncertain and unrecognized taxonomic status (e.g., Erst et al., 2020; Meudt et al., 2025; Wang et al., 2025).
Searches conducted on the ISI Web of Science (www.webofscience.com/wos/alldb/basicsearch, last accessed on 7 August 2025) using “integrative taxonomy” in the title and/or abstract revealed that zoological studies using integrative taxonomy have rapidly increased recently, whereas botanical studies have rarely adopted this method (Figure 1). Recent taxonomic revisions and descriptions of new plant taxa increasingly rely on a combination of morphological traits and genetic data. However, plant systematics frequently encounters taxonomically complex groups (TCGs), in which overlapping evolutionary processes—such as hybridization, polyploidy, asexual reproduction, and/or incomplete reproductive isolation—obscure clear morphological and genetic distinctions (Ennos et al., 2005). These complexities often necessitate the integration of complementary methodologies, including cytogenetic, biochemical, and population genetic analyses, alongside traditional morphological and molecular approaches (Erst et al., 2020).
Diagram showing how many studies using “integrative taxonomy” in the title and/or abstract were conducted in zoology and plant science research from 2010 to 2025.
The Sino-Himalayan region is a temperate biodiversity hotspot with high levels of species endemism (Myers et al., 2000; Sun et al., 2017; Erst et al., 2023). It is also a modern center of distribution and diversification of elements of the temperate flora in the Northern Hemisphere (Peng et al., 2023; Sharples and Manzitto-Tripp, 2024). The Qinghai-Tibet Plateau (QTP), known as the “Roof of the World” and the “Asian water tower,” stands at a minimum elevation of 4,500 m high over a region of approximately 2.5 × 10^6^ km^2^ and consists of three subregions: the Plateau Platform, the Himalaya, and the Hengduan Mountains (Mao et al., 2021; Wu et al., 2022). At present, the QTP houses more than 1,500 genera and at least 12,000 species of vascular plants, of which more than 25% are endemic to this region (Wu and Wu, 1996; Wen et al., 2014). Although the biodiversity of the region has been extensively studied (e.g., Xue et al., 2021; Zhang et al., 2023), new taxa are still being described and the broader QTP is expected to harbor more undescribed taxa for decades to come.
The tribe Cimicifugeae Torr. & A.Gray belongs to the family Ranunculaceae Juss. and includes four genera and more than 49 species (Compton et al., 1998; Wang et al., 2005; Yuan and Yang, 2006; Handel-Mazzetti, 1922). This tribe contains Anemonopsis Siebold & Zucc. (one species), Actaea L. (32 species), Eranthis Salisb. (14 species), and Beesia Balf.f. & W.W.Sm. (two species), and the tribe as a whole exhibits a circumboreal distribution (Compton and Culham, 2002; Ling et al., 2023). Two genera are restricted regional endemics, i.e., Anemonopsis is known only from Japan, and Beesia occurs only in China, India, and Northern Myanmar (Wang et al., 2001; Bhaumik and Krishna, 2024; Kostikova et al., 2024; Liangqian and Tamura, 2001). The genus Beesia was originally described by Balfour and Smith (1915), and the new taxon was thought to be closely related to the Japanese genus Glaucidium Siebold & Zucc. and the Japanese-American genus Hydrastis J.Ellis. In the same publication, the authors also described a new species: Beesia cordata Balf.f. & W.W.Sm. from Northern Burma (Myanmar). Before the introduction of the genus Beesia, B. cordata had already been described by Daniel Oliver based on material collected by Karl Johann Maximowicz (in Hooker’s Icones Plantarum: Oliver, 1888) as Cimicifuga calthifolia Maxim. ex Oliv.; therefore, Beesia cordata is currently recognized as a synonym of B. calthifolia (Maxim. ex Oliv.) Balf.f. & W.W.Sm (Balfour and Smith, 1924; Tiwari and Mao, 2016). The other currently recognized species of Beesia is B. deltophylla C.Y.Wu. Within its range, B. calthifolia is represented by populations that exhibit significant morphological differences, and furthermore, some differences in karyotype structure and ploidy have been noted by previous authors within B. calthifolia (Yang et al., 1995). Recent molecular phylogenetic studies have shown multiple clades among populations of B. calthifolia populations depending on the altitude and climatic conditions of a habitat (Peng et al., 2023).
While the tribe Cimicifugeae has been studied in detail by many authors utilizing individual methods, an integrative approach has not been applied to any members of the group until now. In the present study, we explore multiple levels of diversity of the genus Beesia to forward a taxonomic revision of this group. We utilize an integrative approach involving morphological, phylogenetic, and cytogenetic analyses to evaluate species delimitation and to study the taxonomy of this group. A combination of the above approaches has been successfully applied in previous studies of related taxa (Erst et al., 2020, Erst et al. in review, Eranthis), and other studies have also taken an integrative approach to taxonomy, using different sets of methods (Meudt et al., 2025; Wang et al., 2025, etc.). Based on these multiple lines of evidence, a revised treatment of species boundaries and a revision within this genus are presented. Three suitable lectotypes for Cimicifuga calthifolia Maxim. ex Oliv., Beesia cordata Balf.f. & W.W.Sm., and Beesia elongata Hand.-Mazz. were selected in accordance with articles 9.3 and 9.12 of the ICN (Turland et al., 2025).
Materials and methods
2
Plant material
2.1
More than 100 herbarium specimens from numerous localities were freshly collected during field investigations in China (Sichuan Province and Tibet Autonomous Region) during the 2024–2025 field seasons. Dried and preserved specimens were deposited in the NS (I.M. Krasnoborov Herbarium of the Central Siberian Botanical Garden of the Siberian Branch of the Russian Academy of Sciences, Novosibirsk, Russia), IRK (Herbarium of the Siberian Institute of Plant Physiology and Biochemistry of the Siberian Branch of the Russian Academy of Sciences, Irkutsk, Russia), and LE (Komarov Botanical Institute of the Russian Academy of Sciences, St. Petersburg, Russia) Herbaria (herbarium abbreviations according to Thiers, 2018) (Supplementary Table S1). A review of existing herbarium materials was undertaken through digitized images or physically in Herbaria at ARUN, BM, CAL, E, K, KUN, LE, NS, NY, P, PE, TI, US, and WU. Drawings of Beesia yangii sp. nov. (1), B. calthifolia (2), and B. deltophylla (3) were created based on images of type (1: NS0000490–NS0000492) and non-type specimens (2: NS0003871, NS0003872; 3: NS0033868–NS0033870). Maps of records were prepared with R (v4.4.1; R Core Team, 2024).
Morphological analysis
2.2
A total of 26 different morphological traits were quantitatively measured or otherwise assessed from the above herbarium materials (Table 1). After measurement or assessment of traits, analysis of morphological data was conducted using R with the following packages (v4.4.1; R Core Team, 2024). In case of unavailable or otherwise missing data values, missing-measurement imputation was performed with the missMDA package (v1.19; Audigier et al., 2017). To determine if morphospecies clustered or not, principal component analysis was performed with the help of the FactoMineR package (v2.11; Lê et al., 2008). Visualization of the morphological data was conducted using packages factoextra (v1.0.7; Kassambara and Mundt, 2020), ggplot2 (v3.5.1; Wickham, 2016), and ggpubr (v0.6.0; Kassambara, 2023). For assessment of the specimens’ data distribution, ad hoc (Kruskal–Wallis) and post-hoc (Mann–Whitney–Wilcoxon) tests were carried out with the rstatix package (v0.7.2; Kassambara, 2023).
Molecular analysis
2.3
Taxon sampling and sample sequencing
2.3.1
A total of 27 individuals of Beesia, representing all three species (including the new species), were included in molecular phylogenetic analysis. One species of Actaea, one species of Anemonopsis, and one species of Eranthis, covering all three other genera of Cimicifugeae, were also included. On the basis of other studies (e.g., Wang et al., 2009, 2016), three species—Ranunculus ternatus Thunb., Aquilegia kansuensis (Brühl) Erst, and Caltha palustris L.—were selected as outgroups, and most ingroup samples were obtained from Peng et al. (2023) (Supplementary Table S2). For three newly sequenced individuals, genomic DNA of samples was extracted from silica gel-dried leaves or herbarium specimens and was purified using the Tiangen Isolation/Extraction/Purification Kit (Tiangen Biotech [Beijing] Co., Ltd.). Genomic libraries targeting fragment lengths of 350 base pairs (bp) (via sonication) were prepared off-site for sequencing on the Illumina HiSeq X-Ten platform to assemble plastomes from the above samples based upon paired-end reads with lengths of 150 bp (Novogene [Beijing] Co., Ltd.). Approximately 10 GB of raw sequencing data was newly generated for each sample. To obtain high-quality genome sequences, raw reads were processed using Fastp v0.19.7 (Chen et al., 2018) to remove adapter sequences and low-quality reads. Peng et al. (2023) did not include nuclear data, and thus our new datasets likewise only represented plastid sequences.
Data assembly and annotation
2.3.2
The plastome for newly generated samples was assembled de novo in GetOrganelle v1.7.6.1 (Jin et al., 2020) with default parameters. The obtained plastid contigs were further visualized and scaffolded to complete circular genomes in Bandage v0.8.1 (Suyama et al., 2006). Highly accurate annotation of organelle genomes was performed by using the Organellar Genome GeSeq tool (Tillich et al., 2017) with subsequent manual correction. Two plastomes from Beesia deltophylla (GenBank accession No. NC_072729) and B. calthifolia (NC_041531) were used as reference sequences. We employed OGDRAW v1.3.1 (Greiner et al., 2019) to visualize the circular plastome map. Amino acid sequences of 78 plastid protein-coding-gene regions were extracted for phylogenetic analysis, and each was aligned in MAFFT v7 (Katoh and Toh, 2010). Finally, the corresponding three-codon-aligned DNA sequences were generated from the amino acid alignments using PAL2NAL v14 (Suyama et al., 2006).
Phylogenetic analysis
2.3.3
Phylogenetic analyses of ingroup and outgroup samples were conducted based on concatenated DNA sequences from the coding regions of 78 protein-coding genes. Maximum likelihood (ML) analysis was performed using RAxML v8.2.11 (Stamatakis, 2014) with 1,000 replicates under the GTRGAMMA model.
Cytogenetic analysis
2.4
Cytogenetic analyses were performed on seven populations. Somatic chromosomes of the three species of Beesia were prepared from root meristems. Rhizomes were germinated at room temperature for 1–3 months. Newly formed 1–2-cm-long roots were excised and pretreated with a 0.5% colchicine solution for 3–4 h. Roots were then fixed in a mixture of 96% ethanol and glacial acetic acid (3:1). Chromosomes were stained with 1% aceto-hematoxylin and then subjected to morphometric analyses. Staining and karyotyping were performed according to standard protocols (Smirnov, 1968; Mitrenina et al., 2021). Mitotic metaphase chromosome plates were examined under an Axiostar microscope (Carl Zeiss, Munich, Germany) and photographed using an Axio Imager A.1 microscope (Carl Zeiss, Germany) equipped with an AxioCam MRc5 CCD camera (Carl Zeiss, Germany) at 1,000× magnification by means of AxioVision 4.7 software (Carl Zeiss, Munich, Germany) in the Laboratory for Ecology, Genetics and Environmental Protection (EcoGene) of the National Research Tomsk State University (Tomsk, Russia). KaryoType 2.0 software (Altınordu et al., 2016) was employed for karyotyping, and Adobe Photoshop CS5 (Adobe Systems, USA) and Inkscape 0.92 (USA) for image editing.
Symbols used to describe the karyotypes matched those defined elsewhere (Levan et al., 1964): m = a median centromeric chromosome with an arm ratio (r) of 1.0–1.7 (metacentric chromosome); sm = a submedian centromeric chromosome with an arm ratio of 1.7–3.0 (submetacentric chromosome); st = a subterminal centromeric chromosome with arm ratio 3.0–7.0 (subtelocentric chromosome); t = a terminal centromeric chromosome with arm ratio 7.0 or more (telocentric chromosome); T = a chromosome without an obvious short arm. Mean values of the arm ratio (r) and relative chromosome length (RL) for each chromosome pair and total haploid length (THL) were determined. In addition, we calculated the coefficient of variation of chromosome length (CV_CL_) (Paszko, 2006), the coefficient of variation of the centromeric index (CV_CI_) (Paszko, 2006), and mean centromeric asymmetry (M_CA_) (Peruzzi and Eroğlu, 2013).
Conservation assessment
2.5
To assess the conservation status of species of Beesia, we utilized IUCN Red List criteria for placing taxa into one of six categories: critically endangered (CR), endangered (EN), vulnerable (VU), near threatened (NT), least concern (LC), or data deficient (DD), according to current guidelines and criteria defined by the IUCN Standards and Petitions Committee (2024). In general, there are five evaluation criteria defined by the IUCN Standards and Petitions Committee (2024): population size reduction (criterion A); geographic range size fragmentation, few locations, range decline, or population fluctuations (B); small and declining population size and fragmentation, fluctuations, or few subpopulations (C); very small population or very restricted distribution (D); and quantitative analysis of extinction risk (E). Among these, criterion B—severity of range fragmentation—is suitable for estimating conservation status, even if data are limited and the distribution of a taxon is known from only few georeferenced records. In this study, we used criterion B because distribution records of the three species of Beesia have been relatively well compiled. Specifically, these distribution records were extracted from herbarium labels at the above institutions, including those from our own collections and associated published research. Subcriteria B1 (Extent of Occurrence, EOO) and B2 (Area of Occupancy, AOO) were estimated with the ConR package (Dauby et al., 2017) in R (R Core Team, 2024). The minimum AOO was estimated based on a user-defined grid cell of 2 km^2^ as recommended by the IUCN (2024). In particular, for the AOO and EOO, the ConR package provides the ability to incorporate the unique number of occurrences, the number of subpopulations, and locations of each taxon into analyses.
Results
3
Morphological analysis
3.1
Results of principal component analysis and the significance of interspecies differences suggested recognition of three taxa of Beesia (Figure 2). The largest contribution to the first two principal components (PCs) was made by variables 19 (flower diameter) and 21 (sepal length) (Table 1). Data areas of B. calthifolia and B. yangii sp. nov. were separated with a small intersection. There was separation across gradients of all tested parameter values except for parameters 11 (width of the tooth at the base of the leaf), 14 (width of the tooth middle part of the leaf blade), and 26 (persistent style length) (Table 1). The data area of B. deltophylla was found to be completely separated from the area of B. yangii but exhibited an intersection with the area of B. calthifolia (Figure 2A). Categorical variables 4 (scape pubescence in upper part) and 5 (scape pubescence in lower part) were identical among the examined species.
*Results of principal component analysis. (A) Principal component analysis biplot of measured parameters between different species of Beesia. (B) Dot plot of the significance of the Kruskal–Wallis test [p-value: −log10(P)] of measured parameters. The vertical red dashed line represents the p = 0.05 threshold. (C) A heatmap of adjusted p-values (Benjamini–Hochberg correction) from the interspecies pairwise Mann–Whitney–Wilcoxon test of measured parameters. Asterisks represent adjusted p-values (significance): ns, not significant, *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.00001.
The Kruskal–Wallis test revealed that all the variables have statistically significant interspecies differences (Figure 2B). Pairwise interspecies comparisons of measured parameters are given in Figure 2C.
Molecular analysis
3.2
The plastomes of species of Beesia that were assembled in this study exhibited complete sizes from 157,345 bp in B. yangii sp. nov. to 157,758 bp in B. calthifolia (Figure 3). All plastomes exhibited the quadripartite structure typical of many plastomes, which comprises a pair of IRs (inverted repeats) (26,516–26,584 bp) separated by an LSC (long single copy) region (86,971–87,252 bp) and an SSC (small single copy) region (17,206–17,474 bp; Figure 3). We identified 130 predicted functional genes and one pseudogene (ψycf1 in IR_B_/SSC) for the plastomes of Beesia. These genes were summarized as 112 unique genes (with 18 of them repeated) including 78 protein-coding genes, 30 transfer RNA (tRNA) genes, and 4 ribosomal RNA (rRNA) genes (Figure 3).
Plastome maps of species of Beesia.
The DNA supermatrix comprised 70,009 aligned nucleotides from the 78 protein-coding genes. Maximum likelihood analysis indicated that Cimicifugeae is strongly supported as monophyletic (bootstrap support [BS] = 100%) (Figure 4). Within Cimicifugeae, Actaea clustered with Eranthis (BS = 100%). Beesia was reconstructed as a monophyletic group with strong support (BS = 100%) and as sister taxon to Anemonopsis (BS = 100%). In Beesia, three major clades (1, 2, and 3) were identified, corresponding to the species B. deltophylla (BS = 100%), B. yangii (BS = 100%), and B. calthifolia (BS = 100%), respectively. Beesia yangii and B. calthifolia formed a clade (BS = 68%) that was reconstructed as sister to B. deltophylla (BS = 100%).
The ML tree inferred from the combined 78 plastid protein-coding-gene regions with numbers above branches representing bootstrap values (BS > 50%). The BC1–BC12 and BD1–3 populations correspond to the BC1–12 and BD1–3 populations in Peng et al. (2023). “” denotes BS = 100%.*
Cytogenetic analysis
3.3
Karyotypes of B. calthifolia (Sichuan Province), B. deltophylla (Mêdog County), and B. yangii sp. nov. (Sichuan Province) were newly generated. All the studied specimens were found to be diploid with 2n = 16 (Figure 5). Karyometric analysis was performed on chromosomes with different degrees of compaction (estimated using total haploid length) for more accurate determination of karyotype formulas. The mean values (Table 2) were obtained from measurements in metaphase plates having similar degrees of chromosome compaction, usually the highest among those available on a cytological slide. Intrachromosomal and interchromosomal asymmetry and heterogeneity of the centromeric index were similar among the three examined species of Beesia (Table 2).
Mitotic metaphase chromosomes of the three species of Beesia. (A) B. calthifolia, 2n = 16. (B) B. deltophylla, 2n = 16. (C) B. yangii, 2n = 16. Scale bars = 10 µm.
Karyotype structure of the studied specimens of B. calthifolia corresponds to the formula 2n = 4m + 4m^sat^ + 4sm + 2st + 2t (Figure 6). Metacentric chromosomes differed in length (relative length varies on average from 6.21% to 8.21%), with the second and third pairs slightly more symmetrical than the first and fourth pairs (Table 2). The first and third longest pairs, as a rule, bore secondary constrictions. The fifth and sixth pairs of chromosomes were submetacentric, had similar lengths (relative length is on average 5.45%), but could be well differentiated by the arm ratio. This ratio of the fifth pair was close to a borderline value, and the type of these chromosomes is metacentric according to Levan et al. (1964). The seventh pair was close in length to the submetacentrics (relative length on average 5.39%) but is subtelocentric. The eighth pair was the shortest (relative length 4.61% on average) and is telocentric.
Haploid idiograms for the three species of Beesia. m—metacentric chromosome; sm—submetacentric chromosome; st—subtelocentric chromosome; t—telocentric chromosome; 1–8—chromosome pairs. Scale bar = 10 µm.
The karyotype formula in the studied specimens of B. deltophylla was 2n = 10m + 2sm + 2st + 2st^sat^ (Figure 6). Chromosomes of pairs 1 and 2 were similar in length (relative length of 8.12% and 7.97%, respectively, on average) but usually could be differentiated by the arm ratio (Table 2). Pairs 3, 4, and 5 differed both in length and in the arm ratio. The shortest of them was the most asymmetric. Unequal-armed chromosomes were represented by a pair of submetacentrics and two pairs of subtelocentrics of gradually decreasing length. Chromosomes of the eighth pair were formally classified as the subtelocentric type according to the average arm ratio, although on some metaphase plates, this parameter was very close to a value borderline with the telocentric type of chromosomes: 7.0. This pair of chromosomes was found to have small satellites on the short arm.
Karyotype structure of the studied specimens of B. yangii is expressed by the formula 2n = 6m + 4m^sat^ + 2sm + 4st (Figure 6). Pairs of metacentric chromosomes differed well in length and in the arm ratio. Chromosomes of pairs 3 and 5 bore secondary constrictions. The sixth and seventh pairs were similar in length but differed in the arm ratio: one is submetacentric, and the other is subtelocentric. The eighth pair was the shortest and is subtelocentric.
Discussion
4
Cryptic diversity of Beesia taxa in East Asian region and description of new species
4.1
In this study, a framework for species delimitation in Beesia has been formulated, as derived through utilization of molecular, morphological, and cytogenetic data across populations of its species. Through these data using the integrative taxonomic approach, a new species of the genus Beesia has been uncovered, and lectotypes were selected (see taxonomic treatment below). All approaches, including genomic, cytogenetic, and morphological analyses, support the description of the new species and allow for comprehensive comparison of differences among the three species (Figure 7). Our phylogenetic analyses strongly supported Beesia as monophyletic (Figure 4), in agreement with previous studies (Peng et al., 2023).
Schematic illustration of species delineation within Beesia and various sources of information used to distinguish species.
Recent studies on the biogeographic histories of Megacodon (Hemsl.) Harry Sm. and Beesia by means of various data (ddRAD-seq, chloroplast genomes, molecular fragments, and morphological data) to identify similarities in evolutionary histories between the two genera showed that Beesia and Megacodon began to diverge from the late Miocene onward, with ancient allopatry at lower elevations resulting in narrow species ranges or narrow distributions of relict populations (Peng et al., 2023). Such processes may help explain why the two sister species of Beesia (B. calthifolia and newly described B. yangii) were reconstructed along distinct evolutionary trajectories. Mantel tests between genetic distance and climatic, elevational, or geographic distance revealed an isolation-by-distance pattern in Beesia and Megacodon stylophorus (C.B.Clarke) Harry Sm (Peng et al., 2023). Beesia calthifolia exhibited a genetic divergence pattern along an elevation gradient. Furthermore, we performed morphological measurements on B. calthifolia and found that different elevational groups have distinct leaf shapes (Peng et al., 2023). Nevertheless, the phylogeny of Beesia in this work uncovered genetic heterogeneity within populations of B. calthifolia. The plastome, cytogenetic, and morphological data obtained here allowed for a more robust taxonomic assessment compared with prior work in the group and supported the monophyly of the new species from Sichuan Province (China) and of two related taxa.
For many genera, and especially for endemic and rare taxa poorly represented in herbarium collections, morphological identification criteria are poorly developed (e.g., many representatives of the Ranunculaceae). This work presents new morphological (and geographical) comparisons that are suitable for identifying species of the genus Beesia (see key below). The newly identified trait states, such as those involving leaf texture, the presence of colored leaf veins, shape and size of leaf teeth, width and length of sepals, and the ratio of stamens’ and sepals’ lengths, may now be utilized by field and herbarium botanists alike. Beesia calthifolia differs from other species of the genus by its reniform basal leaves with a light-green (with inconspicuous veins) dull surface, the apex rounded or acuminate, the blade with toothed edges (these teeth sometimes off-center) that are widely crenate with a long mucro at the apex, a yellow or light brown rhizome, and sepals that are shorter than the stamens. In contrast, the other two species produce basal leaves that are cordate or cordate-triangular, with a dark-green [with lighter-green veins] glossy surface and an acuminate apex, the margins toothed to crenately lobed, the teeth narrowly triangular with a long sharp mucro or each lobe with mucronate apical teeth, some lobes doubly lobed, a pink or purple rhizome, and sepals that are longer than the stamens. The new species B. yangii bears greater morphological similarity to B. deltophylla, from which it differs in producing a cordate basal leaf blade, this with margins of narrowly triangular teeth with a long sharp mucro, and narrowly elliptic sepals that are white or white green (vs. cordate-triangular basal leaf blades with crenately lobed margins, the teeth of the margins with a mucronate apex, some lobes doubly lobed, and widely elliptic sepals that are white or pink). Furthermore, each of the three species bears a unique geographical distribution: B. deltophylla and the new species B. yangii occur in more restricted ranges, whereas B. calthifolia is more widespread (Figure 8).
Distribution map of known localities of occurrence for species of Beesia.
Karyotypes of B. calthifolia and B. deltophylla have been previously described in a number of studies (Shang, 1985; Yang et al., 1995; Yang, 1999, 2002, Yuan and Yang, 2006). For B. calthifolia, two cytotypes have been identified: diploid with 2n = 16 and tetraploid with 2n = 32 (Yang, 1999). At first, the karyotype structure of this species was described by the formula 2n = 6m + 4m^sat^ + 2sm + 2sm^sat^ + 2t (Shang, 1985). The karyotype was later revised and determined to exhibit the formula 8m + 4sm + 2st + 2t for diploid individuals and 2n = 16m + 8sm + 4st + 4t for tetraploid individuals (Yang, 1999). In later studies by the same author, for several diploid populations of B. calthifolia, the karyotype formula 2n = 6m + 4m^sat^ + 2sm + 2st + 1t + 1t^sat^ was reported, similarly with Shang’s original report (Shang, 1985; Yang, 2002; Yuan and Yang, 2006). In our study, the karyotype structure in analyzed specimens matches closely with the one from Yang’s earlier study, namely, 2n = 4m + 4m^sat^ + 4sm + 2st + 2t (Yang, 1999). We also detected secondary constrictions on two pairs of metacentric chromosomes. It should be noted that, according to our results and those of Yang, the fifth pair of metacentric chromosomes exhibits an arm ratio close to a borderline value between metacentric and submetacentric types, which may help explain the difference between the karyotype formulas, containing either four or five pairs of metacentrics.
The karyotype structure of B. deltophylla described by us also differs slightly from Yang’s results (Yang et al., 1995). According to his work, this species exhibits the formula 2n = 10m + 4st + 2t^sat^, whereas we obtained a similar formula of 2n = 10m + 2sm + 2st + 2st^sat^. The specimens that we examined are characterized by the presence of a sixth pair of submetacentric chromosomes with an arm ratio of 2.58 ± 0.22. In prior work, this chromosome has an intermediate value between the sm and st types: 3.02. The eighth pair of chromosomes in our specimens displays an arm ratio close to a borderline value with the telocentric type, but we formally assigned it to the submetacentric type. The arm ratio of unequal-armed chromosomes varies more substantially among metaphase plates than that of equal-armed chromosomes, a phenomenon which may be due to uneven compaction when short arms are compared with long arms, thus potentially explaining discrepancies in findings among materials of B. deltophylla. The karyotype of the new species B. yangii is similar to the karyotype of B. deltophylla but shows differences from it in the position of secondary constrictions and in the morphology of fifth-pair chromosomes (see Results). Thus, chromosome sets of the three species of Beesia exhibit species-specific features.
Taxonomic treatment
4.2
The above data allowed us to distinguish a new species of Beesia from specimens previously identified as B. calthifolia.
Genus Beesia Balf.f. & W.W.Sm. 1915, Notes Roy. Bot. Gard. Edinburgh 9: 63; Liangqian and Tamura, 2001, Fl. China 6: 142; Kress et al., 2003, Contr. U.S. Natl. Herb. 45 (Checkl. trees, shrubs, herbs, climbers Myanmar): 333; Mao and Dash, 2020, Fl. Pl. India Annot. Checkl. Dicot. 1: 41.
Type: Beesia cordata Balf.f. & W.W.Sm. (= B. calthifolia (Maxim. ex Oliv.) Balf.f. & W.W.Sm.) (see next).
Description: Perennial herbs. Rhizome creeping or ascending vigorously. Stem erect, simple, subscapose, elongated after flowering with several large membranaceous sheaths at base. Basal leaves 2–4, simple, long petiolate, blades undivided, cordate or cordate-triangular to reniform at base, round to acuminate at apex, regularly dentate, chartaceous, petioles dilated into membranaceous sheath toward base. Cauline leaves present. Lower cauline leaves similar to the basal leaves, but smaller in size, upper cauline leaves 1 or 2, lanceolate. Inflorescence racemiform in upper part, with one to three flowers fasciculate at nodes in the lower part. Scape simple, with membranaceous sheath at base. Cyme compound, with one to three sessile fascicled flowers at several nodes. Bracts lanceolate or subulate. Flowers actinomorphic, opening flat. Sepals 5, petaloid, white, white-green or pink, wide or narrow elliptic, acute at apex, spreading at end of flowering stage. Petals absent. Stamens numerous, shorter, equal or longer than sepals, anthers nearly globose, lateral, filaments filiform or subfiliform. Carpel single. Follicle sessile, transversely veined, directed sideways, with distinct persistent stylodium. Seeds several in follicle, elliptic or ovoid, rugulose.
Distribution: Three species in China and Myanmar.
- Beesia deltophylla C.Y.Wu, 1979, Fl. Reipubl. Popularis Sin. 27: 604; Li and Tamura, 2001, Fl. China 6: 142. (Figures 9A, D, G, 10).
Morphological differences among species of Beesia: (A, D, G) B. deltophylla. (B, E, H) B. calthifolia. (C, F, I) B. yangii. (A), (B), (C) Leaves. (D-F) Flowers. (G-I), (I0, Follicles.
Beesia deltophylla. (A) General view. (B) Edge of the leaf. (C) Flowers. (D) Sepals. (E) Stamen. (F) Follicle.
Type: CHINA. Tibet, Metuo, Lage to Hanmi, 2300 m, in the broad-leaved forest on the hillside, white flowers, 3 August, 1974, P.G. Xiao 74–3975 [in Chinese] (holotype, PE! barcode PE00028304; isotype, PE! barcode PE00440501).
– Beesia calthifolia auct. non (Maxim. ex Oliv.) Balf.f. & W.W.Sm.: U.L. Tiwari & A.A. Mao, 2016, Indian Forester 142(5): 508, with auth. comb. Ulbr.; A.A. Mao & S.S. Dash, 2020, Fl. Pl. India Annot. Checkl. Dicot. 1: 41.
Description: Perennial herbs. Rhizome up to ~10 × 0.3–0.7 cm, pink or purple. Scapes more than 14 cm at anthesis (–34 cm at fruiting) × 1–2 mm, basally subglabrous, apically spreading white pubescent with unicellular lageniform trichomes. Basal leaves 2 or 3; petiole 5.5–14.5 cm long, densely spreading white pubescent, dilated into membranaceous sheath toward base; leaf blade ovate-triangular, 4–9 × 2.8–5.9 cm, both surfaces glabrous, abaxially pale green, adaxially dark green with light green or white veins, glossy; base deeply cordate, margin sparsely 7–16-dentate on each side, teeth 3–8 mm wide at base, apex acuminate to long acuminate. Lower cauline leaves rarely present, same as basal leaves, but smaller in size, upper cauline leaves 1 or 2, lanceolate. Inflorescence paniculate-cymose, 5–10 сm long; bracts subulate, 1–4 mm long. Flowers 6–7 mm in diam.; pedicels 5–10 mm long, densely spreading pubescent. Sepals white or pink, ovate-elliptic, 4–4.5 × 2–2.5 mm, glabrous, apex acute. Stamens 4–6 mm long, equal to or mostly longer than the sepals; anthers yellowish, 0.25 mm long. Follicle green, sometimes turning yellowish brown, ca. 1 cm long, flat, lanceolate-linear, sparsely pubescent, with ca. 4 obliquely transverse veins; style persistent, 1–2 mm long. Seeds several, ca. 2 mm long, obliquely corrugate.
Phenology: Flowering April to June; fruiting late May to September.
Distribution: China, SE Xizang (Medog Xian) (Figure 8).
Habitat: Dense mixed forest, occurring on slopes; elevation: 1500–4000 m.
Additional specimens examined: СHINA, Tibet, Medog County. Nyingchi, near Hanmi village, 2,227 m, 29°22′01.8″″N 95°07′05.2″E, 02 July 2024, J. Zhang, T. Gao, Y.Y. Ling CH2024-22-1 (NS! barcode NS0033870, PE)!; Duoxiong River, 2477 m, 29°24′10.1″N 95°05′16.3″E, 02 July 2024, iidem SP-7-2-1 (NS! barcode NS0033869, PE)!; Nyingchi, near Rizhalu village 2,365 m, 29°23′05.7″N 95°06′07.4″E, 02 July 2024, iidem SP-7-2-2 (NS! barcode NS0033868, PE)!. INDIA, Arunachal Pradesh. Lalung, Pachakshiri [2,438 m], 6 May 1938, F. Ludlow & G. Sherriff 3,714 (BM! barcode BM000078370); 1,829–2,134 m, 28°21′N 96°37″E, 20 July 1928, F. Kingdon Ward 8473 (K! barcodes K004189054 (sheet 1/2) and K004189055 (sheet 2/2)); 2,118 m, 27°34′21.6″N, 93°55′22.7″E, 04 April 2009, A.A. Mao 19143 (ARUN)!; 1,500 – 2,400 m, 28°46′57.1″N 95°11′33.8″E, 21 July 2010, Pathak & Bhaumik 72901 (CAL)!; 2,370–4,000 m, 28°46′59.9″N 95°16′51.6″ E, 11 September 2011, Pathak & Gopal Krishna 54264 (CAL)!.
Preliminary conservation status: Population sizes are likely decreasing with a small number of individuals, with an estimated EOO of 7,633.09 km^2^ and AOO of 44 km^2^. Thus, B. deltophylla was assessed as vulnerable (VU B1a+B2a,b(iii,iv)) at the global level.
- Beesia calthifolia (Maxim. ex Oliv.) Balf.f. & W.W.Sm. 1924, Notes Roy. Bot. Gard. Edinburgh, 14: 110; Ulbr. 1929, Notizbl. Bot. Gart. Berlin-Dahlem 10: 872, isonym; Li and Tamura, 2001, Fl. China 6: 142; Kress et al., 2003, Contr. U.S. Natl. Herb. 45 (Checkl. trees, shrubs, herbs, climbers Myanmar): 333. ≡ Cimicifuga calthifolia Maxim. ex Oliv. 1888, Hooker’s Icon. Pl. 18: pl. 1746 (as calthaefolia). (Figures 9B, E, H, 11).
Beesia calthifolia. (A) General view. (B) Edge of the leaf. (C) Flowers. (D) Sepals. (E) Stamen. (F) Follicle.
Type: CHINA. Szechwan, Mount Omei, 4500 ft, E. Faber 624 (lectotype, K! barcode K004189049, selected here by Erst, Borosova and Tatanov). Syntypes: ibidem, 6,000 ft, idem 625 (var. minor) (K! barcode K004189053, LE! barcode LE01013623, NY! barcode NY00233186 (left-hand fragments on the sheet)); ibidem, summit in ravine, idem 626 (var. minor) (K! barcode K004189052, NY! barcode NY00233186 (right-hand plant on the sheet)); China borealis, prov. Kansu orientali, fl. Dshombunou, prope montem Tschagola, 10 July 1885, G.N. Potanin (LE! barcode LE01013622); China borealis, prov. Kansu orientali, in monte Itschu-schan, 15 July 1885, idem (LE! barcodes LE01013624 and LE01320481); Kansu orientali, 1885, recd. 4 Sept. 1890, idem (K! barcode K004189051, P! barcode P00200370).
= Beesia cordata Balf.f. & W.W.Sm. 1915, Notes Roy. Bot. Gard. Edinburgh 9: 63. – Lectotype: West China. Yunnan, open moist pasture on the margins of thickets on the Kari Pass, Mekong-Yangtze divide, Lat. 27°40″N, Alt. 9,000–10,000 ft, Aug. 1914, G. Forrest 12955 (E! barcode E01150776, selected here by Erst, Borosova and Tatanov). Syntype: Northern Burma [Myanmar], Feng-shui-ling Camp near the Burma-China frontier, among undergrowth of rain forest in deep shade, wet clayey soil, 9,000 ft, fls. white, 9 June 1914, F. Kingdon Ward 1660 (E! barcodes E00078798 (sheet 1/2) and E00078799 (sheet 2/2)).
= Beesia elongata Hand.-Mazz. 1922, Anz. Akad. Wiss. Wien, Math.-Naturwiss. Kl. 59: 245. – Lectotype: China, Yünnan, prope fines Tibeto-Birmanicas inter fluvios Lu-djiang (Salween) et Djiou-djiang (Irrawadi or. Sup.), in jugi Tschiangschel, 27°52″N, 5 July 1916, H.F. Handel-Mazzetti 9156 (WU! barcode WU0033932, selected here by Erst, Borosova and Tatanov; isolectotype, E! barcode E00064955).
– Cimicifuga calthifolia var. minor Maxim. ex Oliv. 1888, Hooker’s Icon. Pl. 18: pl. 1746, nom. inval., without description.
Description: Perennial herbs. Rhizome up to 10 × 0.3–0.5 cm, yellow or light brown. Scapes more than 14 cm at anthesis (– 58 cm at fruiting) × 1–2 mm, basally glabrous, apically densely pubescent with unicellular lageniform trichomes. Basal leaves 2–4; petiole (5.5–)10–26 cm × 0.5–1.5 mm, glabrous, dilated into membranaceous sheath toward base; leaf blade reniform, orbicular-ovate, (1.5–)4.5–9.5 × (1.8–)5.5–16 cm, both surfaces glabrous, rarely pubescent abaxially on veins, light green with inconspicuous veins, dull, base deeply cordate, margin dense with 40–50 teeth on each side, teeth widely crenate with long mucro at apex, sometimes off-center, apex rounded or shortly acuminate. Lower cauline leaves present, same as basal leaves, but smaller in size, upper cauline leaves 1 or 2, lanceolate. Inflorescence cymose, 5.5–9.5 × 1.5–2.5 cm; bracts usually subulate, sometimes lanceolate, rarely spatulate, 1–1.5 mm long, glabrous. Flowers 0.8–1.1 cm in diam.; pedicels 5–10 mm long, densely spreading pubescent. Sepals white or pinkish, narrowly ovate or elliptic, 4–6 × 1.5–2 mm, glabrous, apex acute or rounded. Stamens 3–3.1 mm long, shorter than sepals; anthers 0.3 mm long. Follicle green, 1.1–1.7 cm long, flat, lanceolate-linear, middle part curved, lower part 3–4 mm wide, sparsely pubescent near base, otherwise glabrous, with ca. 8 obliquely transverse veins; style persistent, 1–2 mm. Seeds several, ca. 2.5 mm long, obliquely corrugate.
Phenology: Flowering March to July; fruiting late April to September.
Distribution: China (S. Gansu, N. Guangxi, Guizhou, W. Hubei, W. Hunan, S. Shanxi, Sichuan, N.W. Yunnan); Myanmar (North) (Figure 8).
Habitat: Rainforest and open woodland, occurring on wooded rocky slopes, on river banks, moist slopes along streams, in deep shade amongst undergrowth; elevation: 1,400–4,425 m.
Additional specimens examined. CHINA. [Chongqing] Cheng-kou, 1,450 m, 12 April, 1958, T.L. Tai 102623 (E! barcode E00064958); ibidem, 1,430 m, 26 May 1958, idem 100630 (E! barcode E00064959). Guizhou. Chire-kang-Viheon, May 1908, Cavalerie 2949 (K! barcode K004189038, P! barcodes P00200356 (sheet 1/3), P00200357 (sheet 2/3) and P00200358 (sheet 3/3)). Hupeh [Hubei]. 1885–1888, A. Henry 5957 (BM! barcode BM015177131, E! barcode E01150770, LE! barcode LE01320483, P! barcode P00840973, US! barcode US03558171); ibidem, 1885–1888, idem 5790 (BM! barcode BM015177132, E! barcode E00064956, US! barcode US03558172); ibidem, 1885–1888, idem 5790B (BM! barcode BM000078365, P! barcode P00840975, US! barcode US801343); Patung, May 1888, idem 4669 (K! barcode K004189050, LE! barcode LE01320482); Kuei, recd. March 1889, idem 5790A (BM! barcode BM000078364, K! barcode K004189045, US! barcode US03558173); Ichang, 26 Dec. 1889, idem 5432 (P! barcode P00200369); July 1900, E.H. Wilson 1292 (E! barcode E01150768, K! barcode K004189041, LE! barcode LE01320484, NY! barcode NY04991245, P! barcode P00200363); ibidem, July 1907, idem 2322 (K! barcode K004189044). Kansu [Gansu], Yanchang, Gongjiagou, 2,450 m, 15 July 1989, Y.M. Yuan 1052 (BM! barcode BM015177133, E! barcode E01150778). Shanxi. Wutaishan, above Huadianba, N end of Cangshan, 3,300 m, 20 May 1981, Sino-British expedition to Cangshan 918 (E! barcode E01150777, K! barcode K004189040). [Sichuan] Omei [3,353 m], July 1887, E. Faber 668 (E! barcode E01150780); recd. March 1889, A. Henry 7220 (K! barcode K004189045); Tachienlu [Kangding], [2,743–4,115 m] purchased, Dec. 1890, A.E. Pratt 823 (BM! barcode BM000078371, E! barcode E01150769, K! barcode K004189047, LE! barcode LE01320488, P! barcodes P00200371 (sheet 1/4), P00200372 (sheet 2/4), P00200373 (sheet 3/4) and P00200374 (sheet 4/4)); Mt Omei, June 1904, Wilson (P! barcode P00200375); ibidem, idem 4712 (K! barcode K004189043); 3,800 m, 27 May 1914, C. Schneider 1406 (E! barcode E01041927); Dongrergo, 3,900 m, 23 July 1923, H. Smith 3773 (E! barcode E01150782, K! barcode K004189059); Kulu, E of Muli Gomba, 3650–4425 m, June 1928, F. Rock 16473 (E! barcode E00064957, NY! barcode NY04419612, US! barcode US03558165); Sungpan hsien, 10 Aug. 1928, W.P. Fang 4142 (E! barcode E01150771, K! barcode K004189035, US! barcode US03558163); Omei-shan, 18 Aug. 1928, idem 3161 (E! barcode E01150774); ibidem, 14 June 1930, idem 4731 (US! barcode US03558164); ibidem, 13 July 1930, idem 7468 (K! barcode K004189036); ibidem, 15 July 1930, idem 7606 (K! barcode K004189037); Mt Omei, 2,000 m, 15 July 1931, F.T. Wang 23333 (LE! barcode LE01300485); Tahsiangling, 2,400 m, 26 June 1934, H. Smith 10181 (BM! barcode BM015177135); Sikang, Kangting (Tachienly), 3,300 m, 15 July 1934, idem 10468 (BM! barcode BM015177136, E! barcode E00064960, NY! barcode NY04419611); Tien-chuan-hsien, 2,400 m, 25 May 1936, K.L. Chu 2653 (BM! barcode BM015177139); ibidem, 3,200 m, 13 June 1936, idem 2773 (BM! barcode BM000078363, E! barcode E01150773); Pao-hsing-hsien, 25 July 1936, idem 3292 (BM! barcode BM015177137, E! barcode E01150783); ibidem, 3,000 m, 11 Aug. 1936, idem 3527 (BM! barcode BM015177134); ibidem, 3,000 m, 11 Aug. 1936, idem 3530 (BM! barcode BM015177138, E! barcode E01150779); Kangting, 1963, Kuan & Wang 1303 (K! barcode K004189039); Luding Xian, Moxi, Gongga Shan, E side, 3,000–3,100 m, D.E. Boufford et al., 27382 (E! barcode E00647472, NY! barcode NY04419615, P! barcode P02548475); Dujianyan, from Mashanping to Qinglongzui along the Longxi River, 1,400 m, 11 Sept. 1988, D.E. Boufford et al., 24815 (NY! barcode NY04419613); Mianning Xian, Lamagetou Nature reserve, Yele Xiang 2,790–3,440 m, 10 July 2005, D.E. Boufford et al. 32782 (P! barcode P02548474); Tchen-keou-tin, Su-tchuen oriental., 1,400 m, R.P. Farges 64 (K! barcode K004189042, LE! barcode LE01320486, NY! barcode NY04419610, P! barcodes P00200366 (sheet 1/3), P00200367 (sheet 2/3) and P02527942 (sheet 3/3)); Wenchuan, Ngawa Tibetan and Qiang Autonomous Prefecture, Wolong, 2,803 m, 06 July 2024, A.S. Erst, T.V. Erst, D.A. Krivenko, H.W. Peng CH2024-13 (IRK barcode IRK00049786, NS barcodes NS0003871 and NS0003872, PE). Yunnan. San Chan, May 1906, F. Ducloux 133 (E! barcode E01150781); ibidem, May 1906, idem 4722 (P! barcodes P00200364 (sheet 1/2) and P00200365 (sheet 2/2)); ibidem, 12 June 1907, idem 5188 (P! barcodes P00200359 (sheet 1/2) and P00200360 (sheet 2/2)); Dschungdien (Chuntien) et viscum Djitsung, 4,000–4,100 m, 24 Aug. 1915, H.F. Handel-Mazzetti 7759 (WU! barcodes WU033929 (sheet 1/2) and WU033930 (sheet 2/2)); Landsang-djiang (Mekong) et Lu-djiang (Salween), 3,200–3,500 m, 16 June 1916, idem 8917 (WU! barcode WU033931); [3,353–3,658 m] July 1921, G. Forrest 19707 (CAL! barcode CAL0000041281, E! barcode E01150775, K! barcode K004189058, P! barcode P00200361); [3,353–3,658 m] Aug. 1921, idem 19907 (US! barcode US03558169); Ku-long-tchiang, 800 m, 10 Aug. 1921, R.P. Maire (P! barcode P00200362); Mekong-Salwin divide [3,353 m], Sept. 1921, G. Forrest 20390 (E! barcode E01150772, K! barcode K004189057, US! barcode US03558168); Mountains above Tseku and Tsehchung, Mekong-Salween watershed, 3,048–3,353 m, 1923, J.F. Rock 8729 (US! barcode US03558167, NY! barcode NY04419616); Mount Lauchunshan, SW of the Yangtze bend at Shiku, June 1923, idem 9568 (US! barcode US03558166); Diqing, Zhongdian, below Tian Shu lake, above Xiaozhongdian, 3800 m, 13 June 1993, B. Alden et al., 1231 (E! barcode E011507670); Zhongdian, Tianchi lake, 11 June 1994, Ace 143 (K! barcode K004189048); Shangrila Xian, near Tianchi lake, 3,700 m, 24 July 2007, Tibet-Mac Arthur 1216 (US! barcode US00972018); Muli Xian, near Chang-Hai-Zi lake, 3,674 m, 4 Aug. 2007, idem 1779 (US! barcode US00994674); Shangrila Co., Xiao Zhongdian, on the road to Tianchi lake, 3,834 m, 16 June 2009, idem 2357 (US! barcode US00863424); ibidem, 16 June 2009, idem 2343 (US! barcode US00863410); ibidem, 16 June 2009, idem 2309 (US! barcode US00863295). MYANMAR. Northern Burma, below Feng-shui-ling Camp, near the Burma-China frontier [2743 m], June 1914. F*. Kingdon Ward* 1660 (E! barcode E00078799); Upper Burma, divide between Feng-shui-ling and Lai Kam valleys, 9 Sept. 1919, R.J. Farrer 27382 (E! barcode E00078800). See аlso type specimens above.
Preliminary conservation status: Beesia calthifolia is a widespread species in China. The estimated EOO is 922,109.8 km^2^ and the AOO is 124 km^2^. Thus, B. calthifolia was assessed as least concern at the global level.
- Beesia yangii Erst & K.L. Xiang, sp. nov. (Figures 9C, F, I, 12). urn:lsid:ipni.
Beesia yangii. (A) General view. (B) Edge of the leaf. (C) Flowers. (D) Sepals. (E) Stamen. (F) Follicle.
org:names: 77371025-1
Type: CHINA. Sichuan Province, Dujiangyan Mount, Quingcheng, 970 m, 30°54′18.5″N 103°33′13.6″E, 18 April 2024, Erst A.S., Erst T.V., Zhang J. CH2024-5 (holotype, NS barcode NS0000490; isotypes, IRK barcode IRK00058024, LE barcode LE01323001, NS barcodes NS0000491 and NS0000492). Paratypes: CHINA. Sichuan Province, Pengzhou, Tangba village, 1186 m, 31°11′37.1″N 103°56′29.1″E, 27 May 2025, Erst A.S., Erst T.V., Zhou X.P., Zhu D.L. CH2025-22 (IRK barcode IRK00059177, LE barcode LE01323217, NS barcode NS0000700).
Description: Perennial herbs. Rhizome up to 10 × 0.4–1.1 cm, pink or purple. Scapes more than 40(–75) cm at fruiting × 1–2 mm, basally glabrous, apically densely pubescent. Basal leaves 2–4; petiole (5.5)10–26 cm × 1–2 mm, glabrous, dilated into membranaceous sheath toward base; leaf blade cordate, (1.9–)4.5–13.5 × (3–)5.5–12 cm, both surfaces glabrous, abaxially pale green, adaxially dark-green with light-green or white veins, glossy, base cordate, margin dense with 13–50 teeth on each side, teeth narrowly triangular with long sharp mucro, apex acuminate to acute. Lower cauline leaves same as basal leaves but smaller in size, upper cauline leaves 1 or 2, lanceolate. Inflorescence cymose, 5.5–9.5 × 1.5–2.5 cm; bracts usually subulate, sometimes lanceolate, rarely spatulate, 1–1.5 mm long, glabrous. Pedicels 5–10 mm long, densely spreading pubescent. Sepals white or white-green, narrowly elliptic, 5–9 × 2–4 mm, glabrous, apex acute, or rounded. Stamens 0.4–0.8 mm long, equal to or mostly shorter than sepals; anthers 0.3 mm long. Follicle (0.7–)1.1–1.7(–1.8) cm long, flat, lanceolate-linear, middle part curved, lower part 3–4 mm wide, mostly glabrous, sparsely pubescent near base, with ca. 8 obliquely transverse veins; style persistent, 1–2 mm. Seeds several, ca. 2.5 mm long, obliquely corrugate.
Affinity: The new species is sister to B. calthifolia and B. deltophylla, according to the results of molecular phylogenetic analysis (Figure 4). Beesia yangii is morphologically similar to B. calthifolia and B. deltophylla (Figure 9) in having simple long petiolate basal undivided leaves with cordate or cordate-triangular to reniform blades, crenulate-mucronate leaf margins, paniculate-cymose inflorescences, actinomorphic flowers with sepals, petals lacking, one follicle per flower, and thickened underground creeping rhizomes that produce new aerial shoots. The differences among the three species are presented in Table 1. The new species differs from related species by bearing a cordate basal leaf blade, this with reliably toothed margins, the teeth narrowly triangular with a reliably long sharp mucro, and narrow elliptic, white, or white-green sepals. The sepals are shorter than or equal to the stamens. Additionally, all three species have different distribution patterns (Figure 8).
Phenology: Flowering March to early April; fruiting late April to May.
Distribution: Endemic to Dujiangyan Mount, Sichuan Province, China (Figure 8).
Habitat: Forests, occurring on slopes, elevation: 900–1,700 m.
Etymology: The specific epithet ‘yangii’ of the new species is given in honor of the famous researcher of representatives of the Ranunculaceae family in China: Qin-Er Yang.
Preliminary conservation status: The newly described species is so far known only from two isolated locations; the estimated AOO is 8 km^2^. Thus, B. yangii was assessed as critically endangered (CR B2 a, b (ii, iii, iv, v)) at the global level.
Identification key to species of Beesia
4.3
- Basal leaf blade reniform, light green, with inconspicuous veins, a dull surface, apex rounded or acuminate, edge of basal leaves toothed, teeth widely crenate with long mucro at apex, sometimes off-center; rhizome yellow or light brown; sepals shorter than stamens in open flowers …………………………………………….B. calthifolia
- Basal leaf blade cordate or cordate-triangular, dark green, with lighter green veins, glossy surface, apex acuminate, edge of basal leaves toothed or crenately lobed, teeth narrowly triangular with long sharp mucro or each lobe with mucronate apex, some lobes doubly lobed; rhizome pink or purple; sepals longer than or equal to stamens in open flowers …………….……………………2
- Basal leaf blade cordate-triangular, edge of basal leaves crenately lobed, teeth in each lobe with mucronate apex, some lobes doubly lobed; sepals wide elliptic, white or pink…..…. B. deltophylla
- Basal leaf blade cordate, edge of basal leaves toothed, teeth narrowly triangular with long sharp mucro; sepals narrowly elliptic, white or white green ………………………….…………… B. yangii
Conclusion
5
The integrative taxonomic approach applied in this study using different types of data, including numerous molecular markers together with numerical and qualitative morphological metadata, identified three taxa, including the new cryptic species B. yangii. These data demonstrate that the new taxon is distinct according to comprehensive data showing that it is morphologically and cytogenetically unique, as well as monophyletic. Broader application of integrative approaches may continue to yield important new discoveries in both systematic biology and conservation of biodiversity.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agapow P. M. Bininda-Emonds O. R. P. Crandall K. A. Gittleman J. L. Mace G. M. Marshall J. C. . (2004). The impact of species concept on biodiversity studies. Q. Rev. Biol. 79, 161–179. doi: 10.1086/383542, PMID: 15232950 · doi ↗ · pubmed ↗
- 2Aköz G. Nordborg M. (2019). The Aquilegia genome reveals a hybrid origin of core eudicots. Genome Biol. 20, 256. doi: 10.1186/s 13059-019-1888-8, PMID: 31779695 PMC 6883705 · doi ↗ · pubmed ↗
- 3Ali M. A. Gyulai G. Hidvegi N. Kerti B. Al Hemaid F. M. Pandey A. K. . (2014). The changing epitome of species identification – DNA barcoding. Saudi J. Biol. Sci. 21, 204–231. doi: 10.1016/j.sjbs.2014.03.003, PMID: 24955007 PMC 4061418 · doi ↗ · pubmed ↗
- 4Altınordu F. Peruzzi L. Yu Y. He X. (2016). A tool for the analysis of chromosomes: Karyo Type. Taxon 65, 586–592. doi: 10.12705/653.9 · doi ↗
- 5Audigier V. Husson F. Josse J. (2017). MIMCA: multiple imputation for categorical variables with multiple correspondence analysis. Stat Computing 27, 501–518. doi: 10.1007/s 11222-016-9635-4 · doi ↗
- 6Balfour B. Smith W. W. (1915). A new genus of Ranunculaceae from Burma and Yunnan. Notes R. Botanic Garden Edinburgh 9, 63–64. doi: 10.24823/nrbge.1915.2262 · doi ↗
- 7Balfour I. B. Smith W. W. (1924). “ Beesia calthaefolia, Balf. f. et W. W. Sm,” in Plantae Chinenses Forrestianae. Catalogue of the plants (excluding Rhododendron) collected by George Forrest during his fifth exploration of Yunnan and Eastern Tibet in the years 1921–1922. (Edinburgh: Royal Botanic Garden). vol. 14, 110.
- 8Bhaumik M. Krishna G. (2024). Contribution to the flora of dihang dibang biosphere reserve, India-I. Taiwania 69, 412–419. doi: 10.6165/tai.2024.69.412 · doi ↗