Description on the prevalence of Proteus mirabilis through an integrated sampling framework for health, food, and environment in Northeast India and an integrative review with reference to one health context
Goutam Chowdhury, Tapan Majumdar, Dilem Modi, Karma G. Dolma, Suranjana Chaliha Hazarika, Pallab Sarmah, Samaresh Das, Asish K. Mukhopadhyay, Anup Kumar Ojha, Madhuchhanda Das, Thandavarayan Ramamurthy

TL;DR
This study explores Proteus mirabilis prevalence in health, food, and environment in Northeast India, highlighting its antimicrobial resistance and virulence genes.
Contribution
The study reports the first detection of SPI-1 in P. mirabilis and provides insights into its virulence and resistance patterns across multiple sources.
Findings
P. mirabilis showed high resistance to antibiotics like doxycycline and tetracycline in diarrheal and food isolates.
Whole-genome sequencing identified SPI-1 and other virulence genes in P. mirabilis linked to diarrhea.
Environmental samples had a higher prevalence of P. mirabilis compared to diarrheal cases.
Abstract
Foodborne infections caused by different pathogens are a perennial public health problem in India. An uncommon enteric pathogen Proteus mirabilis has been frequently isolated from the hospitalized diarrheal cases and different food and environmental samples collected from four states in Northeast India. This study was aimed at characterizing P. mirabilis isolates to show its etiological importance in public health and also present a review indicating its global prevalence of antimicrobial resistance and detection virulence genes reported from different sources. In this study, we have screened 6,298 diarrheal stools from hospitalized patients, 12,305 market foods, 4,270 state-specific foods, and 2,130 environmental samples. P. mirabilis was isolated and identified by routine microbiological methods. Representative isolates were examined for antimicrobial susceptibility, putative…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
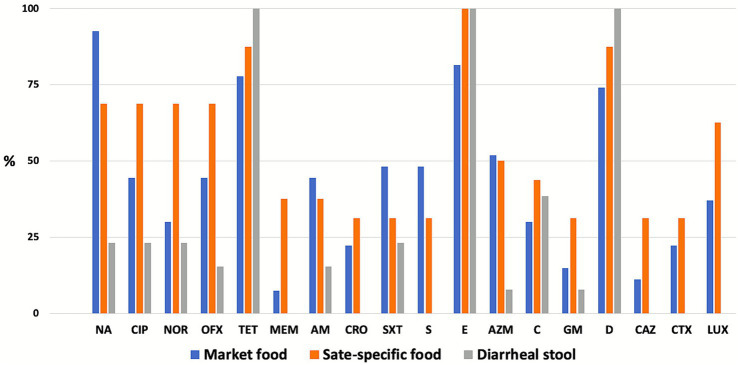 Figure 1
Figure 1| Type of sample | State/percent positive (No positive/total sample) | Total % | |||
|---|---|---|---|---|---|
| Arunachal Pradesh | Assam | Sikkim | Tripura | ||
| Diarrheal stool | 1.9 (11/569) | (0/1051) | (0/3747) | 1.3 (12/931) | 0.4 (23/6298) |
| Market foods | 3.2 (39/1206) | 0.5 (20/4199) | 1.3 (56/4443) | 5.6 (139/2457) | 2.1 (254/12,305) |
| State-Specific Foods | 0.8 (6/712) | 3.9 (74/1894) | 0.8 (7/875) | 0.8 (6/789) | 2.2 (93/4270) |
| Environmental samples | 4.7 (44/926) | (0/178) | 0.0 (0/331) | 3.7 (26/695) | 3.2 (70/2130) |
| Source | No. of samples tested | No of samples positive for |
|---|---|---|
| Environment (cutting surfaces, hand-washing from food-handlers, utensils) | 1,606 | 62 (3.9) |
| Stored water | 524 | 8 (1.5) |
| Total | 2,130 | 70 (3.2) |
| Source | No. of samples tested | No of samples positive for |
|---|---|---|
| Ckung | 69 | 3 (4.3) |
| Khar, Outenga | 16 | |
| Dough and batter | 316 | 4 (1.3) |
| Fermented/processed/preserved | 420 | 3 (0.7) |
| Fruits, vegetables and salads | 2,101 | 20 (0.9) |
| Milk products and sweets | 302 | 4 (1.3) |
| Processed milk products and sweets | 873 | 4 (0.4) |
| Non-vegetarian foods | 1,080 | 27 (2.5) |
| Raw/dried meat | 2,309 | 111 (4.8) |
| Raw/dry fish | 2018 | 43 (2.1) |
| Refrigerated foods | 161 | 6 (3.7) |
| Rice, flour, pulses | 1,467 | 19 (1.3) |
| Kinema | 40 | |
| Spices | 27 | 1 (3.7) |
| Bangoi, godak | 9 | |
| Vegetables | 1,097 | 9 (0.8) |
| Total | 12,305 | 254 (2.1) |
| Source | No. of samples tested | No of samples positive for |
|---|---|---|
| Alcoholic Brew | 23 | |
| Bamboo Shoots | 260 | 4 (1.5) |
| Beer | 32 | |
| Cereals | 716 | 5 (0.7) |
| Fish | 936 | 42 (5.9) |
| Insects | 135 | 8 (5.9) |
| Meat | 147 | 4 (2.7) |
| Milk Products | 122 | |
| Small animals | 185 | 13 (7.0) |
| Soyabean | 128 | 1 (0.8) |
| Vegetables | 943 | 11 (1.2) |
| Wine | 643 | 5 (0.8) |
| Total | 4,270 | 93 (2.2) |
| Sample ID/Source | Center | Virulence gene | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
| ||
| ASM/KMR/RAWFISH/4142 | Assam | |||||||||||
| ASM/KMR/DRYFISH/4141 | Assam | |||||||||||
| ASM/KMR/COOKEDMEAT/4153 | Assam | |||||||||||
| ASM/KMR/COOKEDFOOD/4123 | Assam | |||||||||||
| ASM/KMR/ENVIRONMENT/2961 | Assam | |||||||||||
| ASM/KMR/COOKEDMEAT/3901 | Assam | |||||||||||
| ASM/KMR/RAWFISH/2964 | Assam | |||||||||||
| ASM/KMR/COOKEDMEAT/4355 | Assam | |||||||||||
| ASM/KMR/VEGETABLE/3221 | Assam | |||||||||||
| AR/BPGH/MS/2090 | Arunachal Pradesh | |||||||||||
| ASM/KAM/SSF/S7 | Assam | |||||||||||
| ASM/KAM/SSF/S6 | Assam | |||||||||||
| ASM/KAM/SSF/S39 | Assam | |||||||||||
| ASM/DIB/RAWMEAT/921 | Assam | |||||||||||
| SKM/SSK/RAWMEAT/2517 | Sikkim | |||||||||||
| Source | Virulence | Major virulence encoding gene | Reference |
|---|---|---|---|
| Chicken | Siderophore, proteases, fimbriae, hemolysins |
| |
| Fermented foods | Hemolysin, urease |
| |
| Diarrheal patients | Type IV secretion system, hemolysins, metalloprotease |
|
|
| Chicken | Aggregative adherence in HEp-2, biofilm formation, cytotoxicity in Vero cells |
|
|
| Broiler farms | Siderophore, proteases, fimbriae |
| |
| Animals with diarrhea | Biofilm production |
| |
| Pig farm | Fimbriae, siderophore, siderophore |
|
|
| Chicken | Fimbriae, proteases |
|
|
| ICMR FoodNet ID | Center | Resistance genes | Virulence genes |
|---|---|---|---|
| TRP/DHL/HUMAN/1/1 | Tripura | Chloramphenicol: | TTSS (SPI-1 encode): |
| TRP/DHL/HUMAN/2/1 | Tripura | Chloramphenicol: | TTSS (SPI-1 encode): |
| TRP/DHL/HUMAN/4/1 | Tripura | Chloramphenicol: | TTSS (SPI-1 encode): |
| TRP/DHL/HUMAN/5/1 | Tripura | Chloramphenicol: | TTSS (SPI-1 encode): |
| TRP/DHL/HUMAN/6/1 | Tripura | Chloramphenicol: | TTSS (SPI-1 encode): |
| ASM/KAM/VEGETABLE/ 1,204 | Assam | Chloramphenicol: | TTSS (SPI-1 encode): |
| SKM/SSK/RAWMEAT/4248 | Sikkim | Chloramphenicol: | TTSS (SPI-1 encode): |
| ASM/KAM/SSF/S39 | Assam | Chloramphenicol: | TTSS (SPI-1 encode): |
| Source | Antimicrobial resistance | Predominant AMR encoding gene | Reference |
|---|---|---|---|
| Meat and aquatic products | Trimethoprim-sulfamethoxazole, streptomycin, chloramphenicol, ampicillin, nalidixic acid, norfloxacin, florfenicol, gentamicin, fosfomycin | ICE |
|
| Chicken | Ampicillin, ceftiofur, amoxicillin-clavulanic acid, cephalotin, cefoxitin, ceftazidime, ceftriaxone, cefotaxime, cefepime, nalidixic acid, norfloxacin, enrofloxacin, ciprofloxacin, trimethoprim-sulfamethoxazole, aztreonam, chloramphenicol, gentamicin |
| |
| Broiler farms | Chloramphenicol, ciprofloxacin, trimethoprim-sulfamethoxazole, imipenem, and meropenem |
| |
| Food-producing animals | Ampicillin, amoxicillin-clavulanic acid, cefotaxime, ceftriaxone, chloramphenicol, florfenicol, nalidixic acid, ciprofloxacin, streptomycin, spectinomycin, apramycin, doxycycline, trimethoprim-sulfamethoxazole |
| |
| Pig farms | tetracyclines, macrolides, sulfonamides, |
| |
| Chicken | Beta-lactams, quinolones, sulfonamides |
| |
| Clinical and foods | Tetracycline, minocycline, doxycycline, sulfisoxazole, streptomycin, gentamicin, colistin, and polymyxin B | SGI1 and |
|
| Farm animals | sulfamethoxazole, ampicillin, florfenicol, ciprofloxacin, amoxycillin/clavulanic acid, cefatriaxone, chloramphenicol, levofloxacin, gentamicin, Fosfomycin, tetracycline | SXT/R391 ICEs |
|
| Swine | trimethoprim-sulfamethoxazole, rifampicin, tetracycline, chloramphenicol, fosfomycin | ICE |
|
| Chicken and swine | Amoxicillin-clavulanate, ampicillin, aztreonam, ceftazidime, chloramphenicol, ciprofloxacin, ceftriaxone, cefotaxime, florfenicol, fosfomycin, nalidixic acid, norfloxacin, spectinomycin; streptomycin, trimethoprim-sulfamethoxazole |
| |
| Chicken | Spectinomycin, fosfomycin, cephalosporins, aztreonam, erythromycin, kanamycin, and gentamicin | ICE |
|
| Food animals | ceftazidime, cefotaxime, ceftriaxone |
| |
| Chicken | Chloramphenicol, trimethoprim-sulfamethoxazole, streptomycin, spectinomycin, cephalosporins, aztreonam, fosfomycin, kanamycin, and gentamicin | ICE |
|
| Chicken | Chloramphenicol and sulfonamides |
|
- —Indian Council of Medical Research, New Delhi
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEscherichia coli research studies · Salmonella and Campylobacter epidemiology · Veterinary medicine and infectious diseases
Introduction
1
The genus Proteus includes several species, among which Proteus mirabilis is clinically significant. This opportunistic pathogen is well-known to cause urinary tract infections (UTIs) in humans (Facciolà et al., 2022). Furthermore, several recent reports indicate the potential association between P. mirabilis and diarrheal disease (Müller, 1989; Kharat et al., 2023; Kiiru et al., 2024). P. mirabilis is not typically regarded as a major foodborne pathogen compared to other pathogenic species such as Salmonella or Campylobacter, but it does have important public health implications, particularly in certain populations and settings, as it can also cause gastrointestinal infections. In the context of foodborne illness, P. mirabilis can contaminate improperly handled or undercooked food, especially meat, poultry, and dairy products. Food-producing animals (Guo et al., 2019; Yu et al., 2021; El-Saeed et al., 2024; Ramatla et al., 2024) and fermented foods (Keisam et al., 2019) serve as common sources for the transmission of P. mirabilis. Foodborne outbreaks due to P. mirabilis contamination have also been reported (Cooper et al., 1971; Gong et al., 2019). Infection caused by P. mirabilis in food poisoning cases has several clinical symptoms, including nausea, abdominal pain, and severe diarrhea (Shi et al., 2016). The other important concerns regarding P. mirabilis are its potential to develop resistance to multiple antibiotics and its ability to acquire and transfer antibiotic resistance genes, thereby promoting the development of resistant strains. Due to these reasons, public health surveillance and appropriate treatment strategies for P. mirabilis-associated infections are vital. Several findings indicate the existence of multidrug resistance (MDR) in P. mirabilis isolated from the patients and several food sources (Guo et al., 2019; Facciolà et al., 2022; Kiiru et al., 2024). Although many putative virulence factors have been reported in P. mirabilis, the specific pathogenic mechanism causing diarrhea in humans is still unclear. Considering its role in the One Health approach, systematic surveillance of different sources, proper infection control, and prudent use of antibiotics are important in minimizing its impact on public health.
The present study was conducted to characterize P. mirabilis isolated from the diarrheal cases and different food and environmental samples collected from four states in Northeast India. PCR and whole-genome sequence (WGS) analysis were performed to investigate the genetic features of P. mirabilis isolates with reference to their putative virulence factors and antimicrobial resistance genes (ARGs).
Materials and methods
2
Collection and processing of samples
2.1
As a part of ICMR sentinel diarrheal surveillance, the present study was conducted from 2021 to 2024 in four states of Northeast India, namely Sikkim (Gangtok), Assam (Dibrugarh and Guwahati), Arunachal Pradesh (Pasighat), and Tripura (Agartala). Considering distinct types of samples within each category, a stratified sampling method was used. This strategy leads to more precise estimates for each subgroup. Patients with symptoms suggestive of diarrheal infection, including abdominal pain and watery/bloody stool, were enrolled in this study after obtaining informed consent. For microbial testing, we have screened 6,298 diarrheal stools from hospitalized patients. The stool samples/rectal swabs collected in Carry Blair medium were transported to the Microbiology Laboratory for processing. In addition, 12,305 market foods, 4,270 state-specific foods, and 2,130 environmental samples were collected from shops and vendors, as well as ready-to-eat foods from restaurants and street vendors. The food items included in this study were selected based on high consumption rates, increased risk of contamination, and the fact that they are consumed either raw or in a semi-cooked state, and local culinary practices for consumption and preservation patterns. Environmental samples include stored water, swabs from cutting surfaces/utensils, and hand-wash samples from food handlers.
Isolation and phenotypic characterization of Proteus mirabilis
2.2
After collection, the samples were processed within 2–3 h and tested for identification of enteric pathogens using the cultural and molecular identification methods as per standard operating procedures developed for the sentinel diarrheal surveillance (https://www.icmrfoodnet.in/static/assets/files/ICMR_StandardOperatingProcedures.pdf; Accessed on October 29, 2025). Suspected Proteus spp. isolates were identified using VITEK^®^2 GN Compact system (bioMérieux, Marcy-l’Étoile, France). A total of 43 randomly selected P. mirabilis isolates that cover different sources and states were tested for antimicrobial susceptibility (AST) by the Kirby-Bauer disk diffusion method on Mueller-Hinton agar (Difco). The results were interpreted according to the guidelines set by the CLSI (Clinical and Laboratory Standards Institute) (2022). In the AST, commercially available antibiotic discs were used (BD Co., Sparks, MD, USA) that include ampicillin (AM, 10 μg), azithromycin (AZM, 15 μg), cefotaxime (CTX, 30 μg), ceftazidime (CAZ, 30 μg), ceftriaxone (CRO, 30 μg), chloramphenicol (C, 30 μg), ciprofloxacin (CIP, 5 μg), doxycycline (D, 30 μg), erythromycin (E, 15 μg), gentamicin (GM, 10 μg), levofloxacin (LVX, 5 μg) nalidixic acid (NA, 30 μg), norfloxacin (NOR, 10 μg), meropenem (MEM, 10 μg), ofloxacin (OFX, 5 μg), streptomycin (S, 10 μg), tetracycline (TE, 30 μg), and trimethoprim-sulfamethoxazole (SXT, 1.25 and 23.75 μg). Escherichia coli strain ATCC 25922 was used as a quality control strain to validate test accuracy.
PCR-based detection of virulence encoding genes
2.3
Fifteen randomly selected P. mirabilis isolates from different foods were tested for the major virulence genes encoding extracellular metalloprotease (zapA), flagellin (fliC), hemolysin A of Proteus spp. (hmpA), mannose-resistant *Proteus-*like fimbria (mrpA, mrpH), P. mirabilis fimbriae (pmfA), Proteus toxic agglutinin (ptaA), regulator of swarming behavior (rsbA, rsbR), urease enzyme large subunit (ureC, ureG), and uroepithelial cell adhesin fimbriae (uca), using the published primers and PCR conditions (Pathirana et al., 2018; Mirzaei et al., 2019). Overnight cultures of P. mirabilis isolates grown in Luria-Bertani broth (BD Difco, USA) were used in the PCR assay. For each test, a total of 2.5 μl of the washed bacterial cells in sterile phosphate buffer saline solution was used as a DNA template.
Genomic DNA sequencing and data analysis
2.4
Eight different isolates of P. mirabilis identified from humans, vegetables, raw meat, and state-specific foods were included in the WGS. Genomic DNA was prepared using the QIAamp DNA Mini Kit (Qiagen, Hilton, Germany) following the manufacturer’s guidelines. WGS was performed on the HiSeq 2000 (Illumina, San Diego, CA, United States) platform using a paired-end library with an insert size of 150 bp. The presence of ARGs and virulence encoding genes (VEGs) was detected using the ABRicate V1.0.1 tool.
Results and discussion
3
Prevalence of Proteus mirabilis
3.1
Proteus mirabilis is known to cause UTI using several virulence mechanisms. This pathogen has also been associated with diarrhea in humans through the consumption of contaminated foods. P. mirabilis are ubiquitously present in animals, foods, and environment, and are, therefore, important under One Health context. In addition to diarrheal stool samples, this study was designed to test food samples from the market and different sources in the environment. In any pathogen-specific surveillance, such triangulation of data provides evidence not only to link human illness to food contamination but also improves public health surveillance and guides food safety interventions. In this study, the isolation rate of P. mirabilis remained very low (0.4%) in 6298 clinical samples and was detected only in diarrheal cases from Tripura (1.3%) and Arunachal Pradesh (1.9%) (Table 1). Remarkably, of the 12,305 market samples tested, P. mirabilis was found higher in food samples collected from Tripura and Arunachal Pradesh (5.6% and 3.2%, respectively), as compared to the other two states (Table 1). P. mirabilis was present in 74 (3.9%) state-specific foods collected from Assam.
Proteus mirabilis was detected in 3.2% (70/2130) of the environmental samples that include cutting surfaces/utensils, had-wash samples from food handlers, and stored water (Table 2). Reports on the prevalence of this pathogen in such environmental samples were not found in the existing literature. Raw/dried meat (111/12,305) showed slightly higher positivity for P. mirabilis (Table 3). In several studies, the presence of P. mirabilis has been reported more in animal foods (Sanches et al., 2019; Yu et al., 2021; Ma et al., 2022; El-Saeed et al., 2024), aquatic products (Ma et al., 2022), and abattoirs (Bhandare et al., 2010) than in vegetables (Li et al., 2023). Among the state-specific foods, P. mirabilis was detected more in fermented small animals (13/185) (Table 4). As reported in other studies, the prevalence of P. mirabilis seems high among fermented foods of the Northeast regions in India (Keisam et al., 2019; Kumar et al., 2019; Anand Singh et al., 2023).
Virulence encoding genes (VEGs)
3.2
The pathogenicity of P. mirabilis involves several virulence factors (Chakkour et al., 2024). Of the 15 P. mirabilis isolates tested in the PCR assay, 80% of the isolates were positive for pmfA (P. mirabilis fimbriae), and 73% were positive for mrpA/mrpH, ureC, ptaA, hmpA, zapA, fliC, and rsbAR (Table 5). The uca and ureG genes were detected in 67% and 40% of the isolates, respectively. Most of these genes have been reported in P. mirabilis isolated from UTI cases. Several reports indicated that P. mirabilis isolates harboring ireA (siderophore receptors), ptaA, and zapA, ucaA, pmfA, atfA (ambient temperature fimbriae is a type 1 major fimbrial subunit), and mrpA, fliC, hlyA (hemolysin A), and hpmA genes are strongly associated with their pathogenesis (Barbour et al., 2012; Sanches et al., 2019; Li et al., 2022; Ramatla et al., 2024).
In several studies, the presence of virulence factors and VEGs has been reported in P. mirabilis isolated from diarrheal patients, fermented foods, food animals, and contaminated foods. ureC, hamA, zapA, and rsmA (regulation of swarming motility) were identified in diarrheal cases (Table 6). The virB gene associated with a type IV secretion system was reported to play a role in causing diarrhea in humans (Shi et al., 2016; Gong et al., 2019). P. mirabilis isolated from food poisoning cases demonstrated in vitro and in vivo gastrointestinal pathogenicity, including adherence and invasiveness (Shi et al., 2016). Furthermore, biofilm production was notably associated with the expression of ureC, zapA, rsmA, hmpA, mrpA, atfA, and pmfA genes and was also shown to induce diarrhea in mice (Sun et al., 2020).
P. mirabilis is primarily known for its role in UTI and wound infections, but it has also been implicated in gastrointestinal infections. Unlike Salmonella and Shigella, the diarrheal pathogenesis of P. mirabilis is poorly established. The pathogenesis of diarrheal disease caused by P. mirabilis is multifactorial and includes mucosal lining adherence and colonization, biofilm formation in the gut (Palusiak, 2022), production of urease in causing intestinal dysbiosis (Grahl et al., 2023), secretion of hemolysins (Elhoshi et al., 2023), and several other secreted factors leading to inflammation and diarrhea (Shi et al., 2016).
Using WGS, different VEGs have been identified in P. mirabilis. However, VEGs specific for the diarrheal infection, which are present in the other enteric pathogens, have not been reported in P. mirabilis. In the WGS analysis, we identified potential VEGs associated with diarrheal infections (Table 7). SPI-1 (Salmonella Pathogenicity Island 1) is a gene cluster primarily associated with Salmonella species that enables them to invade host epithelial cells. SPI-1 encodes a Type III Secretion System (T3SS) which injects effector proteins into host cells, triggering rearrangements of the host cell’s cytoskeleton and allowing bacterial internalization. The majority of Salmonella virulence genes are located within these distinct genomic regions of SPIs, which have been acquired by independent horizontal transfer events (Marcus et al., 2000; Galán, 2001). SPI-1 is composed of 39 genes, including inv, hil, prg, org, spa, and sip/ssp genes, and plays a key role in Salmonella pathogenesis (Boyd et al., 1997). The majority of the known invasion genes are located between centisomes 62 and 64 of the chromosome, and SPI-1 is located at centisome 63 (Hansen-Wester and Hensel, 2001). Several SPI-1 gene components including invC, invG, spaP, and SpaQ have been identified in P. mirabilis isolates, which is a significant finding. To the best of our knowledge, this is the first report of SPI-1 in P. mirabilis isolates. Our work is in progress to map the entire SPI-1 gene cluster of P. mirabilis using the WGS data.
A growing body of evidence suggests that these putative VEGs have been identified in several Gram-negative pathogens and are linked with diarrheal infection. In Salmonella enterica, the surface-presenting antigen inv-spa complex has been reported as a potential virulence factor (Boyd et al., 1997). The adherence*-*linked pap (pappyelonephritis-associated pilus) is important in P fimbriae assembly in Escherichia coli and also responsible for causing diarrhea in young animals (Ngeleka et al., 2019; Renzhammer et al., 2020). Lipid A 4′-kinase encoded in the gene ipx acts as an endotoxin, which is expressed in many Gram-negative infections, including septic shock (Garrett et al., 1997; Raetz and Whitfield, 2002). ipx and the lipid-A biosynthesis gene htrB are considered potential virulence factors in Salmonella spp. (Jones et al., 1997) and Campylobacter spp., (Stoakes et al., 2023), respectively. In Campylobacter spp., Salmonella spp., and E. coli, the genes cheR, cheZ and motA regulate adhesion, motility ability, invasion and biofilm formation (Vilas Boas et al., 2024; Koolman et al., 2015). Identification of these VEGs underscores the pathogenic potential of P. mirabilis. However, the role of virulence genes in causing diarrheal infection needs further investigation. Several WGS analyses of P. mirabilis have been made using isolates from humans (Ghiglione et al., 2025; Zamudio et al., 2025), foods (Ma et al., 2022; Tan et al., 2025), and animals/birds (Khudeir et al., 2023; Ma et al., 2023; Su et al., 2025). Some of these studies compared genomic consistency, VEGs, AMR genes, and lineage analysis.
Antimicrobial resistance in Proteus mirabilis
3.3
All the tested P. mirabilis isolates from diarrheal stool samples showed resistance to doxycycline, erythromycin, and tetracycline (Figure 1). However, these isolates were susceptible to tetracycline, ceftazidime, ceftriaxone, and levofloxacin. From market samples, majority of the P. mirabilis isolates are resistant to nalidixic acid (92.6%), erythromycin (81.5%), tetracycline (77.8%), doxycycline (77.8%). Only two isolates showed resistance to meropenem (7.4%; Figure 1). State-specific foods, mostly locally prepared fermented foods, showed a higher prevalence of AMR P. mirabilis than the market food isolates. Higher resistance (>50%) has been seen with quinolone/fluroquinolones, erythromycin, tetracycline, and doxycycline. Meropenem resistance has also been recorded in isolates from market foods and state-specific foods (7.4% and 37.5%, respectively).
Antimicrobial resistance patterns of P. mirabilis isolated from different sources (n = 43). AM, ampicillin; AZM, azithromycin; CTX, cefotaxime; CAZ, ceftazidime; CRO, ceftriaxone; C, chloramphenicol; CIP, ciprofloxacin; D, doxycycline; E, erythromycin; GM, gentamicin; LVX, levofloxacin; NA, nalidixic acid; NOR, norfloxacin; MEM, meropenem; OFX, ofloxacin; S, streptomycin; TE, tetracycline; and SXT, trimethoprim-sulfamethoxazole.
The majority of the clinical isolates were multidrug resistant (MDR). High resistance to quinolone/fluroquinolones, tetracycline has been reported in P. mirabilis isolated from chickens (Ramatla et al., 2024; Sarıçam İnce and Akan, 2024), cotrimoxazole (59.32%) from dairy products (Lordejani et al., 2025), doxycycline, tetracycline, sulfamethoxazole, kanamycin, and cephalothin from diarrhetic animals (Sun et al., 2020), carbapenem resistance from clinical sources (Facciolà et al., 2022; ElTaweel et al., 2024).
We have tested five representative P. mirabilis strains in the WGS analysis. This analysis has identified several ARG alleles such as aminoglycoside acetyltransferase (aadA2, aadA5) aminoglycoside phosphotransferase [aac(3′)-lid, aph(3′)-Ia, aph(3′)-Ib, aph(6′)-Ib10, aph(6′)-Id], beta-lactamase (blaTEM-1, blaCTX-M-3, blaOXA0-1), chloramphenicol acetyltransferase (catA1, catA4, catB3, cmlA) dihydrofolate reductase (dfrA1, dfrA17, dfrA32), erythromycin esterase (ereA) florfenicol resistance (floR), and sulfamethoxazole resistance (sul1, sul2, sul3). Two isolates from diarrheal cases and three food isolates harbored catA4, which encodes resistance to chloramphenicol (Table 7).
Several studies indicate that P. mirabilis isolated from diarrheal patients, food animals, and contaminated foods is MDR (Table 8). ARGs identified from animal foods include aad, aac, aph, blaCTX, blaOXA, cat, dfr, and sul. The self-transmissible mobile genetic element SXT/R391 is an integrative and conjugative element (ICE). Interestingly, P. mirabilis isolates from the non-human sources harbored ICEPmiChnHBRJC2, ICEPmiChnSC1111, ICEPmiChnJZ26, ICEPmiChnChSC1905, ICEPmiChnS012, and ICEPmiJpn1 (Table 8). The SXT/R391 ICEs carry the ARGs encoding resistance to chloramphenicol, kanamycin, streptomycin, trimethoprim-sulfamethoxazole, and tetracycline (Li et al., 2016). In addition to ICEs, P. mirabilis can easily acquire and transfer ARGs to other bacterial species through several mobile genetic elements such as plasmids, insertion sequences, and transposons (Girlich et al., 2020). Salmonella genomic island-1 (SGI1) is an integrative mobile element originally identified in the MDR Salmonella Typhimurium DT104 clone (Boyd et al., 2001). SGI1 and Proteus genomic island (PGI) in P. mirabilis have been found to carry several important ARGs (Boyd et al., 2001; Wang et al., 2019). P. mirabilis isolates are mostly susceptible to fluoroquinolones (Wang et al., 2019). Some of the isolates from chicken sources carried plasmid-mediated qnrD, qnrA1, and qnrS1 genes, which are shown to express resistance to fluoroquinolones (Table 8).
The pathogenesis of diarrheal disease caused by P. mirabilis is complex and involves multiple mechanisms, including adherence to intestinal surfaces, urease production, toxin secretion, and induction of inflammation. Although P. mirabilis is not among the most common causes of gastroenteritis, its ability to disrupt gut homeostasis and to express a repertoire of virulence factors makes it a potential contributor to gastrointestinal diseases.
Limitations of this study
4
There are a few limitations in this study that could be addressed in future research. Although we collected a large number of samples from different sources, statistical analysis was not done because of the high variability in the prevalence of P. mirabilis. WGS data are limited to a few isolates, and large-scale analyses must include identification of sequence types (ST) through core genome multilocus sequence typing (MLST), identification of SPI-1 regulatory genes, and pangenome analysis. Our findings suggest the presence of a Salmonella-like type III secretion module in the genomes of P. mirabilis. However, this study does not present the structural organization of these genes or the genomic island context that would firmly establish an SPI-1-like cluster in P. mirabilis. No in vivo study has been conducted to confirm the functional or virulence expression associated with gastrointestinal pathogenicity.
Contribution and implications
5
This study demonstrates the prevalence of P. mirabilis in diarrheal patients and in food and environmental samples collected in NE India. Market- and state-specific food items showed greater positivity toward this pathogen. The identified P. mirabilis isolates from several sources harbored virulence genes not only responsible for UTI but also capable of causing diarrheal infection. Overall, our findings emphasized the need to include P. mirabilis in routine surveillance as an indicator of food quality. Effective control and prevention strategies for P. mirabilis-associated infections rely on a combination of hygiene practices, antibiotic stewardship, and monitoring in high-risk settings like healthcare facilities and food processing plants. Regular surveillance of healthcare-associated infections can help identify contamination load and outbreaks of P. mirabilis.
Conclusion
6
While P. mirabilis may not be as frequently implicated in foodborne illness as other pathogens, it remains a public health concern due to its potential to cause opportunistic infections. Its role in healthcare-associated infections and the growing issue of antimicrobial resistance. The prevalence of P. mirabilis isolated from several foods from Northeast India is a major concern. Reports from other studies also support the potential of P. mirabilis as a foodborne pathogen. P. mirabilis isolated in this study harbored several VEGs and ARGs and hence may have an association with food contamination/diarrheal disease. However, further detailed studies are warranted to strengthen the hypothesis with well-designed in vitro and in vivo investigations. To control infections caused by P. mirabilis, several public health measures, such as proper sanitation, adequate surveillance, monitoring, and tracking AMR trends, need to be addressed under the One Health approach. Understanding the routes of transmission of P. mirabilis and adhering to best practices in food safety and infection control are key in mitigating its impact.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Anand Singh T. Nongthombam G. Goksen G. Singh H. B. Rajauria G. Kumar Sarangi P. (2023). Hawaijar-an ethnic vegan fermented soybean food of Manipur, India: a comprehensive review. Food Res. Int. 170:112983. doi: 10.1016/j.foodres.2023.112983, 37316061 · doi ↗ · pubmed ↗
- 2Barbour E. K. Hajj Z. G. Hamadeh S. Shaib H. A. Farran M. T. Araj G. . (2012). Comparison of phenotypic and virulence genes characteristics in human and chicken isolates of Proteus mirabilis. Pathog. Glob. Health. 106, 352–357. doi: 10.1179/2047773212 Y.0000000042, 23182140 PMC 4005134 · doi ↗ · pubmed ↗
- 3Bhandare S. Paturkar A. M. Waskar V. S. Zende R. J. (2010). Prevalence of microorganisms of hygienic interest in an organized abattoir in Mumbai, India. J. Infect. Dev. Ctries. 4, 454–458. doi: 10.3855/jidc.998, 20818094 · doi ↗ · pubmed ↗
- 4Boyd E. F. Li J. Ochman H. Selander R. K. (1997). Comparative genetics of the inv-spa invasion gene complex of Salmonella enterica. J. Bacteriol. 179, 1985–1991. doi: 10.1128/jb.179.6.1985-1991.1997, 9068645 PMC 178923 · doi ↗ · pubmed ↗
- 5Boyd D. Peters G. A. Cloeckaert A. Boumedine K. S. Chaslus-Dancla E. Imberechts H. . (2001). Complete nucleotide sequence of a 43-kilobase genomic island associated with the multidrug resistance region of Salmonella enterica serovar typhimurium DT 104 and its identification in phage type DT 120 and serovar Agona. J. Bacteriol. 183, 5725–5732. doi: 10.1128/JB.183.19.5725-5732.2001, 11544236 PMC 95465 · doi ↗ · pubmed ↗
- 6Chakkour M. Hammoud Z. Farhat S. El Roz A. Ezzeddine Z. Ghssein G. (2024). Overview of Proteus mirabilis pathogenicity and virulence. Insights into the role of metals. Front. Microbiol. 15:1383618. doi: 10.3389/fmicb.2024.1383618, 38646633 PMC 11026637 · doi ↗ · pubmed ↗
- 7Chen X. Lei C. W. Liu S. Y. Li T. Y. Chen Y. Wang Y. T. . (2022). Characterisation of novel Tn 7-derivatives and Tn 7-like transposon found in Proteus mirabilis of food-producing animal origin in China. J. Glob. Antimicrob. Resist. 28, 233–237. doi: 10.1016/j.jgar.2022.01.012, 35081393 · doi ↗ · pubmed ↗
- 8Chinnam B. K. Nelapati S. Tumati S. R. Bobbadi S. Chaitanya P. V. Bodempudi B. (2021). Detection of β-lactamase-producing Proteus mirabilis strains of animal origin in Andhra Pradesh, India and their genetic diversity. J. Food Prot. 84, 1374–1379. doi: 10.4315/JFP-20-399, 33725112 · doi ↗ · pubmed ↗