Reporting two novel Kluyvera species, Kluyvera huaxiensis and Kluyvera chengduensis, isolated from human sputa
Lina Liu, Yu Feng, Hongxia Wen, Zhiyong Zong

TL;DR
Scientists discovered two new species of bacteria, Kluyvera huaxiensis and Kluyvera chengduensis, from human sputum samples.
Contribution
The paper introduces two novel bacterial species within the genus Kluyvera based on genomic and phenotypic analysis.
Findings
Strains 142053T and 142359T form distinct branches in the Kluyvera clade based on phylogenomic analysis.
Digital DNA-DNA hybridization and average nucleotide identity values are below species delineation thresholds for both strains.
The two strains differ phenotypically in lipase, arginine dihydrolase, and ᴅ-arabitol utilization.
Abstract
Two Gram-stain negative, facultatively anaerobic, rod-shaped, motile, and non-spore forming bacterial strains, 142053T and 142359T, were isolated from sputum samples obtained from different patients. Growth of strain 142053T and 142359T occurred at 8–42 °C (optimum 28–37 °C), pH 5.0–9.0 (optimum 6.0–8.0) and NaCl concentrations of 0–5% (w/v) (optimum 0–4%). Comparison of 16S rRNA gene sequences showed that strain 142053T and 142359T exhibited high similarity to Kluyvera intermedia NBRC 102594T (99.48%) and Kluyvera georgiana ATCC 51603T (98.68%), respectively. Phylogenomic tree of species from family Enterobacteriaceae revealed that strain 142053T and 142359T formed distinct branches within the Kluyvera clade. The digital DNA–DNA hybridization between 142053T, 142359T, and all type strains of Kluyvera species were ≤39.5% and ≤54.7%, respectively, and the corresponding average…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
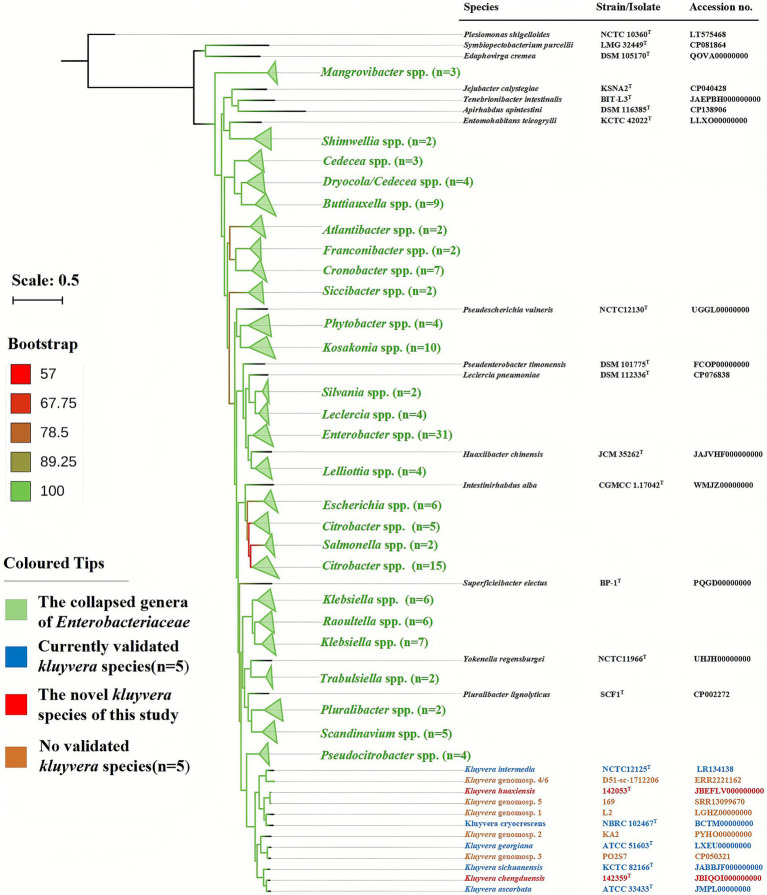 Figure 1
Figure 1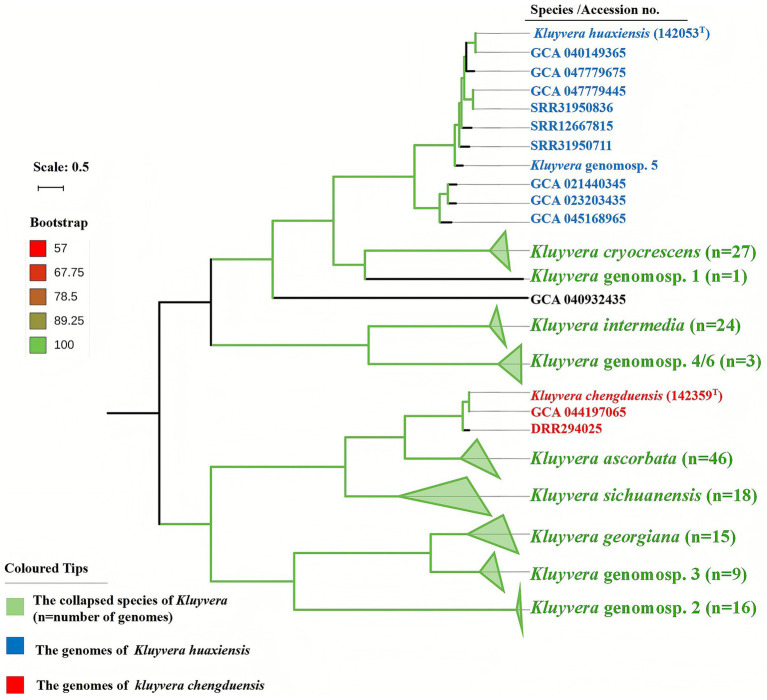 Figure 2
Figure 2| Species | References strain | Accession no. | ANI (%) | POCP (%) | ||||
|---|---|---|---|---|---|---|---|---|
| 142053T | 142359T | 142053T | 142359T | 142053T | 142359T | |||
|
| ATCC 33433T | JMPL01 | 85.98 | 93.85 | 28.9 | 54.7 | 80.51 | 86.15 |
|
| KCTC 82166T | JABBJF01 | 86.30 | 91.31 | 29.7 | 42.8 | 78.30 | 80.47 |
|
| ATCC 51603T | LXEU01 | 85.80 | 87.09 | 28.8 | 30.9 | 79.02 | 78.38 |
|
| NBRC 102467T | BCTM01 | 90.56 | 85.63 | 39.5 | 28.4 | 86.31 | 82.82 |
|
| ATCC 33110T | BCYS01 | 86.66 | 85.45 | 30.4 | 27.8 | 75.82 | 76.49 |
| Genomospecies 1 | L2 | LGHZ01 | 89.69 | 85.58 | 37.4 | 28.3 | 87.03 | 84.03 |
| Genomospecies 2 | KA2 | PYHO01 | 85.54 | 86.61 | 28.2 | 30.1 | 78.54 | 79.77 |
| Genomospecies 3 | PO2S7 | 85.75 | 87.01 | 28.5 | 30.5 | 81.44 | 83.69 | |
| Genomospecies 4/6 | D51-sc-1712206 | ERR2221162 | 86.08 | 85.55 | 29.7 | 27.9 | 76.79 | 79.17 |
| Genomospecies 5 | 169 | SRR13099670 | 98.39 | 86.23 | 87.4 | 29.1 | 85.09 | 77.93 |
|
| 142053T | JBEFLV01 | – | 86.04 | – | 29.0 | – | 80.57 |
|
| 142359T | JBIQOI01 | 86.04 | – | 29.0 | – | 80.57 | – |
| Characteristic | 142053T | 142359T |
|
|
|
|
|
|---|---|---|---|---|---|---|---|
| Motility | + | + | + | + | + | + | + |
| Indole production | + | + | − | + | + | − | + |
| Vojes–Projkauer reaction | − | − | − | − | − | + | − |
| Citrate utilization | + | + | + | + | + | + | + |
| H2S production | − | − | − | − | − | − | − |
| KCN growth in | + | + | + | + | + | + | + |
| Malonate utilization | + | + | + | + | + | + | − |
| NO3 → NO2 | + | + | + | + | + | + | ND |
| ONPG test | + | + | − | + | + | + | + |
| Oxidase | − | − | − | − | − | − | ND |
| Catalase | + | + | − | + | + | + | ND |
| Lipase | W | W | − | − | − | − | − |
| Lysine decarboxylase | + | + | + | + | − | − | + |
| Arginine dihydrolase | + | + | − | − | − | − | − |
| Acid production from: | |||||||
| Sucrose | + | + | − | + | + | + | + |
| Dulcitol | − | − | − | − | − | + | + |
| ᴅ-sorbitol | + | + | + | − | − | + | − |
| ᴅ-arabitol | − | + | − | − | − | − | − |
| Raffinose | + | + | − | + | + | + | + |
| Amygdalin | W | W | + | ND | ND | + | + |
| Glycerol | W | W | − | + | + | + | + |
| ᴅ-mannose | + | + | − | + | + | ND | ND |
| ᴅ-galactose | + | + | + | + | + | ND | ND |
| Methyl α-ᴅ-glucopyranoside | + | + | + | + | + | + | ND |
| ᴅ-lactose | W | + | + | + | + | + | + |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnterobacteriaceae and Cronobacter Research · Infections and bacterial resistance · Fecal contamination and water quality
Introduction
Kluyvera is a genus of Gram-negative, rod-shaped bacteria in the family Enterobacteriaceae (Farmer et al., 1981). Currently, the genus Kluyvera contains five validly published species-Kluyvera ascorbata, Kluyvera cryocrescens, Kluyvera georgiana, Kluyvera intermedia, and Kluyvera sichuanensis and five unnamed genomospecies (1, 2, 3, 4/6, and 5) (Farmer et al., 1981; Müller et al., 1996; Pavan et al., 2005; Liu et al., 2020; Rodriguez and Gutkind, 2024). Kluyvera typically inhabits sewage, hospital sinks, and human sputum, urine, or the gastrointestinal tract (Farmer et al., 1981; Sarria et al., 2001; Li et al., 2019; Alsuheili et al., 2024; Endo et al., 2024). Currently, there is an increasing number of reports on human infections caused by this bacterium, including oral infections, bloodstream infections, enteritis, biliary tract infections, abdominal abscesses, and urinary tract infections. Clinically, these infections pose challenges such as therapeutic difficulty, prolonged treatment duration, and adverse outcomes, including patient death (Mutoh et al., 2019; Zou et al., 2019; Akiki et al., 2023; Inoue et al., 2023; Ulloa-Clavijo et al., 2023). The most frequently reported Kluyvera species associated with human infections are K. ascorbata and K. cryocrescens (Wong, 1987; Lin et al., 2002; Yoshino et al., 2016; Zou et al., 2019), although clinical cases involving K. intermedia and K. georgiana have also been reported in recent years (Inoue et al., 2023; Endo et al., 2024; Xu et al., 2025). Kluyvera possesses a large and significantly flexible gene pool (Yu et al., 2023). Notably, the extended-spectrum β-lactamase gene blaCTX-M and the fosfomycin resistance gene fosA originated from this genus (Humeniuk et al., 2002; Rodríguez et al., 2004; Ito et al., 2018; Rodriguez et al., 2018). Alarmingly, several plasmid-borne carbapenemase genes, including blaKPC (Tiwari et al., 2022), blaOXA-48 (Hernández-García et al., 2018), and blaNDM (Loiodice et al., 2023), and colistin resistance genes mcr (Strepis et al., 2021) have also been identified in Kluyvera species. Taken together, Kluyvera represents an emerging clinical and epidemiological concern due to its dual role as an opportunistic pathogen and as a reservoir of antimicrobial resistance genes. In this study, we conducted a taxonomic analysis on two clinical strains, 142053^T^ and 142359^T^, which were found to represent two novel species within the genus Kluyvera.
Materials and methods
Strains and the study
Strain 142053^T^ and 142359^T^ were isolated from the sputum samples of two different inpatients at West China Hospital in Chengdu (30.6800°N, 104.0811°E), China, in April and August 2024, respectively. The two sputum samples were collected as part of routine care for managing suspected pulmonary infection in the two patients. The sputum samples were vortexed with 0.9% normal saline at a 1:1 ratio for 10 s. The mucoid portion was then picked up with a sterile cotton swab and inoculated onto 5% sheep blood agar (Huankai; Guangzhou, China) plates using the quadrant streak method. The plates were incubated at 37 °C for 48 h, and the colonies of the two isolates were round, smooth, convex, and white. The two isolates were stored long-term at −80 °C in nutrient broth (Huankai) containing 20% glycerol. Preliminary species identification was conducted using matrix-assisted laser desorption ionization time of flight mass spectrum (MALDI-TOF; Bruker; Billerica, MA) with the MBT Compass Library 2023 according to the manufacturer’s instructions. The study has been approved by the Ethical Committee of West China Hospital with waiver of informed consent as this study was to characterize bacterial strains.
Analysis of the 16S rRNA gene sequence
Genomic DNA of strain 142053^T^ and 142359^T^ was extracted using a bacterial genomic DNA extraction kit (Tiangen; Beijing, China). The 16S rRNA gene was amplified using universal primers 27F and 1492R (Moreno et al., 2002), and sequenced bidirectionally using Sanger sequencing (Sangon Biotech; Shanghai, China) for preliminary taxonomic identification. The assembled 16S rRNA gene sequences were compared with the NCBI 16S rRNA database using BLAST, applying a 98% identity threshold to identify closely related genera. Along with those retrieved from type strains of closely related genera, 16S rRNA gene sequences of 142053^T^ and 142359^T^ were aligned using Clustal Omega v1.2.4 (Madeira et al., 2019).
Whole genome sequencing and analysis
Genomic DNA was extracted using the same method as described above. Whole genome sequencing of strain 142053^T^ and 142359^T^ was performed by NovaSeq 6,000 system (Illumina; San Diego, CA, USA). Reads were de novo assembled into contigs using SPAdes v4.0.0 (Bankevich et al., 2012) in isolate modes. The draft genomes of strain 142053^T^ and 142359^T^ were compared with those of type strains of Kluyvera species to assess genomic relatedness using average nucleotide identity (ANI) and in silico DNA–DNA hybridization (isDDH). ANI and isDDH values were calculated using FastANI v1.34 (Jain et al., 2018), OrthoANI (Yoon et al., 2017) and Genome-to-Genome Distance Calculator (formula 2) (Meier-Kolthoff et al., 2013b) with the default settings, respectively. POCP was calculated as described previously (Qin et al., 2014). Sequences were submitted to the online TYGS platform1 to assess their identification at the genus or species level (Meier-Kolthoff and Göker, 2019).
Genome sequences of type strains of all Enterobacteriaceae species were retrieved from GenBank. These sequences were annotated using Prokka v1.14.5 (Seemann, 2014). Orthologues were identified via PIRATE v1.0.5 (Bayliss et al., 2019) to represent the core genome. A maximum-likelihood (ML) phylogenetic tree was inferred from core-genome using IQ-tree v2.1.2 (Minh et al., 2020) with the GTR + GAMMA model and 1,000 bootstrap replicates. Additionally, neighbor-joining (NJ) and maximum parsimony (MP) trees were constructed with 1,000 bootstrap replicates (Kumar et al., 2018) to assess the robustness of the phylogeny. All genomic data analyzed in this study are available in Supplementary 1. Antimicrobial resistance determinants were predicted using ABRicate v1.0.02 against ResFinder databases. Plasmid replicon types were determined using the PlasmidFinder tool available from the Center for Genomic Epidemiology.3
Physiology and chemotaxonomy
Cell motility was examined by observing the bacterial growth and diffusion on the deep semi-solid nutrient medium of 0.3% (w/v) agar (Hopebio; Qingdao, China). Anaerobic growth was examined by streaking the bacterial cultures on brain heart infusion agar plates and placed in the GasPakTM EZ anaerobic bag (BD; Franklin Lakes, NJ, USA) at 37 °C for 3 days. After incubation in nutrient broth at 37 °C for 3 days, the flagella of strain 142053^T^ and 142359^T^ were observed using an H-7650 transmission electron microscope (Hitachi; Tokyo, Japan). Bacterial growth was examined in 5 mL aliquots of nutrient broth. These aliquots were dispensed into tubes with an inner diameter of 16 mm and incubated at temperatures of 4, 8, 18, 28, 32, 37, 42, 45, 48, and 50 °C for 7 days. Meanwhile, salt and pH tolerances were measured using nutrient broth at 37 °C for 7 days at different NaCl concentrations (0, 0.5, 1, 2, 3, 4, 5, 7.5, 10, and 15%, w/v) and at a pH unit of 4.0–12.0 (in increments of 1.0 unit), respectively (Hu et al., 2017). Catalase activity was tested by examining the production of bubbles after addition of 3% (v/v) hydrogen peroxide solution and oxidase activity was measured by using 1% tetramethyl-p-phenylenediamine dihydrochloride solution.
DNase activity was detected using DNase agar (Solarbio; Beijing, China). After incubation at 30 °C for 3 days, 1 M HCl was added for the detection. Malonate, phenylalanine deaminase, and KCN experiments were carried out using biochemical identification tubes (Huankai). Commercially available API 20E, API 50CH, and API ZYM kits (bioMérieux; Marcy l’Etoile, France) were employed to determine biochemical characteristics and enzyme activities. The assays were conducted in accordance with the manufacturer’s instructions, with Escherichia coli strain ATCC 25922 and Pseudomonas aeruginosa strain ATCC 27853 serving as controls. All experiments were performed in triplicate.
Fatty acid analysis
Cellular fatty acid analysis was conducted by the Guangdong Institute of Microbiology (Guangdong, China). In brief, bacteria were inoculated onto nutrient agar plates and incubated at 37 °C for 24 h. Fatty acid methyl esters were extracted and then analyzed via gas chromatography following the guidelines of the Sherlock Microbial Identification System (MIDI Inc.; Newark, DE, USA), as detailed in previous reports (Sasser, 1990). The peaks were automatically integrated, and the proportions of fatty acids were calculated using the MIDI identification database RTSBA6 (version 6.00; MIDI Inc.).
Antimicrobial susceptibility testing
Minimum inhibitory concentrations (MICs) of amikacin, ampicillin, ampicillin-sulbactam, aztreonam, ceftriaxone, ceftazidime, cefepime, cefotaxime, cefuroxime, chloramphenicol, ciprofloxacin, colistin, imipenem, meropenem, piperacillin-tazobactam, sulfamethoxazole-trimethoprim, and tigecycline against these two isolates was determined using the broth microdilution method in accordance with the guidelines of the Clinical and Laboratory Standards Institute (CLSI) (CLSI, 2025). Cation Adjusted Mueller-Hinton Broth (Oxoid, Basingstoke, UK) was used, and antimicrobial powders (purity ≥98%) were purchased from Meilunbio (Dalian, China) or Macklin (Shanghai, China). The breakpoints for colistin and tigecycline were interpreted based on the criteria of the European Committee on Antimicrobial Susceptibility Testing (EUCAST);4 otherwise, the breakpoints defined by the CLSI were applied.
Results
Two novel Kluyvera species were identified
The 16S rRNA gene sequence of strain 142053^T^ showed highest similarity to that of K. intermedia NBRC 102594^T^ (99.48%) and Citrobacter freundii ATCC 8090^T^ (99.22%), while that of strain 142359^T^ was most similar to K. georgiana ATCC 51603^T^ (98.68%) and Klebsiella electrica DSM 102253^T^ (98.11%). A maximum-likelihood tree was constructed based on 16S rRNA gene sequences, incorporating the sequences of the genera Citrobacter, Klebsiella, and Pseudocitrobacter, which are closely related to Kluyvera (Liu et al., 2020). Both strains clustered within the Kluyvera clade. However, they each form relatively long branches distinct from other Kluyvera species, suggesting potential taxonomic novelty (Supplementary Figure S1). Given the limited resolution of 16S rRNA gene-based analyses for differentiating species within the Enterobacteriaceae (Church et al., 2020), whole-genome sequencing was performed to confirm their phylogenomic placement and assess species-level distinctiveness.
Sequencing of strain 142053^T^ yielded 11,580,316 paired-end reads, representing 1.74 Gb of data, with a Q30 score of 93.5% and an average quality score of 26.8. These reads were then assembled into 80 contigs (N50 = 183,280 bp). The draft genome of strain 142053^T^ is 5,003,793 bp in size, with a G + C content of 53.49 mol%. Strain 142359^T^ produced 8,187,706 reads, representing 1.23 Gb of data, with a Q30 score of 95% and an average quality score of 29.2, which were assembled into 52 contigs (N50 = 341,410 bp). The draft genome of strain 142359^T^ is 4,907,633 bp in size, with a G + C content of 54.48 mol%. The POCP values for strains 142053^T^ and 142359^T^ against the reference strains of all Kluyvera species ranged from 75.82 to 87.03% and from 76.49 to 86.15%, respectively, supporting their assignment to the genus Kluyvera. The ANI values between strain 142053^T^ and the type strains of all other Kluyvera species ranged from 85.80 to 90.56%, while strain 142359^T^ showed ANI values between 85.45 and 93.85% relative to these type strains. All values were below the ≥95–96% ANI threshold recommended for species delineation (Meier-Kolthoff et al., 2013b) (Table 1). The OrthoANI values were also below the 95% (Supplementary Figure S2). Correspondingly, the isDDH values between strain 142053^T^ and other Kluyvera type strains ranged from 28.8 to 39.5%, and those for strain 142359^T^ ranged from 27.8 to 54.7%, well below the ≥ 70.0% threshold for species delineation (Richter and Rossello-Mora, 2009; Meier-Kolthoff et al., 2013a). Notably, strain 142053^T^ exhibited high genomic relatedness to the unnamed Kluyvera genomospecies 5, with isDDH and ANI values of 87.4 and 98.39%, respectively (Table 1). These results indicate that K. genomospecies 5 and K. huaxiensis represent the same species. Therefore, K. genomospecies 5 should be formally designated as K. huaxiensis.
A maximum-likelihood phylogenetic tree constructed using 265 core genes from the family Enterobacteriaceae (Figure 1) placed both strain 142053^T^ and 142359^T^ within a clade comprising members of genus Kluyvera. For independent validation, NJ, MP and TYGS-based phylogenomic trees were reconstructed (Supplementary Figures S3–S5), and the results were consistent with the ML phylogeny. This Kluyvera clade was well separated from other related genera, including the most closely related genus Pseudocitrobacter, further supporting that strain 142053^T^ and 142359^T^ belong to the genus Kluyvera. The ML phylogeny inferred from 1,668 core genes within the genus Kluyvera positioned strains 142053^T^ and 142359^T^ in two distinct clades, supporting their classification as novel species, as both were clearly separated from all currently recognized Kluyvera species (Figure 2). Taken together, these genomic and phylogenomic analyses provide robust evidence that strains 142053^T^ and 142359^T^ represent two novel species within the genus Kluyvera.
Phylogenomic tree of strain 142053T, 142359T, and other genera in the family Enterobacteriaceae. The core genome tree was constructed via the maximum-likelihood method based on 1,000 resamplings. Bar, 0.5 substitutions per nucleotide position.
Phylogenomic tree of strain 142053T, 142359T, and other species in the genera Kluyvera. The core genome tree was constructed via the maximum-likelihood method based on 1,000 resamplings. Bar, 0.5 substitutions per nucleotide position.
The physiological and biochemical characteristics of strain 142053^T^ and 142359^T^ are listed in Table 2. Briefly, both strains are positive for indole production, nitrate reduction, citrate utilization, and the activities of lysine decarboxylase, arginine dihydrolase, ornithine decarboxylase, β-galactosidase (ONPG), and catalase. These strains can assimilate aesculin, amygdalin, ᴅ-cellobiose, ᴅ-glucose, ᴅ-galactose, ᴅ-lactose, ᴅ-maltose, ᴅ-melibiose, ᴅ-sorbitol, ᴅ-trehalose, ᴅ-xylose, ʟ-arabinose, ʟ-rhamnose, mannitol, malonate, glycerol, methyl α-ᴅ-glucopyranoside, and salicin. Conversely, they are negative for acetoin production (Voges-Proskauer), DNase activity, H_2_S production, oxidase activity, phenylalanine deaminase activity, and the activities of gelatinase and urease. Moreover, neither strain can utilize adonitol, dulcitol, erythritol, or inositol. In comparison with other species in genus Kluyvera, strains 142053^T^ and 142359^T^ display unique physiological characteristics. Both show weakly positive lipase activity and are positive for arginine dihydrolase. Additionally, the two strains can be distinguished from each another by ᴅ-arabitol utilization, with 142359^T^ but not 142053^T^ can metabolize ᴅ-arabitol. These distinct traits serve as markers for differentiating strains 142053^T^ and 142359^T^ from other Kluyvera species. The cellular fatty acid profiles of two novel strains were analyzed and compared with that of K. sichuanensis GDMCC 1.1872^T^ (Supplementary Table S1). The fatty acid composition of other Kluyvera species has not yet been reported. The major cellular fatty acids of the two strains are C_16:0_, summed feature 3 (C_16:1_ω7c/C_16:1_ω6c), summed feature 8 (C_18:1_ω7c), and C_17:0_ cyclo. Transmission electron micrographs of strains 142053^T^ and 142359^T^ are shown in Supplementary Figsures S6, S7, respectively.
The two strains were resistant to ampicillin but susceptible to ampicillin-sulbactam, aztreonam, ceftriaxone, ceftazidime, cefepime, cefotaxime, ciprofloxacin, imipenem, meropenem, piperacillin-tazobactam, amikacin, colistin, and tigecycline. Strain 142053^T^ was susceptible to cefuroxime but resistant to sulfamethoxazole-trimethoprim, while strain 142359^T^ showed the opposite pattern (Supplementary Table S2). Like other Kluyvera species, both strains possess an intrinsic chromosomal blaCTX-M gene. Strain 142053^T^ encodes CTX-M-3, which is closely related to CTX-M-37 (differing by three amino acids) from Kluyvera genomospecies 5 (now to be considered K. huaxiensis) (Rodríguez et al., 2021), whereas strain 142359^T^ harbors the novel variant CTX-M-283 (GenBank accession no. PV575350). In addition, strain 142053^T^ also carries fosfomycin resistance gene fosA3, sulfonamide-resistance gene sul1, trimethoprim-resistance gene dfrA21, and a plasmid with an IncFIB (pHCM2) replicon, which likely accounts for the last two resistance markers.
Description of Kluyvera huaxiensis sp. nov.
Kluyvera huaxiensis (hua. xi. en’sis N. L. fem. Adj. huaxiensis, referring to West China [“Huaxi” in Chinese], where the type strain was recovered).
The strain 142053^T^ is Gram-negative, facultatively anaerobic, motile, and non-spore-forming rods, measuring 0.6–0.8 μm in width and 1.0–2.5 μm in length. After 12-h incubation on nutrient agar, colonies are round, smooth, convex, and white. Growth occurs at 8–42 °C (optimum, 28–37 °C), pH 5.0–9.0 (optimum, 6.0–8.0), and NaCl concentrations from 0 to 5% (w/v) (optimum, 0–4%). Strain 142053^T^ is positive for indole production, nitrate reduction, citrate utilization, and enzymatic activities including lysine decarboxylase, arginine dihydrolase, ornithine decarboxylase, β-galactosidase (ONPG), and catalase. The strain assimilates aesculin, amygdalin, ᴅ-cellobiose, ᴅ-glucose, ᴅ-galactose, ᴅ-lactose, ᴅ-maltose, ᴅ-melibiose, ᴅ-sorbitol, ᴅ-trehalose, ᴅ-xylose, ʟ-arabinose, ʟ-rhamnose, mannitol, malonate, glycerol, methyl α-ᴅ-glucopyranoside, and salicin. It is negative for acetoin production (Voges-Proskauer test), DNase activity, H_2_S production, oxidase activity, phenylalanine deaminase activity, gelatinase, and urease. No utilization is observed for adonitol, dulcitol, erythritol, or inositol. Notably, the strain exhibits weak lipase activity and is positive for arginine dihydrolase, while ᴅ-arabitol metabolism is not detected. The major cellular fatty acids are C_16:0_ (26.5%), summed feature 3 (C_16:1_ω7c/C_16:1_ω6c, 24.4%), summed feature 8 (C_18:1_ω7c, 18.0%) and C_17:0_ cyclo (9.5%). The genomic DNA G + C content is 53.49 mol%.
The type strain is 142053^T^ (= GDMCC 1.4845^T^ = JCM 37413^T^), isolated from a clinical sputum sample at West China Hospital, Chengdu, China. The GenBank accession number for the 16S rRNA gene sequence is PV388010, and the draft genome sequence has been deposited under JBEFLV000000000 at DDBJ/EMBL/GenBank.
Description of Kluyvera chengduensis sp. nov.
Kluyvera chengduensis (cheng. du. en’sis N. L. fem. Adj. chengduensis, referring to Chengdu, China, where the type strain was recovered).
Cells of strain 142359^T^ are Gram-negative, facultatively anaerobic, motile, non-spore-forming rods, measuring 0.6–0.8 μm in width and 1.0–1.5 μm in length. After 12-h incubation on nutrient agar, colonies are round, smooth, convex, and white. Growth occurs at 8–42 °C (optimum, 28–37 °C), pH 5.0–9.0 (optimum, 6.0–8.0), and NaCl concentrations 0–5% (w/v) (optimum, 0–4%). Strain 142359^T^ is positive for indole production, nitrate reduction, citrate utilization, and the activities of lysine decarboxylase, arginine dihydrolase, ornithine decarboxylase, β-galactosidase (ONPG), and catalase. The strain assimilates aesculin, amygdalin, ᴅ-cellobiose, ᴅ-glucose, ᴅ-galactose, ᴅ-lactose, ᴅ-maltose, ᴅ-melibiose, ᴅ-sorbitol, ᴅ-trehalose, ᴅ-xylose, ʟ-arabinose, ʟ-rhamnose, mannitol, malonate, glycerol, methyl α-ᴅ-glucopyranoside, and salicin. It is negative for acetoin production (Voges–Proskauer), DNase activity, H_2_S production, oxidase activity, phenylalanine deaminase, gelatinase, and urease. The strain does not utilize adonitol, dulcitol, erythritol, or inositol. Additionally, it displays weakly positive lipase activity and is positive for arginine dihydrolase. Strain 142359^T^ can further metabolize ᴅ-arabitol. The major cellular fatty acids are C_16:0_ (24.9%), summed feature 8 (C_18:1_ω7c, 22.7%), summed feature 3 (C_16:1_ω7c/C_16:1_ω6c, 20.0%), and C_17:0_ cyclo (9.7%). The genomic DNA G + C content is 54.48 mol%.
The type strain is 142359^T^ (= GDMCC 1.5094^T^ = JCM 37737^T^), isolated from a clinical sputum sample at West China Hospital, Chengdu, China. The GenBank accession number for the 16S rRNA gene sequence is PV388012, and the draft genome sequence has been deposited under JBIQOI000000000 at DDBJ/EMBL/GenBank.
Discussion
Strains 142053^T^ and 142359^T^ were preliminarily assigned to the Kluyvera species by MALDI-TOF. A maximum-likelihood tree of 265 Enterobacteriaceae core genes placed strains 142053^T^ and 142359^T^ within the Kluyvera clade, clearly separated from related genera. Phylogenomic analysis based on 1,668 Kluyvera core genes further resolved strain 142053^T^ and strain 142359^T^ into distinct branches, supporting their classification as two different species. Although the genus Kluyvera currently comprises only five validly published species, inclusion of five previously proposed but unvalidated genomospecies (1, 2, 3, 4/6, and 5) was necessary to provide a comprehensive phylogenomic framework for the genus. These genomospecies have been consistently recognized in previous taxonomic and genomic studies as distinct Kluyvera lineages, and therefore they were retained in the phylogenetic analysis (Liu et al., 2020; Rodríguez et al., 2021; Rodriguez and Gutkind, 2024). The analysis revealed that K. genomospecies 5 and K. huaxiensis clustered together and exhibited high isDDH and ANI values (Table 1; Figure 2), confirming that they represent the same species. Therefore, the entity previously designated as K. genomospecies 5 should be reclassified as K. huaxiensis. Notably, the genomic sequence named Kluyvera sp. STS39-E (accession no. GCA 040932435) likely represents a novel genomospecies within the genus Kluyvera. As for clinical significance, strain 142053^T^ was isolated from a patient with bilateral interstitial pulmonary fibrosis who did not exhibit any clinical manifestation of pneumonia, and hence it was considered as a colonizer. In contrast, strain 142359^T^ was recovered from a patient with pneumonia, which improved following treatment with piperacillin-tazobactam. However, as the same sputum sample also yielded Klebsiella pneumoniae, the clinical significance of strain 142359^T^ remains uncertain, and it may represent either a pathogen in a mixed infection or a colonizer in a patient with K. pneumoniae-associated pneumonia.
There are limitations of this study. First, only a single isolate was obtained for each of the two proposed species. Although both species are represented by at least two independent genomes, the phenotypic characterization based on a single isolate may not fully capture intraspecies variability. Second, phenotypic features of other Kluyvera species were derived from published studies, which may vary due to difference in experimental conditions. Third, polar lipids, quinones and whole-cell sugar compositions were not analyzed. As such, phenotypic traits used to distinguish the two proposed species from other Kluyvera species should be interpreted with caution.
Nevertheless, the core-genome phylogenetic analysis, together with chemotaxonomic data, clearly places strain 142053^T^ and 142359^T^ within the genus Kluyvera. The ANI and isDDH analyses conclusively demonstrate that these strains represent two novel species, for which the names Kluyvera huaxiensis sp. nov. and Kluyvera chengduensis sp. nov. are proposed.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Akiki M. Ali R. Jamil A. Slim J. Miller R. (2023). Kluyvera ascorbata: an unusual cause of septic shock in a patient with urothelial cancer. Cureus 15:e 51057. doi: 10.7759/cureus.51057, 38269218 PMC 10806381 · doi ↗ · pubmed ↗
- 2Alsuheili A. Z. Alsubaie M. A. Alnajjar A. A. Al-Dajani N. (2024). Kluyvera Cryocrescens urinary tract infection in a neonate: a case report and literature review. Pediatr. Infect. Dis. J. 43, e 461–e 463. doi: 10.1097/inf.0000000000004507, 39265097 · doi ↗ · pubmed ↗
- 3Bankevich A. Nurk S. Antipov D. Gurevich A. A. Dvorkin M. Kulikov A. S. . (2012). SP Ades: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021, 22506599 PMC 3342519 · doi ↗ · pubmed ↗
- 4Bayliss S. C. Thorpe H. A. Coyle N. M. Sheppard S. K. Feil E. J. (2019). PIRATE: a fast and scalable pangenomics toolbox for clustering diverged orthologues in bacteria. Gigascience 8, 1–9. doi: 10.1093/gigascience/giz 119, 31598686 PMC 6785682 · doi ↗ · pubmed ↗
- 5Church D. L. Cerutti L. Gürtler A. Griener T. Zelazny A. Emler S. (2020). Performance and application of 16S r RNA gene cycle sequencing for routine identification of Bacteria in the clinical microbiology laboratory. Clin. Microbiol. Rev. 33:e 00053–19. doi: 10.1128/cmr.00053-19, 32907806 PMC 7484979 · doi ↗ · pubmed ↗
- 6CLSI (Ed.) (2025). Performance standards for antimicrobial susceptibility testing. 35th Edn. Wayne: CLSI supplement M 100. Clinical and Laboratory Standards Institute.
- 7Endo G. Kanai S. Nishio H. Hashimoto H. Higurashi Y. Nomura Y. . (2024). Kluyvera Georgiana bacteremia due to acute cholangitis: a report of the first known case and a literature review. Intern. Med. 63, 2689–2693. doi: 10.2169/internalmedicine.3036-23, 38403769 PMC 11518605 · doi ↗ · pubmed ↗
- 8Farmer J. J. Fanning G. R. Huntley-Carter G. P. Holmes B. Brenner D. J. (1981). Kluyvera, a new (redefined) genus in the family Enterobacteriaceae: identification of Kluyvera ascorbata sp. nov. and Kluyvera cryocrescens sp. nov. in clinical specimens. J. Clin. Microbiol. 13, 919–933. doi: 10.1128/jcm.13.5.919-933.1981, 7240403 PMC 273917 · doi ↗ · pubmed ↗