Therapeutic Potential of Holothuria leucospilota Extract in STZ‐Induced Diabetic Rats: Targeting Inflammation and Apoptosis at the Gene Expression Level
Niloufar Darbandi, Muhanad Yusif Al‐rikabi

TL;DR
A sea cucumber extract reduced diabetes-related inflammation and improved blood sugar in rats, suggesting potential as a natural treatment.
Contribution
The study identifies bioactive compounds in Holothuria leucospilota extract that modulate inflammation and apoptosis in diabetic rats.
Findings
H. leucospilota extract reduced glucose, amyloid beta, and inflammatory markers in diabetic rats.
The extract upregulated anti-inflammatory genes like IL-10 and miR-146a while downregulating pro-inflammatory genes.
GC–MS analysis identified key compounds like olean-12-ene-3,28-diol and oleic acid in the extract.
Abstract
Chronic inflammation is a key contributor to diabetes pathogenesis. Marine‐derived bioactive compounds offer a promising source of therapeutic agents. This study investigated the effects of Holothuria leucospilota n‐hexane extract on biochemical and inflammatory markers in diabetic male rats. Male Wistar rats were divided into four groups: control, diabetic, and diabetic treated with H. leucospilota extract (100 or 200 mg/kg). Diabetes was induced by streptozotocin. Animals received daily intraperitoneal injections of saline or extract. One‐third of the animals in each group (n = 8) were euthanised and sampled on days 1, 15, and 30. The extract composition was analysed by GC–MS. Serum glucose, insulin, amyloid beta, and expression of TGF‐β, TNF‐α, FasL, IL‐10, and miR‐146a were measured in blood, while leptin gene expression was assessed in liver samples. GC–MS revealed major…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
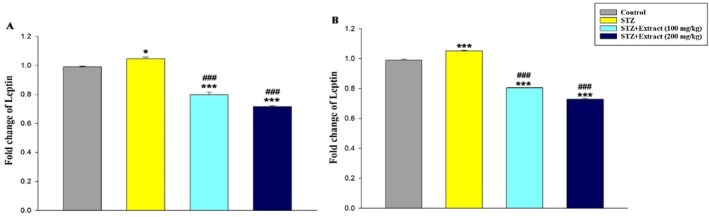 FIGURE 1
FIGURE 1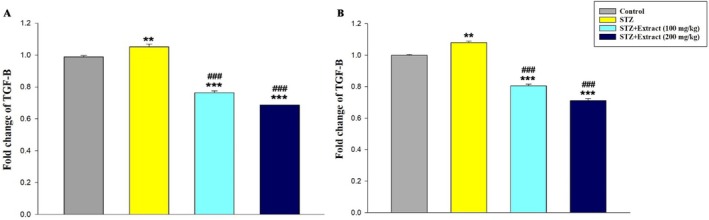 FIGURE 2
FIGURE 2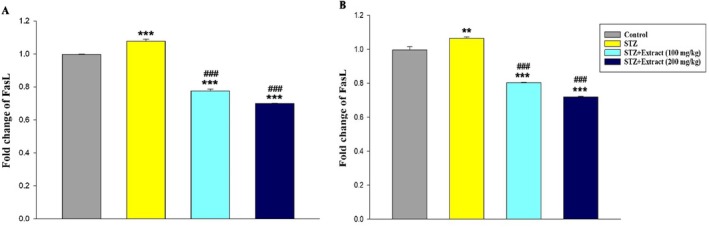 FIGURE 3
FIGURE 3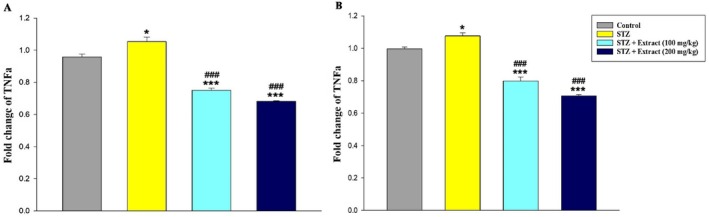 FIGURE 4
FIGURE 4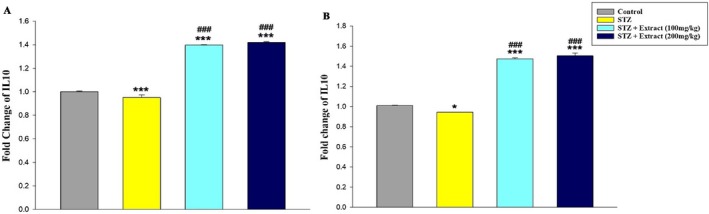 FIGURE 5
FIGURE 5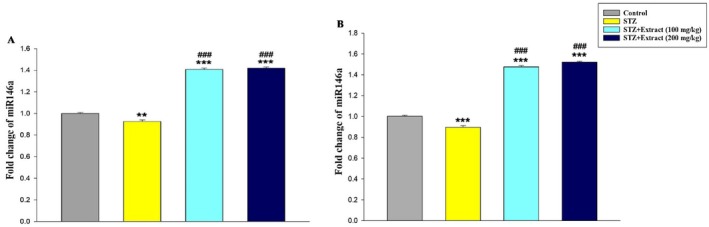 FIGURE 6
FIGURE 6| Gene | Primer direction | Primer sequence (5′ → 3′) | Length (bp) |
|---|---|---|---|
|
| Forward | AGACAGCCGCATCTTCTTGT | 20 |
| Reverse | CTTGCCGTGGGTAGAGTCAT | 20 | |
|
| Forward | AGGTGGAGGTGAACTGGACGG | 21 |
| Reverse | GGCCCACAAAGTCCTCTCAGCAC | 23 | |
|
| Forward | CCCAGCATCTGCAAAGCTC | 19 |
| Reverse | GTCAATGTACAGCTGCCGCA | 20 | |
|
| Forward | GCTCAGCACTGCTATGTTGC | 20 |
| Reverse | TTCATGGCCTTGTAGACACC | 20 | |
|
| Forward | CTGAGGAGAGCCCATTTGAG | 20 |
| Reverse | GGTGTGCACAGAGAGGAGAG | 20 | |
|
| Forward | CATGCAGCAGCCCATGAATTAC | 22 |
| Reverse | CTCTAGGCCCACAAGATGGACAG | 23 | |
|
| Forward | CCGCGCTGAGAACTGAATTCCA | 22 |
| Reverse | AGTGCAGGGTCCGAGGTATT | 20 |
| Category | Compounds | Retention time | Area % |
|---|---|---|---|
| Terpenes and terpenoid | Olean‐12‐ene‐3,28‐diol, (3β) | 10.232 | 14.116 |
| Cyclohexane, 1,1‐bis(5‐methyl‐2‐furyl) | 11.749 | 8.225 | |
| Phorbol | 12.224 | 6.228 | |
| Lumisantonin | 11.071 | 6.130 | |
| Neoisolongifolene, 8‐bromo | 10.893 | 5.992 | |
| Fatty acids | Oleic Acid | 14.68 | 4.753 |
| cis‐13‐Eicosenoic acid | 19.91 | 4.228 | |
| Arachidonic acid | 19.84 | 2.776 | |
| Palmitoleic acid | 17.929 | 2.065 | |
| Eicosanoic acid | 20.9 | 1.560 | |
| 22‐Tricosenoic acid | 25.379 | 1.540 | |
| Others | Tetracyclo [6.1.0.0(2,4)0.0(5,7)] nonane, 3,3,6,6,9,9‐hexaethyl‐, cis, cis, trans— | 11.241 | 8.402 |
| 1‐Heptatriacotanol | 12.434 | 5.035 | |
| 1,4‐Hexadien‐3‐one, 5‐methyl‐1‐[2,6,6‐trimethyl‐2,4‐cyclohexadien‐1‐yl]— | 12.907 | 4.114 | |
| Propanoic acid‐3‐oxo, 3‐(2,4‐dichlorophenyl)‐, ethyl ester | 10.380 | 3.721 | |
| Hexadecanoic acid, 2‐hydroxy‐1‐(hydroxymethyl)ethyl ester | 23.632 | 2.694 | |
| 17‐Pentatriacontene | 24.919 | 2.242 |
| Groups | |||||
|---|---|---|---|---|---|
| Factors | Days | Control | Diabetic | Diabetic + extract (100 mg/kg) | Diabetic + extract (200 mg/kg) |
| Blood glucose (mg/dL) | 1 | 93.1 ± 11.1b | 421.6 ± 8.5a | 440.9 ± 13.5a | 438.6 ± 14.5a |
| 15 | 107.6 ± 8.8d | 431.5 ± 14.6a | 259.2 ± 10.5b | 184.6 ± 5.0c | |
| 30 | 94.3 ± 4.3d | 508.5 ± 11.2a | 233.6 ± 12.0b | 159.5 ± 7.5c | |
| Insulin (micU/mL) | 1 | 13.8 ± 2.6a | 5.8 ± 1.4b | 5.7 ± 1.3b | 5.3 ± 1.7b |
| 15 | 12.5 ± 1a | 5.5 ± 1.3c | 6.4 ± 2.1bc | 7.9 ± 2.0b | |
| 30 | 13.5 ± 1.8a | 4.3 ± 0.8c | 7.4 ± 1.1b | 8.6 ± 2.6b | |
|
Amyloid beta (nmol/mg) | 1 | 0.133 ± 0.01b | 0.359 ± 0.01a | 0.328 ± 0.02a | 0.318 ± 0.03a |
| 15 | 0.114 ± 0.02d | 0.303 ± 0.01a | 0.157 ± 0.03c | 0.204 ± 0.04b | |
| 30 | 0.094 ± 0.01d | 0.381 ± 0.02a | 0.183 ± 0.04c | 0.224 ± 0.02b | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEchinoderm biology and ecology · Seaweed-derived Bioactive Compounds · Protein Tyrosine Phosphatases
Introduction
1
Chronic metabolic and autoimmune disorders, particularly diabetes mellitus, represent a major and growing global health burden worldwide. The development of diabetes is influenced by multiple factors, including genetic predisposition, aging, obesity, and physical inactivity [1]. Type 2 diabetes mellitus (T2DM) is primarily characterised by insulin resistance accompanied by an inadequate compensatory insulin response, which arises from impairments in insulin signalling pathways and subsequent disturbances in glucose and lipid metabolism [2]. Among endocrine regulators, leptin—an adipose tissue–derived hormone—plays a pivotal role in the regulation of body weight, glucose homeostasis, and insulin secretion. Dysregulation of leptin signalling has been closely linked to obesity, systemic inflammation, and the pathogenesis of T2DM [3]. In addition to hormonal alterations, increasing evidence suggests that epigenetic regulators such as microRNAs (miRNAs) contribute to metabolic dysfunction in diabetes. Altered expression of miRNAs in pancreatic β‐cells and insulin target tissues has been reported under diabetic conditions, with miR‐146a emerging as a key mediator linking inflammation and metabolic regulation [4].
Chronic low‐grade inflammation is now recognised as a central mechanism underlying insulin resistance and the progression of diabetes‐related complications [1, 5]. In genetically susceptible individuals, age‐related weight gain is accompanied by sustained activation of inflammatory pathways and increased production of proinflammatory cytokines, particularly tumour necrosis factor‐α (TNF‐α) [6]. TNF‐α disrupts insulin signalling through activation of c‐Jun N‐terminal kinase (JNK) and nuclear factor‐κB (NF‐κB)–dependent pathways, thereby exacerbating insulin resistance and metabolic dysfunction. Conversely, anti‐inflammatory cytokines such as interleukin‐10 (IL‐10) exert protective effects by suppressing proinflammatory mediators and limiting excessive inflammatory responses, highlighting the importance of cytokine balance in metabolic homeostasis [7]. In parallel, transforming growth factor‐β (TGF‐β), a multifunctional cytokine involved in immune regulation, apoptosis, and tissue remodelling, has been implicated in diabetes pathophysiology. TGF‐β1 is expressed in pancreatic β‐cells and has been shown to influence β‐cell survival, while elevated circulating levels of TGF‐β1 have been reported in both type 1 and type 2 diabetes [8, 9]. Moreover, inflammatory conditions promote activation of apoptotic pathways, including Fas ligand (FasL) signalling, which contributes to accelerated β‐cell death and disease progression in diabetes [10].
Despite the availability of various pharmacological agents for diabetes management, effective strategies to prevent or attenuate long‐term hyperglycemic complications remain limited, and many current therapies are associated with adverse effects during prolonged use. Consequently, there is growing interest in identifying novel bioactive compounds with hypoglycemic, anti‐inflammatory, and antioxidant properties [11]. Marine organisms represent a rich source of biologically active natural products with potential metabolic benefits. Sea cucumbers, a group of echinoderms widely used in traditional medicine and functional foods, contain diverse bioactive constituents, including saponins, glycosaminoglycans, sulfated polysaccharides, and essential fatty acids [11, 12]. Experimental studies have demonstrated that sea cucumber–derived compounds, such as fucoidan and glycosaminoglycans, can improve hepatic inflammation, insulin resistance, and glucose metabolism in animal models of metabolic disorders [13]. Given the close interplay between inflammation, apoptosis, and insulin resistance in diabetes, the present study aimed to elucidate the endocrine‐ and inflammation‐related mechanisms underlying the metabolic effects of sea cucumber hexane extract by evaluating inflammatory and apoptotic gene expression in blood and liver tissue, along with serum biochemical parameters, in streptozotocin‐induced diabetic male rats.
Preparation and Extraction of Sea Cucumber Samples
1.1
Deep‐sea sea cucumber samples ( Holothuria leucospilota ) with an average weight of 380 g were collected by divers from the coast of Qeshm, Suza City, and transferred to the laboratory in large containers containing seawater with optimal aeration. The sea cucumber species ( H. leucospilota ) was identified and confirmed using the FAO (2012) identification key for sea cucumbers [14].
In sea cucumber samples, the body wall was separated from other parts by making an incision, divided into small pieces of 1–2 cm, and dried at 45°C until a constant weight was achieved. Approximately 50 g of dried sample was used for extraction. Dry samples were extracted using the Soxhlet method. To this aim, the n‐hexane solvent (Merck, Germany) in a ratio of 1:1.5 was added to Erlenmeyer flasks containing the dry sample and stored at 25°C for 72 h. The use of n‐hexane, a non‐polar solvent, was chosen to selectively extract lipophilic bioactive compounds (e.g., sterols and fatty acids) from sea cucumber, which are known for their anti‐diabetic, anti‐inflammatory, and antioxidant properties. Polar solvents mainly extract hydrophilic compounds, while n‐hexane also ensures compatibility with GC–MS analysis of non‐polar metabolites [15].
After filtering the samples, the solvent was removed using a rotary evaporator (Buchi Rota‐vapour R‐200, Switzerland), and samples were then purified at 45°C and 145 rpm [16]. The chemical composition of the hexane extract of sea cucumber was analysed using a Gas Chromatography–Mass Spectrometry (GC–MS) system (QP2010 SE, Japan). The analysis was performed using a DB‐5MS capillary column (30 m × 0.25 mm × 0.25 μm). Helium was used as the carrier gas at a constant flow rate of 1.2 mL/min. The oven temperature program was as follows: initially held at 50°C for 1 min, then ramped to 280°C at 8°C/min, followed by a further increase to 290°C at 20°C/min, with a final hold for 5 min. To analyse the volatile compounds in the extract, a headspace system was employed, in which the powdered hexane extract was heated at 90°C for 30 min prior to injection. The injector and detector temperatures were set according to manufacturer recommendations, and all analyses were carried out at the University of Tehran. This setup ensured optimal separation and detection of non‐polar bioactive compounds in the sea cucumber extract, compatible with GC–MS analysis.
Laboratory Animals, Grouping, and Treatment
1.2
A total of 96 male Wistar rats, approximately 40 days old and weighing 100–150 g, were purchased from the Razi Vaccine and Serum Research Institute in Tehran. The animals were kept in a 12‐h light–dark cycle with free access to food and water and were randomly divided into four groups (n = 24): control, diabetic, and diabetic + sea cucumber hexane extract (100, 200 mg/kg). The rats were made diabetic through STZ injection intraperitoneally (60 mg/kg). After 3 days, caudal vein sampling and blood sugar level measurement were performed using a glucometer (GlucoDr AGM, 2200, South Korea). An increase in blood sugar of > 300 mg/dL indicated a diabetic animal [17]. On the first day of the treatment phase, following diabetes induction, blood was collected from the heart and liver samples were taken from one‐third of the animals (n = 8) in each group. The remaining 16 animals in each group then entered the treatment phase. The control and diabetic groups received daily intraperitoneal injections of 0.5 cc saline, while the treated diabetic groups received daily intraperitoneal injections of H. leucospilota n‐hexane extract at doses of 100 or 200 mg/kg. In each group, half of the remaining animals (n = 8) were euthanised and sampled on day 15, and the other half (n = 8) on day 30 of treatment [18].
Preparation of Blood and Tissue Samples
1.3
In each group, the animals were anaesthetised with ether 12 h after the last injection, and blood was collected directly from the heart ventricle using a 2 mL syringe. Blood samples were centrifuged at 3000 rpm (Eppendorf, Germany) for 10 min, and the obtained serum was frozen at −80°C [19]. Serum samples were used to examine serum levels of glucose, amyloid beta, and insulin. Whole blood samples were used to examine the expression of TNF‐α, TGF‐β, miRNA‐146a, FasL, and IL‐10 genes. The leptin gene expression was examined using the liver tissue [20].
Serum Glucose, Amyloid Beta, and Insulin Levels
1.4
Serum glucose levels were measured by the enzymatic calorimetric method (Pars Azmoun kit, Iran) using an autoanalyser (Hitachi, Germany). Amyloid beta was quantified by ELISA using an East Biopharm research kit (China) with a sensitivity of 1.08 ng/mL and a coefficient of variation of 10%–12%. Serum insulin levels were measured by radioimmunoassay according to the kit instructions (Diagnostic DSL Systems Laboratories, USA).
RNA Extraction and cDNA Synthesis
1.5
RNA was extracted from blood cells and the liver tissue using an RNA extraction kit (Sambio, Taiwan) according to the manufacturer's protocol. The extracted RNA was stored at −70°C. The RNA concentration of each sample was measured with a NanoDrop spectrophotometer (Titertek Berthold, Germany) at the absorbance ratios of 260/280 and 280/230.
To synthesise cDNA with a kit (BIOFACT, Korea), 1000 ng of extracted RNA was mixed with Master Mix, Random Hexamer, and oligo dT and vortexed for a few seconds. The microtubes were incubated at 37°C for 5 min and then at 50°C for 30 min (for reverse transcription). Finally, the reaction mixture was treated at 95°C for 5 min (to inactivate the reverse transcriptase enzyme). The cDNA samples were stored at −20°C until PCR analysis.
Gene Expression Analysis
1.6
Gene expression levels were measured using qRT‐PCR on an ABI 7300 System (Applied Biosystems, USA). Each 20 μL reaction contained 10 μL BIOFACT Mastermix, 2 μL cDNA, 1 μL forward primer, 1 μL reverse primer, and 6 μL nuclease‐free water. Initial denaturation was performed at 95°C for 15 min, followed by 40 cycles of denaturation at 95°C for 20 s, and annealing/elongation at 60°C for 40 s. Melting curves were generated from 60°C to 95°C to confirm primer specificity. Gene expression was quantified relative to the internal reference gene GAPDH using the 2^−ΔΔCT^ method. Primers were designed and synthesised by KavoshGene Company (Table 1), and their specificity was confirmed by BLAST and melting curve analysis.
Statistical Analysis
1.7
Data were statistically analysed with the one‐way analysis of variance (ANOVA) using SPSS software. Results were reported as mean ± SEM at a significance level of p < 0.05. All measurements were performed in biological triplicates.
Results
2
Results of GC‐MASS Analysis of
H. leucospilota Hexane Extract
2.1
The GC–MS analysis revealed that the extract contained a variety of terpene‐terpenoid compounds, including olean‐12‐ene‐3,28‐diol (3β) (14.116%) and cyclohexane, 1,1‐bis(5‐methyl‐2‐furyl) (8.225%), as well as fatty acids such as oleic acid (4.753%) and cis‐13‐eicosenoic acid (4.228%), which were among the major constituents of the extract (Table 2). The percentages are based on GC–MS peak areas, and only the major compounds are reported.
TABLE 2: The main components identified in the n‐hexane extract of the body wall of H. leucospilota .
Serum Levels of Insulin, Glucose, and Amyloid Beta in the Experimental Groups
2.2
The results of one‐way ANOVA showed statistically significant differences in the mean variables of blood glucose, insulin, and amyloid beta between the experimental groups on days 1, 15, and 30 (p < 0.001) (Table 3). The serum levels of glucose and amyloid beta increased significantly, and the insulin level decreased significantly in the groups receiving STZ compared to the control group. The serum levels of glucose and amyloid beta decreased significantly, and the insulin level increased in diabetic rats treated with H. leucospilota n‐hexane extract (100 and 200 mg/kg) on days 15 and 30 of treatment compared to the diabetic group.
Quantitative Analysis of Changes in Leptin, miR‐146a, TGF‐β, IL‐10, TNF‐α, and FasL Gene Expression Levels Using Real‐Time PCR
2.3
The effect of H. leucospilota n‐hexane extract on the expression of the target genes in diabetic rats was evaluated using real‐time PCR analysis. After confirming the specificity of the bands observed in semi‐quantitative RT‐PCR, the expression of the studied genes was quantified with the PCR technique (35 reaction cycles).
Analysis of melting and amplification curves demonstrated sharp single peaks for leptin, miR‐146a, TGF‐β, IL‐10, TNF‐α, and FasL PCR products, confirming the specificity of amplification. No primer‐dimer artefacts were observed, and negative control samples showed no detectable amplification.
Data analysis indicated that leptin gene expression (Figure 1A,B) in the liver tissue and TGF‐β (Figure 2A,B), FasL (Figure 3A,B), and TNF‐α (Figure 4A,B) genes increased significantly in the blood of diabetic rats in 15‐day and 30‐day periods compared to the control group. According to the results, significant decreases in the expression of these genes were observed in diabetic rats treated with H. leucospilota hexane extract at concentrations of 100 and 200 mg/kg in both periods of 15 and 30 days compared to the diabetic group.
*Expression of leptin gene over a period of 15 (A) and 30 (B) days in different experimental groups using Real‐time‐PCR technique. Data are presented as mean ± SEM of eight animals per group. *p < 0.05, **p < 0.001 as compared with control group. ### p < 0.001 as compared with STZ group.
*Expression of TGF‐β gene over a period of 15 (A) and 30 (B) days in different experimental groups using Real‐time‐PCR technique. Data are presented as mean ± SEM of eight animals per group. **p < 0.01, ***p < 0.001 as compared with control group.
p < 0.001 as compared with STZ group.*
*Expression of FasL gene over a period of 15 (A) and 30 (B) days in different experimental groups using Real‐time‐PCR technique. Data are presented as mean ± SEM of eight animals per group. **p < 0.01, ***p < 0.001 as compared with control group.
p < 0.001 as compared with STZ group.*
*Expression of TNF‐α gene over a period of 15 (A) and 30 (B) days in different experimental groups using Real‐time‐PCR technique. Data are presented as mean ± SEM of eight animals per group. *p < 0.05, ***p < 0.001 as compared with control group.
p < 0.001 as compared with STZ group.*
The analyses also showed that the expression of IL‐10 (Figure 5A,B) and miR‐146a (Figure 6A,B) decreased significantly in the blood of diabetic rats in the periods of 15 and 30 days compared to the control group. In addition, a significant increase in the expression of these genes was seen in diabetic rats treated with H. leucospilota hexane extract at concentrations of 100 and 200 mg/kg in both periods of 15 and 30 days compared to the diabetic group.
*Expression of IL‐10 gene over a period of 15 (A) and 30 (B) days in different experimental groups using Real‐time‐PCR technique. Data are presented as mean ± SEM of eight animals per group. *p < 0.05, ***p < 0.001 as compared with control group.
p < 0.001 as compared with STZ group.*
*Expression of miR‐146a gene over a period of 15 (A) and 30 (B) days in different experimental groups using Real‐time‐PCR technique. Data are presented as mean ± SEM of eight animals per group. **p < 0.01, ***p < 0.001 as compared with control group.
p < 0.001 as compared with STZ group.*
Discussion
3
STZ enters pancreatic beta cells via GLUT2 and causes DNA alkylation and damage. This damage activates the poly ADP ribosylation repair process, which destructs pancreatic tissue by producing free radicals, leading to hyperglycemia [21]. The resulting increase in islet amyloid polypeptide (amylin), produced and secreted along with insulin by pancreatic β cells, promotes the formation of toxic amyloid oligomers. These oligomers contribute to the development of diabetes, apoptosis, and progressive beta cell failure by activating the extrinsic apoptosis pathway through reactive oxygen species generation, impaired mitochondrial function, chromatin condensation, and other apoptotic mechanisms [22, 23].
Sea cucumber extract contains antioxidants and bioactive compounds, including flavonoids, saponins, alkaloids, eicosapentaenoic acid, and docosahexaenoic acid, which have been reported to exert antidiabetic effects [24]. The fatty acids present in the extract may enhance tissue insulin sensitivity by inhibiting alpha‐glucosidase activity [25, 26], while saponins contribute to the reduction of inflammation and improvement of insulin resistance [27]. Fucoidans derived from sea cucumber can promote glucose transport in skeletal muscle and increase insulin sensitivity [28], and glycosaminoglycans may suppress hepatic glucose production [29]. Collectively, these mechanisms are expected to support better glycemic control and modulate factors involved in β‐cell dysfunction [30].
Amplified leptin levels are associated with the risk of diabetes or obesity. Leptin production by white adipose tissue increases significantly in diabetic individuals and leads to reduced glucose tolerance by lowering insulin sensitivity. Elevated leptin levels result in declined responsiveness of pancreatic β cells, inability to suppress insulin secretion, and hyperinsulinemia. This can in turn exacerbate obesity, increase leptin gene expression, and amplify its level in blood serum. In diabetic individuals, therefore, increased leptin expression in a positive feedback loop causes metabolic disorders [31].
Bioactive metabolites present in sea cucumber extract reduce the transcription of appetitive neuropeptides and regulate glucose homeostasis by reducing endoplasmic reticulum stress and phosphorylation of JAK2/STAT3 pathway proteins. On the other hand, they reduce leptin secretion by increasing the levels of stress hormones (catecholamines and cortisol) and inhibiting the phosphodiesterase nucleotide cycle [32].
TGF‐β plays a pivotal role in the development of insulin resistance and obesity. Plasma levels of TGF‐β1 rise significantly in diabetic patients. Elevated blood sugar levels increase the TGF‐β1 gene transcription by increasing protein kinase C (PKC) enzyme activity [11]. Long‐term and chronic inflammation caused by metabolic disorders (e.g., diabetes) increases TGF‐β expression by inducing oxidative stress, leading to excessive deposition of collagen fibres in the extracellular matrix and fibrosis formation. Bioactive metabolites of sea cucumber extract can inhibit tissue fibrosis and reduce TGF‐β gene expression by suppressing the TGF‐β/SMAD pathway [33]. Treatment with eicosapentaenoic acid contained in sea cucumber extract significantly inhibits the activation of TGF‐β and Smad3, increases the phosphorylation level of PI3K/AKT, suppresses NF‐κB activation, reduces the expression of proinflammatory cytokines, and suppresses oxidative stress and the mitochondrial‐mediated apoptosis signalling pathway [34].
FasL seems to be the likely mediator for the lysis of Islets of Langerhans. The level and timing of Fas expression by Islets of Langerhans is induced by incubation with IL‐1β and IFNɣ. A significant increase in Fas gene expression occurs in an inflammatory environment and in response to cytokines, which is accompanied by an accelerated phase of beta cell death [13]. Saponins and fucoidans in sea cucumber extract seem to reduce FasL expression by inhibiting NF‐kB activity in T cells [35].
TNF‐α appears to play a direct role in the development and progression of diabetic complications through cell apoptosis and increased expression of inflammatory factors [36] and inhibits insulin receptor signalling and activity by reducing GLUTA4 [37]. Conversely, IL‐10 has profound and diverse anti‐inflammatory effects; this cytokine inhibits the expression of inflammatory interleukins [7]. In vitro studies have documented that IL‐10 protects against TNF‐α‐dependent insulin resistance in adipocytes [8]. By increasing IL‐10 levels, sea cucumber extract inhibits hepatic inflammation in diabetic rats and improves insulin systemic function [38].
Dysregulation of miRNA expression affects a variety of important cellular functions and significantly impacts health and diabetes development. miR‐146a prevents the increased production of inflammatory cytokines in diabetic patients and restores β‐cell function in T2DM through the NUMB/β‐catenin signalling pathway [39]. High blood sugar levels in diabetic individuals cause a decrease in miR‐146a expression [40]. Fucoidans, polysaccharides found in sea cucumber, have potential useful hypoglycemic effects in diabetes treatment; these compounds can increase miRNA‐29 and miRNA‐146a levels [41]. Saponins also regulate the IRAK1/TRAF6 pathway by increasing the expression of miR‐146a, reducing inflammation, and improving diabetes by inhibiting the NF‐κB pathway [42].
Limitations
4
Although the STZ‐induced diabetes model is widely used, it replicates only certain features of diabetes, and direct extrapolation of the results to humans is limited. The sample size (n = 8 per group) may also restrict the statistical power to detect subtle differences, indicating the need for studies with larger sample sizes. Moreover, in this study, the crude n‐hexane extract of Hikomaruta was used, and it would be preferable in future research to evaluate the effects of individual active compounds separately. Additionally, future investigations should assess the toxicity of the extract and its long‐term effects on diabetes, inflammation, and related molecular pathways.
Conclusion
5
This study highlights the therapeutic potential of H. leucospilota n‐hexane extract as a multifunctional modulator of diabetes‐related metabolic dysfunction. The findings emphasise that the beneficial effects of the extract extend beyond glycemic regulation and involve coordinated regulation of inflammatory, apoptotic, and endocrine‐related molecular pathways. By targeting key mediators that link chronic inflammation, β‐cell dysfunction, and insulin resistance, sea cucumber–derived bioactive compounds may contribute to preserving metabolic homeostasis in diabetes. From a scientific perspective, this work underscores the relevance of marine natural products as sources of bioactive molecules capable of influencing epigenetic and immune‐related mechanisms implicated in diabetes progression. These insights provide a mechanistic framework for future studies aimed at isolating active compounds and exploring their translational potential as adjunct strategies in diabetes management.
Author Contributions
N.D. and M.Y.A.: designed the experiments; N.D.: Supervised, directed and managed the study; M.Y.A.: performed experiments and collected data. N.D.: prepared figures and tables, wrote the main manuscript text; N.D. and M.Y.A.: discussed the results and strategy and final approved of the version to be published.
Funding
The authors have nothing to report.
Ethics Statement
All procedures were approved by the local ethical committee (Research and Ethics Committee of the School of Biology, University of Arak; IR.ARAKMU.REC.1403.039) and were carried out in accordance with the ethical standards and principles of laboratory animal care (NIH publication) and animal protection laws.
Animal Studies: All animal experiments complied with relevant guidelines and regulations.
Consent
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1M. Lotfy , J. Adeghate , H. Kalasz , J. Singh , and E. Adeghate , “Chronic Complications of Diabetes Mellitus: A Mini Review,” Current Diabetes Reviews 13 (2017): 3–10, 10.2174/1573399812666151016101622.26472574 · doi ↗ · pubmed ↗
- 2E. Ahmad , S. Lim , R. Lamptey , D. R. Webb , and M. J. Davies , “Type 2 Diabetes,” Lancet 400 (2022): 1803–1820, 10.1016/S 0140-6736(22)01655-5.36332637 · doi ↗ · pubmed ↗
- 3K. Rehman , M. S. H. Akash , and Z. Alina , “Leptin: A New Therapeutic Target for Treatment of Diabetes Mellitus,” Journal of Cellular Biochemistry 119 (2018): 5016–5027.29236298 10.1002/jcb.26580 · doi ↗ · pubmed ↗
- 4E. Roggli , S. Gattesco , D. Caille , et al., “Changes in micro RNA Expression Contribute to Pancreatic β‐Cell Dysfunction in Prediabetic NOD Mice,” Diabetes 61 (2012): 1742–1751.22537941 10.2337/db 11-1086 PMC 3379668 · doi ↗ · pubmed ↗
- 5B. Shamshadi , R. Askari , R. Rezaei , and A. H. Haghighi , “Investigation of an Increase in the Expression of the Mir 146a Gene in the Hippocampus and a Decrease in the Levels of Blood Sugar, Insulin, and Insulin Resistance in Elderly Diabetic Rats After a Period of High Intensity Interval Training,” Diabetes, Metabolic Syndrome and Obesity: Targets and Therapy 14 (2023): 75–86.
- 6M. Satoh , M. Nakamura , H. Satoh , H. Saitoh , I. Segawa , and K. Hiramori , “Expression of Tumor Necrosis Factor‐Alpha–Converting Enzyme and Tumor Necrosis Factor‐Alpha in Human Myocarditis,” Journal of the American College of Cardiology 36 (2000): 1288–1294.11028485 10.1016/s 0735-1097(00)00827-5 · doi ↗ · pubmed ↗
- 7P. Feng , J. Chai , M. Zhou , N. Simon , L. Huang , and H. Wang , “Interleukin‐10 Is Produced by a Specific Subset of Taste Receptor Cells and Critical for Maintaining Structural Integrity of Mouse Taste Buds,” Journal of Neuroscience 34 (2014): 2689–2701.24523558 10.1523/JNEUROSCI.3074-13.2014 PMC 3921433 · doi ↗ · pubmed ↗
- 8X. Yuan , X. Dai , L. Liu , et al., “Comparing the Effects of 6 Months Aerobic Exercise and Resistance Training on Metabolic Control and β‐Cell Function in Chinese Patients With Prediabetes: A Multicenter Randomized Controlled Trial,” Journal of Diabetes 12 (2020): 25–37.31141300 10.1111/1753-0407.12955 · doi ↗ · pubmed ↗