Effects of Betaine on Performance, Blood Biochemistry, Nutrient Utilization and Gut Health in Coccidia‐Infected Broilers
Abdul Hafeez, Usama Saleem, Shabana Naz, Rifat Ullah Khan, Muhammad Israr, Ala Abudabos, Ibrahim A. Alhidary

TL;DR
This study shows that adding betaine to the diet of chickens infected with coccidia can improve their growth, digestion, blood health, and gut condition.
Contribution
The study demonstrates that betaine at 600 mg/kg is most effective in mitigating coccidiosis effects in broilers.
Findings
Betaine at 600 mg/kg significantly improved growth performance and nutrient digestibility in infected broilers.
Higher betaine doses reduced caecal lesion scores and improved blood lipid profiles in coccidia-challenged chickens.
The study suggests betaine could be a supportive dietary strategy against coccidial infection.
Abstract
The objective of this study was to investigate the ameliorative effect of betaine on the performance, blood biochemistry, intestinal lesion score and nutrient utilization of broiler chickens exposed to coccidian challenge. A total of 600 one‐day‐old broiler chicks (Ross 708) were randomly assigned to five groups (six replicates). The negative control group (NC) remained unchallenged and received non‐supplemented feed, while the positive control group (PC) received non‐supplemented feed and was challenged with oocysts. The other three groups (B2, B4 and B6) were challenged with oocysts and received feed supplemented with 200, 400 and 600 mg/kg betaine, respectively. Results indicated a significant decrease in growth performance, nutrient digestibility and altered blood lipid profiles in the PC group, with the most significant restoration observed in the B6 group. Caecal lesion scores…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
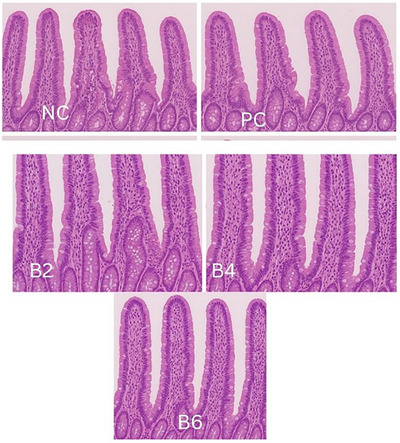 FIGURE 1
FIGURE 1| 1–21 days | 22–45 days | |
|---|---|---|
| Corn | 60.00 | 61.00 |
| Soybean meal (46.8%) | 30.00 | 28.0 |
| Soybean oil | 3.50 | 4.50 |
| Corn gluten meal | 2.00 | 2.70 |
| Limestone | 1.30 | 1.50 |
| Dicalcium phosphate | 2.00 | 1.50 |
| Salt | 0.10 | 0.10 |
| Premix | 0.50 | 0.50 |
| L‐Methionine | 0.13 | 0.19 |
| Calculated nutrient contents | ||
| Crude protein (%) | 22.0 | 19.50 |
| Metabolizable energy (MJ/kg) | 12.30 | 12.50 |
| Lysine (%) | 1.25 | 1.10 |
| Calcium (%) | 1.00 | 0.95 |
| Methionine (%) | 0.45 | 0.40 |
| Available phosphorus (%) | 0.4 | 0.36 |
| Groups | NC | PC | B2 | B4 | B6 | SEM |
|
|---|---|---|---|---|---|---|---|
| Week‐1 | 118d | 116cd | 124c | 130b | 135a | 1.94 | <0.001 |
| Week‐2 | 321b | 324b | 334a | 338a | 343a | 2.32 | <0.001 |
| Week‐3 | 529a | 521b | 537bc | 544ab | 552a | 2.97 | <0.001 |
| Starter (d1–21) | 969d | 961d | 994c | 1012b | 1030a | 7.01 | <0.001 |
| Week‐4 | 837a | 762b | 756bc | 748cd | 740d | 9.33 | <0.001 |
| Week‐5 | 959a | 925b | 924b | 910c | 905c | 5.07 | <0.001 |
| Week‐6 | 1074a | 1011d | 1024c | 1038b | 1072a | 6.80 | <0.001 |
| Finisher (d21–42) | 2869a | 2699c | 2705bc | 2696c | 2717b | 17.8 | <0.001 |
| Overall (d1–42) | 3838a | 3660d | 3699c | 3708c | 3747b | 16.3 | <0.001 |
| Groups | NC | PC | B2 | B4 | B6 | SEM |
|
|---|---|---|---|---|---|---|---|
| Week‐1 | 102d | 102d | 111c | 118b | 125a | 2.47 | <0.001 |
| Week‐2 | 252c | 250c | 262b | 274a | 280a | 3.25 | <0.001 |
| Week‐3 | 356c | 352c | 369b | 376b | 387a | 3.52 | <0.001 |
| Starter (d1–21) | 709d | 705d | 742c | 768b | 793a | 9.09 | <0.001 |
| Week‐4 | 498a | 418c | 426bc | 428bc | 436b | 7.82 | <0.001 |
| Week‐5 | 559a | 495d | 510c | 526b | 532b | 5.82 | <0.001 |
| Week‐6 | 607b | 539e | 570d | 584c | 621a | 7.77 | <0.001 |
| Finisher (d21–42) | 1665a | 1452e | 1507d | 1537c | 1590b | 19.5 | <0.001 |
| Overall (d1–42) | 2374a | 2156d | 2249c | 2305b | 2383a | 22.6 | <0.001 |
| Groups | NC | PC | B2 | B4 | B6 | SEM |
|
|---|---|---|---|---|---|---|---|
| Week‐1 | 1.16a | 1.14ab | 1.12ab | 1.10ab | 1.08b | 0.010 | 0.035 |
| Week‐2 | 1.27ab | 1.29a | 1.27ab | 1.23b | 1.22b | 0.008 | 0.005 |
| Week‐3 | 1.49a | 1.48bc | 1.45bc | 1.45bc | 1.43c | 0.007 | 0.004 |
| Starter (d1–21) | 1.36a | 1.36a | 1.34ab | 1.32bc | 1.30c | 0.007 | <0.001 |
| Week‐4 | 1.68c | 1.82a | 1.78b | 1.75b | 1.70c | 0.014 | <0.001 |
| Week‐5 | 1.71cd | 1.87a | 1.81b | 1.73c | 1.70d | 0.017 | <0.001 |
| Week‐6 | 1.77c | 1.88a | 1.80b | 1.78bc | 1.72d | 0.014 | <0.001 |
| Finisher (d21–42) | 1.72d | 1.86a | 1.79b | 1.75c | 1.71d | 0.015 | <0.001 |
| Overall (d1–42) | 1.62c | 1.70a | 1.64b | 1.61c | 1.57d | 0.011 | <0.001 |
| Groups | NC | PC | B2 | B4 | B6 | SEM |
|
|---|---|---|---|---|---|---|---|
| Dry matter | 72.7a | 52.2d | 60.5c | 65.3bc | 68.3ab | 1.41 | <0.001 |
| Ash | 49.3a | 44.7b | 45.8ab | 49.0ab | 49.2ab | 0.58 | 0.012 |
| Crude protein | 70.8a | 60.3c | 66.3b | 67.8ab | 71.5a | 0.83 | <0.001 |
| Crude Fat | 79.8a | 52.8e | 59.5d | 67.5c | 73.5b | 1.84 | <0.001 |
| Groups | NC | PC | B2 | B4 | B6 | SEM |
|
|---|---|---|---|---|---|---|---|
| Triglycerides (mg/dL) | 47.5cd | 45.0d | 48.8bc | 51.0ab | 53.3a | 0.589 | <0.001 |
| Total cholesterol (mg/dL) | 112ab | 101c | 110b | 112ab | 114a | 0.923 | <0.001 |
| LDL (mg/dL) | 49.0a | 45.5c | 46.8ab | 48.2ab | 47.8ab | 0.383 | 0.029 |
| HDL (mg/dL) | 53.5a | 46.7b | 53.0a | 53.7a | 55.5a | 0.615 | <0.001 |
| Total Protein (g/dL) | 4.27 | 4.42 | 4.55 | 4.40 | 4.45 | 0.034 | 0.122 |
| Caecal lesion score | 0.00c | 2.1a | 1.99a | 1.91b | 1.90b | 0.331 | 0.034 |
- —King Saud University10.13039/501100002383
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCoccidia and coccidiosis research · Animal Nutrition and Physiology · Rabbits: Nutrition, Reproduction, Health
Introduction
1
Coccidiosis, an enteric protozoan parasitic disease in poultry, is associated with substantial economic losses primarily attributed to severe diarrhoea and elevated mortality rates (Rashid and Shnawa 2024; Batool et al. 2025). Most of these economic burdens arise from diminished performance, with the remainder stemming from prophylactic and treatment expenses (Abbas et al. 2025; Nasir et al. 2023). Coccidial infection in broilers induces epithelial cell damage, diarrhoea and intestinal osmotic stress (Khan et al. 2023), subsequently leading to nutrient malabsorption (Saeeda et al. 2023; Almahallawi et al. 2024). Decreases in nutrient and energy digestibility have been documented in birds exposed to coccidia, and the extent of these responses is influenced by diet composition as well as the type and quantity of Eimeria species introduced in the challenge model (Ishaq et al. 2022).
Betaine is a naturally occurring compound first isolated from sugar beet (Beta vulgaris) and widely present in plants, microorganisms and animal tissues (Ghasemi and Nari 2020). Chemically, it is a trimethyl derivative of the amino acid glycine, often referred to as trimethylglycine (Chand et al. 2017). Its unique zwitterionic structure enables it to function both as a methyl donor and as an osmolyte, making it particularly valuable in poultry nutrition (Saeed et al. 2017). Dietary supplementation with betaine has been reported to positively influence growth performance, metabolism and immunity (Chand et al. 2017; Ghasemi and Nari 2020). As a lipotropic agent, betaine reduces hepatic lipid deposition and enhances lipid turnover in the body, proving useful in diets with a low crude protein and high metabolizable energy‐to‐protein ratio. In addition, its osmoregulatory effect supports immune, cardiovascular, nervous and renal functions (Sakomura et al. 2013). These properties enable betaine to alleviate heat stress (Saeed et al. 2017), protect intestinal enzymes and improve nutrient digestibility (Chand et al. 2017).
Despite these established benefits, limited studies have directly examined the role of betaine in mitigating intestinal damage and nutrient malabsorption specifically under Eimeria‐induced coccidiosis in broilers. Most prior work has focused on its effects under nutritional stress or heat stress, leaving a critical gap in understanding its potential as an alternative or adjunct to conventional anticoccidials.
Therefore, this study was designed to address this gap by investigating whether dietary betaine supplementation at different inclusion levels could alleviate intestinal lesions, improve nutrient digestibility and restore growth performance and blood biochemistry in broilers experimentally challenged with Eimeria spp.
Materials and Methods
2
Birds and Treatments
2.1
A total of 600 one‐day‐old broiler chicks (Ross 708) were acquired, each weighing approximately 42 ± 2 g. The chicks were subjected to a lighting program of 23 h of light and 1 h of darkness. They were housed in pens with dried wood shavings, and throughout the study, access to feed and water was provided ad libitum. The broiler chicks were fed a corn‐soybean meal basal diet, free of coccidiostats, which met the recommended requirements outlined in the Ross catalogue for both the starter and finisher phases. The broiler chicks were randomly divided into five groups, each consisting of six replicates and 20 birds per group. The first group served as the negative control (NC), receiving non‐supplemented feed and remaining unchallenged with oocysts. The second group, positive control (PC), received non‐supplemented feed but was challenged with oocysts. The remaining three groups, B2, B4 and B6, were challenged with oocysts and received feed supplemented with 200, 400 and 600 mg/kg betaine, respectively. Betaine (anhydrous betaine, ≥ 99% purity) was procured from Sigma‐Aldrich, St. Louis, MO, USA]. On Day 21, all groups, except the NC, were subjected to a challenge with a mixture of Eimeria species, involving the oral inoculation of a 1.5 mL solution containing sporulated oocysts (2 × 10^5^ oocysts). The Eimeria species composition included Eimeria necatrix (10%), E. acervulina (11%), E. maxima (9%), and E. tenella (70%). The solution also contained 96% natural betaine from Marlborough, UK. In addition, the diets were formulated with chromium dioxide (0.3%) as an inert marker, and the pellets were produced at 70°C. The diets were provided ad libitum (Table 1), and water access was unrestricted throughout the entire trial.
Growth Performance
2.2
Feed intake (FI) was calculated as the difference between the offered and refused feed. Weight gain (WG) of chicks was determined as the difference in bird weight at the end and start of each week. The feed conversion ratio (FCR) was computed by dividing the total amount of feed consumed by chicks by their WG. Mortality was monitored daily, and any dead birds were recorded. Since no mortality occurred during the trial (despite coccidial challenge), growth performance data were not confounded by mortality effects (Hafeez et al. 2020).
Lesion Score
2.3
On Day 7 post‐challenge (Day 21 post‐hatch), two birds were randomly chosen from each replicate cage, euthanized by cervical dislocation, and lesion scoring was performed by a skilled avian veterinarian unaware of treatment allocations. Coccidiosis lesions in the ceca were assessed on a scale of 0–4, following the method outlined by Almahallawi et al. (2024). A score of zero indicated the absence of gross lesions, while 4 denoted extensive haemorrhage or lesions, depending on the Eimeria species involved.
Apparent Total Digestibility
2.4
On Day 42, two birds per replicate were selected, weighed, stunned and euthanized by exsanguination. Faecal samples were collected from the cloaca and subsequently freeze‐dried at −20°C for further chemical analysis. The apparent total digestibility (ATD) of nutrients was determined using the formula (Ullah et al. 2022):
Blood Biochemical Analysis
2.5
On the 42nd day of the trial, blood samples were extracted from the wing veins of two birds per treatment and placed in tubes without anticoagulant. The blood samples underwent centrifugation for 12 min at 1300 × g and 4°C. Subsequently, sera were collected and stored at −20°C for later analysis (Chand et al. 2018). Triglycerides (TG), total cholesterol (CHOL), LDL, HDL and total protein levels were measured using a commercial kit (Biocheck, UK; Cat. No. BC1117, Cholesterol Cat. No. BC1120, LDL Cat. No. BC1122, HDL Cat. No. BC1124, and Total Protein Cat. No. BC1126).
Cecum Histology
2.6
On Day 42, two birds per treatment group were randomly selected and euthanized for histological evaluation of caecal tissues. Segments (approximately 2 cm) from the mid‐jejunum and cecum were excised, gently rinsed in phosphate‐buffered saline (PBS) and fixed in 10% neutral buffered formalin for 48 h. Fixed tissues were dehydrated through a graded ethanol series, cleared in xylene and embedded in paraffin wax. Sections of 5 µm thickness were cut using a rotary microtome and mounted on glass slides. The sections were stained with haematoxylin and eosin (H&E) for general histopathological examination under a light microscope (×40 magnification).
Statistical Analysis
2.7
Data were tested for normality and homogeneity of variance using the Shapiro–Wilk and Levene's tests, respectively. One‐way analysis of variance (ANOVA) was performed using SPSS (version 20.0; IBM, Armonk, NY, USA). When ANOVA indicated a significant treatment effect (p < 0.05), pairwise comparisons were performed using Tukey's Honestly Significant Difference (HSD) post‐hoc test. Data are presented as mean ± SEM. In tables, means within the same row bearing different superscript letters are significantly different (p < 0.05) according to Tukey's HSD.
Results
3
Growth Performance
3.1
Table 2 illustrates the impact of betaine supplementation in the broiler diet on FI. In the first week, FI exhibited a statistically significant increase (p < 0.05) in the B6 treatment compared to the B4 treatment. At Week 2, FI showed a statistically significant increase (p < 0.05) in the B6 treatment, B4 treatment and B2 treatment compared to the PC treatment and NC treatment. By Week 3, FI was significantly (p < 0.05) higher in the B6 treatment, B4 treatment and NC treatment in comparison to the B2 treatment and PC treatment.
During the starter period, FI demonstrated a statistically significant increase (p < 0.05) in the B6 treatment compared to the other treatments. At Week 4, FI exhibited a statistically significant increase (p < 0.05) in the NC treatment compared to the other treatments. In Week 5, FI was significantly (p < 0.05) higher in the NC treatment compared to the other treatments. By Week 6, FI was significantly (p < 0.05) higher in the NC treatment and B6 treatment compared to the other treatments. During the finisher period, FI demonstrated a statistically significant increase (p < 0.05) in the NC treatment compared to the other treatments. For the overall period, FI was significantly (p < 0.05) higher in the NC treatment and significantly (p < 0.05) lower in PC treatment of broilers.
Table 3 outlines the influence of betaine inclusion in the broiler diet on WG. In the initial week, WG displayed a statistically significant elevation (p < 0.05) in the B6 treatment, while being significantly reduced (p < 0.05) in both the NC and PC treatments. In the second week, WG showed a statistically significant increase (p < 0.05) in both the B6 and B4 treatments, with a concurrent significant decrease (p < 0.05) in the PC and NC treatments. By the third week, WG was significantly higher (p < 0.05) in the B6 treatment, while being significantly lower (p < 0.05) in both the NC and PC treatments.
During the starter phase, WG exhibited a statistically significant increase (p < 0.05) in the B6 treatment and significantly (p < 0.05) lower in PC treatment, and NC treatment. At Week 4, WG showed a statistically significant increase (p < 0.05) in the NC treatment compared to the rest of the treatment. In Week 5, WG was significantly (p < 0.05) higher in the NC treatment and significantly (p < 0.05) lower in PC treatment. By Week 6, WG exhibited a statistically significant increase (p < 0.05) in the B6 treatment compared to the NC treatment. During the finisher phase, WG demonstrated a statistically significant increase (p < 0.05) in the NC treatment and significantly (p < 0.05) lower in PC treatment. For the overall period, WG was significantly (p < 0.05) higher in both the NC treatment and B6 treatment and significantly (p < 0.05) lower in PC treatment.
Table 4 outlines the impact of incorporating betaine into the broiler diet on FCR. In the first week, FCR demonstrated a statistically significant decrease (p < 0.05) in the B6 treatment and significantly (p < 0.05) low in the NC treatment. During the second week, FCR exhibited a statistically significant reduction (p < 0.05) in both the B6 treatment and B4 treatment compared to the PC. By Week 3, FCR was significantly (p < 0.05) lower in the B6 treatment compared to the NC treatment.
In the starter phase, the FCR showed a statistically significant reduction (p < 0.05) in the B6 and B4 treatments, while significantly increasing (p < 0.05) in the NC and PC treatments. By Week 4, FCR displayed a statistically significant decrease (p < 0.05) in the B6 treatment and a significant increase (p < 0.05) in the PC treatment. In Week 5, FCR was significantly lower (p < 0.05) in the B6 treatment and significantly higher (p < 0.05) in the PC treatment. By Week 6, FCR exhibited a statistically significant decrease (p < 0.05) in the B6 treatment and a significant increase (p < 0.05) in the PC treatment. During the finisher phase, FCR demonstrated a statistically significant decrease (p < 0.05) in both the B6 and NC treatments compared to the PC treatment. For the overall period, FCR was significantly lower (p < 0.05) in the B6 treatment compared to the PC treatment.
Feed Digestibility
3.2
Table 5 summarizes the impact of incorporating betaine into the broiler diet on the percentage of ATD of nutrients. The ATD% of dry matter (DM) was significantly (*p < *0.05) higher in NC treatment and B6 treatment as compared to B2 treatment and B4 treatment followed by PC treatment. The ATD% of ash was significantly (*p *< 0.05) higher in NC treatment as compared to PC treatment with no differences in B2 treatment, B4 treatment and B6 treatment. The ATD% of crude protein was significantly (*p *< 0.05) higher in B6 treatment and NC treatment as compared to B2 treatment followed by PC treatment, but without any differences with B4 treatment. The ATD% of crude fat was significantly (*p *< 0.05) higher in NC treatment in comparison with B6 treatment followed by B4 treatment, B2 treatment and PC treatment.
Blood Biochemistry and Caecal Lesion Score
3.3
Table 6 and Figure 1, show the effect of inclusion of betaine in broilers diet on blood lipid profile and lesion score. TGs concentration was significantly (*p *< 0.05) higher in B4 and B6 treatment and significantly (p < 0.05) low in PC treatment. Total CHOL concentration was significantly (*p *< 0.05) higher in B4, B6 and NC treatments and significantly (p < 0.05) low in PC treatment. The LDL concentration was significantly (*p *< 0.05) higher in NC treatment in comparison with PC treatment. The HDL concentration was significantly (*p *< 0.05) lower in PC treatment compared to all other treatments. Total protein concentration was not significantly affected (p > 0.05) among all treatments. Lesion score reduced significantly (p < 0.05) in B4 and B6 compared to the PC treatment. No lesions were observed in the intestine or ceca of the unchallenged treatments.
NC showed normal caecal architecture with intact villi and no inflammation. PC exhibited severe necrosis, villus destruction, haemorrhage and heavy inflammatory infiltration. B2 showed moderate villus shortening, crypt hyperplasia and focal necrosis. B4 displayed mild to moderate lesions with improved villi and reduced inflammation. B6 revealed minimal changes, near‐normal villi, and mild inflammatory infiltration.
Discussion
4
Growth Performance
4.1
The results indicated that coccidial infection had a negative impact on the growth performance during the grower and finisher periods (Zhang et al. 2024). Growth performance, a crucial metric in the poultry industry, reflects feed utilization and efficiency (Rahman et al. 2017). Similarly, Batool et al. (2025) observed a significant reduction in the growth performance of broiler chicks challenged with Eimeria spp. Coccidiosis was found to hinder nutrient digestion and absorption, affecting the overall growth performance in broiler chicks (Ishaq et al. 2022). Previous studies have highlighted a connection between impaired performance due to coccidia challenge and a decrease in absorptive surface area, nutrient malabsorption and inflammation (Hafeez et al. 2023; Nasir et al. 2023). In our study, supplementation with 0.6% betaine effectively restored growth performance in challenged broilers. The observed decrease in WG among infected birds in our study seems to be attributed to both compromised FI and feed utilization. Birds experimentally infected with Eimeria spp. has suggested that diminished feed utilization contributes significantly to the decline in WG (Chand et al. 2021; Khan et al. 2023a; S. Khan et al. 2023b). Betaine functions as a methyl group donor (Kar et al. 2025). In conditions where diets are deficient in methionine, the inclusion of betaine serves to compensate for the role of methionine as a methyl donor, leading to improved performance (Alsayeqh and Rao 2023). This ability of betaine to serve as a methyl group donor may explain its positive impact on FCR in birds facing nutritional challenges. Moreover, under challenging conditions such as coccidia infection, there is an increased demand for methyl donors (Kar et al. 2025), highlighting the significance of betaine as a crucial methyl donor under such circumstances.
In this study, coccidia challenge led to notable reductions in the apparent ileal digestibility of DM, ash, crude protein and fat. Coccidiosis‐induced damage to the intestinal lining, particularly the shortening of villi, and decreased activities of digestive enzymes contribute to compromised nutrient digestion and absorption. The observed decline in fat digestibility during coccidia challenge may involve additional mechanisms, such as a reported reduction in bile salt secretion due to mucosal cell damage. Cholecystokinin, responsible for gallbladder contraction and pancreatic enzyme secretion, may be impacted by coccidia challenge. Notably, betaine supplementation demonstrated an improvement in digestibility in birds facing coccidia challenge. Previous research suggests that betaine may influence the crypt‐to‐villus height ratio, providing a potential explanation for the enhanced nutrient digestibility observed with betaine supplementation, as reported by Awad et al. (2022). Betaine, acting as an osmoprotectant, exerts effects on promoting intestinal cell growth, enhancing intestinal cell activity and improving intestinal morphology.
Blood Biochemistry
4.2
The current findings indicate that coccidiosis led to decreased serum concentrations of LDL‐C, TG, VLDL‐C, CHOL and HDL‐C. These findings are consistent with prior studies, which noted decreased plasma levels of TGs and elevated CHOL concentrations in broiler chickens infected with coccidia compared to a non‐infected group. Furthermore, Su et al. (2025) documented diminished serum levels of CHOL, TGs and HDL in coccidia‐infected broiler chicks compared to a control group. As demonstrated in the current study, the addition of dietary betaine resulted in a significant increase in serum TG and showed a tendency towards higher CHOL concentration at Day 42 of age. Yusuf et al. (2018) observed an increase in serum CHOL with dietary betaine supplementation (800 mg/kg) in broilers, while serum TG levels remained unchanged. Ghasemi and Nari (2020) reported a higher lipid profile with no change in blood protein in heat‐stressed broilers fed 1 g/kg betaine supplementation. The observed incremental effect of dietary betaine on serum TG in the current study is likely attributed to its role in mobilizing stored fat into the blood vessels for metabolism and oxidation in tissues.
Caecal Lesions
4.3
In the current study, caecal lesion score was restored in infected broilers received 400 and 600 mg/kg betaine. The protective impact of betaine in coccidial infection stems from its dual functionality as both an osmolyte and a methyl group donor (Kar et al. 2025). Acting as an osmolyte, betaine helps alleviate the osmotic stress experienced in the intestinal tract during coccidiosis (Alsayeqh and Rao 2023). Kar et al. (2025) found that chicks supplemented with betaine exhibited a significant reduction in the intracellular invasion of E. tenella or E. acervulina sporozoites in the intestines compared to control chicks. In its role as a methyl donor, betaine is thought to provide additional methyl groups, particularly to damaged tissues, which have higher requirements for these groups than healthy tissues (El‐Ghany and Babazadeh 2022).
Conclusion
5
Betaine supplementation at 600 mg/kg effectively mitigated the adverse effects of coccidiosis in broilers, improving growth performance, nutrient digestibility, blood lipid profiles and caecal lesion scores. However, these findings were obtained under controlled experimental conditions, which may not fully represent field circumstances where multiple stressors interact. Moreover, only one genetic line of broilers and a single species of Eimeria were tested, limiting the generalizability of the results. Future studies should explore the efficacy of betaine across different broiler strains, ages and production systems, and in combination with other phytogenic or nutritional additives. Long‐term field trials are also needed to validate its potential as a sustainable alternative to conventional anticoccidials.
Author Contributions
Abdul Hafeez: conceptualization. Usama Saleem: software; methodology; data curation. Shabana Naz: visualization; validation. Rifat Ullah Khan: writing – review and editing; writing – original draft. Muhammad Israr: writing – review and editing; writing – original draft. Ala Abudabos: writing – review and editing; writing – original draft. Ibrahim A. Alhidary: funding acquisition.
Funding
This study was funded by The Ongoing Research Funding (ORF‐2026‐833), King Saud University, Riyadh, Saudi Arabia.
Ethics Statement
The Committee on Animal Rights and Welfare, Faculty of Animal Husbandry & Veterinary Sciences, The University of Agriculture, Peshawar, Pakistan approved this study (FAH&VS/126 /2022).
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abbas, A. , Z. Manzoor , M. Seyidov , et al. 2025. “In vivo anticoccidial, growth promoting and biochemical effects of pinus radiata extract against experimental coccidiosis in broiler chickens.” Pakistan Veterinary Journal 45, no. 3: 1383–1388. 10.29261/pakvetj/2025.268. · doi ↗
- 2Aljohani, A. S. M. 2024. “Phenolics of Botanical Origin for the Control of Coccidiosis in Poultry.” Pakistan Veterinary Journal 44, no. 2: 222–228. 10.29261/pakvetj/2024.179. · doi ↗
- 3Almahallawi, R. , N. Al‐Hoshani , E. Al‐Nabati , et al. 2024. “Exploring the Anticoccidial, Growth‐Promoting, Hematological, and Serological Potential Activities of Linum usitatissimum Essential Oil in Broiler Birds.” Pakistan Veterinary Journal 44, no. 1: 117–122. 10.29261/pakvetj/2024.137. · doi ↗
- 4Alsayeqh, A. F. , and Z. A. Rao . 2023. “Nutritional Supplements for the Control of Avian Coccidiosis–A Review.” Annals of Animal Science 23, no. 4: 993–1007.
- 5Awad, W. A. , D. Ruhnau , A. Gavrău , K. Dublecz , and M. Hess . 2022. “Comparing Effects of Natural Betaine and Betaine Hydrochloride on Gut Physiology in Broiler Chickens.” Poultry Science 101, no. 12: 102173.10.1016/j.psj.2022.102173 PMC 957392936228528 · doi ↗ · pubmed ↗
- 6Batool, S. , M. A. Khan , S. Naz , et al. 2025. “Anticoccidial Effect of Bitter Apple (Citrullus colocynthis) Seed Powder and Organic Selenium Nanoparticles in Mitigating Eimeria tenella Induced Challenge in Quails.” Journal of Applied Animal Research 53, no. 1: 2526393. 10.1080/09712119.2025.2526393. · doi ↗
- 7Chand, N. , S. Naz , M. Irfan , R. U. Khan , and Z. ur Rehman . 2018. “Effect of Sea Buckthorn (Hippophae Rhamnoides L.) Seed Supplementation on Egg Quality and Cholesterol of Rhode Island Red x Fayoumi Laying Hens.” Korean Journal of Food Science of Animal Resources 38: 468–475.10.5851/kosfa.2018.38.3.468PMC 604837730018491 · doi ↗ · pubmed ↗
- 8Chand, N. , P. Ali , I. A. Alhidary , et al. 2021. “Protective Effect of Grape (Vitis vinifera) Seed Powder and Zinc‐Glycine Complex on Growth Traits and Gut Health of Broilers Following Eimeria tenella Challenge.” Antibiotics 10: 186. 10.3390/antibiotics 10020186.33672923 PMC 7918881 · doi ↗ · pubmed ↗