Species-specific alternative splicing of SP110 drives tuberculosis susceptibility in cattle
Ajiao Fan, Ying Guo, Zhunan Li, Xiangchen Dong, Zihan Zhang, Xinyu Wang, Yanliang Song, Jing Han, Jing Yang, Haoxin Wang, Xinyan Zhang, Yong Zhang, Yuanpeng Gao

TL;DR
Bovine tuberculosis susceptibility in dairy cattle is linked to a specific alternative splicing pattern of the SP110 gene that reduces its protective function.
Contribution
The study identifies species-specific alternative splicing of SP110 as a novel genetic mechanism driving tuberculosis susceptibility in cattle.
Findings
The truncated bSP110c variant, lacking the SAND domain, is predominantly expressed in cattle and is less effective against M. bovis.
The absence of a pre-SAND exon in ruminants promotes bSP110c production, while introducing human/equine sequences suppresses bSP110c.
This discovery highlights a potential gene editing target for breeding tuberculosis-resistant cattle.
Abstract
Bovine tuberculosis (bTB) is a chronic debilitating zoonotic disease caused by Mycobacterium bovis (M. bovis). M. bovis exhibits a broad host range, but dairy cattle are particularly susceptible. The SP110 (speckled protein 110) gene has been demonstrated to be associated with host susceptibility to TB, but whether the susceptibility of dairy cattle to M. bovis is correlated with the bovine SP110 (bSP110) gene has not been investigated. Here, we demonstrated that the heightened susceptibility of dairy cattle to bTB was driven by species-specific alternative splicing of the bovine SP110 (bSP110) gene, which resulted in predominant expression of a truncated bSP110c variant lacking the SAND domain required for anti-bTB function. Compared to the other two splice variants of bSP110 gene, bSP110a and bSP110b, bSP110c showing significantly reduced resistance to M. bovis. Mechanistically, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
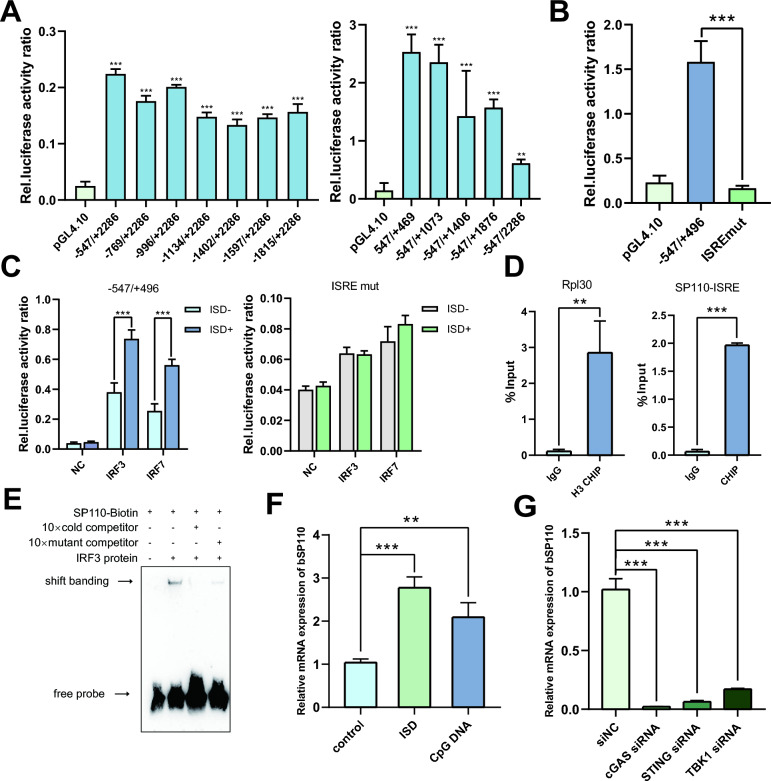 Figure 1
Figure 1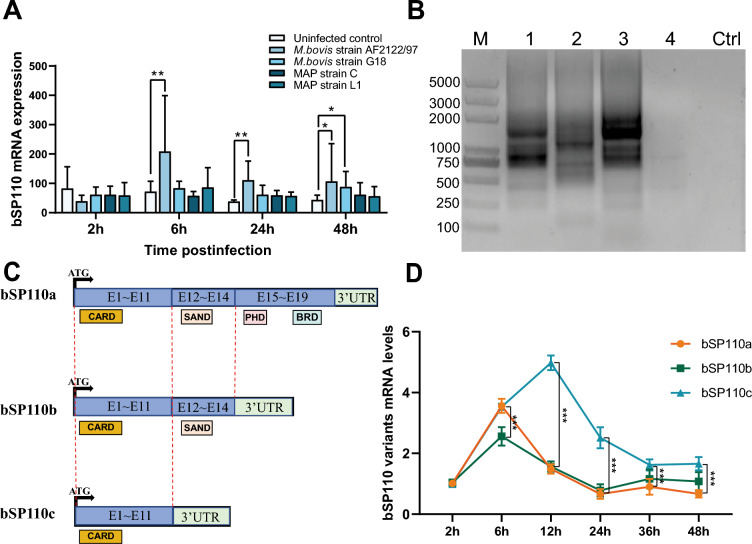 Figure 2
Figure 2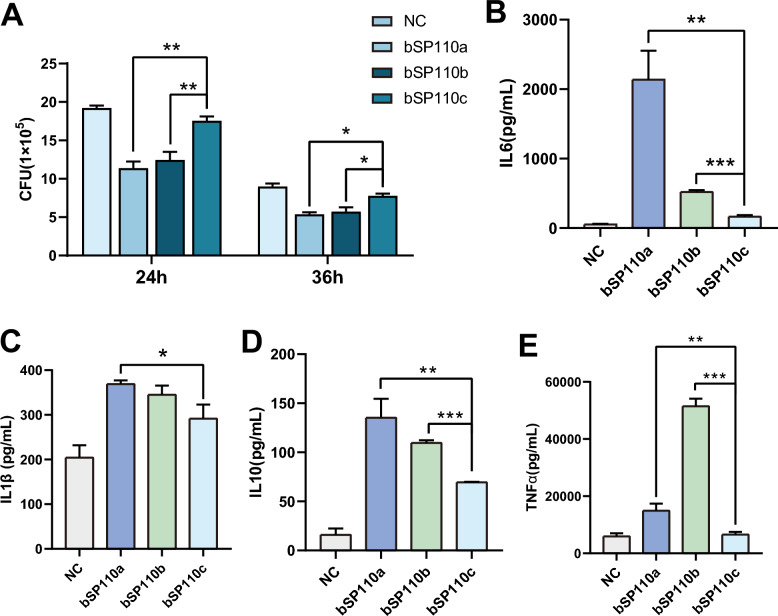 Figure 3
Figure 3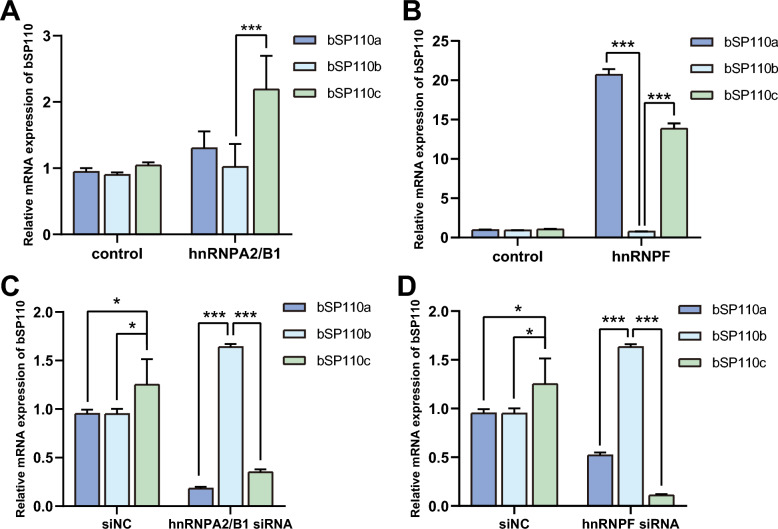 Figure 4
Figure 4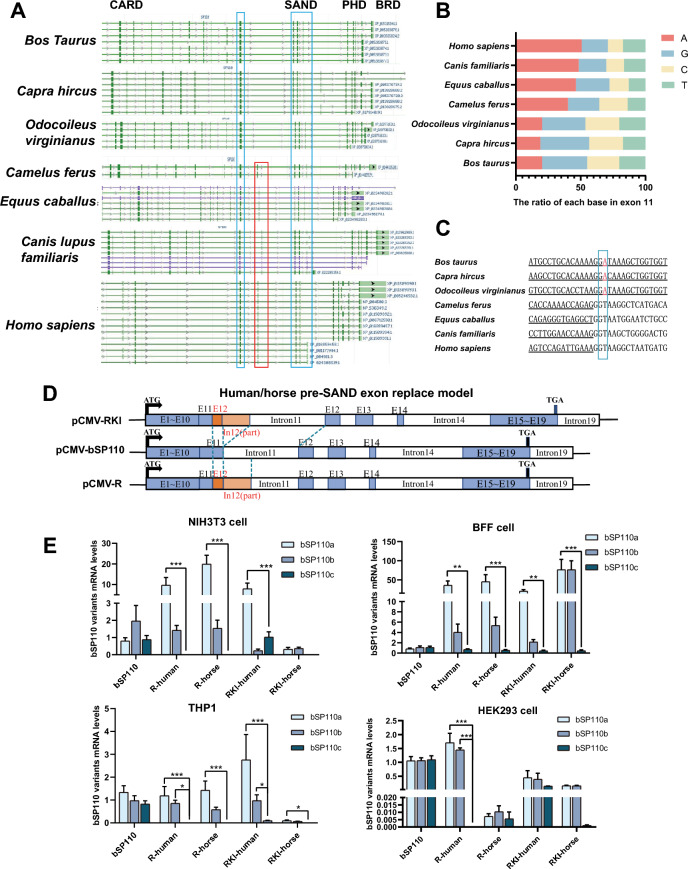 Figure 5
Figure 5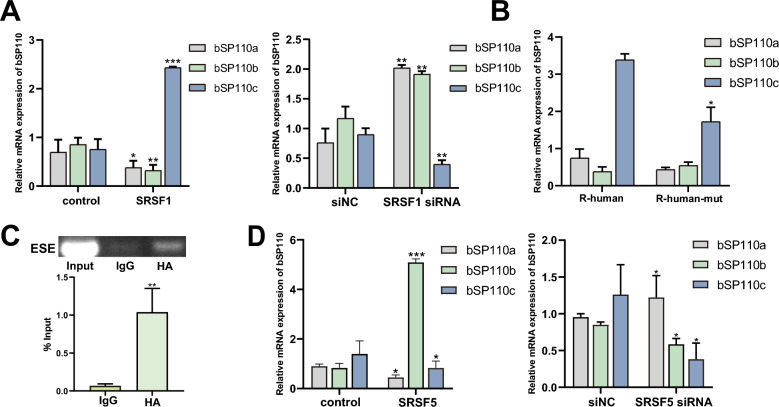 Figure 6
Figure 6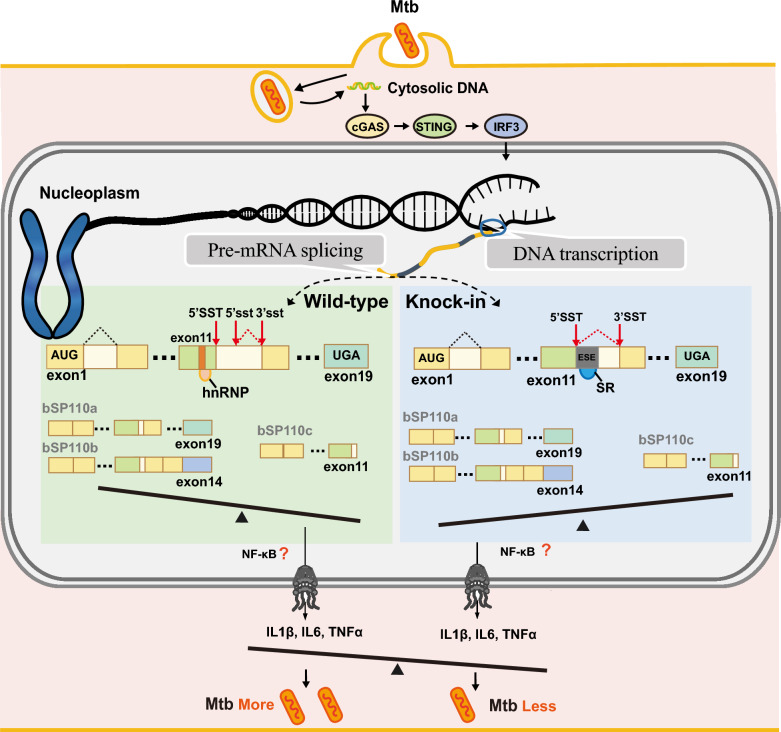 Figure 7
Figure 7- —https://doi.org/10.13039/501100018537National Science and Technology Major Project
- —https://doi.org/10.13039/501100001809National Natural Science Foundation of China
- —National Dairy Industry Technology Innovation Center Project Topics
- —Shaanxi provincial key project
- —https://doi.org/10.13039/501100013290National Key Research and Development Program of China Stem Cell and Translational Research
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTuberculosis Research and Epidemiology · Diagnosis and treatment of tuberculosis · Infectious Diseases and Tuberculosis
Introduction
Bovine tuberculosis (bTB) is a chronic debilitating zoonotic disease caused by Mycobacterium bovis (M. bovis) [1]. Worldwide, approximately 50 million cattle are infected with bTB annually, resulting in $3 billion in economic losses [2], with 10–15% of human tuberculosis cases attributable to M. bovis [3–5]. M. bovis is primarily transmitted through inhalation of respiratory aerosols and has the broadest host range within the Mycobacterium tuberculosis complex (MTBC), infecting cattle, humans, horses, and other species [6, 7]. However, dairy cattle show heightened susceptibility to M. bovis infection compared with other hosts [8–10]. Studies suggest that bTB susceptibility in dairy cattle is associated with specific genomic regions [11]. Identifying genes responsible for susceptibility to bTB in dairy cattle could enhance our understanding of the genetic mechanisms underlying the disease and potentially lead to breeding bTB-resistant dairy cattle [12].
The SP110 (speckled protein 110) gene has been shown to be associated with host susceptibility to TB [13–15]. Research indicates that hSP110B, a splice variant of the human SP110 gene, can interact with NF-κB to resist M. tuberculosis (MTB) infection [16]. The Ipr1 gene, located on mouse chromosome 1 in the super-susceptibility to TB 1 (sst1) locus, is the mouse homolog of SP110 [17, 18]. Site-specific knock-in of the murine Ipr1/SP110 gene mediated by the transcription activator-like effector (TALE) nickase can produce TB-resistant cattle. In vitro and in vivo challenge experiments proved that the transgenic cattle were able to inhibit the growth and multiplication of M. bovis [19]. These studies demonstrated that the SP110 gene plays an important role in TB resistance. However, the SP110 gene exhibits characteristic species-specific structural differences, and can generate splice variants with substantial functional divergence through alternative splicing mechanisms [20–22], potentially influencing the host's susceptibility to TB. The relationship between dairy cattle susceptibility to bTB and the bSP110 gene structure and splice variants in cattle remains to be determined.
To investigate the relationship between the bSP110 gene and dairy cattle susceptibility to bTB, we first characterized the splice variants of bSP110 and their anti-bTB functions. Our study identified three transcript variants of the bSP110 gene: bSP110a, bSP110b, and bSP110c. BSP110c has not been reported in other species, and it confers the weakest bTB resistance compared with bSP110a and bSP110b. The specific structure of the bSP110 gene results in the generation of the bSP110c splicing variant. Substitution of human or equine sequences upstream of the SAND exon (pre-SAND exon) in the corresponding position of the bSP110 gene reduced the expression of bSP110c and improved the resistance to M. bovis in cattle. Our study revealed the genetic factors of bovine susceptibility to M. bovis and provided a gene editing site for breeding bTB-resistant cows.
Materials and methods
Ethics statement
All animal experiments in this study were approved by the Animal Ethics Committee of Northwest A&F University (Approval No. 2021042) and strictly adhered to the “Guidelines for Animal Care and Use for Research Purposes”. The Holstein cattle blood plasma and healthy 3–6-month-old Holstein fetuses were sourced from Yangling Keyuan Co. Every effort was made to minimize animal pain, suffering, and distress, and reduce the number of animals used.
Cell culture and transfection
Hela, HEK293T, RAW264.7, and NIH3T3 cells were cultured in high-glucose DMEM supplemented with 10% fetal bovine serum (FBS), while THP-1 cells were maintained in RPMI 1640 with 10% FBS. Bovine fetal fibroblasts (BFFs) were isolated from 3- to 6 month-old Holstein fetuses (Yangling Keyuan Co.). Tissue from the back of the fetus was removed, minced with scalpels under aseptic conditions, plated on 60-mm Petri dishes (Corning Costar), and cultured in DMEM/F12 with 10% FBS. Peripheral blood mononuclear cells (PBMCs) were isolated from bovine plasma (Yangling Keyuan Co.) and induced to differentiate into bovine monocyte-derived macrophages (bMDMs) using granulocyte–macrophage colony-stimulating factor (GM-CSF). BMDMs were cultured in DMEM/F12 with 10% FBS. Plasmid transfections were performed using Lipofectamine 3000 (Invitrogen) according to the manufacturer's protocol.
3′RACE-PCR (rapid amplification of cDNA ends)
3′RACE primers were designed according to the predicted mRNA sequence of bSP110 in the NCBI database. Total RNA was extracted from bMDMs, reverse transcribed according to the instructions of the SMARTer cDNA amplification kit, followed by 3′RACE-PCR to amplify bSP110 splice variants. The primers used in 3′RACE are shown in Table 1.Table 1Primer sequences of 3′RACE PCR amplificationPrimer namePrimer sequenceSP110-3′RACE-NGSP-1GTCTCCCCTGCCGACAACCCCTACSP110-3′RACE-NGSP-2AAGGAAGGCTCAGTCCAGTGTCCAGSP110-3′RACE-NGSP-3CCTGAACCAAGCGACCCAAAGGASP110-3′RACE-NGSP-4ATGATGTCCCTGAACCAAGCGACCCA
Plasmid construction
Plasmids expressing bSP110 splice variants were constructed by cloning PCR-amplified bSP110a, bSP110b, and bSP110c fragments, which were obtained using primers based on the bSP110 splice variant sequences obtained by 3′RACE, into the pCMV-HA vector. Plasmids expressing IRF family members, STING, TBK1, hnRNPA2/B1, and hnRNPF CDS were constructed by amplifying the target sequences from bovine PBMCs, purifying the PCR products, and cloning them into the pCMV-Flag-N vector. Plasmids expressing SRSF1 and SRSF5 were constructed by PCR amplification from Hela cells followed by cloning into the pCMV-Flag-N vector. Plasmids containing the promoter region of the bSP110 gene were constructed by PCR amplification from bMDMs genomic DNA and subsequent cloning into the pGL4.10 luciferase reporter vector. The bSP110 minigene eukaryotic expression vector, containing exons 1–11 (without introns), partial intron 11, exons 12–14 (with introns), partial intron 14, exons 15–19 (without introns), and 3′UTR sequences, was cloned from the bMDMs genomic DNA into pCMV-Flag-N. To construct chimeric bSP110 mini-gene vectors with human or equine pre-SAND sequence substitutions, the human pre-SAND exon and its adjacent 5′ intronic sequence were PCR-amplified from Hela cell genomic DNA, while the equine pre-SAND exon and flanking 5′ intron were synthesized by Tsingke Biotechnology Co, Ltd. These isolated human or equine sequences were then used to replace the homologous region in the bSP110 mini-gene through homologous recombination. The primers used in plasmid construction are shown in Additional file 1 and Additional file 2.
Electrophoretic mobility shift assay (EMSA)
ISRE upstream and downstream sequences (20 bp) were selected as EMSA probes and the point-mutated ISRE probe sequences were synthesized, and labeled with biotin according to the instructions provided by Beyotime Co. The probes were incubated with the target proteins and EMSAs were performed using a chemiluminescence detection kit according to the instructions. The sequences of EMSA probes are shown in Table 2.Table 2Sequences of EMSA probesProbe nameProbe sequence (5′-3′)SP110-ISRE-FTCCGCAGGATCGGCCCGAGTACTTTCACTTTCACTTTCCTGGAAGCCAGGCCCSP110-ISRE-RGGGCCTGGCTTCCAGGAAAGTGAAAGTGAAAGTACTCGGGCCGATCCTGCGGASP110-ISREmut-Biotin-FTCCGCAGGATCGGCCCGAGTACCCTCATCTTCCTTTCCTGGAAGCCAGGCCC-BiotinSP110-ISREmut-Biotin-RGGGCCTGGCTTCCAGGAAAGGAAGATGAGGGTACTCGGGCCGATCCTGCGGA-BiotinSP110-ISRE-Biotin-FTCCGCAGGATCGGCCCGAGTACTTTCACTTTCACTTTCCTGGAAGCCAGGCCC-BiotinSP110-ISRE-Biotin-RGGGCCTGGCTTCCAGGAAAGTGAAAGTGAAAGTACTCGGGCCGATCCTGCGGA-Biotin
CHIP-qPCR (chromatin immunoprecipitation quantitative PCR)
ChIP assays were performed on human THP-1 cells after transfection with pCMV-Flag-N and pCMV-Flag-IRF3. ChIP DNA was detected by qPCR using primers from the promoter region of hSP110. The assays were performed according to the SimpleChIP^®^ enzymatic chromatin IP kit (CST) instructions. The primer sequences of ChIP-qPCR are shown in Table 3.Table 3Primer sequences of ChIP-qPCRPrimer namePrimer sequencehSP110-CHIP-qpcr-FACTTTCACTTTTCTTTTCTCGGAAGhSP110-CHIP-qpcr-RGGGACAGGGATCACTCCTCAAGATT
MTB culture and infection
M. tuberculosis strain MTB H37Ra was purchased from the American Type Culture Collection (ATCC, 25177), and M. bovis was obtained from the Guo Aizhen group at Huazhong Agricultural University. H37Ra was maintained in 7H9 medium, while M. bovis was cultured in 7H11 medium. Both media were supplemented with 10% oleic albumin dextrose catalase (Solarbio) and 0.5% glycerol. The required number of bacteria was pelleted by centrifugation at 4500 × g for 5 min, washed with phosphate-buffered saline (PBS), and resuspended in PBS for subsequent cell infection. All cells were infected at a multiplicity of infection (MOI) of 10. Post-infection, cells were incubated at 37 °C with 5% CO₂ for 4 h, washed three times with PBS to remove non-internalized mycobacteria, and 50 μg/mL of gentamicin was added to the culture medium for 1 h. Afterwards, the culture medium was replaced with fresh and the cell culture was continued for the specified time.
CFU assay for MTB
RAW264.7 cells were infected with M. bovis at an MOI of 10 for 24 h or 36 h, followed by washing three to five times in PBS and intracellular bacteria were released by cell lysis in PBS containing 0.25% Triton X-100 for 10 min at room temperature. The lysates were serially diluted in PBS and plated in triplicate on 7H11 agar plates. Agar plates were incubated at 37 °C for three weeks and colonies were counted.
Luciferase reporter assay
HeLa cells cultured in 24-well plates were co-transfected with the luciferase reporter vector (90 ng) and pRL-TK control vector (10 ng) using Lipofectamine 3000 (Invitrogen). At 24 h post-transfection, cells were harvested and lysed. The Dual-Luciferase Reporter Assay System (Promega) was employed to sequentially measure firefly and Renilla luciferase activities, with their ratio representing promoter activity. Three independent biological replicates were performed, each with technical duplicates.
Real-time quantitative PCR (RT-qPCR)
Total RNA was isolated using RNAiso Plus (Takara) and reverse-transcribed into cDNA using HiScript III RT SuperMix for qPCR according to the manufacturer’s instructions (Vazyme). ChamQ SYBR qPCR master mix (Vazyme) was used for quantitative real-time PCR analysis. The relative mRNA expression of target genes were normalized to GAPDH and calculated using the 2^−△△Ct^ method. The primers used in RT-qPCR are shown in Table 4.Table 4Primer sequences of qPCR amplificationPrimer namePrimer sequence (5′-3′)qPCR-Va-FCATTTTACAAGGCTTCCGACTTTGGqPCR-Va-RTTATATGGATCCTGAAGTTGGGATGqPCR-Vb-FGCTGCTGCAGAACGGAATTTTGTTCqPCR-Vb-RCACATGGTAACTCTGCAACGTGGTGqPCR-Vc-FCCGGGAAAAAGCGAGCAGCCTCATCqPCR-Vc-RTCACTTTAGATCTACATCACCTGTG
Cytokine ELISAs
RAW264.7 cells were infected with M. bovis at an MOI of 10 for 36 h. The culture supernatants were then collected, filtered through a 0.22 μm filter, and analyzed for mouse IL-1β, TNF-α, IL-10, and IL-6 levels using commercial ELISA kits (Proteintech) following the manufacturer's instructions.
Statistical analysis
The trials were conducted three times, and data are presented as the mean ± SD. Statistical significance was determined using Student’s t test, and P < 0.05 was deemed to be statistically significant.
Results
MTB infection activates bSP110 transcription via the cGAS- STING- IRF3 signaling pathway
Earlier studies showed that the transcript abundance of the bSP110 gene increased rapidly within 2–6 h after infection of bovine monocyte-derived macrophages (bMDMs) with M. bovis [23]. To identify the signaling pathways that activate bSP110 transcription in response to M. bovis infection, we cloned and characterized the bSP110 core promoter. The dual-luciferase reporter assay results demonstrated that the −547/ + 469 region of the bSP110 promoter exhibited the highest transcriptional activity, defining it as the core promoter of the bSP110 gene (Figure 1A). JASPAR database prediction revealed an interferon-stimulated response element (ISRE) within the bSP110 core promoter region. Mutation of this motif significantly impaired promoter-driven gene expression (Figure 1B).Figure 1MTB infection activates bSP110 transcription via the cGAS/STING/IRF3 pathway. A Activity of the truncated promoter, focusing on sequences upstream and downstream of the transcription start site (TSS). B The ISRE plays a key role in ISD induction and regulation of bSP110 transcription. C Effect of IRF3 and IRF7 on ISD induction and regulation of bSP110 transcription. D Enrichment of IRF3 in the promoter region of SP110 was detected by ChIP-qPCR enrichment of the Rpl30 gene of H3 relative to IgG. E IRF3 specifically binds to the SP110 promoter as detected by EMSA. F ISD and CpG DNA can effectively activate bSP110 transcription. G Effect of RNAi of cGAS/STING/TBK1 pathway protein on transcription of bSP110. Data were analyzed by t-test and presented as the mean ± SD of three independent experiments (*P < 0.05, **P < 0.01, ***P < 0.001).
To identify transcription factors binding to the ISRE, we performed JASPAR database and UCSC database association analyses and identified IRF3 and IRF7 as potential transcription factors with the ability to bind to the ISRE, with IRF3 being the most likely (Figure 1C). Electrophoretic mobility shift assay (EMSA) and chromatin immunoprecipitation-quantitative real time PCR (CHIP-qPCR) experiments showed that IRF3 bound to the ISRE in the promoter of bSP110 (Figures 1D, E), supporting the role of IRF3 in bSP110 transcription. The qPCR results showed that interferon-stimulated DNA (ISD) and cytosine-phosphate-guanine DNA (CpG DNA) of mycobacterial origin were able to activate bSP110 transcription (Figure 1F), which suggested that M. bovis infection might induce bSP110 transcription through the cGAS/STING/IRF3 intracellular DNA recognition pathway. To test this hypothesis, we performed RNA interference (RNAi) assays in bMDMs on STING and TANK-binding kinase-1 (TBK1). The results showed that bSP110 mRNA levels were significantly reduced by RNAi silencing of cGAS, STING and TBK1 (Figure 1G). In summary, our findings suggest that transcription of bSP110 can be activated through the cGAS/STING/IRF3 pathway.
Expression levels of the three splice variants of the bovine SP110 gene differ after MTB infection
Transcriptome analysis of NCBI GEO Datasets revealed significant activation of bSP110 transcription by M. bovis infection (Figure 2A). To characterize the splice variants generated following bSP110 transcription, we performed 3′RACE-PCR with sequence analysis of the amplified products and identified three splice variants of bSP110 designated as bSP110a, bSP110b and bSP110c (Figure 2B). Amino acid sequences of the three variants were predicted for conserved domains using the NCBI Conserved Domain Search tool, and the functional domains of the three variants are shown in Figure 2C. Notably, the C-terminal domains were progressively truncated across the variants, with a deletion of the core functional domain, SAND, from bSP110c. Temporal expression profiling in H37Ra-infected bMDMs demonstrated that bSP110c had significantly higher mRNA levels than bSP110a and b at 12 h and 24 h post-infection (Figure 2D).Figure 2Expression levels of the three splice variants of the bovine SP110 gene after MTB infection. A Detection of bSP110 mRNA levels in bovine monocyte-derived macrophages infected with M. bovis and M. avium subspecies paratuberculosis (MAP) GEO accession, GSE104211. B PCR results of 3′RACE; M, Trans 2 K Plus (II) DNA Marker; lanes 1–5, PCR results of different primers; Ctrl, control. C Schematic of the bSP110 domain. D Detection of bSP110 mRNA levels in bMDMs infected by H37Ra. Data were analyzed by t-test and presented as the mean ± SD of three independent experiments (*P < 0.05, **P < 0.01, ***P < 0.001).
Variants bSP110a and bSP110b show greater anti-bTB activity than bSP110c
To explore the bTB-resistance of the three bSP110 splice variants, we overexpressed bSP110a, bSP110b, and bSP110c in the mouse macrophage cell line, RAW264.7, and infected the cells with M. bovis. The CFU results showed reduced intracellular M. bovis infection in all three bSP110 splice variants, but transfection of bSP110a and bSP110b resulted in a significantly larger decrease in intracellular M. bovis compared to bSP110c at 24 h and 36 h after infection (Figure 3A). ELISAs demonstrated that all three variants, bSP110c, bSP110a and bSP110b, significantly promoted the expression of IL6, IL1β, IL10, and TNFα in macrophages after M. bovis infection (Figures 3B–E).Figure 3The variants, bSP110a** and bSP110b, showed greater anti-bTB activity than b**SP110c. A M. bovis CFU from RAW264.7 macrophages overexpressing bSP110a, b, or c after infection for 24 h and 36 h. B–E Expression of IL6, IL1β, IL10, and TNFα in RAW264.7 macrophages overexpressing bSP110a, b and c after M. bovis infection (detected by ELISA). Data were analyzed by t-test and presented as the mean ± SD of three independent experiments (*P < 0.05, **P < 0.01, ***P < 0.001).
The possible presence of ESS in exon 11 of the bovine SP110 gene could lead to the production of bSP110c
Exon splicing silencers (ESS) are often bound by hnRNPs, resulting in exon skipping [24]. To determine whether the production of bSP110c was a result of the presence of an ESS, we constructed a bSP110 minigene vector and overexpressed it with hnRNPA2/B1 and hnRNPF. The results showed that when hnRNPA2/B1 and hnRNPF were overexpressed, bSP110a and bSP110c mRNAs were increased, while bSP110b mRNA was decreased (Figures 4A, B). When hnRNPA2/B1 and hnRNPF expressions were silenced by RNAi, bSP110a and bSP110c mRNA levels were significantly decreased, while bSP110b mRNA was significantly upregulated (Figures 4C, D). The results showed that hnRNPA2/B1 and hnRNPF effectively inhibited the splicing of bSP110a and bSP110c but promoted the splicing of bSP110b, indicating that there may be an ESS binding to the splicing factors hnRNPA2/B1 and hnRNPF in exons 11 and 19 of bSP110, resulting in the formation of bSP110a and bSP110c.Figure 4An ESS was found in exon 11 of the bSP110 gene and resulted in the production of bSP110c. A Effect of hnRNPA2/B1 overexpression on mRNA levels of the three SP110 variants. B Effect of hnRNPF overexpression on mRNA levels of the SP110 variants. C Effect of hnRNPA2/B1 RNAi on mRNA levels of the SP110 variants. D Effect of hnRNPF RNAi on mRNA levels of the SP110 variants. Comparisons were analyzed for significant differences by t-test and presented as the mean ± SD of three independent experiments (*P < 0.05, **P < 0.01, ***P < 0.001).
Substitution of human or equine pre-SAND exon in the corresponding position of the bSP110 gene reduces the expression of bSP110c
We compared the DNA sequences upstream and downstream of the SAND domain exon of the SP110 gene in different species using NCBI and found that, compared with other species, the sequences immediately adjacent to the 5' end of the SAND exon of cows, sheep, and deer were missing (Figure 5A); this region, we designated as the ‘pre-SAND’ exon. The base composition of the pre-SAND exon of non-ruminant species showed typical A-rich features compared with the corresponding sequences of ruminants (Figure 5B). There was a point mutation in the splice recognition site (GU-AG) of the ruminant bSP110 intron 11 (Figure 5C). These results suggest that the ruminant SP110 gene has a unique genetic structure. To determine whether the generation of bSP110c was related to deletion of the pre-SAND exon, we constructed two vectors in which the corresponding position of the bSP110 minigene was replaced by the human and equine pre-SAND exon and the partial sequence of intron 12 and named them bSP110-Rhuman and bSP110-Rhorse. To ensure the retention of the splice element, we also constructed two vectors named bSP110-RKIhuman and bSP110-RKIhorse (Figure 5D). Quantitative PCR results showed that substitution of the human or equine pre-SAND exon sequence in the bovine SP110 gene upstream of the SAND exon effectively reduced expression of the bSP110c splice variant transcript (Figure 5E).Figure 5Replacing the human or horse pre-SAND exon at the corresponding position in the bovine SP110 **gene reduces b **SP110c expression. A Amino acid sequence alignment of SP110 from different species. The red box indicates the 5′-terminal pre-SAND exon adjacent to the SAND domain of the SP110 gene of non-ruminant species. The blue box indicates the homologous exon sequence at both ends of the pre-SAND exon. B Base composition comparison of corresponding sequences of the pre-SAND exon of different species. C Comparison of the intron splice recognition sites associated with the pre-SAND exon of different species. D Diagram showing the structure of the bSP110 minigene vector and the corresponding mutant vector. E Effect of substitution of the human or equine pre-SAND exon on SP110 splicing in different cell lines. Data were analyzed by t-test and presented as the mean ± SD of three independent experiments (*P < 0.05, **P < 0.01, ***P < 0.001).
An exon splicing enhancer (ESE) in the pre-SAND exon of the human and equine SP110 genes regulates alternative bSP110 splicing
To determine why substitution of the human or horse pre-SAND exon sequence in the bSP110 gene reduced bSP110c generation, we first performed exon splicing enhancer (ESE) analysis of the human and equine SP110 pre-SAND exon in ESE Finder software. Both human and equine pre-SAND exons were predicted to have ESE elements that can bind to serine/arginine-rich (SR) proteins (Additional file 3), we selected SRSF1 as a possible candidate for binding to the ESE of the pre-SAND exon of human SP110 and SRSF5 as a possible candidate for binding to the ESE of the equine pre-SAND exon. The results showed that overexpression of SRSF1 effectively promoted the production of bSP110c mRNA and reduced the production of bSP110a and bSP110b. In contrast, when cells were transfected with anti-SRSF1 siRNA, there was a significant decrease in bSP110c mRNA and a significant increase in bSP110a and bSP110b mRNA (Figure 6A). These results indicate that SRSF1 is involved in human SP110 splicing and affects the production of bSP110c. We mutated the ESE in the pre-SAND exon sequence of the human SP110 gene in bSP110-Rhuman and co-expressed it with SRSF1, and the results showed that mutation of the ESE down-regulated bSP110c (Figure 6B). RNA immunoprecipitation (RIP) results showed that SRSF1 specifically bound to this ESE (Figure 6C). We used the same method to determine the effect of splicing factor SRSF5 on the variable splicing of bSP110. The results showed that co-overexpression of SRSF5 and bSP110-Rhorse effectively promoted the production of bSP110b mRNA and reduced the mRNA levels of bSP110a and bSP110c. We then co-expressed anti-SRSF5 siRNA and the bSP110-Rhorse vector, the results were opposite to those of overexpression of SRSF1, with a significant increase in bSP110a and bSP110c mRNA, and a significant decrease in bSP110b mRNA (Figure 6D).Figure 6The ESE in the pre-SAND exons of the human and horse SP110 genes can regulate the alternative splicing of the bovine SP110 gene. A Effect of SRSF1 overexpression and RNAi on SP110 variant mRNA levels. B ESE site mutation can affect SRSF1 regulation of SP110 splicing. C Results of RNA immunoprecipitation. D Effect of SRSF5 overexpression and RNAi on SP110 variant mRNA levels. Data were analyzed by t-test and presented as the mean ± SD of three independent experiments (*P < 0.05, **P < 0.01, ***P < 0.001).
Discussion
Bovine tuberculosis (bTB) is classified by the World Organization for Animal Health as a notifiable animal disease [4, 5]. Although developed countries like the United Kingdom have reduced bTB incidence through long-term implementation of test-and-cull policies, the economic losses from culling may account for 30–50% of the disparity between the market value and slaughter value of dairy or beef cattle [25, 26]. In developing countries, the high prevalence of bTB, compounded by prohibitive control costs, poses significant threats to both animal-derived food safety and public health [27–29]. In addition, the control of bTB is made more difficult by the widespread presence of wildlife hosts, the lack of effective vaccines, and the evolution of MTB drug resistance [30–33]. Studies have shown that dairy cattle are highly susceptible to bTB and that this susceptibility is related to genetic traits [8–11]; therefore, identifying genetic factors of dairy cattle associated with bTB susceptibility and using gene editing to develop bTB-resistant cattle can be a cost-effective strategy for the control of bTB [34–37]. The focus of this research study was to investigate the genetic basis of the susceptibility of dairy cattle to bTB from the perspective of interspecies variation, and to identify potential gene-editing targets for breeding bTB-resistant cattle.
The SP110 gene has been proven to be associated with host susceptibility to TB. SP110 is a member of the speckled protein (SP) family, which contains three nuclear body components: SP100, SP110 and SP140 [38, 39]. SP family proteins are characterized by the presence of CARD, NLS, SAND, PHD and BRD domains [40], and the SAND domain is the core functional domain of SP110 that exerts the anti-TB function [16, 41, 42]. The SP110 gene shows complex alternative splicing [39], and the different splice variants have functional differences [21, 43, 44], affecting resistance to TB infection. Studies have indicated that SP110 gene structure differs among species [20], and there is evidence that the SP110 gene might have selectively evolved in ruminants [45, 46]. However, there has been little research on the bovine SP110 gene, with only a few reports demonstrating that bSP110 gene polymorphisms were associated with MTB susceptibility [47–49]. Our study is the first to examine the splice variants of the bovine SP110 gene. We found that the bSP110 gene can produce three splice variants with sequential deletions in C-terminal functional domains. The most abundant bovine splice variant was bSP110c, which lacks the core TB resistance domain SAND; SP110c has not been reported in other species.
To investigate the generation of bSP110c, we first analyzed the bSP110 gene structure. We found that intron 11 of bSP110 contained not only the canonical 5′ splice site (5′ SST) but also a cryptic 5′ splice site (5′ sst). Competition between these two splice sites can lead to aberrant splicing, generating bSP110c when the spliceosome recognizes 5′ sst. Our study also revealed that hnRNPs regulate bSP110c splicing, suggesting the presence of an exon splicing silencer (ESS) in exon 11 of bSP110 that binds hnRNPs leading to exon skipping and bSP110c formation. We also compared SP110 gene structures across species, and discovered that ruminants lack the pre-SAND exon. Replacing the corresponding region of the bSP110 gene with the pre-SAND exon sequences from humans or horses regulated alternative splicing, leading to a decrease in bSP110c and an increase in bSP110a and bSP110b expression. The results showed that both human and equine pre-SAND exons contained exon splicing enhancers (ESEs) capable of binding SR proteins. This ESE-SR protein interaction modulated alternative splicing of bSP110 (Figure 7).Figure 7Diagram of bSP110** splicing, illustrating how cloning the human or horse pre-SAND exon into the b**SP110 gene can alter variable splicing and enhance the anti-bTB capability of macrophages.
Although this research provides valuable insights, there are some limitations. We demonstrated that insertion of the pre-SAND exon can reduce the production of bSP110c and increase the production of bSP110a and bSP110b, but the effect of the pre-SAND exon on the resistance of macrophages to MTB infection requires further investigation with animal models, and results must be interpreted with caution. We observed that H37Ra, which is deficient in the ESX-1 system, can also significantly activate SP110 gene expression. We hypothesize that this may be associated with mitochondrial stress [41], but this needs to be experimentally tested.
Altogether, these findings increase our understanding of the genetic factors involved in the susceptibility of dairy cattle to M. bovis infection. Our study provides a gene-editing site for the potential breeding of bTB-resistant dairy cattle.
Supplementary Information
Additional file 1 Primer sequences of bSP110 gene PCR amplification. The primers used for constructing the bSP110 minigene eukaryotic expression vector.Additional file 2 Primer sequences of human and horse SP110 gene PCR amplification. The primers used to obtain the human or equine pre-SAND sequence.Additional file 3 Results of ESE analysis of human and horse SP110 pre-SAND exon. The results of the analysis performed using ESE Finder software, revealing ESE motifs within the human and equine SP110 pre-SAND exon that can bind to SR proteins.