Molecular mechanisms of disinfectant resistance in Klebsiella pneumoniae
Daniel J Noel, Alistair Bailey, Benjamin I Nicholas, Paul Skipp, C William Keevil, Sandra A Wilks

TL;DR
This study explores how Klebsiella pneumoniae develops resistance to common disinfectants, revealing molecular mechanisms that could help improve infection control strategies.
Contribution
The study identifies specific molecular adaptations in Klebsiella pneumoniae that confer resistance to multiple disinfectants, including cross-resistance mechanisms.
Findings
Adaptation to BAC, DDAC, and PHMB involves lipid A modification, reducing the bacterial surface's negative charge and disinfectant affinity.
Chlorocresol resistance is linked to increased efflux pump activity and biofilm formation.
Bronopol resistance involves biofilm promotion and thioredoxin upregulation, with NemA potentially degrading bronopol.
Abstract
Chemical disinfectants are critical for infection control in healthcare environments and beyond, as exemplified by their vital role during the COVID-19 pandemic. Despite research repeatedly demonstrating that bacteria can develop adaptations that mitigate the efficacy of chemical disinfectants, the underlying molecular mechanisms remain poorly characterized. This study investigates the mechanisms that underpin resistance demonstrated by disinfectant-adapted Klebsiella pneumoniae NCTC 13443 samples. Resistant samples have previously undergone long-term in vitro adaptation via serial passage in increasing concentrations of common disinfectants benzalkonium chloride (BAC), didecydimethylammonium chloride (DDAC), polyhexamethylene biguanide (PHMB), chlorocresol or bronopol. A multi-omics approach was used to conduct in-depth molecular analyses of the adaptations that contribute to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
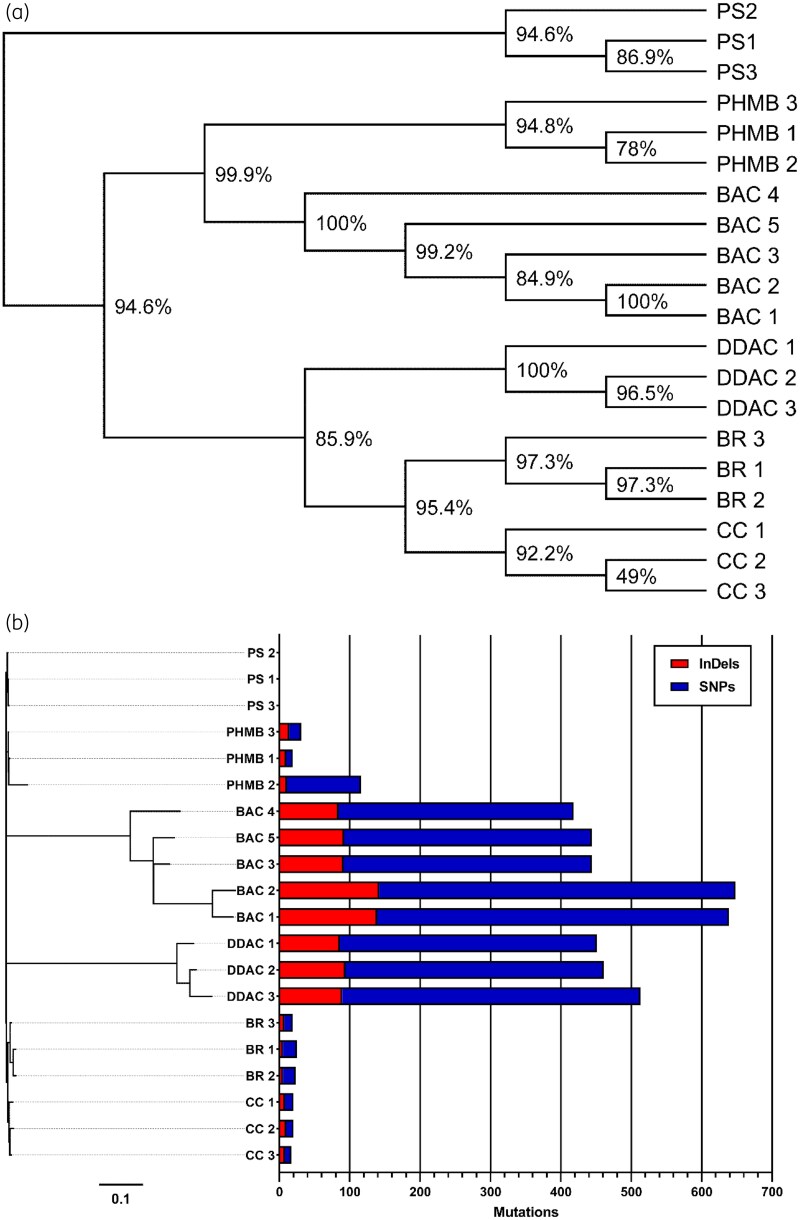 Figure 1
Figure 1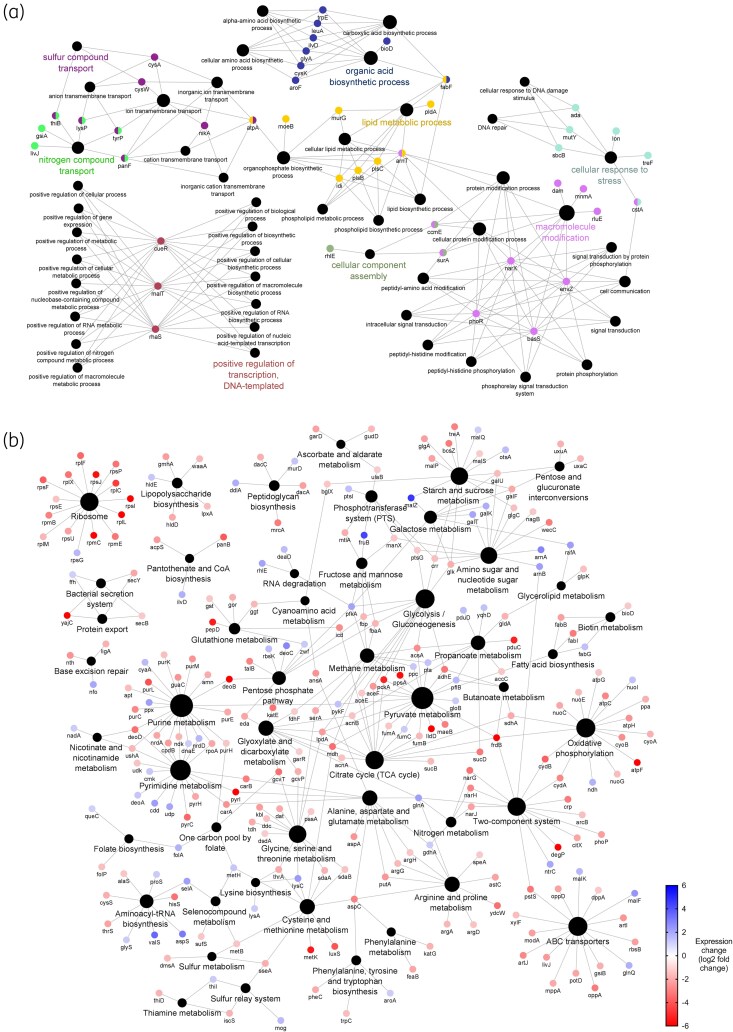 Figure 2
Figure 2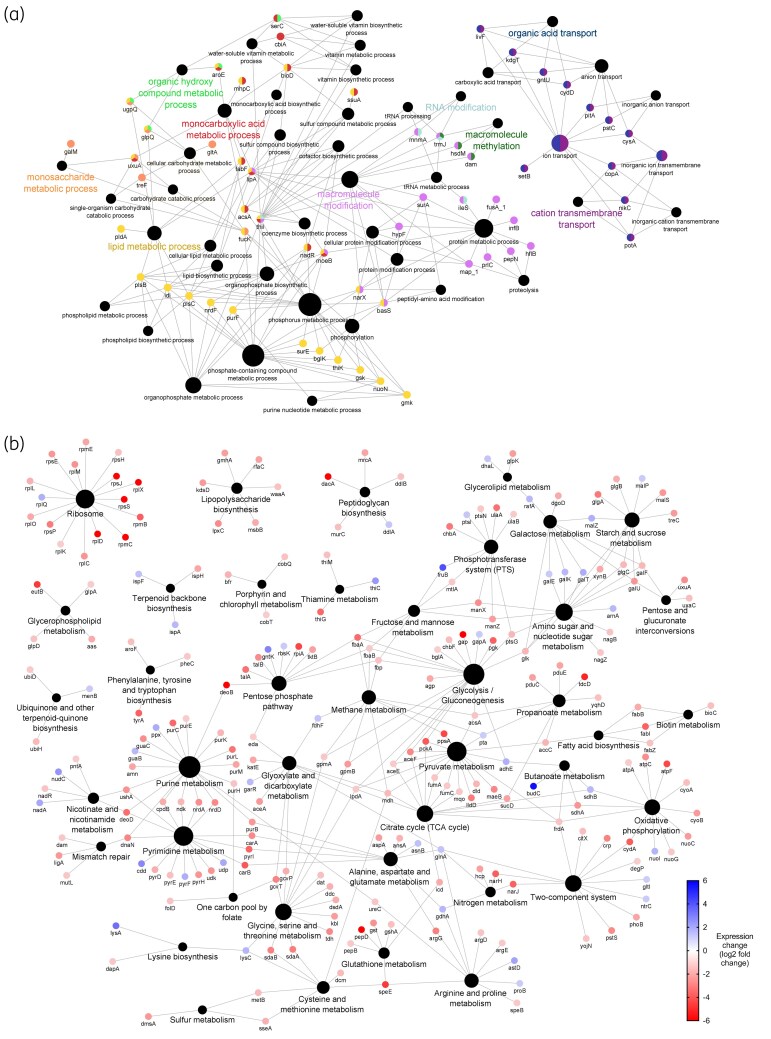 Figure 3
Figure 3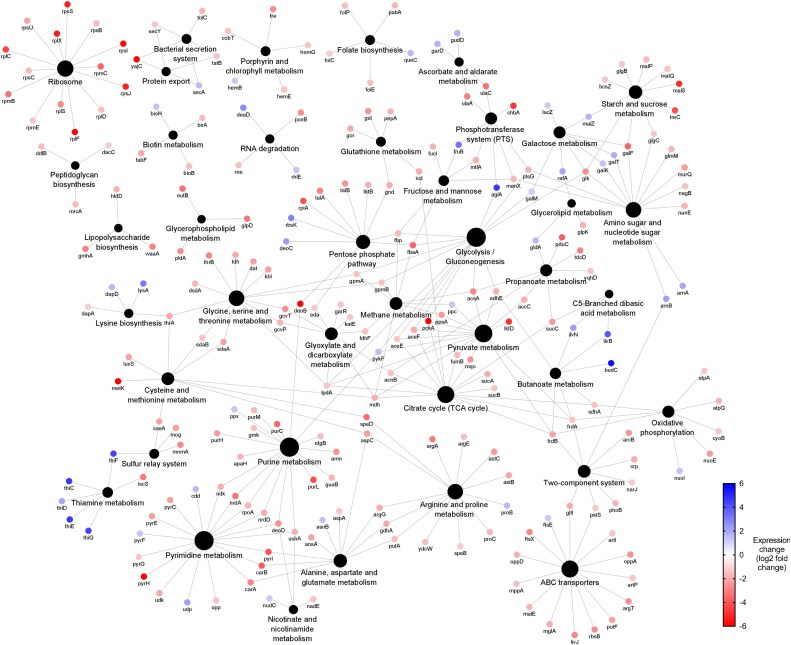 Figure 4
Figure 4 Figure 5
Figure 5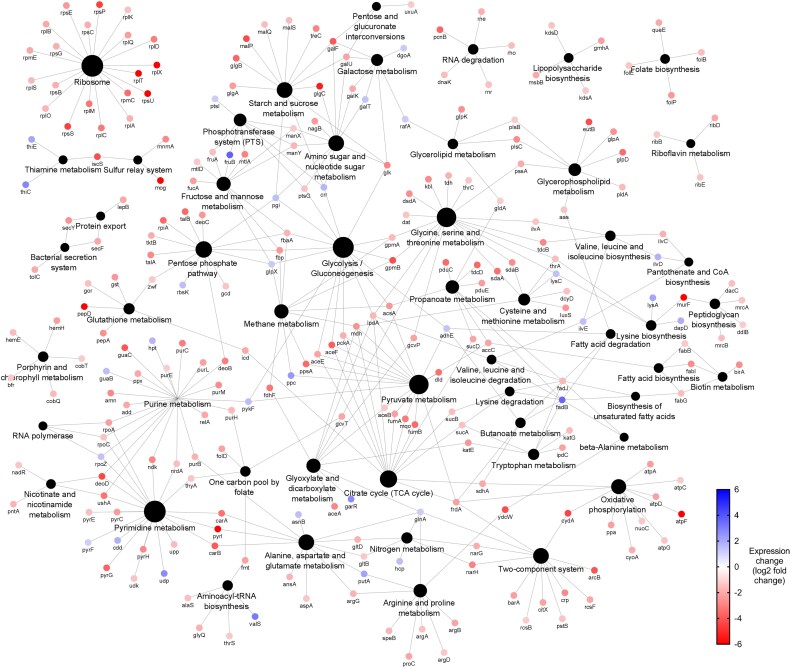 Figure 6
Figure 6| Compound | Cellular target | Antimicrobial mechanism | Applications |
|---|---|---|---|
| BAC | Membrane | Positively charged quaternary nitrogen groups interact with anionic lipids, promoting their own cellular uptake. This interaction facilitates the insertion of hydrophobic tails into the lipid bilayer, disrupting lipid organization and compromising membrane integrity. As a result, low molecular weight substances leak out, the proton motive force is lost, and oxidative phosphorylation becomes uncoupled. | Surface disinfection sprays and wipes, eye/ear drops, burn treatments. |
| DDAC | Membrane | Positively charged quaternary nitrogen groups interact with anionic lipids, promoting their own cellular uptake. This interaction facilitates the insertion of hydrophobic tails into the lipid bilayer, disrupting lipid organization and compromising membrane integrity. As a result, low molecular weight substances leak out, the proton motive force is lost, and oxidative phosphorylation becomes uncoupled. | Surface disinfection sprays and wipes, sterilization of surgical equipment. |
| PHMB | Membrane | The biguanide group sequesters anionic lipids, forming homogenous lipid domains that disturb the structural organization of the membrane. This disruption increases membrane permeability and causes leakage of intracellular content. | Surface disinfection sprays and wipes, wound dressings, contact lens cleaning solution, swimming pool cleaners. |
| Chlorocresol | Membrane | Compromises the permeability barrier through membrane disruption, inducing leakage of low molecular weight components. Causes downstream collapse of proton motive force and uncoupling of oxidative phosphorylation. | Antiseptic, preservative. |
| Bronopol | Proteins. | Catalyses oxidation of thiols to disulphides, cross-linking proteins and impeding functionality. This reaction also generates ROS that cause downstream damage to intracellular components. | Disinfectant, preservative. |
| Disinfectant |
| |
|---|---|---|
| Pre-adaptation | Post-adaptation | |
| BAC | 20 | 56 |
| DDAC | 6 | 14 |
| PHMB | 6 | 9 |
| Chlorocresol | 200 | 260 |
| Bronopol | 8 | 41 |
| Resistant sample | Gene | Variation type | Gene annotation | Protein annotation | Protein product |
|---|---|---|---|---|---|
| PHMB |
| ns-SNP | 469A>C | T157P | Sensor protein BasS/PmrB |
|
| ns-SNP | 374A>G | D125G | Transcriptional regulator KdgR | |
| Chlorocresol |
| fs-ins | 78dupT | T27N | DNA-binding transcriptional repressor MarR |
|
| fs-ins | 290_291insC | M97I | Protein YicC | |
|
| ns-SNP | 163A>C | I55L | RND efflux system | |
|
| ns-SNP | 136A>G | S46G | 3′,5′-cyclic-nucleotide phosphodiesterase | |
|
| ns-SNP | 289G>A | D97N | Fumarate and nitrate reduction regulatory protein | |
|
| ns-SNP | 2233A>C | S745R | Fimbriae usher protein StcC | |
| NCTC13443_06216 | ns-SNP | 104T>G | V35G | Fimbrial-like protein | |
| NCTC13443_06725 | ns-SNP | 1690G>T | A564S | Membrane protein | |
|
| ns-SNP | 742C>T | P248S | Cyclic-guanylate-specific phosphodiesterase | |
|
| nfs-del | 110_121del | 37_41del | Fimbrial protein MrkD | |
| Bronopol |
| ns-SNP | 136A>G | S46G | 3′,5′-cyclic-nucleotide phosphodiesterase |
|
| ns-SNP | 2233A>C | S745R | Fimbriae usher protein StcC | |
| NCTC13443_06216 | ns-SNP | 104T>G | V35G | Fimbrial-like protein | |
|
| ns-SNP | 413GT | R138L | Purine nucleotide synthesis repressor | |
|
| ns-SNP | 664A>C | T222P | PutAP proline dehydrogenase transcriptional repressor | |
|
| ns-SNP | 314G>C | R105P | Negative transcriptional regulator of cel operon | |
|
| ns-SNP | 742C>T | P248S | Cyclic-guanylate-specific phosphodiesterase | |
|
| nfs-del | 110_121del | 37_41del | Fimbrial protein MrkD |
| Resistant samples | Expression change (log2 fold change) | Protein identifier | Full protein name |
|---|---|---|---|
| BAC | 5.64 | BudB | Acetolactate synthase |
| 5.64 | ArnA | Bifunctional polymyxin resistance protein ArnA | |
| 5.64 | YfdX | YfdX-like protein | |
| 5.06 | NCTC13443_01223 | Thioredoxin-like protein | |
| 4.64 | MalZ | Maltodextrin glucosidase | |
| 4.32 | FruB | Multiphosphoryl transfer protein | |
| 3.84 | FruB | Multiphosphoryl transfer protein | |
| 3.64 | AcrB | Efflux pump membrane transporter AcrB | |
| 3.64 | FrlD | Fructosamine kinase FrlD | |
| 3.47 | AcrA | Efflux pump membrane transporter AcrA | |
| DDAC | 5.64 | YfdX | YfdX-like protein |
| 5.64 | GlpK | Glycerol kinase | |
| 5.64 | BudC | Diacetyl reductase [(S)-acetoin forming] | |
| 4.64 | NCTC13443_03659 | Putative NADH:flavin oxidoreductase | |
| 4.06 | FruB | Multiphosphoryl transfer protein | |
| 4.06 | ValS | Valine–tRNA ligase | |
| 3.84 | ScrY | Sucrose porin | |
| 3.32 | DmlA | D-malate dehydrogenase (decarboxylating) | |
| 3.32 | LysA | Diaminopimelate decarboxylase | |
| 3.18 | AldB | Alpha-acetolactate decarboxylase | |
| PHMB | 5.64 | GlpK | Glycerol kinase |
| 5.64 | NCTC13443_03659 | Putative NADH:flavin oxidoreductase | |
| 5.64 | YfdX | YfdX-like protein | |
| 5.64 | NCTC13443_02382 | Putative L-fucose isomerase, C-terminal | |
| 5.64 | NCTC13443_02379 | Putative L-fucose isomerase, C-terminal | |
| 5.64 | BudC | Diacetyl reductase [( | |
| 5.06 | NCTC13443_01223 | Thioredoxin-like protein | |
| 5.06 | SacA | Sucrose-6-phosphate hydrolase | |
| 4.64 | ThiC | Phosphomethylpyrimidine synthase | |
| 4.64 | NCTC13443_02381 | Putative L-fucose isomerase, C-terminal | |
| Chlorocresol | 5.64 | YebE | Inner membrane protein YebE |
| 5.64 | MdtC | Multidrug resistance protein MdtC | |
| 5.06 | YfdX | YfdX-like protein | |
| 5.06 | BudC | Diacetyl reductase [( | |
| 4.64 | AldB | Alpha-acetolactate decarboxylase | |
| 4.64 | BepD | Multidrug resistance protein MdtA | |
| 4.64 | UspG | Universal stress protein G | |
| 4.32 | NCTC13443_03659 | Putative NADH:flavin oxidoreductase | |
| 4.06 | HutU | Urocanate hydratase | |
| 3.84 | FdhF | Formate dehydrogenase | |
| Bronopol | 5.64 | BudB | Acetolactate synthase |
| 5.64 | YfdX | YfdX-like protein | |
| 5.64 | DDJ638005 | — | |
| 4.32 | NCTC13443_03659 | Putative NADH:flavin oxidoreductase | |
| 4.06 | NCTC13443_01223 | Thioredoxin-like protein | |
| 3.64 | FruB | Multiphosphoryl transfer protein | |
| 3.47 | FrlD | Fructokinase | |
| 3.47 | FadB | Fatty acid oxidation complex subunit alpha | |
| 3.32 | NemA |
| |
| 3.32 | DkgB | 2,5-didehydrogluconate reductase DkgB |
- —NIHR Southampton Biomedical Research Centre10.13039/501100022419
- —JVS Products Ltd
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial agents and applications · Antibiotic Resistance in Bacteria · Infection Control in Healthcare
Introduction
Healthcare-associated infections (HAIs) cause hundreds of millions of infections worldwide every year. Klebsiella pneumoniae is a common causative pathogen of HAIs, with Klebsiella spp. accounting for ∼10% of cases in US hospitals.^1^
The COVID-19 pandemic highlighted our reliance on chemical disinfectants for infection control; a dependence likely to grow with the ever-increasing prevalence of antimicrobial resistance (AMR). While AMR is typically associated with antibiotic resistance, evidence shows bacteria can also develop tolerance and resistance towards disinfectants.^2^ This has resulted in calls for antiseptic stewardship^3^ and contributed to the banning of triclosan and other antimicrobials in the USA. Benzalkonium chloride (BAC) and five other antiseptics are currently under review, partly due to concerns surrounding antiseptic resistance and cross-tolerance.^4^
For clarity, tolerance is defined as the ability of an organism to survive transient exposure to otherwise lethal concentrations of disinfectant, while resistance is as an inherited ability of an organism to survive and grow at otherwise lethal concentrations irrespective of exposure time.^5^
Clinical K. pneumoniae samples have shown varying susceptibilities to chlorhexidine,^6^ iodophor^6^ and BAC,^7^ while in vitro experiments have demonstrated K. pneumoniae tolerance to chlorhexidine,^8^ BAC^7^ and polyhexamethylene biguanide (PHMB) in combination with betaine.^9^ We recently demonstrated the ability for K. pneumoniae samples to adapt to otherwise lethal concentrations of quaternary ammonium compounds (QACs) BAC and didecyldimethylammonium chloride (DDAC), the cationic polymer PHMB, the phenol-derivative chlorocresol and the reactive-oxygen species (ROS)-producing bronopol through evolutionary adaptation.^10^ Their characteristics are presented in Table 1, and the respective pre- and post-adaptation MICs in Table 2. These samples were provisionally characterized as disinfectant tolerant, as genotypic adaptations have not been confirmed. Collateral susceptibility was more common that cross-tolerance among these samples, even among disinfectants with similar mechanisms of action (MOA). This is presumably due to the fitness cost of adaptation.
BAC tolerance in K. pneumoniae has been attributed to increased efflux pump activity.^7^ However, chemical inhibition of efflux pump activity had no impact on BAC and PHMB susceptibility of K. pneumoniae isolates,^15^ suggesting other contributing mechanisms. In other species, porin down-regulation and membrane charge alteration via lipid A modification has been associated with BAC tolerance.^16^ Tolerance to PHMB and bronopol has not been investigated in detail, so any mechanisms of tolerance remain unknown. Chlorocresol tolerance has not been reported or investigated, although studies have shown a link between phenolic disinfectant susceptibility and efflux pump activity.^17^
This study aims to characterize the molecular mechanisms enabling these disinfectant-adapted K. pneumoniae samples to survive otherwise lethal disinfectant concentrations via whole-genome sequencing and label-free quantitative proteomics. Addressing these aims will provide valuable insights into how bacteria can adapt to commonly used disinfectants in healthcare, commercial and household environments, while also guiding future decisions on healthcare cleaning routines and antiseptic stewardship policies.
Materials and methods
Bacterial strains and growth media
Whole-genome sequencing and proteomic analysis was performed on three biological replicates of disinfectant-adapted K. pneumoniae samples generated previously.^10^ Owing to minor colour variations between colonies of BAC-adapted replicates plated on CHROMagar^™^ Orientation chromogenic agar, five samples were analysed to check for technical consistency. For a comprehensive description of the adaptation methodology and initial characterization of the samples, see Noel et al.^10^ In brief, samples were passaged daily in Mueller–Hinton broth containing increasing concentrations of disinfectant until no further tolerance developed over 15 consecutive passages.^10^ Pre- and post-adaptation MICs are displayed in Table 2. Total adaptation times varied from 69 to 103 passages.^10^
Adapted samples were cultured overnight at 37°C in Mueller–Hinton broth containing a sub-MIC (post-adaptation) of respective disinfectant (55 mg/L BAC, 13 mg/L DDAC, 8 mg/L PHMB, 240 mg/L chlorocresol, 40 mg/L bronopol)^10^ before protein and DNA extraction. For comparison, three pre-adaptation samples were prepared in the absence of disinfectant.
Stock solutions of antimicrobial compounds
Here, 10 000 mg/L BAC, DDAC, PHMB and bronopol (Thor Specialities) stocks were prepared in ddH_2_O immediately before use. Chlorocresol (Lanxess) was prepared similarly in undiluted DMSO. Working concentrations of DMSO had no detectable impact on K. pneumoniae colony forming unit counts.
Whole-genome sequencing
One millilitre aliquots were washed in PBS three times before DNA extraction via DNeasy PowerSoil Pro Kit (Qiagen) following the manufacturer’s instructions. Lysates were frozen at −20°C until required. Samples were sequenced by Novogene via Illumina^®^ NovaSeq^™^ 6000.
Raw reads were cleaned to remove adapter contamination, reads with >10% uncertain bases or >50% low quality nucleotides, before mapping to the reference genome with BWA. Variants were called using GATK, compared using bcftools and annotated using the ANNOVAR software tool.
Global quantitative proteomics
Samples were washed three times in PBS, pelleted and resuspended in lysis buffer [50 mM tris, 150 mM NaCl, 0.1% w/v SDS, cOmplete^™^ protease inhibitor cocktail (Roche)]. After sonication (120 s total, 12% amplitude, 10-s pulses) and centrifugation (12 000g, 20 min, 4°C), protein concentrations were quantified via BCA assay. Lysates were frozen at −20°C until required.
Volumes containing 100 µg of protein were mixed with 600 µL of methanol, 150 µL of chloroform, 450 µL of dH_2_O and subsequently centrifuged (14 000g, 10 min). After the upper aqueous layer was removed, 450 µL of methanol was added before mixing and centrifugation. Protein pellets were air dried before resuspension in 100 µL of 6 M urea, 50 mM tris-HCl, 5 mM dithiothreitol (pH 8.0), incubated for 30 min at 37°C, before incubation with 15 mM iodoacetamide for 30 min at room temperature. Four micrograms trypsin/Lys-C mix (Promega) were added for 4 hours at 37°C, before dilution in 750 µL of 50 mM tris-HCl (pH 8.0) and overnight incubation. Digestion was terminated by the addition of trifluoroacetic acid before centrifugation (14 000g, 10 min). Peptides were purified using Oasis PRiME HLB 96-well µElution plates (Waters) by elution in 70% acetonitrile and spin-dried under vacuum.
Samples were resuspended in 50 µL of 0.1% v/v formic acid before mass spectrometry (UltiMate 3000 RSLC nano system with Orbitrap Fusion^™^ Tribrid^™^ Mass Spectrometer, Thermo Fisher Scientific).
Peptide/protein identification and area under the curve quantification were performed using PEAKS Studio Xpro (Bioinformatics Solutions). Proteome coverage was 25.6%–28.3%. Proteins were filtered to include proteins identified across all parent and adapted replicates. A 1% false discovery rate and minimum ±2 log_2_ fold change threshold were used to define significant, differentially expressed proteins.
Data analysis
Phylogenetic trees were generated using CSI Phylogeny v.1.4^18^ using default parameters and K. pneumoniae NCTC 13443 as the reference genome. FigTree v.1.4.4^19^ was used for visualization.
Genetic variants were filtered to exclude synonymous single nucleotide polymorphisms (SNPs), non-coding mutations and mutations not conserved across all replicates. Gene Ontology (GO) biological function networks were generated using ClueGO v.2.5.9^20^ in Cytoscape v.3.9.1^21^ with K. pneumoniae strain 342 as the reference genome. Small variant sets are instead presented in Table 3.
GO enrichment of differentially expressed proteins was performed using the Database for Annotation, Visualization and Integrated Discovery (DAVID)^22,23^ with K. pneumoniae MGH 78578 as the background list. Enriched biological process and cellular component GO terms with a P value of ≤0.05 were considered significant and visualized as heatmaps via GraphPad Prism v.9.4.1. Network maps of differentially expressed proteins were generated using ClueGO^20^/Cytoscape^21^ and arranged by Kyoto Encyclopedia of Genes and Genomes (KEGG) biological pathways annotations, with K. pneumoniae strain 342 used as reference.
Results and discussion
Characterization of disinfectant-adapted K. pneumoniae samples
Unrooted phylogenetic trees (Figure 1) group samples into clades by disinfectant treatment, showing that adapted samples have acquired conserved genotypic adaptations facilitating survival in the presence of otherwise lethal concentrations of disinfectant (Table 2) irrespective of exposure time.^10^ We therefore classify the samples as resistant, although only to concentrations of disinfectants significantly lower (Table 2) than those at point of use in commercial products, which are typically in the 10^2^–10^3^ mg/L range.^24,25^ Despite this, it is still pertinent to establish what mechanisms are being used to mitigate the efficacy of disinfectants, especially considering how various factors including dilution factor,^26^ organic load,^27^ exposure time^28^ and residual compound degradation^29^ can effectively reduce the exposure concentration.
Genetic characterization of Klebsiella pneumoniae NCTC 13443 disinfectant-adapted samples. (a) Unrooted, unscaled phylogenetic tree resistant samples. Node values indicate bootstrap values as a percentage. (b) The total number of mutations acquired by Klebsiella pneumoniae NCTC 13443 disinfectant-adapted samples compared to the respective untreated parent samples. The phylogram shows the relative genetic relationships of the samples, with the distance scale indicating the number of nucleotide substitutions per site. InDels, insertions or deletions that are ≤50 base pairs in length. PS: parent samples. BR, bronopol-adapted samples; CC, chlorocresol-adapted samples.
Disinfectant resistant samples displayed short genetic distances from each other, except for QAC-adapted (BAC, DDAC) samples (Figure 1b). Genetic distances between adapted clusters did not align with MOA similarities, with BAC- and DDAC-adapted clades being furthest apart despite both being cationic membrane-active agents with a near-identical general MOA. This suggests small differences in interactions with components of the membrane can manifest distinct adaptations. Although sharing few mutations in common, BAC-adapted samples previously demonstrated cross-resistance to DDAC^10^ implying overlapping mechanisms.
QAC-adapted samples demonstrated a uniquely high number of mutations (Figure 1b), indicating a strong selection pressure. Conversely, PHMB, chlorocresol and bronopol-adapted samples showed fewer mutations and shorter genetic distances between them, despite more varied MOAs. This shows that few mutations are required for K. pneumoniae to adapt to these agents, and suggests that the strength of the selection pressure had more influence on the genotype than MOA similarities.
PHMB-adapted sample 2 accumulated many unique mutations compared with other replicates, indicating inconsistent resistance mechanisms (Figure 1b). PHMB-adapted samples also showed varying cross-resistance profiles,^10^ thought to be due to multiple suggested target sites. While PHMB is primarily regarded as membrane-active, research suggests alternative action via condensing of nucleic acids.^12^ Multiple target sites would allow for different resistance strategies, explaining the lack of homogeneity between the biological replicates. Despite this, all PHMB-adapted replicates remained grouped in a single clade, indicating conserved adaptations.
Quantitative proteomics showed lower expression across all adapted samples (Figures 2b, 3b, 4, 5, 6), with reduced enrichment of arginine biosynthesis, gluconeogenesis, translation and TCA cycle biological process GO terms (Figure S1, available as Supplementary data at JAC-AMR Online), indicating reduced growth and metabolism. This is characteristic of dormant phenotypes, which classically show reduced antimicrobial susceptibility.^30,31^ This also demonstrates the fitness cost of adaptation, with energy being diverted to maintain a resistant phenotype.
Genomic and proteomic characterization of Klebsiella pneumoniae NCTC 13443 benzalkonium chloride-adapted samples. (a) Network diagram of genes that contain conserved mutations across all replicates (n = 5). Genes are indicated by the coloured dots, arranged according to GO biological process annotation. Colours indicate biological process annotation. Black dots indicate biological process annotations, as labelled. Lines connect genes to their annotations. (b) Network diagram of differentially expressed proteins (coloured dots), arranged according to KEGG pathway annotation. Node labels show associated gene name. Blue and red coloration indicates increased or reduced expression, respectively. Black dots indicate KEGG pathway annotations, with size proportional to number of associated differentially expressed proteins. Lines connect genes to their annotations.
Genomic and proteomic characterization of Klebsiella pneumoniae NCTC 13443 didecyldimethylammonium chloride-adapted samples. (a) Network diagram of genes that contain conserved mutations across all replicates (n = 3). Genes are indicated by the coloured dots, arranged according to GO biological process annotation. Colours indicate biological process annotation. Black dots indicate biological process annotations, as labelled. Lines connect genes to their annotations. (b) Network diagram of differentially expressed proteins (coloured dots), arranged according to KEGG pathway annotation. Node labels show associated gene name. Blue and red coloration indicates increased or reduced expression, respectively. Black dots indicate KEGG pathway annotations, with size proportional to number of associated differentially expressed proteins. Lines connect genes to their annotations.
Network diagram of differentially expressed proteins in polyhexamethylene biguanide-adapted Klebsiella pneumoniae NCTC 13443 samples. Differentially expressed proteins are indicated by the coloured dots, arranged according to KEGG pathway annotation. Blue and red coloration indicates up or down expression, respectively. Black dots indicate KEGG pathway annotations, as labelled. Lines connect proteins to their annotations. This network map was generated by Cytoscape v.3.9.1 using the ClueGO v.2.5.9 plugin.
Network diagram of differentially expressed proteins in chlorocresol-adapted Klebsiella pneumoniae NCTC 13443 samples. Differentially expressed proteins are indicated by the coloured dots, arranged according to KEGG pathway annotation. Blue and red coloration indicates up or down expression, respectively. Black dots indicate KEGG pathway annotations, as labelled. Lines connect proteins to their annotations. This network map was generated by Cytoscape v.3.9.1 using the ClueGO v.2.5.9 plugin.
Network diagram of differentially expressed proteins in bronopol-adapted Klebsiella pneumoniae NCTC 13443 samples. Differentially expressed proteins are indicated by the coloured dots, arranged according to KEGG pathway annotation. Blue and red coloration indicates up or down expression, respectively. Black dots indicate KEGG pathway annotations, as labelled. Lines connect proteins to their annotations. This network map was generated by Cytoscape v.3.9.1 using the ClueGO v.2.5.9 plugin.
K. pneumoniae resistance to QACs
Mutated genes in QAC-adapted samples were linked to lipid metabolic and macromolecule modification cellular processes (Figures 2a and 3a). Both BAC and DDAC-adapted samples contained a conserved non-synonymous SNP in histidine kinase basS (polymyxin resistance protein B, PmrB), which positively regulates arnABCDEFT genes responsible for 4-amino-4-deoxy-L-arabinose (L-Ara4N) synthesis. ArnT modifies lipid A with L-Ara4N, neutralizing the negative charge of the 4′-phosphate group and decreasing the net-negative charge of the outer leaflet.^32^ This reduces the affinity of cationic peptides including colistin and polymyxin B, facilitating resistance.^32^
The mutation causes a A68V substitution within the transmembrane region (Tables S1 and S2), responsible for physiological signal detection and conformational changes activate arnABCDEFT genes.^33^ Similar mutations in this region constitutively activate E. coli PmrB and cause increased expression of arnT.^34^ Proteomics reveals increased expression of downstream ArnAB in BAC-adapted samples (Figure 2), and ArnA in DDAC-adapted samples (Figure 3), suggesting the A68V substitution constitutively activates BasS.
BAC-adapted samples contained a conserved N457S substitution in arnT (Table S1) in transmembrane domain helix 13 adjacent to the lipid A binding cavity, specifically the polar region that associates with 3-deoxy-D-manno-oct-2-ulosonic acid sugars.^35^ Asparagine and serine are polar uncharged residues, so this relatively subtle substitution may alter lipid A binding affinity and contribute to BAC resistance.
BAC-adapted samples also displayed a conserved V187M substitution in the linker region of histidine kinase envZ. This protein regulates OmpF/C expression via the activation of OmpR in response to changes in osmolarity. No downstream proteins were detected via proteomics due to protein coverage limitations, so the impact remains unclear. The absence of this mutation in DDAC-adapted samples suggests it is unique to BAC.
Both BAC and DDAC-adapted samples contained mutations in DNA repair genes ada and mutY, while BAC also had a mutation in sbcB. DDAC-adapted samples showed decreased expression of methyl-directed mismatch repair system protein MutL. Loss of DNA repair function contributes to hypermutable phenotypes,^36^ allowing for greater stochastic development of beneficial mutations and explaining the large number of QAC-associated mutations.
Regarding efflux pumps, all QAC-adapted samples carried an E130K substitution in AcrB, the inner membrane component of multidrug efflux pump AcrAB-TolC. Located in the PN1 subdomain of the porter domain where both proximal and distal binding pockets are located,^37^ similar substitutions can alter substrate specificity,^38^ resulting in AMR.^39^ DDAC-adapted samples also had conserved substitutions in resistance-nodulation-division family efflux pumps MdtC and BepE, plus a F81S substitution proximal to the ligand binding site of AcrAB regulator AcrR.^40^ BAC-adapted samples showed increased AcrAB and BepE expression, further reinforcing the importance of efflux pumps in QAC resistance.
DDAC resistance appeared to require additional efflux pump modifications, probably reflecting necessary modifications to substrate specificity. This explains the lack of reciprocation in cross-resistance profiles between BAC and DDAC-adapted samples.^10^
Collectively, K. pneumoniae QAC resistance occurs via modification of lipid A with L-Ara4N, lowering cell surface negative charge and reducing affinity of cationic QACs. As this is a well-established mechanism facilitating resistance to cationic peptides including polymyxin B and colistin, it is highly likely that this mechanism confers cross-resistance. QAC resistance is also associated with efflux pump activity, with DDAC requiring additional efflux modifications compared with BAC. Adaptation is further supported by reduced DNA repair functionality, giving rise to a hypermutable K. pneumoniae phenotype.
K. pneumoniae resistance to PHMB
Similar to QAC-adaptation, molecular analysis of PHMB-adapted K. pneumoniae showed links to lipid A modification. PHMB-adapted K. pneumoniae samples displayed only two conserved mutations across all biological replicates, in basS and kdgR (Figure 4, Table 3, Table S3). The BasS adaptation causes T157P in the dimerization and histidine phosphotransferase domain, responsible for phosphotransferase, phosphatase and autokinase activities. As this domain modulates protein activity, and expression of downstream ArnA and ArnB was increased (Figure 4), this mutation probably increases activity or constitutively activates BasS.
Lipid A modification was also seen in BAC- and DDAC-adapted samples and is known to underpin polymyxin and colistin resistance.^41^ As this exact mutation has previously been attributed with colistin resistance,^41^ it probably provides cross-resistance between PHMB and colistin. Collectively, these data suggest that increased L-Ara4N modification of lipid A is a common mechanism for resistance to cationic antimicrobials in general, raising questions regarding cross-resistance between cationic disinfectants in healthcare and last-resort antibiotics such as polymyxins.
PHMB-adapted samples also showed a conserved D125G substitution in 2-keto-3-deoxygluconate (KDG) regulon repressor KdgR, which regulates KdgATK proteins responsible for KDG transportation and catabolism. Interestingly, a SNP causing D11G in KdgR was also found in all BAC-adapted samples (Table S1). Downstream expression of eda (KdgA) was lower in PHMB and BAC-tolerant samples, suggesting increased KdgR repressor activity. The exact impact this has on BAC and PHMB resistance is unclear.
K. pneumoniae resistance to chlorocresol
Chlorocresol-adapted samples contained conserved mutations in cpdA and yjcC (Table 3), which regulate intracellular cAMP and c-di-GMP levels, respectively. The mutations resulted in S46G in CpdA and P248S in YjcC.
yjcC (synonymous pdeC) encodes a phosphodiesterase that hydrolyses c-di-GMP when dimerized,^42^ negatively regulating type 3 fimbriae expression and biofilm formation.^43^ The observed substitution is in the second transmembrane region, responsible for dimerization and protein activation.^42^ We hypothesize the substitution of this conserved proline^42^ impedes dimerization, increasing intracellular c-di-GMP, enhancing biofilm formation and type 3 fimbriae expression.
CpdA hydrolyses cAMP to 5′-adenosine monophosphate (AMP). Deletion of this protein causes intracellular cAMP to increase up to 4-fold,^44^ promoting biofilm formation in K. pneumoniae via type 3 fimbriae production.^45^ The adaptations may therefore increase biofilm formation, probably relating to mutations in fimbriae-related genes htrE, fim_1 and NCTC13443_06216 (Table 3), which shares sequence homology with type 3 fimbria minor subunit MrkB.
Also conserved was a frameshift insertion in marR, truncating the marRAB operon repressor (T27N.fsX3) (Table 3). This protein is responsible for AMR-related changes in expression including down-regulation of OmpF, increased expression of AcrAB-TolC and resistance to oxidative stress via transcriptional activator SoxS.^46^ MarR truncation probably increases expression of AcrAB-TolC and SoxS, contributing to chlorocresol resistance through efflux and activation of the superoxide response regulon. However, downstream changes could not be confirmed by proteomics due to protein coverage limitations.
Interestingly, loss of MarR function has been linked to increased WaaY expression, responsible for phosphorylation of the inner core of LPS, increasing the net-negative charge of the bacterial outer surface and susceptibility to antimicrobial peptides.^47^ Therefore, truncation of MarR probably contributes to the collateral susceptibility of chlorocresol-adapted samples to BAC and DDAC observed previously,^10^ and potentially polymyxin B.
Chlorocresol-adapted samples also showed increased expression of the MdtC and BepD (MdtA) proteins (Figure 5, Table 4), all components of tripartite efflux complex MdtABC. This complex is responsible for resistance to novobiocin and detergent deoxycholate via TolC-dependent efflux,^48^ and is associated with resistance to multiple antibiotics in K. pneumoniae clinical strains.^49^ Therefore, this probably contributes to chlorocresol resistance.
It should be noted that sub-inhibitory concentrations of DMSO have been shown to alter gene expression,^50^ mitigate ROS damage^50^ and reduce biofilm formation in P. aeruginosa.^51^ As such, the specific impact of the presence of low DMSO concentrations on the observed adaptations remain unclear, particularly those associated with increased biofilm formation.
K. pneumoniae resistance to bronopol
Bronopol-adapted samples carried conserved SNPs in transcriptional regulators putA, rhaS and purR (Table 3). putA and rhaS_2 encode for regulators of the put and cel operons, respectively. PutA oxidizes proline and acts as an auto repressor of putA and putP. Increased PutA expression indicates the mutation impedes the repressor functionality.
The conserved R138L substitution in the purine biosynthesis (pur) operon repressor PurR is proximal to the E. coli PurR corepressor binding site. Nearby mutations can broaden corepressor binding specificity and enhance repression activity.^52^ As purBCEHLM were all significantly down-regulated (Figure 6), the mutation likely enhances PurR-mediated repression. This response has also been observed in E. coli exposed to oxidative and antibiotic stressors,^53^ although the specific mechanisms are unknown. As bronopol induces oxidative stress through the generation of ROS, these data support the hypothesis that PurR has a key regulatory role associated with oxidative stress response.
Bronopol-adapted samples also displayed conserved mutations in the secondary-messenger regulators yjcC and cpdA, alongside in fimbriae-associated htrE, fim_1 and NCTC13443_06216 genes (Table 3). This suggests resistance through enhanced biofilm formation as seen with chlorocresol. Similarly, oxidative-stress-inducing hypochlorite and tellurite are associated with increased intracellular c-di-GMP levels in P. aeruginosa,^54,55^ alongside increased diguanylate cyclase activity,^54,55^ surface attachment^54^ and biofilm formation.^55^ This collectively highlights a link between intracellular c-di-GMP concentration, biofilm formation and bacterial resistance to oxidative stress-inducing antimicrobials such as bronopol.
Bronopol-adapted samples showed increased expression of thioredoxin-like protein NCTC13443_01223 and flavin oxidoreductase-like protein NCTC13443_03659 (Table 4). Thioredoxin proteins can reduce the disulphide bonds formed by the MOA of bronopol, mitigating bronopol-induced oxidative stress in adapted K. pneumoniae samples. As flavin oxidoreductase knockouts demonstrate high susceptibility to oxidative stress in E. coli^56^ and Streptococcus pneumoniae,^57^ increased expression probably has the opposite effect.
The flavin-dependant N-ethylmaleimide reductase NemA was among the top proteins up-regulated in bronopol-resistant samples (Table 4). This protein has previously been shown to be capable of breaking down electrophiles^58^ including 24,6-trinitrotoluene (TNT)^59^ in E. coli. As bronopol and TNT both contain nitro electrophilic groups, the significant up-regulation of NemA suggests a potential role in bronopol resistance via enzymatic degradation. The requirement of flavin cofactor explains the associated up-regulation of NADH:flavin oxidoreductase. The reduced expression of catalases KatE, KatG and superoxide dismutase SodB (Figure 6) supports the hypothesis that NemA can break down bronopol before it is able to form ROS.
Conclusion
Chemical disinfectants are relied on worldwide across healthcare, industrial settings, the food sector and household environments. Understanding mechanisms that mitigate their efficacy is critical to combating the hundreds of millions of annual HAI cases. This molecular analysis of K. pneumoniae adaptation to common disinfectants provides novel insights into potential disinfectant resistance mechanisms of HAI-associated pathogens. The findings highlight similarities between mechanisms facilitating resistance to cationic antibiotics and disinfectants, raising questions about the risk of cross-resistance that can be expanded on in future work. Genetic variation between BAC and DDAC-adapted samples show how minor differences between similar agents can manifest distinct adaptations, as demonstrated by conserved efflux pump adaptations in DDAC-adapted samples that are not necessary for BAC resistance.
Chlorocresol and bronopol resistance has not been investigated previously. K. pneumoniae resistance to chlorocresol was associated with marR loss of function, increased MdtABC efflux complex expression and promoting biofilm formation, a mechanism shared with bronopol resistance alongside mitigation of cross-linking damage. Increased expression of N-ethylmaleimide reductase NemA may facilitate bronopol resistance via enzymatic degradation.
Collateral susceptibility of chlorocresol-resistant K. pneumoniae to QACs and cationic agents via MarR truncation highlights how adaptations to one antimicrobial often leaves organisms vulnerable to others, knowledge that can be used to improve efficacy of cleaning routines and infection control.
Future studies should validate the mechanisms implicated in this work. The impacts of QAC and PHMB adaptations associated with lipid A modification can be assessed via zeta potential analysis and plasmid-based complementation, while the extent of QAC-polymyxin cross-resistance can be confirmed through antimicrobial susceptibility assays. The role of NemA in bronopol resistance can be investigated through enzymatic activity assays.
This study identifies molecular mechanisms of disinfectant resistance in K. pneumoniae samples generated via stepwise adaptation, deepening our understanding of the potential routes that HAI pathogens can exploit to mitigate disinfectant efficacy.
Supplementary Material
dlaf247_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Magill SS, O’Leary E, Janelle SJ et al Changes in prevalence of health care-associated infections in U.S. hospitals. N Engl J Med 2018; 379: 1732–44. 10.1056/NEJ Moa 180155030380384 PMC 7978499 · doi ↗ · pubmed ↗
- 2Maillard JY, Pascoe M. Disinfectants and antiseptics: mechanisms of action and resistance. Nat Rev Microbiol 2024; 22: 4–17. 10.1038/s 41579-023-00958-337648789 · doi ↗ · pubmed ↗
- 3Maillard JY, Kampf G, Cooper R. Antimicrobial stewardship of antiseptics that are pertinent to wounds: the need for a united approach. JAC Antimicrob Resist 2021; 3: dlab 027. 10.1093/jacamr/dlab 02734223101 PMC 8209993 · doi ↗ · pubmed ↗
- 4U.S. Food and Drug Administration . FDA Response to ACI July 2020 Progress Report (ethanol bzk pmcx bzec pi). Document ID: FDA-2015-N-0101-1493. November 18, 2020. Available at: https://www.regulations.gov/document/FDA-2015-N-0101-1493.
- 5Krewing M, Mönch E, Bolten A et al Resistance or tolerance? Highlighting the need for precise terminology in the field of disinfection. J Hosp Infect 2024; 150: 51–60. 10.1016/j.jhin.2024.05.00638782058 · doi ↗ · pubmed ↗
- 6Guo W, Shan K, Xu B et al Determining the resistance of carbapenem- resistant Klebsiella pneumoniae to common disinfectants and elucidating the underlying resistance mechanisms. Pathog Glob Health 2015; 109: 184–92. 10.1179/2047773215 Y.000000002226184804 PMC 4530556 · doi ↗ · pubmed ↗
- 7Abdelaziz A, Sonbol F, Elbanna T et al Exposure to sublethal concentrations of benzalkonium chloride induces antimicrobial resistance and cellular changes in Klebsiellae pneumoniae clinical isolates. Microb Drug Resist 2019; 25: 631–8. 10.1089/mdr.2018.023530614757 · doi ↗ · pubmed ↗
- 8Bock LJ, Wand ME, Sutton JM. Varying activity of chlorhexidine-based disinfectants against Klebsiella pneumoniae clinical isolates and adapted strains. J Hosp Infect 2016; 93: 42–8. 10.1016/j.jhin.2015.12.01926899354 · doi ↗ · pubmed ↗