Two new saprobic species of Helicosporium and Dictyosporium and a new host record of Neohelicomyces guizhouensis isolated from Phellodendron amurense (Rutaceae, Tracheophyta) in China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
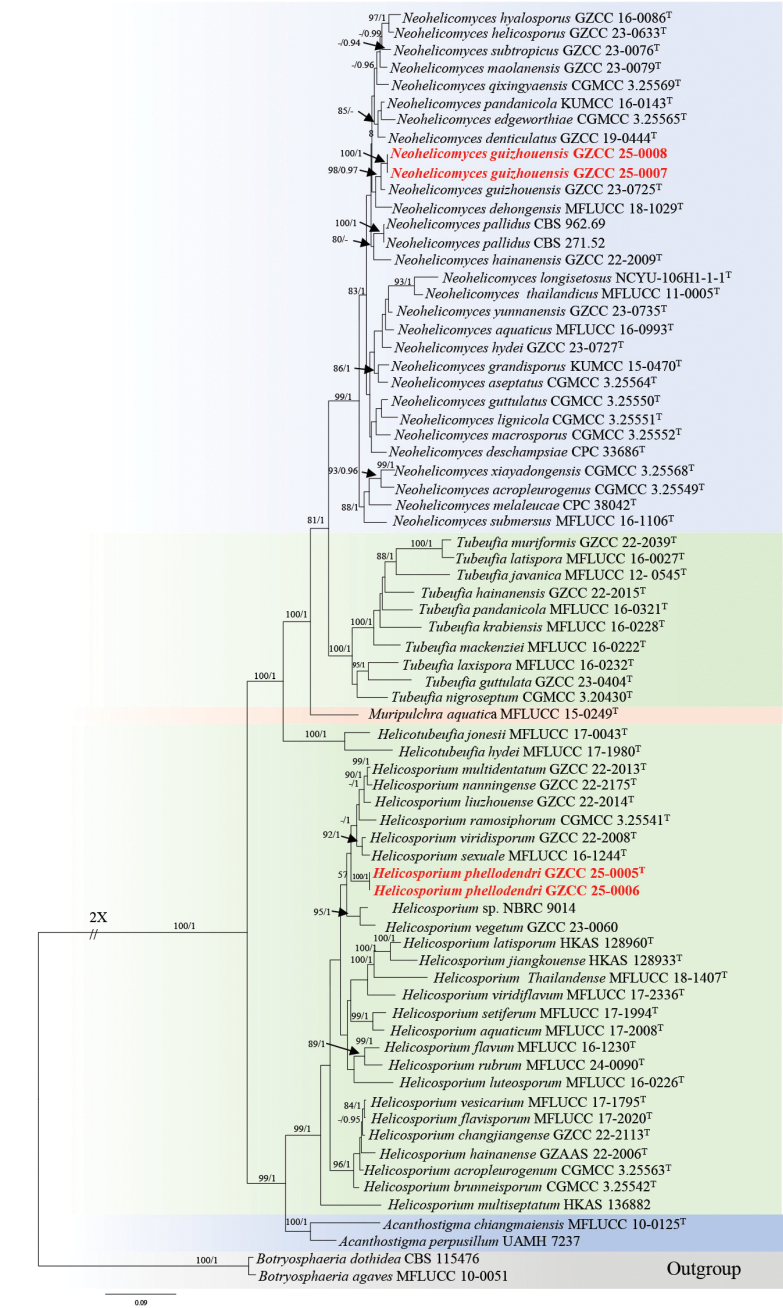 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5| Taxon | Strain | GenBank Accession numbers | |||
|---|---|---|---|---|---|
| LSU | ITS | ||||
|
| MFLUCC 10-0125T |
|
|
| – |
|
| UAMH 7237 |
|
| – | – |
|
| MFLUCC 10-0051 |
|
| – | – |
|
| CBS 115476 |
|
|
|
|
|
| CGMCC 3.25563T |
|
|
|
|
|
| MFLUCC 17-2008T |
|
|
|
|
|
| CGMCC 3.25542T |
|
|
|
|
|
|
|
|
|
| |
|
| MFLUCC 17-2020T |
|
|
|
|
|
| MFLUCC 16-1230T |
|
|
| – |
|
| GZAAS 22-2006T |
|
|
|
|
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| MFLUCC 16-0226T |
|
|
|
|
|
|
|
|
|
| |
|
| HKAS_136882 T |
|
|
| – |
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CGMCC 3.25541T |
|
|
|
|
|
| MFLUCC 24-0090T |
|
|
|
|
|
| MFLUCC 17-1994T |
|
|
|
|
|
| MFLUCC 16-1244T |
|
|
|
|
| NBRC 9014 |
|
| – | – | |
|
| MFLUCC 18-1407T |
|
|
| – |
|
|
|
|
|
| |
|
| MFLUCC 17-1795T |
|
|
|
|
|
| MFLUCC 17-2336T | – |
|
|
|
|
|
|
|
|
| |
|
| MFLUCC 17-1980T |
|
|
|
|
|
| MFLUCC 17-0043T |
|
|
|
|
|
| MFLUCC 15-0249T |
|
| – | – |
|
| CGMCC 3.25549T |
|
|
|
|
|
| MFLUCC 16-0993T |
|
|
|
|
|
| CGMCC 3.25564T |
|
|
|
|
|
| MFLUCC 18-1029T |
|
|
| – |
|
|
|
| – | – | |
|
| CPC 33686T |
|
| – | – |
|
| CGMCC 3.25565T |
|
|
|
|
|
| KUMCC 15-0470T |
|
| – |
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| CGMCC 3.25550T |
|
|
| – |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
| – |
|
| |
|
| CGMCC 3.25551T |
|
|
|
|
|
| NCYU-106H1-1-1T | – |
| – | – |
|
| CGMCC 3.25552T |
|
|
|
|
|
|
| – |
|
| |
|
| CPC 38042T |
|
|
| – |
|
| CBS 271.52 |
|
| – | – |
|
| CBS 962.69 |
|
| – | – |
|
| KUMCC 16-0143T |
|
|
| – |
|
| CGMCC 3.25569T |
|
|
|
|
|
| MFLUCC 16-1106T |
|
| – |
|
|
|
|
|
|
| |
|
| MFLUCC 11-0005T |
|
| – | |
|
| CGMCC 3.25568T |
|
|
|
|
|
|
|
| – | – | |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
| MFLUCC 12-0545T |
|
|
| – |
|
| MFLUCC 16-0228T |
|
|
|
|
|
| MFLUCC 16-0027T |
|
|
|
|
|
| MFLUCC 16-0232T |
|
|
|
|
|
| MFLUCC 16-0222T |
|
|
|
|
|
|
|
|
|
| |
|
| CGMCC 3.20430T |
|
|
|
|
|
| MFLUCC 16-0321T |
|
|
|
|
| Taxon | Strain | GenBank Accession numbers | ||
|---|---|---|---|---|
| LSU | ITS | |||
|
| MFLUCC 17-2080 |
|
|
|
|
| MFLUCC 17-1318 |
|
|
|
|
| ATCC 34953 T |
|
| – |
|
| MFLUCC 17-2259 T |
|
| – |
|
| MF1318 T | – |
| – |
|
| HKUCC 8797 | – |
| – |
|
| yone221 |
|
|
|
|
| HUEST 24.0140 |
|
|
|
|
| KH 401 |
|
|
|
|
| yone 280 |
|
|
|
|
| KUNCC 23-15949 |
|
|
|
|
|
|
|
| |
|
| FMR 13125 |
|
| – |
|
| NBRC 32502 T |
|
| – |
|
| BCC 11346 | – |
| – |
|
| ZHKUCC 24-0002 T |
|
| – |
|
| MFLUCC 1- 0258 T |
|
|
|
|
| CBS 396.59 | – |
| – |
|
| KUMCC 17-0268 |
|
|
|
|
| K 1847 |
|
|
|
|
| KUNCC 23-15923 |
|
| – |
|
| MFLU 16-1890 T |
| – |
|
|
| SNC182 | – |
|
|
|
| MFLUCC 10-0131 T |
|
| – |
|
|
|
|
| |
|
| MFLUCC 17-2053 T |
|
|
|
|
| BCC 3555 | – |
| – |
|
| KH 375 T |
|
|
|
|
| CBS-H 22129 |
| – | – |
|
| MFLU 16-1886 T |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| MFLUCC 10-0127 T |
|
| – |
| J15 | – |
| – | |
| J16 | – |
| – | |
| KT 2025 | – |
| – | |
| MFLUCC 15-0629 |
|
|
| |
| 19VA07 | – |
| – | |
|
| KUNCC 23-17212 |
|
|
|
|
| CCFC 241241 T |
| – | – |
|
| CBS 123359 T |
| – | – |
|
| K 2865 |
|
|
|
|
| MFLUCC 13-0773 T |
|
| – |
|
| CBS 209.65 |
|
| – |
|
| MFLUCC 17-2052 T |
|
|
|
|
| MFLUCC 17-2056 |
|
| – |
|
| MFLUCC 15-0631 T |
|
|
|
|
| ZHKUCC 24-0003 T |
|
|
|
|
| CGMCC 3.18703 T | – |
| – |
|
|
|
| – | |
|
| MW–2009a T | – |
| – |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Fungal Biology and Applications
Introduction
Phellodendron amurense (Rutaceae, Tracheophyta), a deciduous plant, is mainly distributed in East Asia. It is rich in alkaloids, flavonoids, triterpenoids, coumarins and other bioactive compounds (Chen et al. 2024). In China, it is renowned as one of the most important traditional Chinese medicine plants, listed in the Chinese Pharmacopoeia (2020 edition) and has been widely used for thousands of years (Xian et al. 2014; Wang et al. 2015; Dulan et al. 2022; Zhang et al. 2024). Medicinal plants are recognised as potential reservoirs for uncovering fungal diversity (Du et al. 2024). Endophytic and pathogenic fungi associated with P. amurense have been documented (Cao and Qi 1990; Li 2006; Li et al. 2009, 2018; Wen et al. 2022). However, saprobic fungi occurring on this medicinal host remain poorly explored.
Tubeufiaceae was established by Barr (1979) to accommodate the type genus Tubeufia and comprises mainly saprobic taxa inhabiting decaying wood. Members of the family are widely distributed in tropical and subtropical regions (Rossman 1987; Kirk et al. 2001; Lumbsch et al. 2010; Boonmee et al. 2014; Luo et al. 2017; Lu et al. 2018a, b), occurring in both terrestrial wood and aquatic environments (Hyde et al. 2016, 2017; Brahamanage et al. 2017; Chaiwan et al. 2017; Lu et al. 2017a, b, c, 2018a, b; Liu et al. 2018; Hongsanan et al. 2020). Morphologically, members of the family are characterised by helicosporous conidia in the asexual morph (Linder 1929; Samuels and Muller 1979; Spatafora et al. 2006;Tsui et al. 2006; Tsui Barr Lu et al. 2018b; Dong et al. 2020;Zhang et al.2020) and by superficial ascomata, bitunicate asci and hyaline to pale brown, elongate to obovoid, septate ascospores in the sexual morph (Barr 1980; Kodsueb et al. 2006; Boonmee et al. 2011, 2014; Brahamanage et al. 2017; Lu et al. 2018b; Karimi et al. 2025; Sun et al. 2025). Currently, Tubeufiaceae comprises 54 genera (Lu et al. 2018b; Liu et al. 2019a, b, c, d; Wijayawardene et al. 2022; Karimi et al. 2025), amongst which Helicosporium and Neohelicomyces represent common helicosporous hyphomycetes within the order Tubeufiales (Lu et al. 2018b).
Dictyosporiaceae was introduced by Boonmee et al (2016) to accommodate most cheirosporous hyphomycetous genera, with Dictyosporium designated as the type genus. According to Wang et al (2025), the family includes 23 genera and 178 species, excluding one novel species proposed in this study. Members of Dictyosporiaceae are saprobes on plant litter and decaying wood in both terrestrial and aquatic habitats worldwide (Boonmee et al. 2016; Fu et al. 2021; Shen et al. 2022). The type genus Dictyosporium is characterised by its cheiroid conidia (Corda 1836).
During a taxonomic survey of saprophytic fungi, associated with Phellodendron amurense in Guizhou Province, China, six fresh collections were obtained from the decaying branches in two forests sites. Morphological observations combined with multigene phylogenetic analyses revealed two novel species, Helicosporium phellodendri and Dictyosporium phellodendri and a new host record, Neohelicomyces guizhouensis. Detailed morphological descriptions, illustrations and molecular evidence are provided herein.
Materials and methods
Sample collection, specimen examination and isolation
Fresh specimens were collected from decaying wood of Phellodendron amurense in Anshun City and Guiyang City, Guizhou, China, in October 2024. Relevant collection data were recorded in the field. Specimens were examined and observed using a stereomicroscope (SMZ-168, Nikon, Japan). Micromorphological features were observed and photographed with an ECLIPSE Ni compound microscope (Nikon, Tokyo, Japan), equipped with a Canon 90D digital camera. Photoplates were prepared using Adobe Photoshop CC 2019 (Adobe Systems, USA) and Tarosoft® Image Frame Work.
Single spore isolations were performed following the method described by Senanayake et al. (2020). Germinated conidia were transferred on to fresh potato dextrose agar (PDA) plates and incubated in a 25 °C for colony development.
Material deposition
Dried specimens were deposited in the Herbarium of Cryptogams Kunming Institute of Botany, Academia Sinica (HKAS), Kunming, China. Living cultures were deposited in the Guizhou Culture Collection (GZCC), Guizhou, China. New taxa were registered on the Index Fungorum database (Index Fungorum 2025).
DNA extraction, PCR amplification and sequencing
Fresh mycelia were scraped using a sterile toothpick and transferred to 1.5 ml microcentrifuge tube. Genomic DNA was extracted using the Trelief Hi-Pure Fungus Genomic DNA Extraction Kit (Tsingke, China) according to the manufacturer’s instructions. Four genetic loci (LSU, ITS, tef1-α and rpb2) were amplified using the primers LR0R/ LR5 (Vilgalys and Hester 1990), ITS5/ ITS4 (White et al. 1990), EF1-983F/EF1-2218R (Rehner and Buckley 2005) and fRPB2-5F/fRPB2-7cR (Liu et al. 1999), respectively. PCR amplification was conducted in a 25 μl reaction mixture containing 21 μl of 1.1 × T3 Super PCR Mix (Tsingke Biotech, Chongqing, China), 2 μl of DNA template and 1 μl of each primer. The cycling parameters for LSU, ITS, tef1-a and rpb2 followed the protocols of Ma et al. (2024b). Purified PCR products were sequenced by Tsingke Biological Engineering Technology and Services Co., China.
Phylogenetic analyses
Sequence quality was verified using BioEdit v. 7.0.5.3 (Hall 1999). Forward and reverse reads were assembled with SeqMan v. 7.0.0 (DNASTAR, Madison, WI, USA; Swindell and Plasterer 1997) and deposited in GenBank. Similar sequences were retrieved using the BLASTn tool in NCBI (https://www.ncbi.nlm.nih.gov/). Reference sequences used for phylogenetic analyses are listed in Tables 1, 2. Each locus was aligned using MAFFT v.7.473 (Katoh et al. 2019) on the online server (https://mafft.cbrc.jp/alignment/server/) and ambiguous regions were trimmed using trimAl v.1.2rev59 (Capella-Gutiérrez et al. 2009). The alignments were concatenated using SequenceMatrix-Windows-1.7.8 (Vaidya et al. 2011).
Maximum Likelihood (ML) analyses were performed with the IQ-TREE web server (http://iqtree.cibiv.univie.ac.at/, Nguyen et al. 2015; Zeng et al. 2023) using default parameters. Bayesian Inference (BI) analyses followed the procedures described by Lu et al (2022). The best-fit substitution model for each gene partition was determined using MrModelTest 2.3 under the Akaike Information Criterion (AIC) (Nylander et al. 2008).
Phylogenetic results
The first phylogenetic tree represents the phylogenetic positions of the new collections within Tubeufiaceae, based on four loci: LSU, ITS, tef1-α and rpb2.The concatenated dataset comprised 3,411 characters (LSU: 1–851, ITS: 852–1,441, tef1-α: 1,442–2,353, rpb2: 2,354–3,411), including gaps. A total of 73 strains, including isolates obtained in this study and two outgroups, Botryosphaeria agaves (MFLUCC 10-0051) and B. dothidea (CBS 115476), were analysed. Both ML and BI analyses yielded similar tree topologies. Fig. 1 illustrates the best- scoring ML tree, with a final likelihood value of -29709.023. In our phylogenetic tree (Fig. 1), our strains (GZCC 25-0007 and GZCC 25-0008) clustered within Neohelicomyces, while GZCC 25-0005 and GZCC 25-0006 clustered within Helicosporium in the family Tubeufiaceae.
Maximum Likelihood (ML) phylogenetic tree, based on the combined dataset of LSU, ITS, tef1-α and rpb2 sequences. Bootstrap support values for ML analysis (≥ 75%) and posterior probability (PP) values from Bayesian inference (≥ 0.95) are shown at the corresponding nodes. Ex-type strains are indicated by “T” and newly-generated isolates are highlighted in bold red.
Phylogenetic analysis of Dictyosporium was based on three loci: LSU, ITS and tef1-α. The dataset comprised a total of 51 strains, which included isolates obtained in this study as well as the two outgroup taxa, Aquadictyospora clematidis (MFLUCC 17-2080) and A. lignicola (MFLUCC 17-1318). The concatenated sequence matrix contains 2,352 characters (LSU: 1–851, ITS: 852–1,442, tef1-α: 1,443–2,352) including gaps. The phylogenetic tree (Fig. 2) reveals that GZCC 25-0522 and GZCC 25-0523 cluster together with maximal support values (ML/PP = 100/1), forming a distinct and well-supported monophyletic lineage within Dictyosporium.
Maximum Likelihood (ML) phylogenetic tree, based on the combined dataset of LSU, ITS and tef1-α sequences. Bootstrap support values for ML analysis (≥ 75%) and posterior probability (PP) values from Bayesian inference (≥ 0.95) are shown at the corresponding nodes. Ex-type strains are indicated by “T” and newly-generated isolates are highlighted in bold red.
Taxonomy
Helicosporium
Taxon classificationFungiTubeufialesTubeufiaceae
Nees, Syst. Pilze (Würzburg): 68 (1816) [1816–17]
62429C69-8BC8-5A47-9600-40500E1D39D3
Notes.
Helicosporium, one of the three earliest described helicosporous genera (Helicoma, Helicomyces and Helicosporium) (Link 1809; Nees 1817; Corda 1837), was established by Nees (1817) with H. vegetum as the type species. To date, 118 species are recorded in the Index Fungorum (2025). However, based on combined morphological and molecular evidence, only about 32 species are currently accepted in this genus (Lu et al. 2018b; Xiao et al. 2023; Ma et al. 2024b; Bai et al. 2025; Peng et al. 2025; Sun et al. 2025). Amongst these, sexual morphs have been confirmed for only five species: H. flavum, H. sexuale, H. vegetum, H. rubrum and H. multiseptatum (Boonmee et al. 2014, 2021; Brahmanage et al. 2017; Peng et al. 2025; Sun et al. 2025). Most species of Helicosporium are saprobes inhabiting freshwater and terrestrial environments, with a broad geographical distribution (Morgan-Jones and Goos 1992; Zhao et al. 2007; Lu et al. 2017a, 2018b, 2022; Dong et al. 2020; Boonmee et al. 2021; Hsieh et al. 2021; Xiao et al. 2023; Ma et al. 2024b). The majority of Helicosporium species have been isolated from decaying wood of unidentified plant hosts, while H. multiseptatum as the saprobe and H. thailandicum as the pathogen obtained from Poaceae sp. and Elaeis sp. (Arecaceae), respectively (Sun et al. 2025). In this study, we introduce a new Helicosporium species which was isolated from medicinal plants.
Helicosporium
phellodendri
Taxon classificationFungiTubeufialesTubeufiaceae
S.P. Zou, Y. P. Xiao & Y.Z. Lu sp. nov.
3979AD23-9DE3-552F-8105-B96E019FAFEC
Index Fungorum: IF904507
Etymology.
the specific epithet “phellodendri” refers to the host genus name “Phellodendron” from which the fungus was isolated.
Holotype.
HKAS 145879.
Description.
Saprobic on dead wood of Phellodendron amurense. Sexual morph Undetermined. Asexual morph Hyphomycetous, helicosporous. Colonies on natural substrate superficial, effuse, gregarious, with masses of crowded, glistening conidia, yellowish-green. Mycelium partly immersed, partly superficial, composed of pale brown to brown, branched, septate, guttulate, smooth hyphae. Conidiophores 89–97 × 4–5 μm (x̄ = 94 × 4.5 μm, n = 20), macronematous, mononematous, erect, cylindrical, straight or broadly curved, simple or occasionally branched, septate, brown at base, paler towards the apex, thick-walled. Conidiogenous cells 9.5–10.5 × 3.5–4.8 μm (x̄ = 10 × 4.3 μm, n = 15), holoblastic, monoblastic, integrated, intercalary, cylindrical, pale brown to brown, smooth-walled. Conidia solitary, pleurogenous, helicoid, tapering towards the ends, 10.5–12.5 μm diam. and conidial filament 1.5–2 μm wide (x̄ = 11.5 × 1.8 μm, n = 25), 60–65 μm long (x̄ = 63 μm, n = 25), tightly coiled 2½–3 times, not becoming loose in water, multi-septate, hyaline, smooth-walled.
Helicosporium phellodendri (HKAS 145879, holotype). a, b. Colonies on the natural substrate; c–f. Conidiophores and conidiogenous cells; g–j. Conidia; k, l. Surface and reverse view of colonies on PDA after 36 days of incubation at 25 °C. Scale bars: 30 μm (c–f); 5 μm (g–j).
Culture characteristics.
Conidia produce germ tubes on PDA within 14 hours of incubation at 25 °C. Colonies on PDA are circular, with a flat surface and entire edge, reaching a diameter of 26 mm after 36 days of incubation at 25 °C. The colony appears dark brown to black at centre, brown at outer ring on the top view and brown on the reverse view.
Material examined.
China • Guizhou Province, Guiyang City, Qingzhen City, Maige Miao and Bouyei Ethnic Township, Xiaochong Village, 26°74′N, 106°37′E, on the rotting wood of Phellodendron amurense, 1 October 2024, Shi-Ping Zou, HB-Z17-1 (HKAS 145879, holotype), ex-type culture GZCC 25-0005 • Ibid., HB-Z17-2 (HKAS 145878), living culture GZCC 25-0006.
Notes.
Morphological examination indicated that Helicosporium phellodendri conforms to the generic concept of Helicosporium, characterised by pale yellow to yellow-green colonies on the natural woody substratum, erect setiferous cylindrical conidiophores with denticulate conidiogenous cells and hyaline to yellow-green helicoid conidia which are mainly becoming loosely coiled in water (Ma et al. 2024b). Helicosporium phellodendri is morphologically similar to H. multidentatum, but differs in having longer conidia (105–130 μm vs. 60–65 μm), with more coils (3¼–3¾ vs. 2½–3). Furthermore, the conidia of H. multidentatum become loosely coiled in water, while those of H. phellodendri do not loosen (Xiao et al. 2023). Phylogenetic analysis (Fig. 1) also supports that H. phellodendri and H. multidentatum are distinct species. Strains GZCC 25-0005 and GZCC 25-0006 formed a well-supported, independent lineage within Helicosporium (Fig. 1). Helicosporium viridisporum and H. sexuale are closely related to H. phellodendri. However, H. viridisporum can be distinguished from H. phellodendri by its longer conidiophores (69–189 × 3–5 μm vs. 89–97 × 4–5 μm), denticulate conidiogenous cells and acropleurogenous, aseptate conidia (Lu et al. 2022). Helicosporium sexuale displays sexual morph (Boonmee et al. 2021), whereas, H. phellodendri exhibits asexual morph. Therefore, H. phellodendri is introduced here as a novel species.
Neohelicomyces
Taxon classificationFungiTubeufialesTubeufiaceae
Z.L. Luo, Bhat & K.D. Hyde, Cryptog. Mycol. 38(1): 39 (2017)
98E12299-ADBF-5FF2-82E7-9876BBCF2FF2
Notes.
Neohelicomyces was proposed by Luo et al. (2017) with N. aquaticus designated as the type species. To date, 30 species have been recorded in Index Fungorum (2025). All known taxa represent asexual morphs (Ma et al. 2024a, b), except for N. sexualis, which was characterised and linked to its sexual morph, based on both morphological and molecular evidence (Sun et al. 2025). The asexual morphs of Neohelicomyces species are characterised by macronematous, mononematous, erect, septate conidiophores with holoblastic conidiogenous cells and loosely or tightly coiled helicoid conidia (Hsieh et al. 2021; Lu et al. 2022; Yang et al. 2023; Ma et al. 2024a, b). In contrast, the sexual morph produces scattered, superficial, solitary, subglobose, light brown ascomata on natural substrates, lacking setae. Ascospores are multiseriate, narrowly cylindrical, straight to slightly curved, hyaline to pale brown, septate and rough (Sun et al. 2025). Species of Neohelicomyces are saprobic, typically inhabiting freshwater or terrestrial environments on decaying wood in Asia, Europe and North America, including records from China, Germany and Thailand (Ma et al. 2024a; Sun et al. 2025). Only a few host plants have been documented for this genus (Ma et al. 2024a).
Neohelicomyces
guizhouensis
Taxon classificationFungiTubeufialesTubeufiaceae
J. Ma, Y.Z. Lu & K.D. Hyde, MycoKeys 105: 323 (2024)
FAB80A6E-7F0B-5E3F-B38C-5FFF2F74DE23
901915
Description.
Saprobic on dead wood of Phellodendron amurense. Sexual morph Undetermined. Asexual morph Hyphomycetous, helicosporous. Colonies on natural substrate superficial, effuse, gregarious, with mass of crowded conidia, white. Mycelium partly immersed, partly superficial, composed of hyaline to pale brown, branched, septate, guttulate, smooth hyphae. Conidiophores 154–175 × 4–5 μm (x̄ = 161 × 4.5 μm, n = 20), macronematous, mononematous, erect, cylindrical, tapering towards the apex, straight or slightly flexuous, unbranched, septate, subhyaline to pale brown, thick-walled. Conidiogenous cells 11–19 × 2.5–4 μm (x̄ = 14.5 × 3.5 μm, n = 20), holoblastic, monoblastic or polyblastic, integrated, terminal or intercalary, cylindrical, with denticles, subhyaline to pale brown, smooth-walled. Conidia solitary, acropleurogenous, helicoid, developing on tooth-like protrusion, 14–20 μm diam. and conidial filament 1.5–3 μm wide (x̄ = 16 × 2 μm, n = 20), 94–133 μm long (x̄ = 113 μm, n = 25), tightly coiled 3–3½ times, not becoming loose in water, guttulate, hyaline, smooth-walled.
Neohelicomyces guizhouensis (HKAS 145881). a, b. Colonies on the natural substrate; c–g. Conidiophores; h–k. Conidia; l, m. Surface and reverse view of colonies on PDA after 45 days of incubation at 25 °C. Scale bars: 50 μm (c–f); 20 μm (g); 10 μm (h–k).
Culture characteristics.
Conidia produce germ tubes on PDA within 10 hours of incubation at 25 °C. Colonies on PDA are circular, with a raised centre and undulate edge, reaching a diameter of 30 mm after 45 days of incubation at 25 °C. The colony appears grey to pale brown on the top view, with the reverse side ranging from pale brown to brown.
Material examined.
China • Guizhou Province, Guiyang City, Qingzhen City, Maige Miao and Bouyei Ethnic Township, Xiaochong Village, 26°74′N, 106°37′E, on the rotting branch of Phellodendron amurense, 1 October 2024, Shi-Ping Zou, HB-Z15-1 (HKAS 145881), living culture GZCC 25-0007 • Ibid., HB-Z15-2 (HKAS 145880), living culture GZCC 25-0008.
Notes.
Phylogenetic analysis showed that our isolates (GZCC 25-0007 and GZCC 25-0008) grouped with N. guizhouensis (GZCC 23-0725) by strong support (ML/PP = 98%/0.97; Fig. 1). Pairwise comparison of the ITS, LSU, tef1-α and rpb2 sequences between GZCC 25-0007 and N. guizhouensis (GZCC 23-0725) revealed minimal nucleotide differences: 5 bp/424 bp (1.2%, including two gaps), 3 bp/859 bp (0.3%, including two gaps), 2 bp/892 bp (0.2%, no gaps) and 2 bp/903 bp (0.2%, no gaps), respectively. Morphologically, our new collections share identical characteristics of conidiogenous cells and conidia with the holotype of N. guizhouensis (Ma et al. 2024a). Therefore, GZCC 25-0007 and GZCC 25-0008 are identified as N. guizhouensis. Neohelicomyces guizhouensis was originally described by Ma et al. (2024a) from submerged wood in a stream in China and our study represents its first record associated with Phellodendron amurense.
Dictyosporium
Taxon classificationFungiPleosporalesDictyosporiaceae
Corda, Weitenweber’s Beitr. Nat. 1: 87 (1837)
27F8C3BB-10E6-5CCA-8810-F65D61BFCFFB
Notes.
Dictyosporium was proposed by Corda (1836) with D. elegans designated as the type species. Species of Dictyosporium are distributed across the globe in both freshwater and terrestrial habitats, functioning as saprobes and frequently found on decaying wood, plant litter,and in soil (Van Emden 1975; Boonmee et al. 2016; Yang et al. 2018; Zhang et al. 2023). To date, a total of 91 species has been recorded within this genus (Index Fungorum 2025).
Dictyosporium
phellodendri
Taxon classificationFungiPleosporalesDictyosporiaceae
S.P. Zou, Y. P. Xiao & Y.Z. Lu sp. nov.
B02086B7-AC53-5678-B33B-6E7F31AC07BD
Index Fungorum: IF904506
Etymology.
The specific epithet “phellodendri” refers to the host genus name “Phellodendron” from which the fungus was isolated.
Holotype.
HKAS 149888.
Description.
Saprobic on dead wood of Phellodendron amurense. Sexual morph Undetermined. Asexual morph Hyphomycetous. Colonies on natural substrate forming sporodochial conidiomata, superficial, punctiform, scattered, black, glistening. Mycelium immersed, pale to brown. Conidiophores 3–5 μm wide, micronematous, simple, aseptate, cylindrical, hyaline to pale brown, thin-walled. Conidiogenous cells 5–8 × 3–6 μm (x̄ = 6.3 × 4.7 μm, n = 10), holoblastic, hyaline or pale brown, cylindrical. Conidia 17–44 × 15–27 μm (x̄ = 30 × 21 μm, n = 30), acrogenous, cheiroid, complanate, smooth-walled, pale brown or pale yellow, mostly consisting of four closely adpressed rows of cells, rarely with three or five rows, 33–38 ×4–6 μm (x̄ = 35.5 × 5.5 μm, n = 30), 4–10-euseptate in each row, with hyaline, tubular, elongated apical appendages arising from two outer rows, 13–58 × 3–6 μm (x̄ = 32.9 × 4.2 μm, n = 15).
Dictyosporium phellodendri (HKAS 149888, holotype). a, b. Colonies on the natural substrate; c, d. Squash mount of a sporodochium; e–h. Conidia with conidiophores; i–m. Conidia with appendages; n–q. Conidia at different stages; r. Germinated conidium; s, t. Surface and reverse view of colonies on PDA after 39 days of incubation at 25 °C. Scale bars: 50 μm (c, d); 10 μm (e); 20 μm (f–m); 10 μm (n); 20 μm (o–r).
Culture characteristics.
Conidia produce germ tubes on PDA within 18 hours of incubation at 25 °C. Colonies on PDA are circular, with a flat surface and entire edge, reaching a diameter about 28 mm after 39 days of incubation at 25 °C. The colony appears white on the top view, while the reverse side with pale grey centre and yellow edge.
Material examined.
China • Guizhou Province, Guiyang City, Anshun City, Xixiu District, Yaowang Valley, 26°16'17"N, 106°7′0"E, on the rotting branch of Phellodendron amurense, 15 October 2024, Shi-Ping Zou, HBYWGC13 (HKAS 149888, holotype), ex-type culture GZCC 25-0522 • Ibid., HBYWGC25 (149889), living culture GZCC 25-0523.
Notes.
In the phylogenetic analysis, Dictyosporium phellodendri (GZCC 25-0522 and GZCC 25-0523) formed a sister clade to D. muriformis with 80% ML/0.98PP (Fig. 2), but D. muriformis, possesses smaller conidia (20–30 × 11–14.5 μm vs. 26–32 × 15–25 μm), darker pigmentation (median brown vs. pale brown or pale yellow) and the absence of appendages (Hyde et al. 2020). Morphologically, D. phellodendri is most similar to D. alatum (ATCC 34953) in conidial shape. However, it is distinguished by its larger conidia (17–44 × 15–27 μm vs. 26–32 × 15–25 μm), paler colouration and 4–10 septa per row (Van Emden 1975). Furthermore, comparisons of sequences between D. phellodendri and D. alatum (ATCC 34953) revealed differences in the ITS region of 13% (64/507 bp, including 11 gaps) and in the LSU region of 3% (18/573 bp, with 4 gaps). We, therefore, designated our collections as a new species, based on morphological and phylogenetic evidence.
Discussion
In this study, three saprobic fungal taxa were collected from terrestrial habitats in Guizhou Province, China. Based on multigene phylogenetic analyses combined with morphological comparative evidence, Helicosporium phellodendri and Dictyosporium phellodendri are proposed as two novel species, while Neohelicomyces guizhouensis is identified as a new host record.
The family Tubeufiaceae (Tubeufiales, Dothideomycetes, Ascomycota) represents an important lineage of helicosporous fungi. Most genera and species in the family exhibit distinct asexual morphs, characterised by filamentous, helicoid (spirally coiled) conidia, which are key diagnostic features for species identification (Barr 1979). To date, 33 Helicosporium species, including the new species introduced herein, have been recognised (Bai et al. 2025), while 30 species of Neohelicomyces have been accepted (Ma et al. 2025). Members of both genera are commonly isolated from decaying wood or plant litter of unidentified hosts in freshwater and terrestrial habitats (Peng et al. 2025; Sun et al. 2025). Most known species are represented only by their asexual morphs; to date, four species of Helicosporium and one of Neohelicomyces have been reported in their sexual morphs (Brahmanage et al. 2017; Xiao et al. 2023; Sun et al. 2025).
Dictyosporium species play a crucial ecological role as decomposers of decaying wood and plant litter and are reported worldwide from both terrestrial and aquatic environments (Dubey 2022). Ho et al. (2002) investigated the seasonality and succession of fungi on submerged wood and found that Dictyosporium species occurred on diverse woody substrates in Tai Po Kau Forest Stream. Furthermore, Dictyosporium has been shown to enhance plant growth and resistance to pathogen infection (Peitl et al. 2020; Jiang et al. 2023). Dictyosporium phellodendri, isolated from decay branch of Phellodendron amurense, suggests that this fungal community plays a functional role in wood decomposition of Phellodendron amurense in the ecosystem. Future research may reveal that species of this genus participate in a broader range of ecological functions.
The members of Tubeufiaceae, especially helicosporous taxa, are known to produce diverse bioactive secondary metabolites (Lu et al. 2018b; Ma et al. 2024b). Likewise, Dictyosporium species has been recognised as sources of novel natural products (Tran et al. 2019, 2020). All strains isolated in this study originated from the dead branches of P. amurense, a medicinal plant rich in bioactive compounds, such as berberine, polyphenols, fibre, phytosterols and carotenoids. Extracts of this plant exhibit a wide range of pharmacological properties, including anti-inflammatory, antibacterial and anticancer activities and are used in the treatment of pneumonia, diarrhoea and circulatory disorders (Kim et al. 2011; Steinmann et al. 2012; Velmurugan et al. 2017; Wang et al. 2017; Sun et al. 2019; Balážová et al. 2022; Erhan et al. 2023; Feng et al. 2023). The isolation of these saprobic fungi from P. amurense not only expands our understanding of their biodiversity, but also provides valuable resources for exploring novel bioactive metabolites and elucidating the ecological interactions between fungi and medicinal host plants.
Supplementary Material
XML Treatment for Helicosporium
XML Treatment for Helicosporium phellodendri
XML Treatment for Neohelicomyces
XML Treatment for Neohelicomyces guizhouensis
XML Treatment for Dictyosporium
XML Treatment for Dictyosporium phellodendri
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bai S Wang F Wan SR Lv XK Chen LJ Wu R Ma J (2025) Two novel Helicosporium species (Tubeufiaceae, Tubeufiales) from southern China based on morphological and molecular evidence.Myco Keys 124: 159–176. 10.3897/mycokeys.124.16893341179845 PMC 12576478 · doi ↗ · pubmed ↗
- 2Barr ME (1979) A classification of Loculoascomycetes.Mycologia 71(5): 935–957. 10.1080/00275514.1979.12021099 · doi ↗
- 3Barr ME (1980) On the family Tubeufiaceae (Pleosporales).Mycotaxon 12(1): 137–167. 10.5962/p.417317 · doi ↗
- 4Boonmee S Zhang Y Chomnunti P Chukeatirote E Tsui CKM Bahkali AH Hyde KD (2011) Revision of lignicolous Tubeufiaceae based on morphological reexamination and phylogenetic analysis.Fungal Diversity 51(1): 63–102. 10.1007/s 13225-011-0147-4 · doi ↗
- 5Boonmee S Rossman AY Liu JK Li WJ Dai DQ Bhat JD Jones EBG Mc Kenzie EHC Xu JC Hyde KD (2014) Tubeufiales, ord. nov., integrating sexual and asexual generic names.Fungal Diversity 68(1): 239–298. 10.1007/s 13225-014-0304-7 · doi ↗
- 6Boonmee SD’souza MJ Luo ZL Pinruan U Tanaka K Su H Bhat DJ Mc Kenzie EHC Jones EBG Taylor JE Phillips AJL Hirayama K Eungwanichayapant PD Hyde KD (2016) Dictyosporiaceae fam. nov.Fungal Diversity 80(1): 457–482. 10.1007/s 13225-016-0363-z · doi ↗
- 7Boonmee S Wanasinghe DN Calabon MS Huanraluek N Chandrasiri SK Jones GE Rossi W Leonardi M Singh SK Rana S Singh PN Maurya DK Lagashetti AC Choudhary D Dai YC Zhao CL Mu YH Yuan HS He SH Phookamsak R Jiang HB Martín MP Dueñas M Telleria MT Kałucka IL Jagodziński AM Liimatainen K Pereira DS Phillips AJL Suwannarach N Kumla J Khuna S Lumyong S Potter TB Shivas RG Sparks AH Vaghefi N Abdel-Wahab MA Abdel-Aziz FA Li GJ Lin WF Singh U Bhatt RP Lee HB Nguyen TTT Kirk PM Dutta AK Acharya K Sarma VV Niranjan M Rajeshkumar KC Ashtekar N Lad S Wijayawardene NN Bhat JD Xu RJ Wijesinghe SN Shen HW Luo ZL Zhang JY Sysouphanth · doi ↗ · pubmed ↗
- 8Brahamanage RS Lu YZ Bhat DJ Wanasinghe DN Yan JY Hyde KD Boonmee S (2017) Phylogenetic investigations on freshwater fungi in Tubeufiaceae (Tubeufiales) reveals the new genus Dictyospora and new species Chlamydotubeufia aquatica and Helicosporium flavum.Mycosphere: Journal of Fungal Biology 8(7): 917–933. 10.5943/mycosphere/8/7/8 · doi ↗