Rhododendron mulunense (Ericaceae), a new species from the karst mountains of Guangxi and Guizhou provinces, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2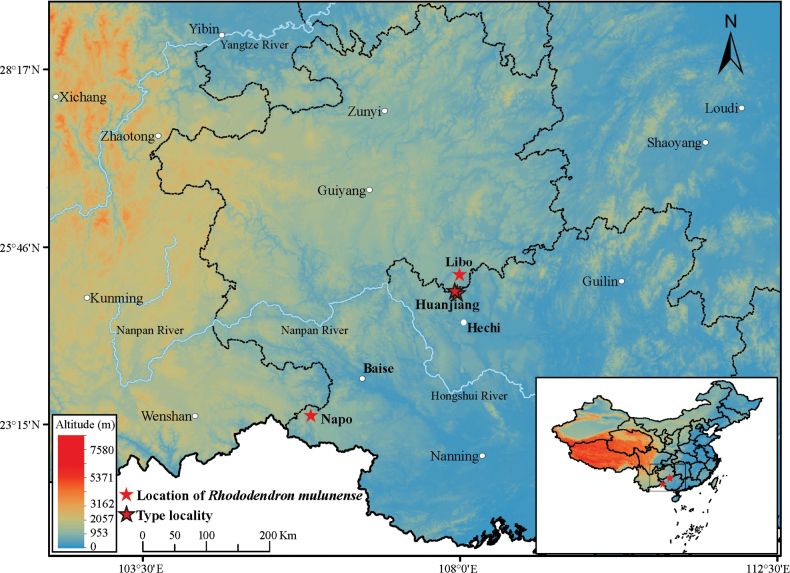 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5| Characters |
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|---|
| Young shoot | hairless | sparsely setose | hairless | densely setose | hairless | sparsely setose | hairless |
| Leaf shape | elliptic, oblong-lanceolate | elliptic, oblong-obovate | oblong-lanceolate, obovate | oblong-lanceolate, narrowly obovate | oblong | elliptic, elliptic-obovate | oblong-elliptic |
| Leaf size (cm) | 4.2–7.4 × 2.3–3.8 | 3.5–4.5 × 1.8–2.4 | 5.8–10.9 × 2.4–5.3 | 4.5–7.0 ×1.6–4.5 | 7.2–16.3 × 2.1–5.0 | 4.7–8.2 × 1.9–4.1 | 4.0–5.1 × 1.8–2.5 |
| Leaf margin | hairless | densely setose | hairless | densely setose | hairless | densely setose | hairless |
| Scales × (own diameter apart) | 1–4 | 1–3 | 1 or contiguous | 0.5–1 | 1–3 | 1–1.5 | 1–2 |
| Petiole | hairless | densely setose | nearly hairless | densely setose | hairless | sparsely setose | sparsely setose |
| Inflorescence | 2–4-flowered | 1–2-flowered | 2–4-flowered | 2–3-flowered | 2–3-flowered | 2–4-flowered | 2-flowered |
| Calyx margin | hairless | hairless | sparsely setose | densely setose | hairless | densely setose | hairless |
| Calyx lobe length (mm) | 0.5–1.0 | 0.7–1.2 | 1.0–1.6 | 1.0–3.0 (6.0) | 10–15 | 8–10 | 7–8 |
| Corolla shape | tubular-funnelform | broadly funnelform | broadly funnelform | broadly funnelform, funneled-campanulate | tubular-campanulate | broadly funnelform | broadly funnelform |
| Corolla size (cm) | 5.9–7.5 × 4.7–7.0 | 3.1–4.5 × 3.3–4.9 | 4.5–6.5 × 4.2–5.8 | 3.7–5.7 × 3.4–5.4 | 8.0–9.5 × 6.5–8.3 | 4.5–6.1 × 4.9–5.5 | 4.1–6.2 × 4.3–5.1 |
| Tube size (cm) | 2.9–3.9 × 0.5–1.2 | 1.9–2.5 × 1.7–2.4 | 0.6–1.3 × 1.7–2.9 | 0.5–1.2 × 1.7–2.6 | 3.5–4.0 × 2.0–3.5 | 0.5–0.8 × 1.8–2.3 | 0.4–0.7 × 1.9–2.2 |
| Color of corolla tube outside | white flushed with pink | yellowish green | white, sometimes flushed with pink | white flushed with pale red | white to yellowish green | white, sometimes flushed with pink | white flushed with light pink |
| Corolla tube outside | not scaly, pubescent | sparsely scaly, hairless | densely scaly and pubescent at base | scaly and pubescent at base | densely scaly, hairless | sparsely scaly, hairless | sparsely scaly, hairless |
| Flowering time | March to April | March to April | April to May | April to May | May to June | April to May | April to May |
| Geographic distribution | NW and SW Guangxi, SE Guizhou | Central Guangxi | SW and W Guizhou, NW, W, S, SE, and Central Yunnan | W Guizhou, NE Yunnan | N, NE, and NW Guangxi, Guizhou, Hunan, SE Yunnan | Guangxi, Guizhou, Guangdong, Fujian, Jiangxi, Hunan | Guangxi, Jiangxi, Zhejiang, Fujian |
| 1 | calyx lobes usually less than 3 mm in length (rarely much longer) |
|
| – | calyx lobes far more than 6 mm in length |
|
| 2 | corolla tubular-funnelform with a slender tube |
|
| – | corolla broadly funnelform with a robust tube |
|
| 3 | young shoot and leaf margin sparsely to densely setose |
|
| – | young shoot and leaf margin hairless |
|
| 4 | scales on abaxial leaf surface generally 2–3 or 4 × their own diameter apart |
|
| – | scales on abaxial leaf surface less than one (rarely one) × their own diameter apart |
|
| 5 | corolla tubular-campanulate, generally 8–10 cm in length |
|
| – | corolla broadly funnelform, less than 7 cm in length |
|
| 6 | young shoot, petiole, and leaf margin sparsely or densely setose |
|
| – | young shoot, petiole, and leaf margin hairless; usually epiphytic |
|
| 7 | young shoot and leaf margin hairless, calyx margin sparsely setose |
|
| – | young shoot and leaf margin densely setose, calyx margin setose |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBioactive Natural Diterpenoids Research · Plant and Fungal Species Descriptions · Ginger and Zingiberaceae research
Introduction
Rhododendron L. (1753: 392) is the largest genus in the family Ericaceae, comprising approximately 1,150 species worldwide (Khan et al. 2021) and over 1,000 species in the northern hemisphere (Feng 1983; Chamberlain et al. 1996; Gibbs et al. 2011; Geng 2014; Yu et al. 2017; Mao et al. 2021). The species were traditionally divided into eight subgenera (Chamberlain et al. 1996), of which only five are supported by phylogenetic studies (Goetsch et al. 2005; Ma et al. 2022; Xia et al. 2022, 2024). The great majority of Rhododendron species occur in China and the Malay Archipelago (Fang and Min 1995; Brown et al. 2006). In China, there are ca. 600 species of the genus documented in most provinces except for Ningxia and Xinjiang (Fang et al. 2005; Cheng et al. 2021), among which ≥ 400 species are endemic, with the greatest species diversity in the south and southwest of the country (Chen and Fang 1957; Yang et al. 1994; Li 1995; Fang et al. 2005; Cheng et al. 2021).
Field investigations in Guangxi and Guizhou Province, southern China, have led to the discovery of several new Rhododendron species in recent years (Chen and Lan 2003; Gao and Li 2004; Chen et al. 2010a, b; Ma et al. 2015; Dai et al. 2020; Tong et al. 2020; Chang et al. 2021; Deng et al. 2025), indicating that the species diversity of Rhododendron in these areas has been underestimated. During fieldwork investigating Rhododendron diversity in Guangxi Mulun National Nature Reserve from 2021 to 2024, we found an unknown Rhododendron species growing as an upright shrub on a karst mountain. It had scaly and broad leaves, terminal and umbellate inflorescences, large-sized and tubular-funnelform corollas with five lobes, 10 unequal stamens, scaly and slender styles, and large and scaly capsules – typical of subsection Maddenia (Hutch.) Sleumer (1949: 525) in subgenus Rhododendron. Then, in late March 2025, we obtained photographs of Rhododendron plants with matching morphological characteristics from the karst mountains of Napo County, southwest Guangxi, and from Maolan National Nature Reserve, southeast Guizhou. We compared the plants both to descriptions in the literature and to herbarium specimens of species in subsect. Maddenia (Fang et al. 2005; Li 2008; Geng 2014; Chang et al. 2021; Yang et al. 2022), concluding that they represent a species that is new to science. We formally describe and illustrate the species here.
Materials and methods
Morphological studies of the newly identified species were conducted through field investigations during flowering and fruiting times at type localities, and through thorough examinations of herbarium specimens of related species in the herbaria of the Guangxi Institute of Botany (IBK), South China Botanical Garden Herbarium (IBSC), Kunming Institute of Botany (KUN), and Institute of Botany (PE), Chinese Academy of Sciences (Qin et al. 2019), as well as online images of specimens in the herbaria of The Natural History Museum (BM), Royal Botanic Garden Edinburgh (E), and Royal Botanic Gardens (K) (https://sweetgum.nybg.org/science/ih/), and on the website of JSTOR Global Plants (https://plants.jstor.org/). Voucher specimens of the new species are deposited in the Herbarium of the Guangxi Institute of Botany (IBK), Chinese Academy of Sciences (Qin et al. 2019). In addition to consulting relevant publications and herbarium specimens (including types) of the closely related species Rhododendron wumingense (Fang 1983) in the herbarium (IBK), we conducted field observations for R. wumingense in spring 2024. We then performed comparative analyses of morphological characteristics and phenological patterns for the newly described species and related species. The conservation status of the new species was assessed based on the IUCN Red List Categories and Criteria (Gibbs et al. 2011; IUCN 2012).
Taxonomic treatment
Rhododendron
mulunense
Taxon classificationPlantaeEricalesEricaceae
Y.S.Huang & X.X.Mao sp. nov.
13369960-870B-5345-83A5-6540EBDEEDD2
urn:lsid:ipni.org:names:77374716-1
Type.
China. • Guangxi: Hechi City, Huanjiang County, Mulun National Nature Reserve, 25°06'N, 107°57'E, on the limestone under sparse forest, in summits of karst mountains, elev. 641 m, 3 March 2021, Ying Qin et al., GXQY20210303021 (holotype: IBK! 00470814, isotype: IBK! 00470813) (Fig. 1)
Holotype specimen of Rhododendron mulunense (IBK).
Diagnosis.
Resembles Rhododendron wumingense Fang but distinguishable by the absence of indumentum on young shoots, leaf margins, and petioles (vs. sparsely setose), longer and larger leaves (4.2–7.4 × 2.3–3.8 vs. 3.5–4.5 × 1.8–2.4 cm), relatively sparser scales on abaxial leaf surfaces (1–4 vs. 1–3 × their own diameter apart), corollas that are tubular funneled (vs. broadly funneled), longer and wider (5.9–7.5 × 4.7–7 vs. 3.1–4.5 × 3.3–4.9 cm) with slender tubes (2.9–3.9 × 0.5–1.2 vs. 1.9–2.5 × 1.7–2.4 cm), outer surface of corolla tubes white flushed with pink (vs. yellowish green) and pubescent but not scaly (vs. sparsely scaly and hairless) (Figs 1, 2, 4; Table 1).
Table 1.: Comparisons of morphological characteristics, phenology, and geographic distributions of Rhododendron mulunense and similar species.
Rhododendron mulunense. A. The habitat of this new species in type localities; B. Flowering plants; C. Leaves; D–F. Flowering branchlets, inflorescences, and flowers; G, H. Dissection of corolla; I. Flower buds; J. Stamens and style; K. Styles and ovaries; L. Dissection of ovary; M. Stigmas; N. Poorly developed stamens; O. Normal stamens; P, Q. Anther abaxial and adaxial surface; R. Tube base; S. Flower bud scales; T. Dehiscent capsule. Photographs by Yu-Song Huang.
Description.
Shrubs, 0.5–2 m tall; young shoots stout, short, and relatively sparse, brownish scaly, hairless. Stems robust, erect, much-branched and sparsely scaly. Leaf-bud scales acuminate, elongated at flowering time. Petioles densely scaly, hairless, 0.5–0.8 cm long. Leaf leathery, elliptic, oblong-lanceolate, 4.2–7.4 × 2.3–3.8 cm, apex acute or obtuse, mucronate, base tapering or narrowly cuneate, margin slightly revolute and glabrous; adaxial surface green with minute and transparent scales, somewhat shining, hairless; abaxial surface usually gray-white, densely scaly, scales one to four times their own diameter apart, pale brown or brownish-red; midrib sunk adaxially, conspicuously raised abaxially, and lateral veins sunk adaxially but not obvious abaxially. Inflorescences terminal, umbellate, 3-flowered, sometimes 2- or 4-flowered; pedicel 0.5–1 cm long, densely scaly, hairless. Calyx 5-lobed, lobes 0.5–1 × 0.5–0.8 mm, rounded or triangular, outside densely to moderately scaly, margins glabrous. Corolla tubular-funnelform, 5.9–7.5 × 4.7–7 cm, narrowly tubular near the middle and lower part of corolla, 5-lobed; corolla lobes white, with yellow markings inside; corolla tubes slender, 2.9–3.9 × 0.5–1.2 cm, white flushed with pink and not scaly but pubescent outside, glabrous with yellow markings inside. Stamens 10, unequal, 3.7–5.6 mm long; filaments densely pubescent at the base. Ovary ovoid, 5-locular, 0.7–1.2 mm long, densely scaly, hairless. Styles scaly and pubescent near the base, curved upwards, extend beyond the corollas. Capsule ovoid, 1.4–2.1 × 0.4–0.7 cm, densely scaly, hairless, calyx persistent.
Phenology.
Flowering from March to April. Fruiting from July to August.
Etymology.
The specific epithet of this newly described species refers to its type localities at Guangxi Mulun National Nature Reserve, part of the South China Karst landscape registered in UNESCO’s World Heritage List.
Vernacular name.
Simplified Chinese: 木论杜鹃; Chinese pinyin: mù lùn dù juān.
Distribution and ecology.
This new species is known from five localities to date. Three localities are fragmentally distributed in the same province, including two localities in Huanjiang County, Hechi, NW Guangxi, and one smaller locality at Molu Mountain (105°52'E, 23°23'N) in Napo County, Baise, SW Guangxi. The remaining two localities were distributed at Dongli (107°59'E, 25°22'N) in Mao-lan National Nature Reserve, Libo County, S Guizhou (Fig. 3). The plants were found on limestone under sparse, mixed, broad-leaf forest on the karst mountaintop (Figs 2A, 4A) at elevations from 640 to 1350 m (Fig. 3). The associated plants include Calocedrus rupestris Aver., T.H.Nguyên & L.K.Phan (Cupressaceae), Zanthoxylum dimorphophyllum Hemsl. (Rutaceae), Platycarya strobilacea Siebold & Zucc. (Juglandaceae), Polygala wattersii Hance (Polygalaceae), Acer sycopseoides Chun (Sapindaceae), Alyxia schlechteri H.Lév. (Apocynaceae), Leptopus pachyphyllus X.X.Chen (Phyllanthaceae), Rhododendron liboense Zheng R.Chen & K.M.Lan (Ericaceae), Paphiopedilum micranthum Tang & F.T.Wang (Orchidaceae), and Ophiorrhiza japonica Blume (Rubiaceae).
The known geographical distribution of Rhododendron mulunense in Guangxi and Guizhou Province, SW China (two localities at Libo in Guizhou are superimposed). Photograph by Xing-Xing Mao.
Comparisons of Rhododendron mulunense sp. nov. (A–D) and R. wumingense (E–H). A, E. Habitat; B, F. Flowers per inflorescence; C, G. Abaxial and adaxial leaf surface; D, H. Micrograph of the scales on the abaxial leaf surface. Photographs by Yu-Song Huang and Xing-Xing Mao.
Systematic position in Rhododendron.
Subsect. Maddenia Sleumer, Sect. Rhododendron, Subgen. Rhododendron.
Conservation status.
According to multiple fieldwork expeditions for Rhododendron plants around the karst mountains of Guangxi and Guizhou, five subpopulations or localities of R. mulunense have been discovered. Fewer than 30 mature individuals were found at two localities of less than 100×100 m^2^ in Huanjiang County, nine mature individuals were found at Molu mountain, and approximately 150 mature individuals were found in two localities at Mao-lan National Nature Reserve. Based on our investigations, the habitat quality and population size of R. mulunense have a continuing downward trend. Although two subpopulations, or all individuals at type localities from Huanjiang, are protected well, there are severe threats in other localities. Two subpopulations are situated in very small rocky hills in Mao-lan, with limited area encircled by urban roads, residential communities, and farmlands. Another subpopulation faces more severe threats in Napo, including its few mature individuals, limited distribution, and degraded habitat due to deforestation and wind power projects. Based on these findings, the conservation status of this new species should be assessed as Endangered (EN) under the IUCN Red List criteria C2a(i); D (Gibbs et al. 2011; IUCN 2012). This recommendation reflects the extremely limited distribution and very small population sizes of R. mulunense, combined with an observed continuing decline in numbers of mature individuals resulting from its close association with human activities and vulnerability to habitat loss.
Conservation studies of Rhododendron species indicated that the member botanic gardens of BGCI (Botanic Garden Conservation International, Richmond, UK) and the Global Conservation Consortium for Rhododendron play important roles in the conservation of species of subsect. Maddenia (BGCI 2020; Hu et al. 2024). Notably, a study focusing on conservation of Maddenia species in global botanic gardens suggested that ex situ living collections are urgently required to conserve wild endemic Maddenia species in countries of origin, and broader sampling of wild populations should be conducted for these threatened species (Hu et al. 2024). We propose extending the ecogeographical range of wild collections of R. mulunense and establishing ex situ collections in three member gardens of BGCI in China (Kunming Botanical Garden, Guilin Botanical Garden, and Lushan Botanical Garden). Meanwhile, given the threats to this endangered species, we urge local authorities to carry out habitat restoration and protection of the subpopulations in Mao-lan and Napo.
Additional specimens examined.
China. Guangxi Zhuang Autonomous Region: Hechi City, Huanjiang County, Chuanshan Town, Mulun National Nature Reserve, alt. 641 m, 3 March 2021, Y. Qin et al. GXQY20210303022 (IBK), GXQY20210303023 (IBK), GXQY20210303024 (IBK, IBSC), GXQY20210303025 (IBK, IBSC), GXQY20210303026 (IBK, PE), GXQY20210303027 (IBK, PE), GXQY20210303028 (IBK, PE), GXQY20210303029 (IBK); the same town, Jianfulun, alt. 1000 m, 22 April 2025, J.H. Liang, M.L. Mo & Q.M. Wei YB0225 (IBK); the same town, Zhonglun, alt. 970 m, 29 March 2025, Y.S. Huang Y25032906 (IBK); the same locality, elev. 980 m, 23 April 2025, J.H. Liang, M.L. Mo & Q.M. Wei YB0264 (IBK).
Notes.
The species of Rhododendron subsect. Maddenia show high interspecific diversification in morphological traits, including whether shoots and leaves have indumentum, leaf shape and size, scale density on the abaxial leaf surface, corolla shape and size, indumentum and scales on the corolla and tube exterior, and the color of the corolla and tube exterior (Min and Fang 1990; Fang et al. 2005). Some of these divergences between species may be attributed to heterogeneous habitat conditions. In south China, certain Maddenia species growing in dry conditions or subhumid habitats generally have hairless shoots and leaves, such as R. pachypodum Balf.f. & W.W.Sm., R. liliiflorum Levl., and R. kiangsiense Fang, while the shoots and leaves of species growing in moister or humid conditions tend to be covered with setose hairs; for example, R. wumingense Fang, R. ciliicalyx subsp. lyi (Levl.) R.C.Fang, and R. levinei Merr. (Table 1). Therefore, species distributed across different conditions or habitats have distinct morphological traits in subsect. Maddenia. Meanwhile, this indicates that habitat conditions can be used to discriminate specific species in this group.
Among the morphological traits mentioned above, the presence or absence of indumentum on shoots and leaves, the shape and size of the corolla and tube, the presence or absence of indumentum and scales on the corolla and tube exterior, the presence or absence of indumentum on calyx margins, the size of the calyx lobe, and the scale density on the abaxial leaf surface are the most taxonomically important. Both the morphological characteristics and geographic distribution of R. mulunense are most similar to those of R. wumingense, from which it can nevertheless be readily distinguished (Figs 2, 4; Table 1). Moreover, R. mulunense grows on limestone under subhumid conditions with sparse forest cover (Figs 2A, B, 4A), while R. wumingense is found on granite rocks under moister conditions in dense forest on montane slopes (Fig. 4E) or on open wet cliffs.
To assess whether it should be recognized as a new species, we compared R. mulunense with other similar species (Fig. 5; Table 1). Morphologically, this newly discovered species is similar to R. pachypodum and R. dendricola Hutch.: it has oblong-lanceolate leaves, an undeveloped calyx, and white flowers. However, it can be distinguished from these species by its tubular-funnelform corolla with an obviously slender tube lacking scales. R. pachypodum has a later flowering period, and R. dendricola has a narrow, non-overlapping distribution in the Himalaya–Hengduan Mountains, where it generally grows among conifers (Gibbs et al. 2011). In terms of the number of flowers per inflorescence and geographical distribution, R. mulunense is similar to other species of subsect. Maddenia, including R. ciliicalyx subsp. lyi, R. liliiflorum (including the synonym R. chunienii Chun and W.P.Fang), R. levinei, and R. kiangsiense (Li 2008; Geng 2004, 2014; Yang et al. 2022), but it can be readily distinguished by its tubular-funnelform corolla with the tube pubescent and not scaly, its undeveloped calyx, and an earlier flowering period.
Photographs of other species similar to Rhododendron mulunense. A.R. dendricola; B.R. pachypodum; C.R. ciliicalyx subsp. lyi; D.R. liliiflorum; E.R. levinei; F.R. kiangsiense. Photographs by Xing-Xing Mao, Yu-Song Huang, De-Tuan Liu, and Bing-Hua Chen.
Hu et al. (2023) showed that there have been polyploidization events in multiple species within subgen. Rhododendron, which could result in phenotypes distinct from their diploid progenitors. Nevertheless, polyploidization seldom occurs in subsect. Maddenia, and all related species, including R. wumingense, R. pachypodum, R. dendricola, R. ciliicalyx, R. liliiflorum, R. levinei, and R. kiangsiense, have been confirmed as diploids (Hu et al. 2023). Based on these findings, our study concludes that R. mulunense represents a new species in subsect. Maddenia of subgen. Rhododendron. Detailed comparisons of morphology, phenology, and distributions of R. mulunense and related species are summarized in Table 1. In addition, an identification key is provided to facilitate discrimination of R. mulunense from similar Maddenia species.
Key to Rhododendron
mulunense and similar Maddenia species
**: **
Supplementary Material
XML Treatment for Rhododendron mulunense
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1BGCI (2020) Compilation of ex situ collection records of Rhododendron from global botanic gardens. Unpublished report. Botanic Garden Conservation International, Richmond, UK.
- 2Brown GK Craven LA Udovicic F Ladiges PY (2006) Phylogeny of Rhododendron section Vireya (Ericaceae) based on two noncoding regions of cp DNA.Plant Systematics and Evolution 257: 57–93. 10.1007/s 00606-005-0367-8 · doi ↗
- 3Chamberlain DF Hyam R Argent G Fairweather G Walter KS (1996) The genus Rhododendron: its classification and synonymy. Royal Botanic Garden Edinburgh, Edinburgh.
- 4Chang YH Yao G Neilsen J Liu DT Zhang L Ma YP (2021) Rhododendron kuomeianum (Ericaceae), a new species from northeastern Yunnan (China), based on morphological and genomic data.Plant Diversity 43: 292–298. 10.1016/j.pld.2021.04.00334485771 PMC 8390914 · doi ↗ · pubmed ↗
- 5Chen WY Fang WP (1957) Materials for the flora of south China: New species of Gwangsi plants.Zhiwu Fenlei Xuebao 6(2): 159–173.
- 6Chen ZR Lan KM (2003) Rhododendron liboense, a new species of the Ericaceae from Guizhou, China.Zhiwu Fenlei Xuebao 41(6): 563–565. 10.1071/SB 00005_CO · doi ↗
- 7Chen X Huang JY Consaul L Chen X (2010 a) New taxa of Rhododendron (Ericaceae) from China.Annales Botanici Fennici 47(5): 397–402. 10.5735/085.047.0510 · doi ↗
- 8Chen X Huang JY Consaul L Chen X (2010 b) Two new species of Rhododendron (Ericaceae) from Guizhou, China.Novon 20: 386–391. 10.3417/2008102 · doi ↗