First country record of Metopiellus Raffray, 1908 (Staphylinidae, Pselaphinae, Metopiasini) from Ecuador, with description of two new species

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1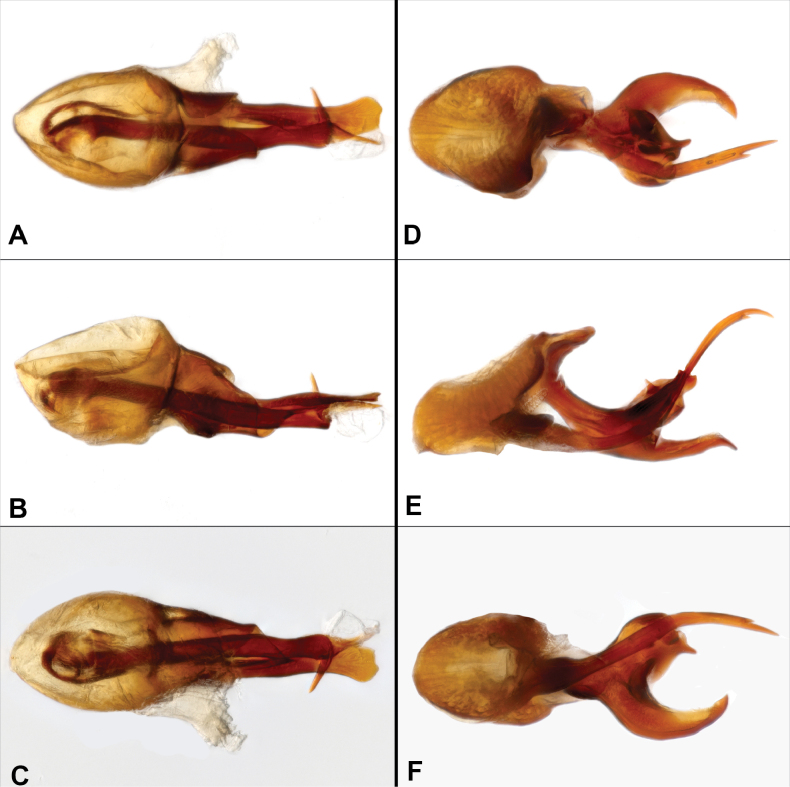 Figure 2
Figure 2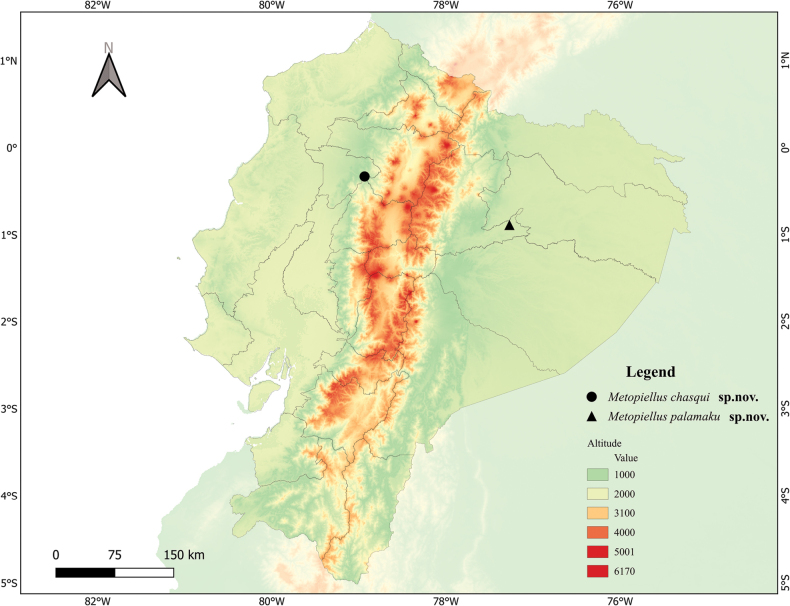 Figure 3
Figure 3| 1 | Vertex with projections; pronotum with two pairs of mediolateral thick, spinose projections, either simply convex or with a series of well-defined tumuli |
|
| – | Vertex without projections |
|
| 2 | Vertex with one horn-like projection |
|
| – | Vertex with a pair of horn-like projections |
|
| 3 | Pronotum with one pair of mediolateral, thick, spinose projections (Fig. |
|
| – | Pronotum with two pairs of mediolateral, thick, spinose projections; basal elytral fovea absent |
|
| 4 | Vertexal projections horn-like, apically acute; pronotum with two pairs of mediolateral, thick, spinose projections and one lateral process on each side; sternite VI with a pair of mediolateral setose processes and tergite VIII with a medial subquadrate thick process in the male |
|
| – | Vertexal projections forming transverse pair of ridge-like projections (Fig. |
|
| 5 | Head similar in width to pronotum; eyes absent |
|
| – | Head narrower than pronotum; eyes small, but conspicuous or very reduced, inconspicuous |
|
| 6 | Antennomere II about half as long as scape; antennomere V almost as long as antennomeres III and IV together |
|
| – | Antennomere II less than half as long as scape; antennomere V shorter than the length of antennomeres III and IV together |
|
| 7 | Antennomere VII rounded; dorsolateral margins strongly constricted anterad the eyes; paramere with apex bifurcated; median lobe curved and edge with long line of small teeth |
|
| – | Antennomere VII rectangular; dorsolateral margins of head gradually converging anterad the eyes; paramere not bifurcated apically; median lobe almost straight, with the apex curved, without small teeth along its length |
|
| 8 | Antennomere VIII transverse; eyes small, but conspicuous |
|
| – | Antennomere VIII obconical; eyes very reduced, inconspicuous |
|
| Species |
|
|
|
|
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1.75 | 2.91 | 1.00 | 0.86 | 1.00 | 0.58 | 0.55 | 0.26 | 0.58 | 0.64 | |
| 1 | 1.72 | 2.52 | 0.98 | 0.83 | 0.98 | 0.58 | 0.49 | 0.30 | 0.63 | 0.67 | |
| 1 | 2.16 | 3.24 | 1.07 | 0.94 | 1.07 | 0.56 | 0.56 | 0.25 | 0.52 | 0.70 | |
| 1 | 2.02 | 3.24 | 1.03 | 0.96 | 1.03 | 0.64 | 0.56 | 0.24 | 0.58 | 0.67 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsColeoptera Taxonomy and Distribution · Coleoptera: Cerambycidae studies · Spider Taxonomy and Behavior Studies
Introduction
Metopiasini is a tribe of Pselaphinae rove beetles suspected to be myrmecophiles (Parker 2016). This tribe comprises nine genera, eight of which are restricted to the Neotropical region (Mário Chaul and Lopes-Andrade 2024). One of these Neotropical genera is Metopiellus Raffray, 1908, which is distributed from Colombia to Argentina (Fiorentino et al. 2022; Mário Chaul and Lopes-Andrade 2024). This genus is composed of seven described species, five of them (M. aglenus Reitter, 1885, M. hirtus Reitter, 1885, M. painensisAsenjo et al., 2017, M. crypticusAsenjo et al., 2023, M. emavieirae Mário Chaul & Lopes-Andrade, 2024) from Brazil, one from Colombia (M. guanano Fiorentino, Tocora & Ramirez, 2022), and a final one from Argentina (Metopiellus silvaticus Bruch, 1933). Metopiellus is distinguished by a second antennomere much longer than the third, protibia carinated anteromedially, metacoxae contiguous or nearly so, and pronotum without sharp spines on the lateral edge, at most having thickened spinose protuberances mediolaterally on the pronotal disc (Mário Chaul and Lopes-Andrade 2024).
Until now, Metopiellus has not been reported from Ecuador. Herein, we officially describe two species of this genus, from both sides of the Andes: M. palamaku sp. nov. from an Amazon ecosystem in eastern Ecuador and M. chasqui sp. nov. from a montane cloud forest in western Ecuador.
Material and methods
Some specimens of the new species were obtained by sifting forest-floor leaf litter from the Minga Reserve in Napo Province, Ecuador (0.88853°S, 77.26833°W) and from the Otongachi Reserve in Pichincha Province, Ecuador (0.330510°S, 78.934420°W). One was hand-collected at Tiputini Biodiversity Station (0.6376°S, 76.1499°W) under permit FAUNA (X) 0020-MA-DPO-PNY (Orellana Province). Sifted litter was processed using Winkler extractors, with specimens collected, stored in 95% EtOH, and finally deposited in the National Biodiversity Institute collection (MECN) and the Pontificia Universidad Católica del Ecuador (QCAZ). Genitalia were cleared using 10% KOH, rinsed in water, and stored in glycerin in microvials pinned beneath the specimens. Photos were taken with a Macropod Pro photographic system (Macroscopic Solutions LLC), using a Canon MP-E 1–5X macro lens, combining the resulting stacks with Zerene Stacker (Richland, WA, USA).
Species recognition was based on unique combinations of external and male genitalic characters. The abbreviations used in the text and figures are: AL—abdomen length, BL—body length (from the anterior margin of the prolongation of the head to the posterior margin of tergite VIII), BW—body width (maximum width of elytra in dorsal view), EL—elytral length (maximum in dorsal view), EW—elytral width (maximum in dorsal view), HL—head length (from the anterior margin of the prolongation of the head to the posterior margin of the head disc in dorsal view), HW—head width (maximum, including eyes, in dorsal view), NW—neck width (minimum in dorsal view), PL—pronotum length (maximum in dorsal view), PW—pronotum width (maximum in dorsal view).
Results
Key to species of Metopiellus
Note: Adapted from Asenjo et al. (2017), Fiorentino et al. (2022), and Mário Chaul and Lopes-Andrade (2024). See those references for additional images and illustrations.
**: **
Taxonomy
Metopiellus
palamaku
sp. nov.
Taxon classificationAnimaliaColeopteraStaphylinidae
ECADB036-E18D-5186-866F-C8C250313EE4
https://zoobank.org/F67D4393-8C7F-4839-B576-FFC41B318BDD
Type material.
Holotype ♂ (MECN-EN 23780): “Ecuador. Napo: Chontapunta, R. Minga, -0.88692, -77.27049, 300 m. Winkler. 07-feb-2023, Díaz-Guevara & N. Berrazueta” / “MECN-EN 23780”; deposited in MECN.
Additional, non-type material.
Ecuador • 1 ♀; Orellana; 0.6376°S, 76.1499°W; Tiputini Biodiv. Sta.; 2-9.vi.2011; general hand collecting, AT1341, M. Caterino, A. Tishechkin; Caterino DNA voucher, Ext. MSC-12663; ZSFQ-i16787 • 1 ♀; Panayacu, Yasuní, 0°42'17.6"S, 76°40'15.7"W; 340 m; Winkler; 17 Jul. 2021; Pazmiño-Palomino; MECN-EN 48730.
Comparative diagnosis.
Metopiellus palamaku sp. nov. strongly resembles M. guanano, having a vertex with a single horn-like projection. However, M. palamaku can be easily differentiated by its single pair of mediolateral, thick, spinose pronotal projections (vs. two pairs in M. guanano), the presence of basal elytral foveae (absent from M. guanano), and by aedeagal characters including a more elongate basal bulb, and differently shaped articulated armature (Fig. 2), bifid at the tip with subapical hook bent at ~90° (vs. aedeagal armature simply tapered, slightly sinuate at apex in M. guanano; aedeagal armature with four spatulate, curved tips in M. emavieirae).
Description.
Avg. BL = 2.7 mm (N = 2; see Table 1 for other measurements). Body (Fig. 1A–C) densely setose and brown; eyes prominent, comprising ~11 ommatidia; head pyriform with two vertexal foveae near posterior margin with a single median process similar to M. guanano; antennae with 11 antennomeres, antennal scape base thick and slightly narrowed before the apex (in dorsal view), antennomere I as long as rest of the antennomeres combined, antennomere II about as long as III–VIII combined, III-IV and VI slightly longer than wide, V distinctly longer, VII globular, VIII small, IX and X slightly transverse, XI almost as long as IX–X combined, tapered in apical half; pronotum trapezoidal, wider anteriorly and narrower posteriorly, with two pairs of mediolateral, thick, spinose projections; each elytron with two basal foveae, and with longitudinal impressions extending from them posteriorly approximately two-thirds elytron length. Aedeagus (Fig. 2A–C) with large, elongate basal bulb, slightly asymmetrical, with smoothly rounded basal margin; dorsal diaphragm large, elongate oval; tegmen narrowed toward flat, paddle-shaped apex; elongate armature articulated within basal bulb, apex bifid, with acute, subapical hook bent at ~90°, other tip slightly diverging to left, acute.
Habitus of new Metopiellus species. A–C.M. palamaku sp. nov. (female dorsal, female lateral, male holotype ventral); D–F.M. chasqui sp. nov. (male holotype dorsal, female paratype head + pronotum, male non-type head only). Scale bars: 1 mm (unless otherwise indicated).
Aedeagus of new Metopiellus species (dorsal, lateral, ventral). A–C.M. palamaku sp. nov.; D–F.M. chasqui sp. nov.
Distribution.
This species is known from the type locality in Chontapunta, Ecuador (Fig. 3), and probably also from the Tiputini region of Orellana (if the associations of the females under ‘other material’ are correct). The sites are separated by about 125 km, but are all on the same (south) side of the Rio Napo.
Map of Ecuador showing type localities of the two new species.
Etymology.
The specific name derives from a Kichwa legend in which Palamaku is the divine source of insects.
Metopiellus
chasqui
sp. nov.
Taxon classificationAnimaliaColeopteraStaphylinidae
FABB52E7-EA8A-580E-8CCD-D2A5995C0AFA
https://zoobank.org/636C2C0D-06BF-4B70-A4C3-4B1DA9D2F850
Type material.
Holotype ♂ (QCAZ 280376): “Ecuador. Pichincha: Otongachi Reserve -0.330510, -78.934420, 1087m.20-Feb-2020, E. Tapia & N. Dupérré”; deposited in QCAZ. Paratypes (4 ♀, same general locality as type): -0.330510, -78.934420, 1087 m (QCAZ 280377 to 280380).
Additional, non-type material.
Ecuador • 4 ♂, 4 ♀: Zamora Chinchipe; Las Orquídeas; -4.2482, -78.6595; 877 m; E. Tapia; Sifting litter; 29-Jul-2024; QCAZ 280381 to 280388.
Comparative diagnosis.
Metopiellus chasqui resembles M. emavieirae in the presence of a pair of (rather than a single) vertexal structures and by having two pairs of mediolateral pronotal projections. However, it can be distinguished from it by the vertexal and median pronotal processes being transversely ridge-like, and by the lateral pronotal processes having a posterior acute point (vs. horn-like vertexal and pronotal structures in M. emavieirae). The aedeagi differ strongly, with that of M. chasqui having a strongly bifurcate dorsal armature, and that of M. emavieirae being subdivided subapically into four curved, spatulate tips.
Description.
Avg. BL = 3.2 mm (N = 2; see Table 1 for other measurements). Body (Fig. 1D–F) densely setose and brown-reddish; eyes prominent, borne on small elevated base, comprising ~8 ommatidia; head widest at base, rounded, narrowed anterad, with two posterior vertexal foveae (Fig. 1E) immediately in front of two low posterior vertexal ridges; antennae with 11 antennomeres, antennal scape about as long as antennomeres II–VIII combined, antennomere II as long as III–VII combined, III–VI elongate, V longer than others, VII globular, VIII smallest, IX–XI forming weak club, IX and X transverse, XI almost as long as IX and X combined, tapered in apical two-thirds; pronotum densely setose, with 4 acute lobes, lateral lobes each with posterior acute point, median ones forming oblique ridges, prominent at their inner corners; each elytron with two basal foveae, and with longitudinal impressions extending from them posteriorly approximately two-thirds elytron length; protibia rather slender. Aedeagus (Fig. 2D–F) with basal bulb asymmetrical and globose, with basal rounded margin, elevated distad, dorsal diaphragm elongate oval; tegmen with ventral portion long, slender, curved dorsad and then distad apically; articulated armature robust, deeply bifurcated, one bifurcation is hook-like, longer and evenly tapered, other more robust with small projection oriented towards distal apex of basal bulb.
Distribution.
This species probably only occurs in the Otongachi Reserve in northern Ecuador, in the province of Pichincha (Fig. 3). We believe the ‘additional material’ cited from Zamora-Chinchipe to be mislabeled, since that locality is over 400 km to the south, and located on the Amazonian side of the Andean crest, an unlikely distribution for a flightless beetle species. The male genitalia is identical between the two alleged localities.
Etymology.
The specific epithet “chasqui” refers to the agile messengers of the Inca Empire, evoking the beetle’s slender posterior legs.
Discussion
Metopiellus now contains nine known species. These two new species extend the geographic range of the genus into Ecuador. The closest previous record was from Amazonian Colombia, while the bulk of the species are known from eastern and southeastern Brazil. Given the existence of the Colombian species (M. guanano), a record of the genus from the Ecuadorean Amazon shouldn’t be too surprising. However, the new species from the cloud forests of the western slope of the Andes represents an important extension not only geographically but also ecologically. Clearly, further sampling of leaf-litter habitats will result in continued discovery of new species across the Neotropical region.
Mário Chaul and Lopes-Andrade (2024) considered their M. emavieirae to be closely related to M. guanano, but our M. palamaku is even closer to M. guanano, differing almost exclusively in fairly minor aedeagal characters. So if species groups are to be delineated, the three should be grouped together. Metopiellus chasqui also shares the horn-like projections of the vertex that these three do, and would appear to be closely related as well, although its aedeagus is radically different from any of the others. Of these four, only M. emavieirae exhibits any significant male secondary sexual characters, with projections of the terminal tergite and sternites.
We have little to contribute to speculation on the general natural history of species in the genus, with all our specimens coming from fairly generic leaf-litter samples. Early records of some species associated with ants’ colonies have not been repeated, and the collection of one Metopiellus species (M. painensis; Asenjo et al. 2017) from caves suggests something rather different: that members of the genus may have subterranean propensities, habits that would be consistent with their generally reduced eyes and elongated appendages. We can only hope that future direct collections take closer note of more significant natural history associations to further illuminate the biology of these impressive pselaphine beetles.
Supplementary Material
XML Treatment for Metopiellus palamaku
XML Treatment for Metopiellus chasqui
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Asenjo A Ferreira RL Zampaulo RDA (2017) Description of Metopiellus painensis sp. nov. (Coleoptera, Staphylinidae), first troglobitic Pselaphinae from Brazil.Zootaxa 4269(1): 115–123. 10.11646/zootaxa.4269.1.528610342 · doi ↗ · pubmed ↗
- 2Asenjo Ade Oliveira MPA Oliveira RRM Pires ES Valois M Oliveira G Vasconcelos S (2023) The complete mitochondrial genome and description of a new cryptic Brazilian species of Metopiellus Raffray (Coleoptera: Staphylinidae: Pselaphinae). Peer J 11: e 15697. 10.7717/peerj.15697 PMC 1038723137525659 · doi ↗ · pubmed ↗
- 3Fiorentino G Tocora MC Ramirez S (2022) A new species of Metopiellus (Coleoptera, Staphylinidae, Pselaphinae) from the northern Colombian Amazon.Zoo Keys 1108: 1–9. 10.3897/zookeys.1108.7607736760694 PMC 9848935 · doi ↗ · pubmed ↗
- 4Mário Chaul JC Lopes-Andrade C (2024) A new species of Metopiellus Raffray (Coleoptera: Staphylinidae: Pselaphinae) from the Atlantic Forest of Brazil, with keys to species of the genus and to the genera of Metopiasini.Studies on Neotropical Fauna and Environment 59(3): 1114–1128. 10.1080/01650521.2024.2333103 · doi ↗
- 5Parker J (2016) Myrmecophily in beetles (Coleoptera): Evolutionary patterns and biological mechanisms.Myrmecological News 22: 65–108. 10.25849/myrmecol.news_022:065 · doi ↗