In vitro selection of cefiderocol-resistant mutants in Acinetobacter baumannii harbouring the most common carbapenemase genes
Otávio Hallal Ferreira Raro, Jacqueline Findlay, Laurent Poirel, Jean-Winoc Decousser, Patrice Nordmann

TL;DR
This study shows how Acinetobacter baumannii can develop resistance to the antibiotic cefiderocol through mutations in global regulators that affect iron uptake genes.
Contribution
The study identifies BfmS and OxyR as novel global regulators linked to cefiderocol resistance via reduced expression of iron uptake genes.
Findings
Mutations in BfmS and OxyR were found in cefiderocol-resistant isolates.
Complementation with wild-type BfmRS or OxyR reversed resistance.
BfmS and OxyR reduced expression of piuA and pirA, linking to resistance.
Abstract
Carbapenem-resistant Acinetobacter baumannii (CRAB) represents a critical global health threat due to its high mortality rates in infections, limited treatment options and resistance to last-resort antibiotics. Cefiderocol, a novel siderophore-conjugated cephalosporin, has emerged as a promising therapeutic agent against such infections. However, resistance to this antibiotic has already been reported. To investigate whether the production of common carbapenemases affects cefiderocol susceptibility in A. baumannii and to identify the genetic mechanisms underlying resistance development. Using isogenic A. baumannii CIP 7010 strains carrying carbapenemase genes commonly identified in that species, namely blaNDM-1, blaOXA-23, blaOXA-40 or blaOXA-58, mutants were selected under increasing cefiderocol pressure. Mutants obtained were subjected to MIC determinations, WGS, complementation…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
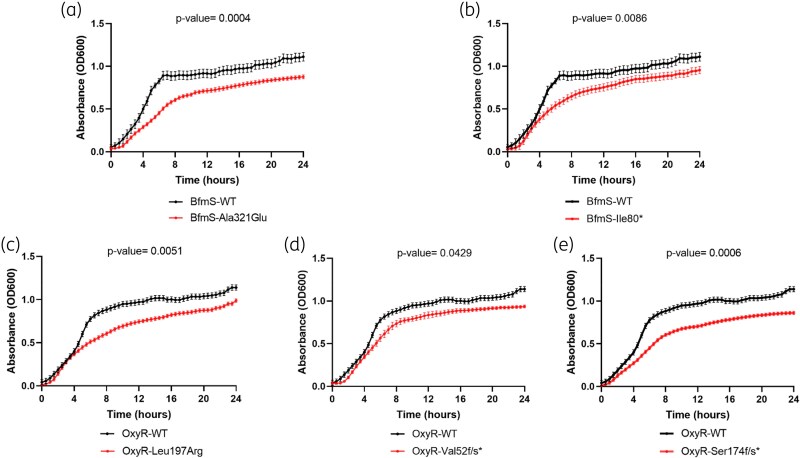 Figure 1
Figure 1| Strain | FDC selection, mg/L | Protein sequence changes | MIC, mg/L | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BfmS | OxyR | FDC | PIP | CAZ | FEP | IPM | MEM | SAM | SUL/DUR | CST | TET | CIP | AMK | ||
| CIP 7010 | — | — | — | 0.12 | 16 | 1 | 1 | ≤ | 0.25 | 2 | ≤0.12 | ≤0.12 | 0.5 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 | — | — | — | 0.12 | 16 | 1 | 1 | 0.12 | 0.25 | 2 | 0.5 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 | 8 | Ala321Glu | — | >128 | 16 | 1 | 1 | 0.12 | 0.25 | 2 | 0.5 | ≤0.12 | 0.5 | 0.25 | 1 |
| CIP 7010 + pVRL1 + NDM-1 | — | — | — | 1 | 128 | >128 | 32 | 4 | 8 | 16 | 8 | ≤0.12 | 0.5 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + NDM-1 | 8 | Ala321Glu | — | >128 | >128 | >128 | 32 | 8 | 8 | 16 | 8 | ≤0.12 | 0.5 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + OXA-23 | — | — | — | 0.12 | >128 | 2 | 2 | 2 | 2 | 8 | 1 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + OXA-23 | 8 | — | Leu197Arg | >128 | >128 | 2 | 2 | 2 | 2 | 8 | 1 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + OXA-40 | — | — | — | 0.12 | >128 | 2 | 2 | 0.5 | 1 | 8 | 1 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + OXA-40 | 64 | — | Val52f/s | >128 | >128 | 2 | 2 | 0.5 | 1 | 8 | 1 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + OXA-58 | — | — | — | 0.12 | >128 | 2 | 2 | 0.5 | 1 | 8 | 1 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + OXA-58 | 8 | Ile80 | — | >128 | >128 | 2 | 2 | 0.5 | 1 | 8 | 1 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| CIP 7010 + pVRL1 + OXA-58 | 64 | — | Ser174f/s | >128 | >128 | 2 | 2 | 0.5 | 1 | 8 | 1 | ≤0.12 | 0.25 | ≤0.12 | 1 |
| Strain | Protein sequence changes | FDC MIC, mg/L |
|---|---|---|
| pVRL1–8 mg/L | BfmS—Ala321Glu | >128 |
| pVRL1–8 mg/L/pUBYT | >128 | |
| pVRL1–8 mg/L/pUBYT- | 1 | |
| OXA-58–8 mg/L | BfmS—Ile80 | >128 |
| OXA-58–8 mg/LpUBYT | ≥128 | |
| OXA-58–8 mg/LpUBYT- | 2 | |
| OXA-23–8 mgL | OxyR—Leu197Arg | >128 |
| OXA-23–8 mg/L/pUBYT | ≥128 | |
| OXA-23–8 mg/LpUBYT- | 0.5 | |
| OXA-40–64 mg/L | OxyR—Val52f/s | >128 |
| OXA-40–64 mg/LpUBYT | >128 | |
| OXA-40–64 mg/LpUBYT- | 0.5 | |
| OXA-58–64 mg/L | OxyR—Ser174f/s | >128 |
| OXA-58–64 mg/LpUBYT | ≥128 | |
| OXA-58–64 mg/LpUBYT- | 0.5 |
| Strain | FDC selection, mg/L | Protein sequence changes | 2−ΔΔCT | |
|---|---|---|---|---|
|
|
| |||
| CIP 7010 | — | — | 1 | 1 |
| pVRL1 | 8 | BfmS—Ala321Glu |
|
|
| OXA-58 | 8 | BfmS—Ile80 |
|
|
| OXA-23 | 8 | OxyR—Leu197Arg |
|
|
| OXA-40 | 64 | OxyR—Val52f/s |
|
|
| OXA-58 | 64 | OxyR—Ser174f/s |
|
|
- —University of Fribourg10.13039/501100005869
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotics Pharmacokinetics and Efficacy · Antibiotic Use and Resistance
Introduction
The ability of Acinetobacter baumannii to cause severe human infections, acquire antimicrobial resistance genes and persist in hospital environments, has led to carbapenem-resistant A. baumannii (CRAB) being classified as ‘critical’ in the WHO bacterial priority pathogens list for research and development of new medicines, diagnostics and prevention tools.^1^
CRAB strains are responsible for a wide range of infections including hospital- and community-acquired pneumonia, bloodstream infections, urinary tract infections and wound infections. These infections often progress to severe stages among intensive care patients, particularly in low- to middle-income countries, leading to alarmingly high mortality rates.^1–3^ CRAB was listed among the six leading pathogens associated with deaths due to antimicrobial resistance in 2019 and 2021, with attributable mortality of 57 700 and 78 100 deaths, respectively.^2,4^
Despite the critical threat posed by CRAB, the high morbidity and mortality, and the increased reports of resistance to clinically relevant antibiotics, therapeutic options remain severely limited, and antibiotic development has stagnated. Notably, no clinically effective agents are currently available against MBL-producing CRAB strains, underscoring the urgent need for sustained investment in antimicrobial research and development.^5,6^
The management of CRAB infections remains challenging due to the toxicity and limited efficacy of last-resort agents. Agents such as colistin and polymyxin B are frequently associated with nephrotoxicity, and tigecycline is associated with poor penetration in infection sites like the lungs and the urinary tract.^7^ The recently developed sulbactam/durlobactam combination shows promise; however it is ineffective against MBL-producing strains, leaving a critical therapeutic gap.^8^
These challenges highlight the urgent need for novel antimicrobial agents active against resistant A. baumannii. Cefiderocol, a siderophore-conjugated cephalosporin, represents a novel therapeutic approach that exploits the bacterial iron uptake systems. It binds extracellular iron and is actively transported into Gram-negative bacteria via catecholate siderophore receptors, while also diffusing through outer membrane porins thus enhancing access to target sites into the periplasmic space of the cell.^9^ Cefiderocol has been approved for treating adults with complicated Gram-negative urinary tract infections, hospital-acquired pneumonia and ventilator-associated pneumonia.^9,10^
However, infections caused by cefiderocol-resistant CRAB strains have already been reported. Acquired resistance has been associated with mutations in genes involved in iron acquisition, including pirA (siderophore gene) and piuA (TonB-dependent siderophore receptor), β-lactamases genes (NDM-like, SHV-12 and PER-like), and point mutations in PBP1a and PBP3.^11–14^ To better understand the influence of carbapenemase types on cefiderocol resistance in A. baumannii, our study aimed to select cefiderocol-resistant mutants from a collection of isogenic recombinant A. baumannii CIP 7010 isolates carrying the most frequently acquired carbapenemase genes in this species, namely blaNDM-1, blaOXA-23, blaOXA-40 or blaOXA-58, and to identify the mechanisms responsible for resistance development.
Materials and methods
Construction of the A. baumannii CIP 7010 library containing carbapenemase genes
The strain A. baumannii CIP 7010 (ATCC 15151) was selected for this study because it is a fully sequenced, antibiotic-susceptible reference strain of A. baumannii commonly used in comparative genomics and resistance mechanism studies.^15^ To generate the isogenic recombinant strains, primers were used to amplify blaNDM-1, blaOXA-23, blaOXA-40 or blaOXA-58, the resulting products were ligated into the shuttle vector pVRL1,^16^ and transformed into Escherichia coli TOP10, using 15 mg/L gentamicin for selection. Plasmids were then extracted using Zyppy Plasmid Miniprep Kit (Zymo Research, Irvine, CA, USA) and transformed into A. baumanniii CIP 7010 by electroporation. Primers are shown in Table S1 (available as Supplementary data at JAC Online).
Susceptibility testing
Susceptibility testing was performed by broth microdilution using an in-house-prepared iron-depleted Mueller–Hinton broth (ID-MHB) to determine the MIC of cefiderocol. MHB was used for all the other antibiotics for all isolates according to the EUCAST guidelines.^17,18^ Interpretation of the results for cefiderocol was performed in accordance with the pharmacokinetic/pharmacodynamic breakpoints from EUCAST (susceptible ≤2 mg/L; resistant >2 mg/L).^17,18^ Since there is a lack of breakpoints available for A. baumannii, the reference strains E. coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853 and A. baumannii NCTC 13304^19^ were used for broth microdilution (BMD) quality control.
Selection of cefiderocol-resistant mutants
Cefiderocol-resistant A. baumannii mutants were selected in ID-MHB using several passages with increasing concentration of cefiderocol. Briefly, a single colony of each strain was first grown in LB broth overnight at 37°C without antibiotics. The strains were then inoculated in a 1:100 ratio in 5 mL of ID-MHB, with an initial concentration of cefiderocol of 0.5 mg/L, for 24 h. Then strains were plated onto LB agar plates, and two single colonies were recovered and stored at −80°C. Additionally, from the remaining broth with 0.5 mg/mL of cefiderocol, the strains were transferred (1:100) to 5 mL of fresh media containing double the initial concentration of cefiderocol. After that, both processes (single colony recovery and increasing cefiderocol concentration) were repeated every 24 h until no further bacterial growth was observed in both LB broth and agar.
WGS
Genomic DNA (gDNA) was isolated from overnight-grown bacterial cultures using the QIAamp DNA Mini Kit (Qiagen, Valencia, CA, USA). WGS was carried out using the Illumina MiSeq (San Diego, CA, USA) platform. Libraries were prepared with the Nextera DNA sample preparation protocol, generating 2 × 300 bp paired-end reads with a minimum depth of 30× coverage. Short-read data were quality-trimmed using Trimmomatic and assembled with the Shovill pipeline (https://github.com/tseemann/shovill).^20^ Resulting contigs were annotated using Prokka.^21^ Resistance gene detection and species confirmation were conducted via the Center for Genomic Epidemiology’s tools: ResFinder 4.1 and KmerFinder 3.2 (https://www.genomicepidemiology.org/).^22,23^ Snippy (https://github.com/tseemann/snippy) was used to search for substitutions such as SNPs and multiple nucleotide polymorphisms, and for insertions or deletions between the reference genome (A. baumannii CIP 7010 plus the pVRL1 empty vector) and the mutant isolates. Sequencing datasets from this study were deposited in the NCBI Sequence Read Archive under BioProject accession number PRJNA1338351.
bfmRS and oxyR cloning and complementation
Wild-type bfmRS and oxyR were amplified from A. baumannii CIP 7010 and cloned into pUBYT using the primers listed in Table S1,^24^ to generate plasmids pUBYT-bfmRS and pUBYT-oxyR. Both the original plasmid, pUBYT, and recombinant plasmids were electroporated into strains and mutants, with selection on LB agar supplemented with 50 mg/L kanamycin.
mRNA extraction and cDNA synthesis
Strains were inoculated in ID-MHB and incubated at 37°C with shaking until reaching the OD of 0.5. The total RNA was extracted using the Quick-RNA MiniPrep kit (Zymo Research, Irvine, CA, USA). To remove contaminating DNA from RNA preparations, the Turbo DNA-free kit (Invitrogen, Waltham, MA, USA) was used. cDNA synthesis was performed by using LunaScript RT SuperMix kit (New England BioLabs, Ipswich, MA, USA). cDNA sample concentrations were measured using the NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
RT-qPCR
Quantitative real-time PCR experiments were performed using the Rotor-Gene Q cycler (Qiagen, Hilden, Germany). The primers used in the experiments were targeting the genes rpoB (reference), piuA and pirA (primers are shown in Table S1). Reactions were set up in a total volume of 20 µL with a GoTaq qPCR Master Mix kit (Promega, Madison, WI, USA). The cycle threshold values were analysed by the 2^−ΔΔCT^ method.^25^ Relative expression levels were calculated by comparison with the control samples, and the condition values were corrected with the appropriate reference gene. Experiments were performed on three independent biological replicates with two technical replicates of each reaction. Data were analysed by one-way analysis of variance (ANOVA) corrected for multiple comparisons using statistical hypothesis testing with Tukey using GraphPad Prism Software version 9.5.1. Statistically significant results were defined as P value <0.05.
Fitness evaluation
Measurements of growth rates were performed with the A. baumannii CIP 7010 strain in comparison with the mutated versions of the respective genes obtained after in vivo selection with cefiderocol. Growth curves were obtained following standard protocols and performed in ID-MHB in 96-well microplates. Bacterial growth was accessed by measuring OD at 600 nm wavelength (OD_600_) using a BioTek Cytation 5 plate reader (Agilent Technologies, Santa Clara, CA, USA) over a 24 h period. Experiments were performed on three independent replicates with two technical replicates of each. Statistical analyses were conducted using two-tailed parametric t-tests, and statistical significance was defined as a P value <0.05. Analyses were performed using GraphPad Prism version 9.5.1.
Results and discussion
For cefiderocol mutants obtained from each genetic construct, three independent colonies were initially selected for sequencing and antimicrobial susceptibility testing. Colonies that displayed identical resistance profiles and mutations were considered equivalent and represented by only a single isolate or at most two per strain to avoid redundancy and facilitate data interpretation. According to this selection process and the variable selective endpoints, the following mutants were chosen: one mutant carrying the empty vector pVRL1 (pVRL1), one with pVRL1 harbouring blaNDM-1 (pVRL1 + NDM-1), one with pVRL1 harbouring blaOXA-23 (pVRL1 + OXA-23), and one with pVRL1 harbouring blaOXA-58 (pVRL1 + OXA-58a), all recovered at 8 mg/L of cefiderocol. Additionally, mutants carrying pVRL1 with blaOXA-40 (pVRL1 + OXA-40) and a second colony with blaOXA-58 (pVRL1 + OXA-58b), were selected at 64 mg/L of cefiderocol. The MIC values and corresponding mutations for all selected mutants are summarized in Table 1. The obtained mutations conferred resistance to cefiderocol without cross-resistance to the other antibiotics tested. This was particularly evident for antibiotics currently considered potential alternative treatments for infections caused by MDR A. baumannii, namely ampicillin/sulbactam, sulbactam/durlobactam and colistin according to the IDSA.^26^
Genes encoding the global regulators BfmS or OxyR were regularly altered across several cefiderocol-resistant mutants, regardless of the carbapenamase carried. Two distinct alterations were observed in BfmS across different parental stains: Ala231Glu (pVRL1-derived and NDM-1-derived) and Ile80-stop [stop codon; (OXA-58-derived)]. Similarly, three other differences were also observed in OxyR: Leu197Arg (OXA-23-derived), Val52f/s-stop [f/s, frameshift (OXA-40-derived)] and Ser174f/s-stop (OXA-58-derived). All six mutants harbouring mutations in either oxyR or bfmS exhibited cefiderocol MIC >128 mg/L (Table 1). The same types of mutations in the bfmS and oxyR regulatory genes were obtained regardless of the carbapenemase gene content. Therefore, the expression of the NDM gene did not influence the type of mutations obtained, although it has been reported as a favourable background for the development of reduced susceptibility to cefiderocol.^11^
Complementation assays were performed by introducing WT bfmRS or oxyR into one representative of each mutation, resulting in five cefiderocol-resistant mutants being tested to assess any impact on MICs of cefiderocol. In all five mutants a significant decrease in cefiderocol MICs to parental MICs was observed in most cases (Table 2). These results confirmed that the identified mutations directly impacted cefiderocol susceptibility. Notably, the presence of the different carbapenemase enzymes did not influence the level of cefiderocol resistance among the mutants.
RT-qPCR assays were then performed to evaluate the impact of these mutants on the expression of genes involved in iron acquisition in A. baumannii. The WT A. baumannii CIP 7010 strain served as a control, whereas one representative of each oxyR or bfmS mutation was analysed for the expression of the main iron transport genes, namely piuA and pirA. A statistically significant reduction in piuA and pirA expression was observed for all mutants compared with the control strain. Using A. baumannii CIP 7010 as the control (2^−ΔΔCT^: 1), piuA expression ranged from 0.03 to 0.07 in oxyR mutants and from 0.37 to 0.51 in bfmS mutants whereas pirA expression ranged from 0.09 to 0.37 and 0.12 to 0.42, respectively (Table 3).
These findings confirm that mutations in oxyR and bfmS indirectly promote cefiderocol resistance in A. baumannii by down-regulating the expression of a siderophore gene (pirA) and a TonB-dependent siderophore receptor (piuA), both previously associated with cefiderocol resistance.^13,14^ In contrast to prior reports describing direct substitutions in iron transporters piuA (Val216Gly, Thr280Ser, Asn381Ser, Ser412Thr) and pirA (Leu275Phe, I277Val),^27^ our study identified mutations in the regulatory genes oxyR and bfmS that indirectly reduce pirA and piuA expression. Similar mutations in iron transporters have also been linked to cefiderocol resistance in other species such as P. aeruginosa (e.g. piuA, piuC, pirA, pvdS) and Enterobacterales (e.g. cirA, fiuA).^28–30^
BfmS is a sensor kinase component of the two-component regulatory system BfmRS, which modulates virulence, antimicrobial resistance and environmental adaptation in A. baumannii.^31^ Geisinger et al.^32^ showed that A. baumannii lacking bfmS (ΔbfmS) exhibit increased resistance to several β-lactams, including aztreonam, ampicillin, carbenicillin, cephalexin, ceftazidime, mecillinam and sulbactam. However, in our study, bfmS mutants did not display increased MICs to β-lactams, suggesting no global cross-resistance effect. This supports the hypothesis that the observed increase in cefiderocol MICs is specifically related to impaired drug entry mediated by iron transport pathways involving piuA and pirA. Additionally, another two-component regulatory system, BaeSR, has been previously reported to mediate cefiderocol resistance.^33^ Liu et al.^33^ showed that BaeSR mutations promote resistance through strong up-regulation of the MacAB-TolC and an MFS efflux pump and activation of the phenylacetic acid operon (paa). Both BfmRS and BaeSR systems modulate envelope-stress responses, yet they employ distinct downstream effectors: whereas the first limits drug entry, the second promotes drug efflux, highlighting a parallel regulatory resistance mediation with mechanistic divergence.^32,33^
OxyR is a LysR-type transcriptional regulator that functions as a master activator of the oxidative stress response, detecting and detoxifying hydrogen peroxide (H_2_O_2_) via alkyl hydroperoxide reductase (Ahp) and catalase (Kat), converting H_2_O_2_ to water.^34^ In A. baumannii, OxyR contributes to survival, virulence and genetic mobility (integrons or IS elements) through SOS and DNA repair responses.^35,36^ Inactivation of OxyR can trigger compensatory activation of other regulators (SoxR, RpoS, MarA), restoring oxidative stress tolerance and enhancing antibiotic resistance through efflux, envelope remodelling or DNA repair mechanisms.^37,38^ Although no previous studies have shown a direct role of OxyR in regulating iron uptake in A. baumannii, OxyR in E. coli has been reported to induce transcription of dps, which encodes a ferritin-like protein that protects DNA by sequestering iron.^39^ Moreover, an OxyR ortholog in Cupriavidus pinatubonensis positively regulates the type VI secretion system (T6SS1) by binding its promoter region, promoting iron import under oxidative stress.^40^
Growth curve analyses revealed that mutations in either bfmS or oxyR significantly reduced bacterial fitness compared with WT strains (Figure 1). Although no direct evidence links BfmS or OxyR activation to iron availability in A. baumannii, our observations suggest that mutants harbouring these mutations may have an increased dependence on iron to maintain regulatory and stress-response pathways. Under limited iron availability in ID-MHB medium, these mutants are likely to face higher energetic demands to sustain regulatory activity, explaining their impaired growth.
Growth curves of the BfmS-WT and OxyR-WT Acinetobacter baumannii CIP7010 and mutant isolates. (a) BfmS-WT versus BfmS-Ala321Glu. (b) BfmS-WT versus BfmS-Ile80. (c) OxyR-WT versus OxyR-Leu197Arg. (d) OxyR-WT versus OxyR-Val52f/s*. (e) OxyR-WT versus OxyR-Ser174f/s*. Data are shown as mean ± SD. f/s, frameshift; , stop codon.
BfmR, the response regulator of the BfmRS system, is negatively regulated by BfmS and directly controls genes involved in siderophore biosynthesis and transport.^41^ Given the essential role of iron in cellular function, our findings align with the hypothesis that restricted iron acquisition diminishes cell fitness.^42^ Additionally, OxyR has been shown to influence bacterial fitness and virulence: A. baumannii ΔoxyR strains are less abundant and less capable of disseminating in the liver and lungs of infected mice compared with WT oxyR strains.^43^
Conclusions
Despite being crucial for metabolic and regulatory processes in A. baumannii, BfmS and OxyR have not been previously associated with resistance to cefiderocol. In this study, we demonstrated that the global regulators BfmS and OxyR are responsible for decreasing the expression of piuA and pirA genes, thereby contributing to cefiderocol resistance. This evolutionary analysis enhances our understanding of the mechanisms underlying cefiderocol resistance and identifies potential molecular targets for the development of new therapeutics against CRAB.
Supplementary Material
dkaf462_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization . 2024. Bacterial priority pathogens list, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. https://iris.who.int/bitstream/handle/10665/376776/9789240093461-eng.pdf?sequence=1
- 2Antimicrobial Resistance Collaborators . Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 2022; 399: 629–55. 10.1016/S 0140-6736(21)02724-035065702 PMC 8841637 · doi ↗ · pubmed ↗
- 3Jiang Y, Ding Y, Wei Y et al Carbapenem-resistant Acinetobacter baumannii: a challenge in the intensive care unit. Front Microbiol 2022; 13: 1045206. 10.3389/fmicb.2022.104520636439795 PMC 9684325 · doi ↗ · pubmed ↗
- 4GBD 2021 Antimicrobial Resistance Collaborators . Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. Lancet 2024; 404: 1199–226. 10.1016/S 0140-6736(24)01867-139299261 PMC 11718157 · doi ↗ · pubmed ↗
- 5O’Neill J. 2014. Antimicrobial resistance: tackling a crisis for the health and wealth of nations—review on antimicrobial resistance. https://amr-review.org/sites/default/files/AMR%20Review%20Paper%20-%20Tackling%20a%20crisis%20for%20the%20health%20and%20wealth%20of%20nations_1.pdf
- 6Patel J, Harant A, Fernandes G et al Measuring the global response to antimicrobial resistance, 2020–21: a systematic governance analysis of 114 countries. Lancet Infect Dis 2023; 23: 706–18. 10.1016/S 1473-3099(22)00796-436657475 · doi ↗ · pubmed ↗
- 7Kollef M, Dupont H, Greenberg DE et al Prospective role of cefiderocol in the management of carbapenem-resistant Acinetobacter baumannii infections: review of the evidence. Int J Antimicrob Agents 2023; 62: 106882. 10.1016/j.ijantimicag.2023.10688237301312 · doi ↗ · pubmed ↗
- 8Barnes MD, Kumar V, Bethel Moussa SH et al Targeting multidrug-resistant Acinetobacter spp.: sulbactam and the diazabicyclooctenone β-lactamase inhibitor ETX 2514 as a novel therapeutic agent. m Bio 2019; 10: e 00159-19. 10.1128/m Bio.00159-1930862744 PMC 6414696 · doi ↗ · pubmed ↗