mGem: Faecalibacterium, an important protector of gut health
Carlos Sabater, Xenia Vázquez, Sylvia H. Duncan

TL;DR
Faecalibacterium, a gut bacteria, produces butyrate and may protect against gut diseases, with potential for microbiome therapies.
Contribution
Genomic analysis reveals Faecalibacterium's biosynthetic gene clusters and potential antimicrobial properties against E. coli.
Findings
Faecalibacterium produces butyrate via the butyryl-CoA:acetate CoA transferase pathway.
Faecalibacterium strains often lack antibiotic resistance genes, indicating a favorable safety profile.
Biosynthetic gene clusters in Faecalibacterium may produce antimicrobial compounds like ranthipeptides.
Abstract
Faecalibacterium is among the most abundant bacterial genera in the healthy human colon, comprising approximately 10–15% of the total gut microbiota. Species within this genus ferment complex carbohydrates, including pectin, to produce butyrate, a short-chain fatty acid with anti-inflammatory and anti-carcinogenic properties. Butyrate is the primary energy source for colonocytes and in Faecalibacterium is synthesized via the butyryl-CoA:acetate CoA transferase pathway. Reduced levels of Faecalibacterium are often associated with increased abundance of Escherichia coli and may be linked to early-onset colorectal cancer. Here, genomic analysis of Faecalibacterium strains revealed that several lack antibiotic resistance genes, suggesting a favorable safety profile. Additional genome mining revealed multiple biosynthetic gene clusters (BGCs) involved in the synthesis of secondary…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
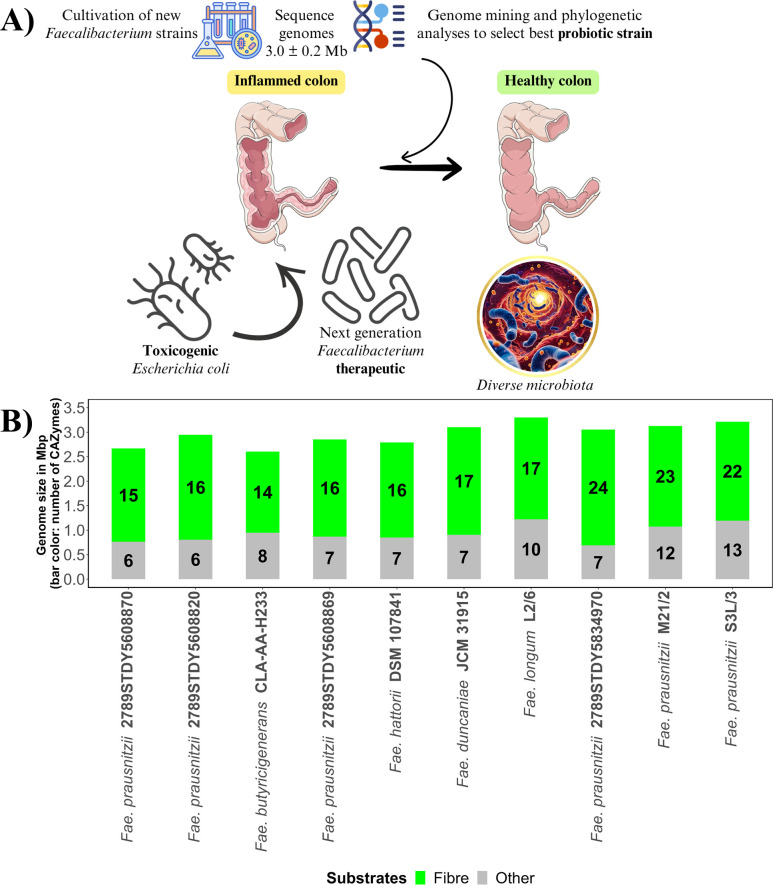 Fig 1
Fig 1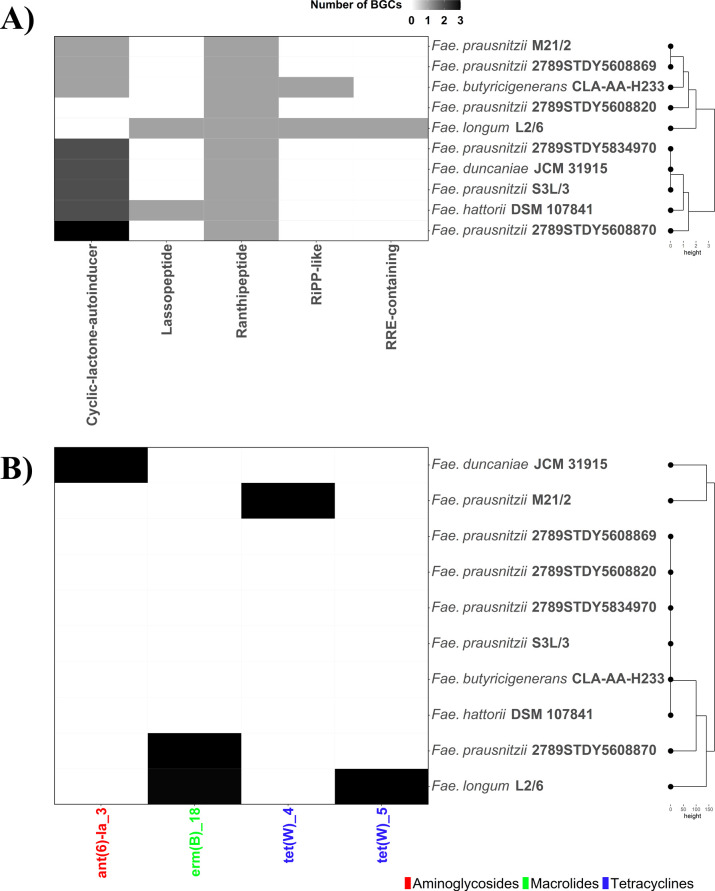 Fig 2
Fig 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Gut microbiota and health · Bacterial Genetics and Biotechnology
PERSPECTIVE
FAECALIBACTERIUM: A KEYSTONE GENUS
The adult human colon hosts a complex and dense ecosystem of bacteria, fungi, and viruses, and the former are comprised of many hundreds of different bacterial species (1). Despite considerable interindividual variation in microbial composition, the microbiota tends to be quite stable in most adults (2) but can show changes along the life course. In particular, the frail elderly tend to have a decline in species diversity (3).
The composition of each person’s microbiota can, however, be driven by a number of factors, including diet (4, 5), colonic pH, oxygen gradients in the colon, and bile salt levels (6–10). These microbes can play a key role in health and disease, including gut health and also a range of other health factors, such as gut-brain axis signaling (11, 12).
In most healthy adults, the single most abundant genus is Faecalibacterium, which has been detected in over 85% of human intestinal samples (13), routinely making up around 5–10% of the total microbiota and can be higher in certain individuals (1, 14). In disease states, its abundance can often be much lower (15). The Faecalibacterium genus, first described by Duncan et al. (16), belongs to the Bacillota (previously Firmicutes) phylum (15) with a genome size of approximately 3.0 ± 0.2 Mb (Fig. 1A). Species within the Faecalibacterium genus include the human-derived species F. prausnitzii, F. butyricigenerans, F. duncaniae (17), F. longum (18), and F. hattorii, while F. gallinarum has been reported in chickens (15).
(A) Use of Faecalibacterium strains as next-generation probiotics and their role as anti-inflammatory gut microbes. This figure has been designed using resources from flaticon.com (free icons “Bacteria” by cube29, “Wand free” by Sir.Vector, “Experiment” by Paul J., and “Dna Strand” by Freepik) and bioicons.com (“healthy-colon-3d” and “crohns-disease” icon by Servier, licensed under CC-BY 3.0). (B) Total number of carbohydrate-active enzymes (CAZymes) acting on fiber (defined as the sum of pectin, xylan/arabinoxylan, mannan, glucan/xyloglucan, fructan, lignocellulosic materials, resistant starch, and malto-oligosaccharides) and other carbohydrate substrates (“Other”) determined in the genome sequences of currently recognized human gut Faecalibacterium strains. CAZymes were annotated using “run_dbcan” software (19), which maps the samples against the CAZy database (http://www.cazy.org/ last accessed: 16 June 2025). Genome sequences were retrieved from the National Center for Biotechnology Information (NCBI) repository.
Faecalibacterium strains are non-spore forming and non-motile rod-shaped cells (16, 20). Moreover, it is a strict anaerobe (16), surviving for less than two minutes following exposure to air (21). Paradoxically, it can, however, be found in sites near the colon wall, where there is diffusion of oxygen and survival could be due to a mechanism that involves extracellular electron shuttling (22). Faecalibacterium strains that have been tested have revealed a degree of sensitivity to bile salts (9). Moreover, certain Faecalbacterium strains showed auxotrophy for vitamins and amino acids (23), suggesting reliance on other microbes for key vitamin and growth factor requirements. Given these findings, it is remarkable that Faecalibacterium can survive and maintain a dominant presence in the human colon.
Dietary substrates fermented by Faecalibacterium strains, determined using in vitro analyses and genome mining, include pectin (24), xylan, and mannan (25) derivatives (see Fig. 1B). Faecalibacterium strains showing a wide range of carbohydrate-active enzymes (CAZymes) acting on fiber include F. prausnitzii strains 2789STDY5834970, M21/2, and S3L-3 (n = 22–24) (Fig. 1B). Fermentation of simple sugars, such as glucose, and other dietary residues that escape digestion by host enzymes results in the formation of butyrate as a major fermentation end product, with minor amounts of other acids, including formate and lactate (16).
ROLE OF FAECALIBACTERIUM IN HUMAN HEALTH
Faecalibacterium is likely to be a major contributor to human health, in part due to its fermentative abilities, which include butyrate formation. As most colonic butyrate producers, Faecalibacterium employs the butyryl CoA:acetate CoA transferase route for butyrate formation. Butyrate is the main energy source for colonocytes and has anti-inflammatory and anti-carcinogenic properties. Moreover, butyrate has a role in host protection via epigenetic changes during the differentiation of monocytes to macrophages. Butyrate induces the latter to upregulate antimicrobial proteins in the gut, such as calprotectin, which plays a crucial role in inflammatory responses and is used as a biomarker to detect intestinal inflammation (26). Faecalibacterium species and other dominant butyrate producers, such as Eubacterium rectale and Roseburia species (27), also act as anti-inflammatory gut microbes (28). Further, there is a correlation between high populations of F. prausnitzii, low IL-12 abundance, and higher IL-10 production (24, 29). A protein (referred to as the MAM protein), which is produced by F. prausnitzii, has been linked to its anti-inflammatory effects (30).
With regard to disease conditions, lower-than-usual levels of F. prausnitzii have been associated with inflammatory bowel disease (IBD) and Crohn’s disease (29) (Fig. 1A). Intestinal disorders, such as Crohn’s disease, found reduced levels of F. prausnitzii in both fecal and mucosal samples (31). The lower abundance of these bacteria is not only associated with the risk of developing IBD, but also with the chance of relapsing after successful treatment. Individuals with lower abundance of F. prausnitzii were six times more likely to relapse in the future (29, 32). Moreover, lower levels of Faecalibacterium generally correlate with elevated levels of Escherichia coli (7). In particular, exposure to colibactin-producing E. coli, particularly in early life, may be a contributing factor in the increasing early onset of colorectal cancer (32). A reduction in Faecalibacterium abundance and its ability to generate secondary metabolites is likely to benefit the survival of pathogenic species.
Several biosynthesis gene clusters (BGCs) involved in the synthesis of secondary metabolites have been identified in the genome sequences of different Faecalibacterium strains (Fig. 2A). Specifically, Faecalibacterium strains have ranthipeptide BGCs, while lassopeptide BGCs were characteristic of the F. prausnitzii strain L2/6. Ranthipeptides are a subclass of ribosomally synthesized and post-translationally modified peptides that share certain similarities to lanthipeptides. The latter have been shown to disrupt bacterial membranes and inhibit bacterial cell wall synthesis in Gram-negative bacteria including E. coli (33). Ranthipeptides have also been reported to play an important role in quorum sensing mechanisms (34), while lassopeptides show biological effects against E. coli, Salmonella, Klebsiella, and Shigella species (35, 36).
(A) Identification of secondary metabolite biosynthesis gene clusters (BGCs) in the genome sequences of currently recognized human gut Faecalibacterium strains using antiSMASH software (37). RiPP-like: other unspecified ribosomally synthesized and post-translationally modified peptide product (RiPP). RRE-containing: RiPP precursor recognition element (RRE)-element containing cluster. (B) Identification of antimicrobial resistance genes (ARGs) in the genome sequences of currently recognized human gut Faecalibacterium strains using abricate software and ResFinder database (38). Genome sequences were retrieved from the National Center for Biotechnology Information (NCBI) repository.
F. prausnitzii therefore shows excellent promise as a bacterium for supporting good health and improving gut barrier function (39, 40) (Fig. 1A). In addition, this bacterium improves the gut barrier by improving the permeability and the expression of tightly bound proteins, including occludin, and both of these increase the tight junctions between cells, thereby strengthening gut barrier function and alleviating inflammation (41).
Supporting populations of Faecalibacterium in the colon may require a number of different approaches, such as interactions with other bacterial species to create a suitable environment, including reducing the redox potential and altering the composition of nutrients (42). Studies show that F. prausnitzii interacts with other bacteria, which influences its butyrate production and survival. As an example, co-culturing F. prausnitzii and bifidobacteria enhanced butyrate production by F. prausnitzii (43) due to the supply of acetate and other nutrients. The supply of acetate helps fuel the butyryl CoA:acetate CoA pathway for butyrate formation, and certain bacteria may also supply other key growth factors, such as vitamins. F. prausnitzii is auxotrophic for most vitamins, including biotin, folate, and thiamine, as well as the amino acid tryptophan (23), reveals its likely reliance on other vitamin- and nutrient-synthesizing gut microbes. Cross-feeding is therefore likely to be essential for Faecalibacterium strains to maintain their dominance in the human colon.
Where Faecalibacterium is in low abundance, it is becoming increasingly important to consider how best to ameliorate this and how to deliver this bacterium to the colon. One option may be fecal microbiota transplantation (FMT), which is the transfer of fecal microbiota from a healthy donor into another individual to promote health. This method has been used with some success to treat a few health conditions, including bowel disorders and infections, particularly those caused by Clostridioides difficile (44, 45); however, it can have side effects, such as bacteremia (46). Given the reported relative success of FMT and the likely prevalence of Faecalibacterium in stool samples from healthy donors, there is considerable credible support for the safety and efficacy of developing Faecalibacterium as a single strain or defined microbial consortia for human use. Using single strains or defined bacterial consortia allows for a much more detailed analysis of the strains of interest, including their antibiotic resistance profiles, providing much more confidence in the use of next-generation probiotic products (47).
The use of well-characterized strains is therefore a much safer, more acceptable, and targeted approach compared to FMT to treat gut health conditions. The sensitivity of Faecalibacterium to oxygen and how best to deliver this anaerobe, either alone or in combination with other strains, to the intestine, however, requires further exploration. When considering the use of Faecalibacterium strains as a next-generation probiotic or biotherapeutic (Fig. 1A), each strain needs to be thoroughly assessed for safety and other factors, such as carriage of antibiotic resistance genes (ARGs), given the level of antibiotic resistance in intestinal bacteria. Aminoglycoside (strain JCM 31915), macrolide (strains L2/6 and 2789STDY5608870), and tetracycline (strains L2/6 and M21/2) resistance genes have been annotated in the genome sequence of Faecalibacterium strains (Fig. 2B). However, no multidrug-resistant Faecalibacterium strains have been identified. Moreover, several strains had no evidence of ARGs (Fig. 2B). It is important to consider that the strains have all factors required for growth and may therefore require adding vitamins and/or another strain (co-culture) such as Bifidobacterium adolescentis which could provide growth factors such as acetate to fuel the butyrate pathway (43). Given that Faecalibacterium does not sporulate and is sensitive to air, if using a microencapsulation approach, these factors also need to be taken into consideration, and there are methods currently used for anaerobes (48). With the increasing number of Faecalibacterium strains that have been isolated and genome-sequenced, it is timely to carry out a deep dive into these strains to determine how many different species exist and to compare important traits across strains, in order to optimize the choice of strain(s) for development as next-generation probiotics to best promote and potentially identify prebiotic approaches for promoting gut health. Moreover, despite growing interest in microbial metabolites, our understanding of the capacity of the human gut microbiota to produce secondary metabolites and inhibit pathogens remains limited. Uncovering this microbial “dark matter” is essential for advancing therapeutic strategies and deepening our understanding of host–microbe interactions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Flint HJ, Scott KP, Duncan SH, Louis P, Forano E. 2012. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 3:289–306. doi:10.4161/gmic.1989722572875 PMC 3463488 · doi ↗ · pubmed ↗
- 2Faith JJ, Guruge JL, Charbonneau M, Subramanian S, Seedorf H, Goodman AL, Clemente JC, Knight R, Heath AC, Leibel RL, Rosenbaum M, Gordon JI. 2013. The long-term stability of the human gut microbiota. Science 341:1237439. doi:10.1126/science.123743923828941 PMC 3791589 · doi ↗ · pubmed ↗
- 3O’Toole PW, Jeffery IB. 2015. Gut microbiota and aging. Science 350:1214–1215. doi:10.1126/science.aac 846926785481 · doi ↗ · pubmed ↗
- 4David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, Ling AV, Devlin AS, Varma Y, Fischbach MA, Biddinger SB, Dutton RJ, Turnbaugh PJ. 2014. Diet rapidly and reproducibly alters the human gut microbiome. Nature 505:559–563. doi:10.1038/nature 1282024336217 PMC 3957428 · doi ↗ · pubmed ↗
- 5Duncan SH, Conti E, Ricci L, Walker AW. 2023. Links between diet, intestinal anaerobes, microbial metabolites and health. Biomedicines 11:1338. doi:10.3390/biomedicines 1105133837239009 PMC 10216541 · doi ↗ · pubmed ↗
- 6Duncan SH, Louis P, Thomson JM, Flint HJ. 2009. The role of p H in determining the species composition of the human colonic microbiota. Environ Microbiol 11:2112–2122. doi:10.1111/j.1462-2920.2009.01931.x 19397676 · doi ↗ · pubmed ↗
- 7Lopez-Siles M, Martinez-Medina M, Surís-Valls R, Aldeguer X, Sabat-Mir M, Duncan SH, Flint HJ, Garcia-Gil LJ. 2016. Changes in the abundance of Faecalibacterium prausnitzii Phylogroups I and II in the intestinal mucosa of inflammatory bowel disease and patients with colorectal cancer. Inflamm Bowel Dis 22:28–41. doi:10.1097/MIB.000000000000059026595550 · doi ↗ · pubmed ↗
- 8Lopez-Siles M, Martinez-Medina M, Abellà C, Busquets D, Sabat-Mir M, Duncan SH, Aldeguer X, Flint HJ, Garcia-Gil LJ. 2015. Mucosa-associated Faecalibacterium prausnitzii phylotype richness is reduced in patients with inflammatory bowel disease. Appl Environ Microbiol 81:7582–7592. doi:10.1128/AEM.02006-1526296733 PMC 4592880 · doi ↗ · pubmed ↗