The Pseudomonas aeruginosa Cpx system provides a cyclic-di-GMP independent link between cell envelope stress and surface sensing
Megan R. O'Malley, Hailey N. Dearing, Xuhui Zheng, Alyssa N. Kretschmer, Timothy H.-S. Cho, Tracy L. Raivio, Matthew R. Parsek

TL;DR
This paper shows that the Cpx system in Pseudomonas aeruginosa helps the bacteria sense surfaces and form biofilms without relying on a key signaling molecule, expanding our understanding of how these bacteria cause infections.
Contribution
The study identifies a novel Cpx signaling pathway in Pseudomonas aeruginosa that enables surface sensing independently of cyclic-di-GMP.
Findings
The P. aeruginosa Cpx system includes two unique adaptor proteins, cpxM and cpxH.
Cpx is activated by surface attachment through a mechanism independent of cyclic-di-GMP.
Cpx influences gene expression related to antibiotic resistance, biofilm matrix production, and redox homeostasis.
Abstract
Bacteria surveil their cell envelope through a network of envelope stress response systems (ESRs). Beyond regulation of envelope maintenance, ESRs influence expression of a range of virulence traits among pathogenic bacteria. The Cpx two-component system, a conserved ESR, responds to envelope stress generated by bacterial contact with a solid surface. This feature is particularly interesting in the context of bacterial biofilm formation, a key virulence trait of the opportunistic pathogen Pseudomonas aeruginosa, which initiates biofilm formation upon detecting envelope stress-related cues of surface adhesion. While a putative Cpx system is present in P. aeruginosa, it exhibits dissimilarities from orthologous systems and has not been evaluated for its roles in stress signaling and/or biofilm formation. Here, we found that the P. aeruginosa Cpx system includes two previously…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
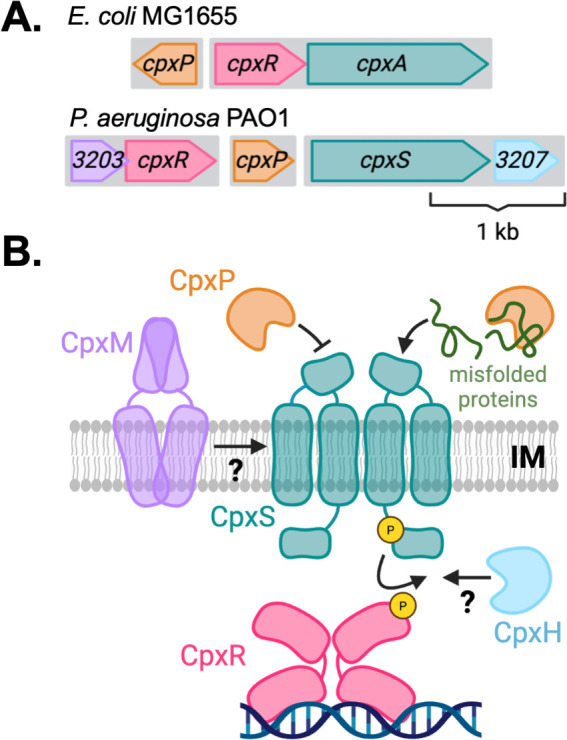 Fig 1
Fig 1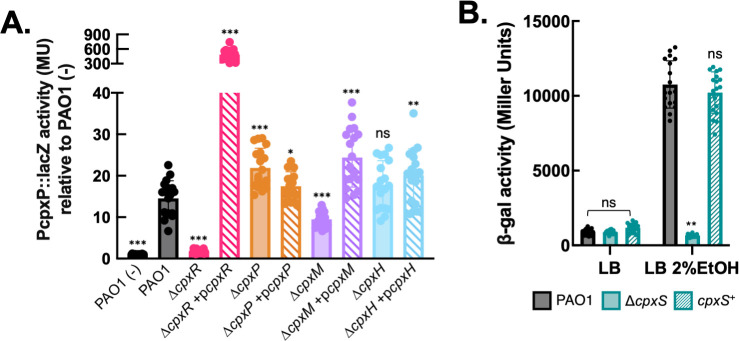 Fig 2
Fig 2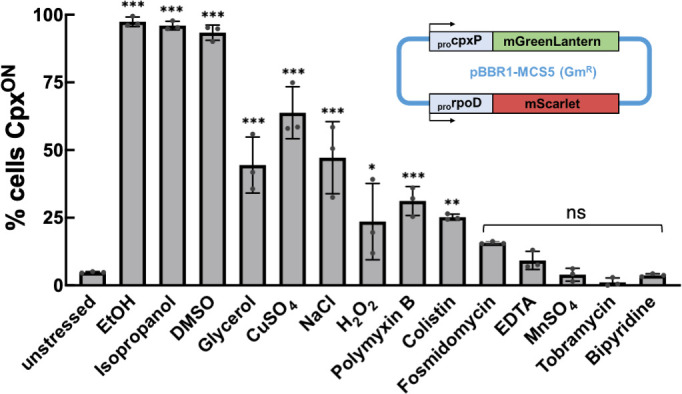 Fig 3
Fig 3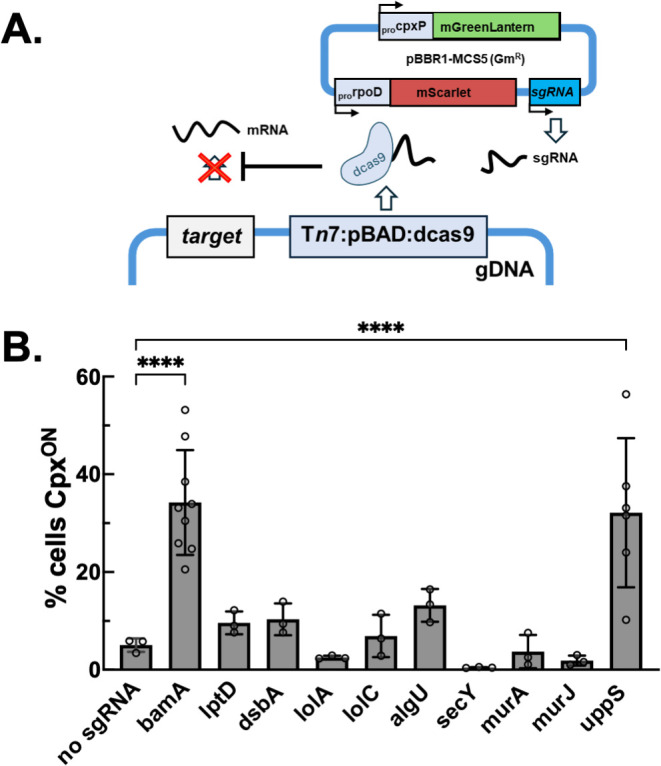 Fig 4
Fig 4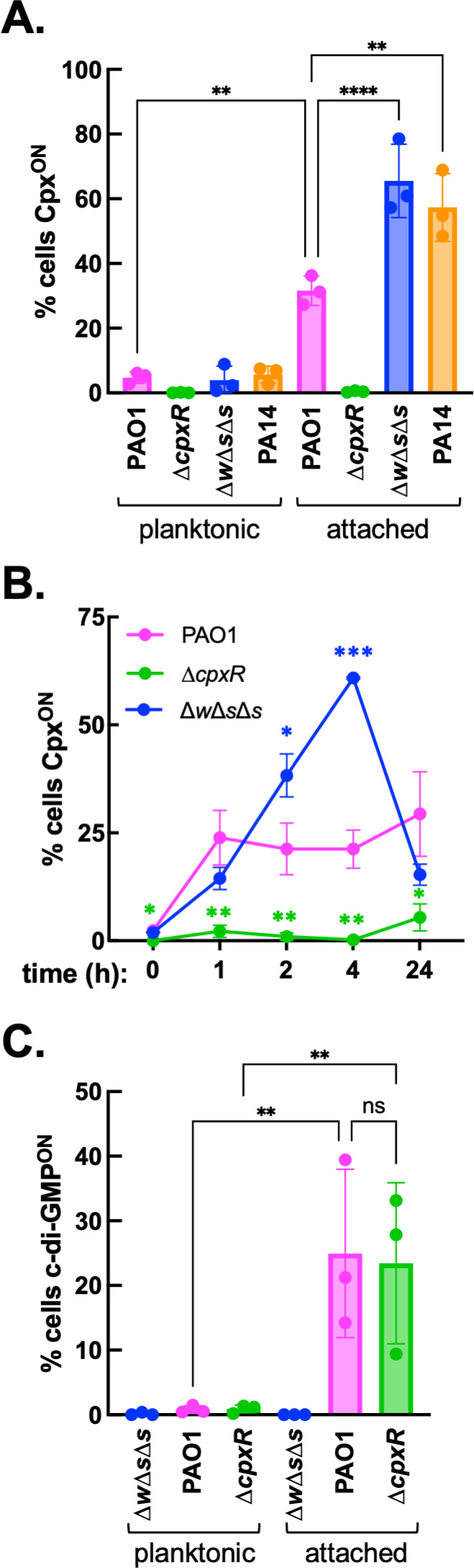 Fig 5
Fig 5| Locus | Gene | Gene function | Dist. from ATG | Predicted CpxR site | ID motif | ID DAP-Seq |
|---|---|---|---|---|---|---|
| PA0425 | mexA | RND efflux protein | −246 |
| y | y |
| PA0502 | nirQ | Transcriptional regulator, denitrification | −87 |
| y | n |
| PA0527 | dnr | Transcriptional regulator, denitrification | −129 |
| n | y |
| PA0546 | metK | Methionine adenosyltransferase | −86 |
| y | n |
| PA0865 | hpd | 4-Hydroxyphenylpyruvate dioxygenase | −194 |
| n | y |
| PA0873 | phhR | Transcriptional regulator, phenylalanine catabolism | −124 |
| n | y |
| PA0888 | aotJ | Periplasmic arginine transport protein | −258 |
| n | y |
| PA0931 | pirA | Ferric enterobactin outer membrane receptor | −195 |
| n | y |
| PA1003 | mvfR | Transcriptional regulator, PQS quorum sensing | −366 |
| n | y |
| PA1156 | nrdA | Ribonucleotide reductase | −196 |
| y | n |
| PA1097 | fleQ | Transcriptional regulator, flagellar motility/adhesion | −212 |
| n | y |
| PA1345 | Hypothetical ATP-binding protein | −69 |
| y | n | |
| PA1528 | zipA | Inner membrane cell division protein | −147 |
| n | y |
| PA2017 | Hypothetical inner membrane beta-propeller protein | −164 |
| n | y | |
| PA2231 | pslA | Exopolysaccharide psl biosynthesis protein | −181 |
| n | y |
| PA2523 | czcR | Two-component response regulator, metal sensing | −22 |
| n | y |
| PA2528 | muxA | RND efflux protein | −78,−89 | y | y | |
| PA2558 | Hypothetical inner membrane MgtC-like transport protein | −105 |
| n | y | |
| PA2637 | nuoA | NADH dehydrogenase I subunit | −363 |
| y | n |
| PA2936 | Cytochrome b561 | −325 |
| n | y | |
| PA3064 | pelA | Exopolysaccharide pel biosynthesis protein | −147 |
| n | y |
| PA3155 | wbpE | Lipopolysaccharide biosynthesis protein | −317 |
| y | n |
| PA3205 | cpxP | Periplasmic cpx repressor protein | −81 |
| y | y |
| PA3268 | Siderophore outer membrane receptor | −53 |
| y | y | |
| PA3575 | yceJ | Cytochrome b561 | −63,−74 | y | y | |
| PA3762 | Hypothetical metal-binding protein | −97 |
| y | y | |
| PA3795 | Cytoplasmic oxidoreductase | −144 |
| y | y | |
| PA3965 | Transcriptional regulator | −315,−326 | y | y | ||
| PA3978 | Hypothetical SEL1 repeat family protein | −93 |
| y | y | |
| PA4075 | Hypothetical methyltransferase | −61 |
| n | y | |
| PA4168 | fpvB | Ferric pyoverdine outer membrane receptor | −111 |
| y | y |
| PA4315 | mvaT | Transcriptional regulator, exotoxin production | −66 |
| n | y |
| PA4431 | Hypothetical inner membrane iron-sulfur protein | −274 |
| y | n | |
| PA4513 | piuC | Inner membrane siderophore oxidoreductase | −131 |
| y | y |
| PA4514 | piuA | Siderophore outer membrane receptor | −91 |
| y | y |
| PA4628 | lysP | Inner membrane lysine permease | −148 |
| y | n |
| PA4798 | Hypothetical protein | −368 |
| y | n |
| Locus | Gene | Description | log2FC |
|
|---|---|---|---|---|
| DEGs upregulated in PAO1 | ||||
| Cpx locus | ||||
| PA3205 | cpxP | cpx periplasmic accessory protein | 8.79 | <0.001 |
| PA3206 | cpxS | Sensor histidine kinase | 5.48 | <0.001 |
| PA3207 | cpxH | cpx accessory protein | 3.88 | <0.001 |
| PA3208 | ydjA | NADPH nitroreductase | 2.65 | <0.001 |
| PA3209 | ykgJ | Cysteine cluster protein | 1.56 | <0.001 |
| Efflux | ||||
| PA2525 | opmB | Outer membrane efflux protein | 3.37 | <0.001 |
| PA2526 | muxC | RND efflux protein | 3.36 | <0.001 |
| PA2527 | muxB | RND efflux protein | 3.53 | <0.001 |
| PA2528 | muxA | Efflux membrane fusion protein | 3.97 | <0.001 |
| PA0425 | mexA | Efflux membrane fusion protein | 1.85 | <0.001 |
| PA0426 | mexB | RND efflux protein | 1.64 | <0.001 |
| PA0427 | oprM | Outer membrane efflux protein | 1.52 | <0.001 |
| PA4596 | esrC | Transcriptional regulator of efflux | 1.08 | <0.001 |
| Siderophore-mediated iron uptake | ||||
| PA4513 | piuC | Inner membrane siderophore oxidoreductase | 1.97 | <0.001 |
| PA4217 | phzS | Flavin-containing monooxygenase | 0.56 | 0.0022 |
| PA4218 | ampP/fptX | Pyochelin membrane transporter | 1.84 | <0.001 |
| PA4219 | ampO | Inner membrane protein | 1.85 | <0.001 |
| PA4222 | pchI | Pyochelin ABC transport protein | 1.66 | <0.001 |
| PA4223 | pchH | Pyochelin ABC transport protein | 1.68 | <0.001 |
| PA4224 | pchG | Pyochelin biosynthesis protein | 2.01 | <0.001 |
| PA4225 | pchF | Pyochelin biosynthesis protein | 1.92 | 0.01 |
| PA4229 | pchC | Pyochelin biosynthesis protein | 2.07 | <0.001 |
| PA4231 | pchA | Pyochelin biosynthesis protein | 1.33 | <0.001 |
| PA3465 | yfiS | Enterobactin efflux transporter-like protein | 0.53 | 0.0018 |
| Ion transport and redox homeostasis | ||||
| PA0545 | Ferric reductase | 2.98 | <0.001 | |
| PA3575 | yceJ | Cytochrome b561/ferric reductase | 4.3 | <0.001 |
| PA3576 | Periplasmic lipocalin-like protein | 2.36 | <0.001 | |
| PA3963 | Probable iron transporter | 2.54 | <0.001 | |
| PA3963a | Readthrough variant of PA3963 with part of PA3964 disordered protein | 5.41 | <0.001 | |
| PA3795 | Cytoplasmic oxidoreductase | 2.66 | <0.001 | |
| PA2524 | czcS | Metal ion-sensing sensor histidine kinase | 1.27 | <0.001 |
| PA0809 | mtnH2 | Divalent metal cation transporter | 0.66 | <0.001 |
| BqsSR Fe(II)-sensing two-component system regulon | ||||
| PA2656 | bqsS/carS | Iron-sensing sensor histidine kinase | 3.13 | <0.001 |
| PA2657 | bqsR/carR | Iron-sensing response regulator | 3.9 | <0.001 |
| PA2658 | bqsQ | BqsSR accessory protein | 4.95 | <0.001 |
| PA2659 | bqsP | BqsSR accessory protein | 4.47 | <0.001 |
| PA0102 | pSCA1 | Beta-carbonic anhydrase | 1.24 | <0.001 |
| PA0320 | carO | Periplasmic substrate binding protein | 7.4 | <0.001 |
| PA0321 | Acetylpolyamine amidohydrolase | 3.93 | <0.001 | |
| PA0322 | Polyamine/amino acid permease | 1.17 | <0.001 | |
| PA0327 | carP | Inner membrane beta-propeller protein | 4.61 | <0.001 |
| PA0328 | aaaA | Arginine autotransporter | 1.64 | <0.001 |
| Adhesins | ||||
| PA3059 | pelF | Pel glycosyltransferase | 0.63 | 0.0033 |
| PA3061 | pelD | Pel biosynthesis protein | 1.07 | <0.001 |
| PA3063 | pelB | Pel biosynthesis protein | 0.82 | <0.001 |
| PA4624 | cdrB | Outer membrane adhesin export protein | 0.60 | <0.001 |
| SOS Response | ||||
| PA0612 | ptrB | Pyocin regulator | 1.07 | <0.001 |
| PA0613 | Pyocin protein | 0.98 | <0.001 | |
| PA0614 | Pyocin protein | 1.18 | <0.001 | |
| PA0615 | Pyocin protein | 0.69 | <0.001 | |
| PA0616 | Pyocin protein | 1.18 | <0.001 | |
| PA0617 | Pyocin protein | 1.28 | <0.001 | |
| PA0618 | Pyocin protein | 1.17 | <0.001 | |
| PA0619 | Pyocin protein | 1.08 | <0.001 | |
| PA0620 | Pyocin protein | 1.01 | <0.001 | |
| PA0621 | Pyocin protein | 1.0 | <0.001 | |
| PA0622 | Pyocin protein | 1.13 | <0.001 | |
| PA0623 | Pyocin protein | 1.19 | <0.001 | |
| PA0624 | Pyocin protein | 1.13 | <0.001 | |
| PA0625 | Pyocin protein | 1.03 | 0.0012 | |
| PA0626 | Pyocin protein | 0.97 | 0.0018 | |
| PA0628 | Pyocin protein | 1.13 | 0.0036 | |
| PA0630 | Pyocin protein | 1.07 | <0.001 | |
| PA0631 | Pyocin protein | 1.06 | 0.0064 | |
| PA0633 | Pyocin protein | 1.27 | <0.001 | |
| PA0635 | Pyocin protein | 1.23 | <0.001 | |
| PA0636 | Pyocin protein | 1.11 | <0.001 | |
| PA0637 | Pyocin protein | 1.13 | <0.001 | |
| PA0638 | Pyocin protein | 1.11 | <0.001 | |
| PA0639 | Pyocin protein | 1.16 | <0.001 | |
| PA0640 | Pyocin protein | 0.98 | 0.0029 | |
| PA0641 | Pyocin protein | 1.01 | <0.001 | |
| PA0643 | Pyocin protein | 0.66 | 0.0005 | |
| PA0669 | dnaE2 | Damage-inducible polymerase | 0.46 | 0.0015 |
| PA0670 | DNA repair protein | 0.67 | 0.0022 | |
| PA0787 | RecF-like DNA repair protein | 0.40 | <0.001 | |
| PA0807 | ampDh3 | Cell wall hydrolase | 0.98 | <0.001 |
| PA0907 | alpA | Autolysis protein | 0.69 | <0.001 |
| PA0910 | alpD | Autolysis protein | 0.88 | <0.001 |
| PA0911 | alpE | Autolysis protein | 0.87 | <0.001 |
| DEGs downregulated in PAO1 | ||||
| Cellular respiration and denitrification | ||||
| PA0509 | nirN | Heme d (1) biosynthesis protein | −1.72 | <0.001 |
| PA0510 | nirE | Heme d (1) biosynthesis protein | −1.56 | 0.0013 |
| PA0511 | nirJ | Heme d (1) biosynthesis protein | −0.92 | 0.0058 |
| PA0520 | nirQ | Denitrification regulatory ATPase protein | −1.24 | <0.001 |
| PA0521 | nirO | Cytochrome c oxidase subunit III | −1.24 | 0.0015 |
| PA0522 | nirP | Cytochrome c oxidase subunit IV | −1.68 | <0.001 |
| PA0523 | norC | Nitric oxide reductase subunit C | −1.16 | 0.0026 |
| PA0524 | norB | Nitric oxide reductase subunit B | −1.12 | 0.0075 |
| PA4129 | NirA-associated oxidoreductase | −1.38 | <0.001 | |
| PA4130 | nirA | Nitrite reductase | −1.29 | <0.001 |
| PA4131 | ccoG | Cytochrome | −1.07 | 0.0058 |
| PA4132 | mpaR | Transcriptional regulator | −0.66 | <0.001 |
| PA4133 | ccoN4 | Cytochrome | −0.86 | <0.001 |
| Fe-S cluster biogenesis | ||||
| PA1847 | nfuA | Fe-S biogenesis protein | −0.85 | <0.001 |
| PA3811 | hscB | Fe-S cofactor chaperone protein | −0.62 | <0.001 |
| PA3813 | iscU | Fe-S assembly scaffold protein | −0.56 | 0.0021 |
| PA3814 | iscS | Fe-S biogenesis protein | −0.81 | <0.001 |
| PA3815 | iscR | Transcriptional regulator | −0.66 | 0.0026 |
| PA5275 | cyaY | Fe-S biogenesis protein | −0.68 | <0.001 |
| Carbohydrate uptake | ||||
| PA3186 | oprB | Outer membrane glucose porin | −1.22 | <0.001 |
| PA3187 | gltK | Glucose transport protein | −1.44 | <0.001 |
| PA3188 | gltG | Glucose transport protein | −1.21 | <0.001 |
| PA3189 | gltF | Glucose transport protein | −1.34 | <0.001 |
| PA3190 | gltB | Glucose permease | −1.74 | <0.001 |
| PA2291 | oprB2 | Outer membrane glucose porin | −0.34 | 0.0039 |
| Motility and chemotaxis | ||||
| PA0395 | pilT | Twitching motility protein | −0.32 | <0.001 |
| PA0413 | chpA | Pil-Chp chemosensory protein | −0.51 | <0.001 |
| PA0414 | chpB | Pil-Chp methylesterase | −0.50 | <0.001 |
- —Cystic Fibrosis Foundationhttp://dx.doi.org/10.13039/100000897
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —Cystic Fibrosis Foundationhttp://dx.doi.org/10.13039/100000897
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Legionella and Acanthamoeba research · Cancer Research and Treatments
INTRODUCTION
Chronic Pseudomonas aeruginosa infection is often associated with the prolific ability of this pathogen to form biofilms on host surfaces, in which sessile, semi-dormant bacteria are embedded in a self-produced extracellular matrix (1, 2). Sensing initial surface adhesion is generally thought to act as an inducing stimulus for bacteria to adopt the biofilm lifestyle (3–6). P. aeruginosa senses perturbations to the cell envelope generated by surface contact, which stimulates a rapid increase in the intracellular concentration of the signaling molecule cyclic-di-GMP (c-di-GMP) (7, 8). C-di-GMP is a global biofilm regulator that suppresses cellular motility and stimulates matrix production. The molecular stress-related signal(s) arising from surface adhesion are not fully understood, though stimuli including contact-induced membrane deformation, disruption of cell envelope protein folding, and/or physiochemical properties of the surface environment which affect envelope function (i.e., local pH or osmolarity) have been discussed as potential mechanisms (9, 10). A broader exploration of P. aeruginosa surface-sensing mechanisms is currently limited by the fact that envelope stress signaling in this pathogen has not been extensively characterized, with most molecular knowledge derived from enteric model organisms such as Escherichia coli.
The Gram-negative cell envelope, consisting of an inner and outer membrane with an intervening peptidoglycan cell wall, is intrinsic to bacterial cell integrity and survival. Envelope homeostasis is impacted by various stressors inherent to the host environment, such as changes in temperature and redox potential encountered during infection, and is also affected by several classes of antibiotics (11). In the model organism E. coli, a network of envelope stress response systems (ESRs) monitors envelope integrity (e.g., the σ^E^, Cpx, Bae, Rcs, and Psp systems) (12). These ESRs play somewhat specialized roles in sensing distinct stimuli arising from envelope stress; for example, misfolded outer membrane proteins are detected by σ^E^, while cell-surface lipopolysaccharide (LPS) defects trigger the Rcs system (13, 14). In addition to remediating envelope damage repair, ESRs have been found to elicit broader effects on the cell, including regulation of virulence gene expression (15).
The conserved Cpx ESR is particularly notable for its effects on virulence. Discovered in E. coli, CpxRA is a classic two-component system comprised of an inner membrane sensor histidine kinase CpxA, which modulates the activity of an OmpR-type response regulator CpxR through phosphorylation (16). The CpxA sensor integrates multiple stress-related signals in E. coli, including interactions with envelope proteins that are misfolded or aberrantly localized at the inner membrane due to defects in protein homeostasis and/or trafficking (17–22). Accordingly, the first identified regulatory targets of CpxR in E. coli were “quality control” factors (i.e., spy, dsbA, ppiA, degP/htrA) involved in remediating or recycling envelope proteins (23–26). Cpx-mediated tolerance of envelope stress is essential for various pathogenic and mutualistic bacterial species to survive within host systems (27–30). The Cpx response has since been found to modulate the expression of a wide variety of genes in many Proteobacteria, including virulence factors such as flagella, pili, chemotaxis systems, biofilm matrix components, exotoxins, and secretion systems (31–35). Both direct transcriptional regulation by CpxR and indirect effects on the stability of large envelope-localized protein complexes due to Cpx-regulated homeostasis factors influence virulence factor biogenesis (36–38). As such, Cpx-associated virulence phenotypes are often complex and vary widely across systems.
E. coli Cpx is also induced by cellular surface adhesion, making this system a compelling target for study in the context of P. aeruginosa surface sensing and biofilm formation. While a putative homologous Cpx ESR has been identified in P. aeruginosa (39), its roles in stress signaling and virulence have not been fully determined. However, spontaneous adaptive mutations in the Cpx sensor kinase are frequently recovered from clinical and antibiotic resistance-evolved populations of P. aeruginosa, suggesting pathogenic relevance of this system (40–44). Several of these mutants, predicted to confer constitutive activation of Cpx signaling, exhibit enhanced antibiotic resistance due to elevated expression of cellular efflux pumps, which are directly regulated by Cpx in both P. aeruginosa and E. coli (39, 45–47). Transcriptomic studies have also reported elevated Cpx-associated gene expression in P. aeruginosa upon surface attachment, including attachment to a variety of abiotic surfaces (48) and pulmonary epithelial tissue (49).
In the present study, we characterize the P. aeruginosa Cpx system. P. aeruginosa encodes two previously uncharacterized, Pseudomonas-specific proteins within the conserved cpx locus, including a putative inner-membrane adaptor protein CpxM (PA3203) required for full Cpx system activity. We demonstrate that Cpx signaling is activated under cell envelope stress, likely related to an outer membrane protein-related signal. Through analysis of genomic CpxR promoter-binding sites and transcriptomic profiling, we find that the PAO1 Cpx response impacts cellular efflux, biofilm matrix production, and iron acquisition and participates in complex regulation of energy metabolism. The P. aeruginosa Cpx regulon notably lacks many of the canonical Cpx targets involved in envelope protein homeostasis previously identified in E. coli, suggesting diversification of Cpx stress signaling in this organism. Our findings further demonstrate that the Cpx surface response constitutes a c-di-GMP-independent surface-sensing mechanism.
RESULTS
Genetic analysis of the Cpx two-component system reveals multiple putative adaptors that impact signaling
The Cpx system consists canonically of a sensor histidine kinase, annotated either CpxA or CpxS by species convention, a response regulator CpxR, and a conserved periplasmic adaptor protein, CpxP. Homologous Cpx systems are present throughout the α-, β-, and γ-Proteobacteria classes. Among representative genomes, the majority of cpx loci exhibit an organization similar to E. coli K-12, in which the cpxP gene is encoded in diverging orientation from an inferred cpxAR bicistronic operon (Fig. 1A) (50). By contrast, in the Pseudomonadaceae family, cpxS and cpxR are encoded separately on opposite sides of cpxP (Fig. S1). Cpx homologs in the model isolate P. aeruginosa PAO1 are encoded by PA3204 (CpxR, 64.5% protein similarity to E. coli K12 MG1655), PA3205 (CpxP, 35.6%), and PA3206 (CpxS, 43.4%). Within the Pseudomonas genus exclusively, additional genes are associated with the cpx locus (Fig. 1A; Fig. S1). Hypothetical protein-coding gene PA3203 forms an operon with cpxR and is generally conserved throughout the genus Pseudomonas, with few exceptions (i.e., Pseudomonas putida KT2440). An additional protein-coding gene, PA3207, is present exclusively within the P. aeruginosa species cluster in an operon with cpxS. PA3203-cpxR and cpxS-PA3207 operons were predicted by the Database of Prokaryotic Operons (DOOR, Pseudomonas Genome Database) (51, 52), and validated using Rockhopper operon analysis (53) from PAO1 RNA-Seq data generated in this study (Table S1).
An extended Cpx system in P. aeruginosa. (A) Schematic of the cpx genetic locus from E. coli MG1655 and P. aeruginosa PAO1 to approximate scale. Cpx protein-coding sequences are indicated by colored arrows in the direction of transcription. Shaded boxes indicate transcriptional units as inferred for the E. coli Cpx system (50) and predicted by Rockhopper for PAO1 (53). Hypothetical protein-coding genes cpxM and cpxH were previously annotated as PA3203 and PA3207 in PAO1, respectively. Scale bar indicates 1 kb. (B) Hypothetical model of Cpx complex formation and envelope stress signaling in PAO1. Two-component sensor histidine kinase CpxS (teal) phosphorylates response regulator CpxR (magenta) from the inner membrane (IM), stimulating CpxR binding to regulatory DNA targets. Repressor protein CpxP modulates signaling by interacting with the periplasmic signaling domain of CpxS, and/or through chaperone activity on misfolded proteins in the periplasm. Prospective interactions between CpxM/H and the two-component system are indicated. Graphics were generated using BioRender.
PA3203 is a small inner membrane protein, predicted by AlphaFold2 to harbor two linked transmembrane and periplasmic α-helices which homodimerize (54). PA3207 has a predicted α/β fold structure similar to those found among bacterial histidine kinase catalytic/ATP-binding domains (55), and harbors a potential catalytic histidine residue (His72). Models of PA3207 exhibit a disordered N-terminal domain consistent in size (21aa residues) to a typical signal peptide; however, this sequence does not match any known signal peptide consensus sequences (SignalP 6.0, PSORTb), and full-length PA3207 was predicted to localize to the cytoplasm by the MatureP classifier (56–58). Based on these predictions, we hereafter refer to PA3203 as CpxM (Cpx membrane protein) and PA3207 as CpxH (Cpx histidine kinase-like protein).
P. aeruginosa CpxR has been previously shown to interact with a “Cpx box” promoter motif originally identified in E. coli and regulates the transcription of target genes including cpxP. As cpxP promoter activity has been widely used as a readout of Cpx signaling in other systems (59–62), we evaluated the activity of a chromosomal PcpxP::lacZ transcriptional reporter in PAO1 and cpx gene deletion mutants. While basal activity of this reporter was low in unstressed LB growth medium (Fig. S2), activity was stimulated ~15-fold during growth in the presence of 2% ethanol, a known Cpx inducer in E. coli (61). This response was fully abrogated in both PAO1 ∆cpxR and ∆cpxS mutants (Fig. 2). Complementation of the ∆cpxR mutant by plasmid-based overexpression of cpxR drastically increased reporter activity, likely due to the high dosage of the complementing allele as noted in previous studies (63, 64). By contrast, the PAO1 ∆cpxS stress response could not be restored by plasmid-based overexpression cpxS allele (data not shown). We hypothesize that overexpression of CpxS may lead to an accumulation of the sensor kinase in its inactive conformation, as CpxS/CpxA orthologs exhibit dual kinase/phosphatase activity toward CpxR (65); as sensor kinases depend on homodimerization for catalytic activity, perturbations to the stoichiometry of the system by overexpression may further complicate plasmid-based complementation. As an alternative approach, we used allelic exchange to reintroduce the wild-type cpxS allele to the PAO1 ∆cpxS mutant strain. The resulting strain, PAO1 cpxS^+^, exhibited a fully restored Cpx response (Fig. 2B). We note that PAO1 ∆cpxS displays slightly elevated basal activity in unstressed media when assessed with an alternative reporter system (pBBR1 PcpxP::mGreenLantern/PrpoD::mScarlet-I, described in later sections), which is more sensitive to heterogeneous patterns of gene expression within a population, consistent with the notion that CpxS may exert complex positive and negative regulation of Cpx signaling in P. aeruginosa as in other bacterial systems.
*The PAO1 cpx locus encodes a multi-adapter two-component regulatory system. (A) Genetic analysis of the PAO1 Cpx signaling system. Activity of a PcpxP::lacZ transcriptional reporter integrated at the neutral Tn7 site was measured in PAO1 and cpx gene deletion mutants, expressing either an unmodified pJN105 plasmid (GmR) or pJN105 expressing complementing alleles of cpx genes under arabinose-inducible control. Reporter activity was quantified by standard β-galactosidase assay (Miller units, MU) on cell lysates from bacterial cultures grown to exponential phase (4 h) in the presence of 0.1% arabinose and 100 µg/mL gentamicin. Cellular stress was induced by the addition of 2% ethanol. Graphed are means (± standard deviation, SD) of PcpxP reporter activity relative to a baseline level defined by PAO1 grown in unstressed LB (-). Asterisks denote statistical significance based on pairwise t-tests between mutant and ethanol-stressed PAO1 reporter activity levels, ***P < 0.001, **P < 0.01, *P < 0.05, “ns” no significant difference. Data are pooled from three independent experiments; n = 18. (B) Complementation of PcpxP::lacZ reporter activity in a PAO1 ∆cpxS mutant by restoring the wild-type allele to the chromosome through allelic exchange (cpxS+). Mean (± standard deviation, SD) MU are shown. Asterisks denote statistical significance based on pairwise t-test between mutant and wild-type PAO1 reporter activity levels, **P > 0.001, “ns” no significant difference. Data are pooled from three independent experiments; n = 18. Graphing and statistical analysis were performed in GraphPad Prism v10.1.
Using this reporter system, we similarly assessed the role(s) of CpxP and the hypothetical proteins CpxM and CpxH. In E. coli, CpxP binds misfolded periplasmic proteins and the sensor domain of CpxA, functioning largely as a negative autoregulator of Cpx activity (59, 66, 67). CpxP is the only PAO1 homolog to E. coli CpxP and its paralog, Spy (66, 68); we note that P. aeruginosa CpxP exhibits stronger protein similarity to Spy (47.4%) than to E. coli CpxP. A PAO1 ∆cpxP mutant displays elevated reporter activity, indicating that, as expected, CpxP dampens Cpx activity in P. aeruginosa. Complementation of PAO1 ∆cpxP restored near wild-type levels of activity. PAO1 ∆cpxP did not exhibit constitutive activation during unstressed growth in LB (Fig. S2), suggesting that de-repression in the absence of CpxP is not alone sufficient to activate the Cpx system. Meanwhile, a PAO1 ∆cpxM mutant strain displayed a significantly reduced response under ethanol stress, which was restored by cpxM complementation. A PAO1 ∆cpxH mutant strain exhibited an ethanol response similar to wild-type PAO1, though cpxH plasmid-based complementation significantly elevated reporter activity, suggesting that CpxH is nonessential but may play an accessory role in enhancing Cpx signaling. We note that plasmid-based expression of complementing cpx gene alleles significantly elevated Cpx reporter activity above PAO1 levels, which was likely an effect of gene dosage due to plasmid copy number and/or gene expression levels under inducible promoter control. Our genetic characterization of the PAO1 Cpx system suggests that the core CpxRS two-component system is required for activation of the Cpx stress response, while accessory proteins CpxM and CpxP function, respectively, as positive and negative regulators of Cpx activity.
The PAO1 Cpx system functions as a cell envelope stress sensor
Several envelope stress-related stimuli, including alterations in membrane composition and toxic protein accumulation, have been previously shown to induce Cpx signaling in various bacteria (18, 69–71). To determine the Cpx stimulon in P. aeruginosa, we screened a variety of chemical stressors with known impacts on envelope homeostasis. To effectively compare the impact of various stressors on Cpx signaling, we utilized chemical concentrations that uniformly suppressed growth rate to around 20% of unstressed PAO1 growth (Fig. S3). Cpx activity in PAO1 was evaluated by a pBBR1 plasmid-based bicolor fluorescence reporter containing the transcriptional fusions PcpxP::mGreenLantern (mGL) and PrpoD::mScarlet-I, the latter a constitutively active reporter used to normalize PcpxP reporter signal (6, 8). Fluorescence intensity was quantified from single cells imaged by microscopy. We found PrpoD::mScarlet-I reporter activity to be consistent across stress conditions, including those that impacted cell growth (Fig. S4 and S9).
Through this approach, we identified a variety of stressors that elicit the Cpx response in PAO1 (Fig. 3). As PcpxP::mGL reporter activity showed significant heterogeneity within the population, we scored the impact of stressors based on the percentage of individual cells with high Cpx reporter activity (Cpx^ON^) based on an experimentally determined threshold (see Materials and Methods) (72). Protein denaturants such as ethanol, isopropanol, and DMSO were the strongest inducers, with over 90% of cells designated Cpx^ON^. Other stressors exerted milder but significant impacts on Cpx activity, including oxidative and osmotic stress. Reporter activity was also stimulated by polymyxins, a class of antibiotics that impact outer membrane integrity by disrupting cell-surface LPS stability (73), suggesting that outer membrane dysbiosis is also relevant to Cpx stress signaling. Importantly, we observed that multiple stressors that impacted PAO1 cell growth did not elicit a significant Cpx response, demonstrating that this response was not simply attributable to a general stress response or altered growth.
*Chemicals that disrupt envelope homeostasis impact Cpx signaling. Activity of a bicolor fluorescent reporter plasmid (inset), containing transcriptional fusions of PcpxP::mGreenLantern (mGL) and the housekeeping promoter PrpoD::mScarlet, was assessed in PAO1 after 4 h of growth in the presence of select chemical stressors. Fluorescence intensity was quantified from single bacterial cells imaged by microscopy. Normalized mGL/mScarlet values were compared to those of unstressed cells; cells were considered CpxON when normalized reporter activity exceeded twofold the mean value from the unstressed cell population. Graphed are means (± SD) of the percentage of CpxON cells in the imaged population from data pooled across three independent experiments, with >100 total cells imaged per condition. Chemical stressors were applied at the following concentrations: 2% (vol/vol) ethanol, 1% (vol/vol) isopropanol, 4% (vol/vol) DMSO, 8% (wt/vol) glycerol, 3 mM CuSO4, 400 mM NaCl, 12.5 mM hydrogen peroxide, 0.5 µg/mL polymyxin B, 0.38 µg/mL polymyxin E (colistin), 32 µg/mL fosmidomycin, 0.3 mM EDTA, 5 mM MnSO4, 4 µg/mL tobramycin, and 1.4 mM bipyridine. One-way ANOVA with multiple comparison test was performed in GraphPad Prism v10.3.0; *P < 0.05, **P < 0.01, **P < 0.001, “ns” no significant difference.
While these findings substantiated that the PAO1 Cpx system generally responds to cell envelope stress, the pleiotropic effects of many chemical stressors on the cell envelope limit the interpretation of these results. We therefore employed an additional CRISPRi approach to conditionally deplete expression of genes involved in cell envelope homeostasis. This system is comprised of an integrated spdcas9 enzyme under inducible promoter control; when recruited to target genes by a single guide RNA (sgRNA) probe, spdcas9 inhibits transcript elongation by steric hindrance (Fig. 4A) (74, 75). To assess the impact of CRISPRi gene knockdown on Cpx signaling, sgRNAs were expressed from the same pBBR1 plasmid encoding P_cpxP_::mGL / P_rpoD_::mScarlet-I reporters. Induction of spdcas9 expression in the absence of a functional sgRNA did not impact growth, indicating that this system is not toxic to producer cells (Fig. S5 and S6). We screened a panel of sgRNAs, spanning various essential cell envelope functions including general secretion (secY), periplasmic protein folding (dsbA), envelope stress signaling (algU), LPS biogenesis (lptD, uppS), outer membrane protein folding (bamA), lipoprotein trafficking (lolA, lolC), and peptidoglycan biosynthesis (murA, murJ, uppS). The effectiveness of CRISPRi gene depletion was validated by the severe growth phenotypes incurred when spdcas9 was induced in the presence of sgRNAs targeting essential functions and/or phenotypic effects of sgRNA targeting for non-essential genes (dsbA and algU), (Fig. S7).
*CRISPRi knockdown of key envelope homeostasis regulatory genes impacts Cpx signaling. (A) Schematic of the CRISPRi experimental approach. PAO1 strains with an integrated, Tn7::araBAD::spdcas9 cassette expressed a bicolor fluorescent reporter pBBR1 plasmid, containing transcriptional fusions of PcpxP::mGreenLantern and PrpoD::mScarlet-I. sgRNAs targeting the indicated genes were constitutively expressed under SpeI promoter control from the same plasmid. (B) Cells (>100 per condition) were imaged by microscopy after 4 h incubation with 0.2% arabinose to induce spdcas9 expression. Single-cell fluorescence intensity quantified with MicrobeJ and normalized PcpxP::mGreenLantern/ PrpoD::mScarlet values were compared to those from cells expressing a fluorescent reporter plasmid with no sgRNA. Cells were considered CpxON when normalized reporter activity exceeded twofold the mean value from the control population (no sgRNA). One-way ANOVA with multiple comparisons test was performed in GraphPad Prism v10.3.0; ***P < 0.001. Data are pooled from three biological replicates, except for the bamA and uppS conditions, which represent 9 and 6 biological replicates, respectively.
Most sgRNAs did not induce Cpx signaling (Fig. 4B; Fig. S8). However, we observed a significant increase in Cpx activity when either the bamA or the uppS gene expression was depleted by CRISPRi. BamA is a subunit of the conserved Bam complex, which mediates the insertion of β-barrel proteins into the outer membrane (76). UppS is a cytoplasmic enzyme that catalyzes the formation of undecaprenyl pyrophosphate (often referred to as C55-PP). C55-PP is a precursor to lipid II, an inner membrane “carrier lipid” which mediates the export of hydrophilic envelope components, including peptidoglycan and LPS (77, 78). However, we did not observe Cpx induction by CRISPRi attenuation of other genes involved in cell wall or LPS biogenesis (lptD, murA, murJ), suggesting that defects in the cell wall and/or cell surface LPS are not alone sufficient for Cpx activation. While these findings substantiate a role of Cpx in envelope stress signaling, further study is necessary to determine the exact molecular signal(s) produced by bamA and uppS depletion responsible for Cpx activation.
The Cpx system regulates gene expression related to efflux, iron acquisition, adhesion, and redox stress in PAO1
Around 100 promoters have been identified as targets of CpxR in E. coli based on the presence of a CpxR conserved direct-repeat DNA-binding motif (5′-GTAAA-N_5_-GTAAA-3′), which the P. aeruginosa PA14 CpxR homolog binds to in vitro (26, 39, 79). We queried the PAO1 genome for CpxR motifs present in gene promoter regions and identified 37 transcriptional units in PAO1 potentially under Cpx regulation (Table 1). Prospective CpxR-regulated genes in PAO1 included previously established CpxR targets in E. coli, including cpxP, the efflux system muxABC-opmB, a putative cytochrome b561 yceJ (PA3575), and NADH dehydrogenase nuoAB (61, 80, 81). However, PAO1 orthologs of many canonical E. coli CpxR targets (dsbA, ppiA, algU [rpoE], algW [degP/htrA], PA2604 [yccA], PA2830 [htpX], and PA3712 [yebE]) lacked any evident CpxR promoter-binding sites, suggesting that the Cpx regulon may differ substantially between these organisms.
To investigate the Cpx regulon, we profiled the global transcriptome of a PAO1 strain with a Cpx-activating mutation in the sensor kinase CpxS (PAO1 cpxS^T163P^). A spontaneous missense mutation in CpxS was originally identified in an in vitro evolved isolate of P. aeruginosa PA14 under antibiotic stress (40). Adaptive cpxS mutations have been frequently identified in various stress-evolved and host-derived P. aeruginosa populations, many of which similarly confer transmembrane proline substitutions (43, 44). We introduced this mutation into our PAO1 wild-type strain and found that it drastically elevated PcpxP::lacZ reporter activity in a cpxR-dependent manner (Fig. S10). Using RNA-Seq to identify differentially expressed genes (DEGs) between PAO1 vs. PAO1 cpxS^T163P^, we identified a total of 176 DEGs (Padj <0.01), with 110 upregulated and 66 downregulated in PAO1 cpxS^T163P^ (Table 2; Table S2) (83). qRT-PCR was used to validate the expression level of select DEGs (Fig. S11). Several predicted direct targets of CpxR were upregulated in PAO1 cpxS^T163P^, including cpxP, the efflux systems mexAB-oprM and muxABC-opmB, czcS, yceJ (PA3575), PA3795, piuC (PA4513), and genes from the pel exopolysaccharide operon. These results coincide with previous findings that the Cpx response upregulates the mexAB-oprM and muxABC-opmB efflux systems in both E. coli and PA14. Cpx regulation of pel, a biofilm matrix component in Pa, also echoes other studies in which the Cpx system has been found to regulate a variety of cell-surface adhesins (31, 33, 35, 84, 85).
Many of the genes significantly upregulated in PAO1 cpxS^T163^ are related to iron acquisition. Iron is essential for bacterial growth, but its abundance is often limited during infection due to host competition and limited solubility of oxidized iron (Fe(III)) in aerobic environments. P. aeruginosa scavenges extracellular iron by producing a variety of secreted high-affinity Fe(III)-binding siderophores. Genes involved in the production and assimilation of the siderophore pyochelin were upregulated, as well as a variety of redox enzymes involved in reducing Fe(III) to Fe(II), which has improved bioavailability and can be readily assimilated by the cell. Cpx-activating mutations have also been associated with increased secretion of the siderophore pyoverdine via the MuxABC-OpmB pump, suggesting a potentially multifaceted role of Cpx in iron regulation (44, 86). Of the genes predicted as CpxR direct targets that did not exhibit significant differential expression in PAO1 cpxS^T163P^, we note that several (pirA/PA0931, PA3268, fpvB/PA4168, piuA/PA4514) encode cell-surface siderophore receptors, which often require the presence of their ferrisiderophore cargo for full expression, a feature likely limited under our iron-rich conditions (87). A previous transcriptomic analysis of the Cpx response in Vibrio cholerae El Tor C6706 similarly revealed upregulation of iron acquisition factors, including several orthologous genes to those identified here (Table S3) (64).
PAO1 cpxS^T163P^ also exhibits elevated expression of genes encoding the metal ion-induced two-component systems CzcRS and BqsRS. CzcRS is a predicted target of CpxR and regulates cellular efflux and outer membrane permeability in response to high concentrations of various heavy metal ions (88, 89). The BqsRS (CarRS) system senses reduced iron (Fe(II)) (90, 91); bqsRS and its regulon were among the highest upregulated genes identified in this study and were similarly upregulated by Cpx in Vibrio cholerae (63). Given the abundance of iron acquisition and redox factors upregulated in PAO1 cpxS^T163P^, we hypothesize that elevated Cpx activity increases the cellular concentration of bioavailable Fe(II) iron, leading to induction of BqsRS. Elevated cellular iron in PAO1 cpxS^T163P^ may also account for the broad upregulation of genes associated with the cellular response to oxidative damage, including a large stress-inducible R2/F2-type pyocin gene cluster, as free iron and ferrisiderophores generate damaging reactive oxygen species through Fenton/Haber-Weiss chemistry (92–95).
We observed a significant reduction in the expression of the denitrification regulatory gene nirQ, a predicted CpxR target, indicating that PAO1 CpxR can exert both stimulatory and inhibitory effects on target promoters. We observed that the predicted CpxR-binding site upstream of the nirQ gene overlaps a known binding site for the global denitrification regulator Anr, which is essential for expression of both diverging nirQ and nirS genes, the latter encoding the precursor for nitrite reductase (96). Other denitrification-related genes, particularly those involved in the production of heme-containing and iron-sulfur enzymatic cofactors, were downregulated in cpxS^T163P^. Suppression of these functions may serve as a protective mechanism for P. aeruginosa under envelope stress, where these chemically labile cofactors can damage cell structures through Fenton chemistry (95, 97, 98).
The Cpx system is activated by surface attachment and is not linked to c-di-GMP-based surface sensing
The E. coli Cpx system is generally considered to be induced upon bacterial surface contact, though this phenomenon significantly varies based on experimental conditions such as surface type (22, 85, 99, 100). In the current model, E. coli Cpx senses surface adhesion through a mechanism involving the CpxA periplasmic sensor domain and outer membrane-localized proteins, including the lipoprotein sensor NlpE and the outer membrane β-barrel protein OmpA (22, 100, 101). P. aeruginosa lacks an obvious NlpE homolog, and whether the Cpx system is induced by surface attachment is unclear. To investigate, we assessed the activity of a PcpxP::mGL fluorescent reporter among cells from planktonic and surface-attached populations. A similar approach was previously used in our lab to monitor a temporal increase in c-di-GMP signaling following P. aeruginosa surface contact, in which the activity of a fluorescent c-di-GMP-sensitive promoter (PcdrA) was quantified in surface-attached cells at a liquid:solid interface (8).
We found that PcpxP::mGL fluorescent signal was substantially elevated in surface-attached P. aeruginosa (Fig. 5A); strong single-cell heterogeneity in reporter activity was observed, particularly among attached cells, with many individuals exhibiting at least a fivefold increase in normalized PcpxP::mGL activity relative to a ∆cpxR-negative control population. For strain PAO1, we found that around a quarter of the attached cell population showed high Cpx activity (Cpx^ON^) after 4 h surface attachment. Another widely studied P. aeruginosa isolate, PA14, exhibited a stronger Cpx surface response, with >60% attached cells Cpx^ON^. A time-course analysis of PcpxP::mGL reporter activity in PAO1 further indicated that the Cpx response is elicited rapidly upon initial surface contact; PAO1 reporter activity rose to ~25% cells Cpx^ON^ 1 h post-surface attachment and remained constant at later time points (Fig. 5B). We similarly assessed Cpx surface signaling among various PAO1 ∆cpx gene deletion mutants and found that both the core two-component system CpxRS and adaptor CpxM are required for PAO1 to mount a full Cpx surface response (Fig. S12A). A functional Cpx signaling system was not required for effective PAO1 colonization of surfaces at the liquid:solid interface (Fig. S12B), suggesting that the Cpx system is important post-attachment.
*The Cpx system is induced in surface-attached P. aeruginosa independent of c-di-GMP. (A) Single-cell activity of a PcpxP::mGreenLantern/PrpoD::mScarlet fluorescent reporter plasmid was quantified by microscopic imaging of fixed P. aeruginosa. Bacteria were grown to mid-exponential phase in LB for 3 h (Abs600 ~ 0.5) and either fixed immediately (planktonic cell fraction) or exposed to a LB 1.5% agarose surface for an additional 4 h, after which the attached cell fraction was harvested. Wild-type PAO1 and isogenic mutants PAO1 ∆cpxR and PAO1 ∆wspR∆sadC∆siaD (∆w∆s∆s), as well as model isolate PA14, were analyzed. Cells were designated CpxON when normalized PcpxP reporter activity exceeded 5× an experimentally determined arbitrary threshold, defined by the mean normalized reporter activity from the planktonic PAO1 ∆cpxR population. Graphed are means (± SD) of the %CpxON cells per condition, with asterisks denoting statistical significance based on ordinary one-way ANOVA with multiple comparisons test; **P < 0.01, ****P < 0.0001. (B) Time-course analysis of PcpxP reporter activity in PAO1 and derivative mutant strains following exposure to a liquid:agarose interface, with 0 h representing planktonic cells from the initial inoculum. Asterisks denote statistical significance based on pairwise t-test between PAO1 and mutant strains at the indicated time points; *P < 0.05, **P < 0.01, ***P < 0.001. (C) PAO1 and derivative mutants expressing a PcdrA::mGreenLantern/PrpoD::CFP fluorescent reporter plasmid were assessed for surface-induced activity of the c-di-GMP sensitive promoter PcdrA as described in (A), normalizing PcdrA to PrpoD. Cells were considered PcdrA/c-di-GMPON when normalized PcdrA activity exceeded 5× the mean value from the planktonic PAO1 ∆w∆s∆s population. Graphed are means (± SD) of the %c-di-GMPON cells per condition, asterisks based on one-way ANOVA with multiple comparisons test; *P < 0.01, “ns” no significant difference. All data are pooled from three independent experiments, with >100 cells assessed per biological replicate.
To determine whether the Cpx surface response was affected by c-di-GMP levels, we similarly tested reporter activity of a PAO1 ∆wspR∆sadC∆siaD (∆w∆s∆s) mutant lacking the three major diguanylate cyclases responsible for intracellular c-di-GMP accumulation in response to surface contact (8, 102). This strain displays an overall reduction in initial (1–4 h) surface colonization (Fig. S13), consistent with previous observations that P. aeruginosa with low c-di-GMP levels typically exhibit increased cellular motility and fail to remain stably adhered to a surface (72). PAO1 ∆w∆s∆s exhibited a strong Cpx surface response similar to PA14 (>60% Cpx^ON^), significantly exceeding Cpx reporter levels observed in the PAO1 parental strain (Fig. 5A). Time-course analysis of PcpxP reporter activity further revealed that PAO1 ∆w∆s∆s exhibited a transient increase in Cpx signaling 2–4 h post-attachment, which returned to PAO1 levels by 24 h (Fig. 5B). While the molecular basis of this phenotype is subject to ongoing investigation, these data indicate that Cpx surface sensing does not involve the cyclases linked to the c-di-GMP surface response, but may be promoted during the transient, unstable surface interactions characteristic of c-di-GMP-defective P. aeruginosa. Correspondingly, surface-induced activity of the c-di-GMP reporter PcdrA::mGL did not significantly differ between a PAO1 ∆cpxR mutant and its parental strain (Fig. 5C), similarly suggesting that Cpx signaling does not impact the c-di-GMP surface response. Together, these findings suggest that the Cpx and c-di-GMP surface responses function independently.
DISCUSSION
Relative to the well-characterized landscape of ESRs in Enterobacteriaceae, envelope stress signaling in P. aeruginosa is less understood. In the E. coli paradigm system, the cell envelope is surveilled by a network of ESRs, including σ^E^, Cpx, Bae, Psp, and Rcs (12). Based on sequence homology, P. aeruginosa PAO1 possesses complete orthologs of only the Cpx and σ^E^ (AlgU) systems. The homologous σ^E^ and AlgU extracytoplasmic sigma factors similarly regulate expression of genes involved in assembly of the cell wall, LPS, and membrane proteins, many of unknown function (103–105). Outside of this core set of envelope-related functions, the full σ^E^ regulon appears to vary within the E. coli species group and across bacterial species (106, 107), including regulation of diverse virulence traits such as the biofilm exopolysaccharide alginate, an important factor in chronic P. aeruginosa pulmonary infections (108). In this study, we found that the Cpx system of P. aeruginosa also exhibits significant divergence from its E. coli ortholog, including novel Pseudomonas-specific Cpx adaptor proteins and evident differences in the Cpx regulon. The dissimilar envelope stress responses in E. coli and P. aeruginosa may reflect the significant ecological and pathogenic differences of these bacteria (an obligate animal commensal vs. an environmentally acquired opportunistic pathogen, respectively).
These apparent species-level differences highlight a need for further in situ investigation of Pseudomonas ESR signaling networks. The exceptionally large repertoire of two-component systems in P. aeruginosa (>60 vs. ~30 in E. coli K-12) (109) potentially contains various novel ESRs, with several systems involved in envelope stress tolerance already identified (i.e., PhoPQ, PmrAB, AmgRS, DsbRS) (110–115). Of these, AmgRS and DsbRS have been shown to directly regulate envelope homeostasis factors (i.e., htpX, dsb) that are typically regulated by the Cpx system in E. coli, but absent from the Cpx regulon in P. aeruginosa. Compartmentalization of the envelope stress response across a multiplicity of sensory systems may enable P. aeruginosa to fine-tune its response to specific stress-related stimuli, and functional redundancy among systems may permit evolution of specialized regulators linking cell ESR signaling to other biological processes, including virulence and biofilm regulation. Diversification of two-component signaling with adaptor proteins may further expand the P. aeruginosa ESR network. The incorporation of Pseudomonadaceae-specific accessory genes into broadly conserved two-component systems (i.e., BqsRS, CbrAB) has been described previously (90, 116). In this study, we investigated the function of two novel Cpx adaptor genes, cpxM (PA3203) and cpxH (PA3207), finding that cpxM is required for full Cpx signaling in PAO1, including Cpx induction in response to P. aeruginosa surface contact. The predicted periplasmic domain of the CpxM protein may facilitate interaction(s) with Cpx-inducing ligands, and/or this adaptor may potentiate CpxS activation through interactions within the inner membrane. While the histidine phosphotransferase-like protein CpxH is nonessential for the Cpx response, this factor may enhance Cpx phosphorelay activity or facilitate regulatory crosstalk with other systems. As this protein is present only in P. aeruginosa, it is likely a more recent acquisition of the Cpx system and represents an intermediate state in the evolution of a novel two-component signaling mechanism. Further investigations into the cellular functions of CpxM and CpxH are currently ongoing.
Our work substantiates that the P. aeruginosa Cpx system, as in other species, is induced by cell envelope stress. By targeting specific envelope functions with CRISPRi, we furthermore discovered that OMP dysbiosis (i.e., due to bamA transcriptional arrest by CRISPRi) is a major inducing cue of the PAO1 Cpx system. When the Bam function is disrupted, uninserted β-barrel OMPs likely accumulate in the periplasm, where they may activate Cpx due to misfolding or erroneous insertion in the inner membrane (117, 118). The E. coli Cpx system is similarly induced under conditions where OMP assembly is dysregulated, though the underlying mechanism(s) are not fully understood (119, 120). The P. aeruginosa Cpx system may be regulated by a dedicated outer membrane protein sensor, similar to interactions previously described between E. coli CpxA and the porin OmpA (22, 121). Our observation that the P. aeruginosa CpxP adaptor protein bears strong similarity to E. coli Spy, a periplasmic chaperone known to bind unfolded outer membrane proteins, may further suggest that P. aeruginosa Cpx signaling involves an unknown outer membrane protein factor.
The P. aeruginosa Cpx system was also stimulated by CRISPRi targeting of uppS, which depletes cellular availability of the inner membrane carrier lipid C55-PP. The molecular basis of this response is unclear, particularly considering that direct inhibition of cell envelope biogenesis processes requiring carrier lipids, such as cell wall peptidoglycan and LPS production, did not induce Cpx signaling to the same degree. Pleiotropic effects of uppS deletion, impacting the stability of multiple envelope compartments, may contribute to Cpx activation. We note that the sensitivity of our reporter is potentially limited to detecting large changes in Cpx signaling. We observed significant Cpx reporter induction in response to depletion of bamA, consistent with a recent RNA-Seq analysis using the same bamA sgRNA in PAO1, which reported a ~20-fold upregulation in cpxP transcript levels (75); however, that study also reported ~6-fold upregulation of cpxP under lptD depletion, which did not significantly impact activity of our reporter. Another prior study reported that cpxP is upregulated under inhibition of lipoprotein trafficking in PAO1 (122), in contrast to our findings that lolA and lolC depletion do not elicit Cpx reporter activity, though substantial differences in experimental approach may account for this discrepancy (123, 124). Further investigation is therefore necessary to determine the scope and nature of Cpx-inducing stress stimuli in P. aeruginosa.
Consistent with the E. coli literature, we also observed that surface attachment stimulates the Cpx system in P. aeruginosa through a mechanism independent of the global biofilm regulator c-di-GMP. The magnitude of the Cpx surface response differed between isolates PAO1 and PA14, which are often used as representatives of the two major phylogenetic clades of P. aeruginosa (125) and exhibit distinct patterns of surface attachment and biofilm formation (126). PAO1 cells rapidly form stable associations with a surface, often “laying down” along the cell body, accompanied by an early accumulation of c-di-GMP (i.e., induction of a c-di-GMP-sensitive fluorescent reporter after 4 h surface attachment) (8, 72). Meanwhile, PA14 favors initial surface adhesion at the cell pole and exhibits a relatively prolonged period of “sampling” the surface through transient, reversible attachment (127). This behavioral pattern slows the initial progression of surface colonization, with c-di-GMP not significantly accumulating in PA14 until ~24 h (128). Overall, our findings suggest that the Cpx response is rapidly induced upon initial surface contact and predominates in the early stages of P. aeruginosa surface interaction. The heightened Cpx surface response observed among strains lacking an early c-di-GMP surface response (PA14, PAO1 ∆w∆s∆s) may therefore accumulate through multiple successive “surface-sampling” contact events, as cells engage in transient attachment events before committing to biofilm formation.
The consequences of Cpx surface signaling on biofilm formation remain to be fully explored. Our data suggest that Cpx may regulate the production of the cell-associated exopolysaccharide Pel, providing an additional, c-di-GMP-independent link between surface sensing and biofilm matrix production. Unlike c-di-GMP, however, Cpx does not appear to broadly suppress the expression of genes involved in motility (i.e., flagella, pili) in P. aeruginosa. An alternative mechanism for surface-engaged cells to initiate exopolysaccharide production, while retaining some level of cellular motility, may enable a bacterial population to effectively distribute and arrange itself as it colonizes a new surface to maximize overall biofilm fitness. Other prospective Cpx-regulated processes identified in this study, such as iron acquisition, are also associated with P. aeruginosa biofilm formation. Given that sufficient concentrations of iron are required for surface-associated P. aeruginosa cells to commit to biofilm formation (129, 130), Cpx activation may “prime” transient or peripherally attached cells to increase the uptake of this key nutrient. This initial surface response may therefore lay the foundation for the progression of biofilm formation once other environmental criteria for biofilm initiation, such as adequate levels of environmental iron, are met. Other Cpx-regulated factors, such as those involved in redox homeostasis, may enable cells to acclimate to the unique physiological conditions of the surface, which can differ significantly from aqueous environments (9, 131). Future studies will investigate the effects of the Cpx surface response on individual cell fate during surface colonization, as well as the coordination of Cpx with other surface-sensing systems, to evaluate this model.
MATERIALS AND METHODS
Strains and growth conditions
P. aeruginosa and E. coli strains were stored at −80°C in 25% glycerol and routinely streaked onto Lennox LB medium 1.5% agar plates. Plates were incubated either overnight at 37°C or (for P. aeruginosa) 2 days at room temperature. Bacteria were inoculated aseptically into 2 mL sterile LB broth in borosilicate glass test tubes and grown overnight (~18–20 h) at 37°C with 225 RPM agitation. When necessary, LB was supplemented with gentamicin to concentrations of 30 µg/mL for chromosomal markers and 100 µg/mL for plasmid-based markers in P. aeruginosa, and 10 µg/mL for E. coli.
Genetic analysis of PAO1 Cpx
Gene deletion mutants were constructed in PAO1 by allelic exchange with the nonreplicative vector pEX18 as previously described (132, 133). Mutant genotypes were confirmed by PCR and Sanger sequencing. For complementation, wild-type gene alleles were PCR amplified from PAO1 genomic DNA (gDNA) using primers containing an artificial strong bacterial ribosome-binding (RBS) sequence to enhance transgene expression and cloned into an L-arabinose-inducible expression cassette (araBAD) in the replicative vector pJN105 (Gm^R^) using Gibson assembly (134). Plasmids were transformed into competent PAO1 strains by electroporation (135). 187 bp upstream of the cpxP (PA3205) start codon was PCR amplified from PAO1 gDNA, using primers to introduce an artificial strong bacterial RBS, and Gibson cloned upstream of a lacZ reporter gene into a linearized nonreplicative vector pUC18-miniTn7T (Gm^R^) (136). Tn7 integrated transformants were cured of the selective marker by FRT recombination with the plasmid pFLP (Cb^R^), which was subsequently cured by sacB counterselection (137). Overnight bacterial cultures were subcultured in LB containing gentamicin (for pJN105 plasmid retention) and 0.1% L-arabinose, as well as 2% ethanol to induce cellular stress where indicated, and grown 3 h at 37°C with 225 RPM agitation. 50 µL aliquots were transferred to a 96-well microplate, and turbidity (Abs_600_) was measured in a CLAIROstar PLUS microplate reader (BMG Labtech). Cells were lysed by a 1:2 dilution in B-PER II protein extraction reagent (Thermo Scientific). P_cpxP_::lacZ reporter activity was quantified by a standard β-galactosidase assay (138).
Fluorescence microscopy of Cpx reporter activity
We adapted a pBBR1-MCS5-based (Gm^R^) reporter plasmid described previously (8, 139), using Gibson assembly to fuse the cpxP gene promoter (187 bp) to a green fluorescent protein mGreenLantern (140, 141). This plasmid also contained a PrpoD::mScarlet-I red fluorescent protein housekeeping reporter for normalization (6, 142). Short half-life (ASV) variants of fluorescent proteins were utilized (8, 141), and all promoter fusions contained an artificial RBS to enhance reporter expression. This construct was used to report Cpx activity in both PAO1 and PA14, as the cpxP promoter region is > 99% identical between strains. Reporter activity was quantified by imaging cells with a Nikon Ti2 Eclipse widefield inverted microscope with a 100× objective oil-immersion lens, using phase contrast and fluorescence filters for GFP (Ex 466/40, Em 525/50) and TRITC (Ex 554/23, Em 609/54). Light intensity and exposure time were adjusted as needed to optimize visibility. Prior to imaging, cells were fixed in 2.5% paraformaldehyde in 1× phosphate-buffered saline (PBS), incubated >15 min at 37°C with agitation, and stored at 4°C for <48 h. 1–5 µL of fixed cells were spotted onto a glass coverslip and overlaid with a pad of 1% low melting point agarose in 1× PBS. At least 100 individual cells were imaged for all biological replicates. Images were processed using the MicrobeJ plugin for the Fiji distribution of ImageJ (143), with pre-processing to detect bacterial cells performed as described previously (8). Background-extracted mean fluorescence intensity values for individual cells were analyzed with Microsoft Excel or R. GraphPad Prism v10.3.0 was used for graphing and statistical analysis.
Cpx elicitor screen
Bacteria expressing the pBBR1 PcpxP::mGreenLantern/PrpoD::mScarlet-I reporter were grown overnight in LB with gentamicin selection and subcultured 1:100 in fresh LB supplemented with the indicated chemical stressors. After 4 h growth at 37°C with 225 RPM agitation, culture turbidity (Abs_600_) was measured by Genesys20 tube spectrophotometer, and cells were fixed and imaged by Nikon Ti2 Eclipse (10% light intensity, with 200 ms exposure for the GFP channel and 400 ms for the TRITC channel). We filtered out non-fluorescing cells due to plasmid loss, excluding all cells with <800 arbitrary fluorescence units (AFUs) in the PrpoD::mScarlet channel from further analysis. Bacteria were designated as Cpx^ON^ when normalized PcpxP::mGreenLantern/PrpoD::mScarlet fluorescent signal exceeded 2× the mean value from a control population grown in unstressed LB (72).
CRISPRi
Our approach is based on that described previously (75). An araBAD::spdcas9 cassette was PCR amplified from an integrated attB::araBAD::spdcas9 construct in PAO1, gifted by the Mekalanos lab. The araBAD::spdcas9 cassette was amplified from gDNA by PCR and cloned into the nonreplicative Tn7 integration plasmid, pUC18-miniTn7T (GmR), by Gibson assembly. A cassette containing both a 25-nucleotide sgRNA and a gRNA scaffold was expressed under SpeI constitutive promoter control on a pPSV37 plasmid; vectors containing bamA, lptD, and secY sgRNAs were also received from the Mekalanos Lab. All other sgRNA vectors were generated in this study by PCR amplification of the pPSV37 backbone with Gibson overhangs introducing sgRNA sequences, which were then DpnI treated and re-circularized by Gibson ligation. For validation of CRISPRi target inhibition, bacterial growth (Abs_600_) was assessed by CLAIROstar PLUS microplate reader (BMG Labtech), using overnight bacterial cultures diluted 1:100 into fresh LB media with gentamicin and 0.2% L-arabinose. To confirm inhibition of the nonessential gene dsbA by phenotypic screening, 5 µL of bacteria was spotted onto Vogel-Bonner Minimal Medium 1.5% agar plates containing 80 µg/mL Congo Red, 30 µg/mL Brilliant Blue, 100 µg/mL gentamicin, and 1% L-arabinose (144). To confirm inhibition of algU, the mucA22 mutation was introduced into PAO1 Tn7::araBAD::spdcas9 by allelic exchange, conferring a mucoid phenotype (145, 146). Strains were plated onto Pseudomonas Isolation Agar with gentamicin and 1% L-arabinose to confirm that sgRNA targeting of algU reversed the mucoid phenotype. To assess the impact of CRISPRi on Cpx activity, sgRNA expression cassettes were Gibson cloned into pBBR1 PcpxP::mGreenLantern/PrpoD::mScarlet-I. Bacterial cultures were diluted 1:100 in fresh LB with gentamicin and 0.2% L-arabinose and grown 4 h at 37°C with 225 RPM agitation. Cells were fixed and imaged by a Nikon Ti2 Eclipse as described above, using 20% light source intensity with 200 ms exposure for both GFP and TRITC channels. Due to plasmid loss among cells expressing toxic transgenes, we excluded non-fluorescing cells (<100 AFUs in the PrpoD::mScarlet channel) from further analysis. Bacteria were designated as Cpx^ON^ when the normalized PcpxP::mGreenLantern/PrpoD::mScarlet fluorescence intensity exceeded 2× the mean value from a control PAO1 population, which contained the Tn7::araBAD::spdcas9 cassette and fluorescent reporter plasmid with no sgRNA.
CpxR-binding site prediction
We queried the PAO1 genome (NC_002516.2, Pseudomonas Genome Database) for the CpxR consensus sequence (5′-GTAAA-(N)4-8-GTAAA-3′) (39), and a near-perfect 5′-GTAAA-(N)4-8-GTAA-3′ motif previously identified in Cpx-regulated promoters in E. coli (23). We included two additional CpxR motifs (5′-GTCAA-(N)4-8-GTAAA-3′ and 5′-GTAAA-(N)4-8-GTCAA-3′) identified through exponential enrichment (HT-SELEX) of DNA binding interactions by PAO1 CpxR in vitro (79). Of the 40 total sites identified by this method, 22 potential CpxR-regulated gene targets were selected based on the presence of at least one CpxR-binding motif within 500 bp upstream of the translation initiation codon. We additionally surveyed potential CpxR targets identified by a DAP-Seq analysis of the genomic binding profiles of >50 response regulators in PAO1 (82). To limit analysis to high-likelihood targets, we considered only binding sites that showed 10× or greater enrichment when precipitated with CpxR relative to a negative DAP-Seq control. Of 28 potential CpxR targets, 15 had already been identified by our analysis of CpxR consensus motifs. For the remaining 13 targets predicted by Trouillon et al., we estimated potential CpxR promoter-binding motifs based on the location of DAP-Seq peak summit positions, identified by the original authors using MASC2 followed by BEDtools analysis (147, 148). These sites typically contained one CpxR direct-repeat sequence (5′-GTAAA-3′) or similar motif. In cases where predicted CpxR sites fell within an intergenic region with two potential gene targets, we favored the gene in which the binding site was positioned approximately 50–200 bp upstream of the start codon, as binding site location is a strong predictor of Cpx regulation in E. coli (26).
RNA-Seq
Bacterial overnight cultures were subcultured 1:100 in LB and grown to an optical density (Abs_600_) of 0.52 ± 0.01 as measured by a Genesys20 test tube spectrophotometer. Cultures were normalized to Abs_600_ = 0.1. 1 mL (~1 × 10^8^ cells) was sedimented by centrifugation for 5 min at 17,000 × g and pellets were flash frozen in liquid nitrogen. Standard RNA preparation, including rRNA depletion and Illumina library preparation, was performed by Azenta Life Sciences, and paired-end 150 bp sequencing was performed on an Illumina HiSeq 3000. Trimmomatic was used to trim TruSeq3 adapter sequences and to filter low-quality reads (149). Reads were aligned to the reference PAO1 genome (NC_002516.2) with STAR (150, 151). Htseq-count was used to quantify reads overlapping genomic features as defined by a GFF3 annotation file obtained from Pseudomonas Genome DB (152). Differential gene expression (DGE) analysis was performed with DESeq2 (83). Operon prediction was performed on trimmed reads using Rockhopper (53).
qRT-PCR validation of DGE
PAO1 and PAO1 cpxS^T163P^ were grown to mid-exponential phase as described above for RNASeq sample preparation. 2 mL culture was pelleted by centrifugation and flash frozen in liquid nitrogen. Pellets were resuspended in 500 µL TRIzol (Invitrogen) and extracted with 100 µL chloroform. Samples were centrifuged at 17,000 × g for 15 min at 4°C to separate phases, and the aqueous phase was combined with an equal volume of ice-cold 100% ethanol. RNA was purified by the RNeasy Mini column-based kit (Qiagen). Nucleic acid was eluted in DEPC-treated ddH_2_O, and contaminating gDNA was removed using a TURBO DNA-free kit (ThermoFisher). Purified RNA was quantified by Qubit RNA HS assay using a Qubit 3 fluorometer (Invitrogen by ThermoFisher). Samples were normalized to 3 µg total RNA in DEPC-treated ddH_2_O prior to cDNA synthesis with an AffinityScript QPCR cDNA Synthesis kit (Agilent). qRT-PCR was performed using the SsoAdvanced Universal SYBR Green Supermix (BioRad) and a BioRad CFX384 thermocycler. DGE was calculated by the 2^−∆∆Ct^ formula (153).
Cpx surface sensing assay
P. aeruginosa with pBBR1 PcpxP::mGreenLantern/PrpoD::mScarlet-I was diluted 1:100 from overnight cultures grown at 37°C in LB broth for 3 h. 1 mL culture aliquots (~ 0.5 × 10^8^ CFUs) were sedimented by centrifugation for 10 min at 10,000 × g, and either fixed immediately in 2.5% paraformaldehyde in 1× PBS (planktonic cell fraction) or transferred to a sterile LB 1.5% agarose surface overlaid with 1 mL ddH_2_O in a 24-well microtiter plate. Plates were incubated statically at 37°C for 4 h. The supernatant was then decanted, and the agarose surface gently washed with 1 mL 1× PBS. The remaining surface-attached cell fraction was then resuspended in 200 µL of 2.5% paraformaldehyde by shaking for 20 min at 37°C. Cells were either imaged immediately or stored at 4°C for <48 h. Fluorescence microscopy was performed with a Nikon Ti2 Eclipse as described above, with exposure time set to 200 ms and Sola light source intensity set to 25% (GFP) and 10% (TRITC). Single-cell fluorescence intensity was quantified with MicrobeJ as previously detailed. Cells with PrpoD::mScarlet mean fluorescence intensity <100 AFU were omitted from further analysis; at least 200 individual cells were analyzed per biological replicate. Reporter activity was quantified by normalizing PcpxP::mGreenLantern to PrpoD::mScarlet fluorescence intensity values for each cell. Activity of a plasmid-based c-di-GMP responsive fluorescent reporter (pBBR1 PcdrA::mGreenLantern/PrpoD::CFP) was assessed by the same methods, using P_rpoD_::CFP signal for normalization (8). Both Sola (GFP) and Xylis (CFP) light sources were set to 20% intensity for experiments using this reporter. For each cell, the fold change in reporter activity was calculated relative to the mean value of an experimentally defined negative control population (reporter^OFF^), with cells exhibiting a >5× fold change designated reporter^ON^. For Cpx reporter assays, the planktonic cell population of PAO1 ∆cpxR was used to represent the reporter^OFF^ condition; for c-di-GMP reporter assays, the planktonic population of a c-di-GMP-deficient strain, PAO1 ∆wspR∆sadC∆siaB, defined the reporter^OFF^ condition.
Surface attachment assay
PAO1 and derivative mutant strains carrying the pBBR1 PcpxP::mGreenLantern/PrpoD::mScarlet-I reporter plasmid were subcultured (1:100) from overnight cultures into fresh LB broth and grown 3 h at 37°C. 1 mL exponential-phase culture was combined with 1 mL ddH_2_O in a 12-well polystyrene microplate, and 22 × 22 mm glass coverslips (VWR) were submerged vertically in wells. Plates were incubated statically at 37°C, and coverslips were removed at the indicated times and submerged in a reservoir of 0.5% crystal violet stain in ddH_2_O for 15 min. Coverslips were washed thrice in ddH_2_O to remove residual staining and destained in 0.5 mL 30% acetic acid solution. Attachment was quantified by measuring absorbance at 562 nm of destained solution in a CLAIROstar PLUS microplate reader.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Donlan RM. 2001. Biofilm formation: a clinically relevant microbiological process. Clin Infect Dis 33:1387–1392. doi:10.1086/32297211565080 · doi ↗ · pubmed ↗
- 2Costerton JW, Stewart PS, Greenberg EP. 1999. Bacterial biofilms: a common cause of persistent infections. Science 284:1318–1322. doi:10.1126/science.284.5418.131810334980 · doi ↗ · pubmed ↗
- 3Laventie BJ, Jenal U. 2020. Surface sensing and adaptation in bacteria. Annu Rev Microbiol 74:735–760. doi:10.1146/annurev-micro-012120-06342732905753 · doi ↗ · pubmed ↗
- 4Schniederberend M, Williams JF, Shine E, Shen C, Jain R, Emonet T, Kazmierczak BI. 2019. Modulation of flagellar rotation in surface-attached bacteria: a pathway for rapid surface-sensing after flagellar attachment. P Lo S Pathog 15:e 1008149. doi:10.1371/journal.ppat.100814931682637 PMC 6855561 · doi ↗ · pubmed ↗
- 5Luo Y, Zhao K, Baker AE, Kuchma SL, Coggan KA, Wolfgang MC, Wong GCL, O’Toole GA. 2015. A hierarchical cascade of second messengers regulates Pseudomonas aeruginosa surface behaviors. m Bio 6:e 02456-14. doi:10.1128/m Bio.02456-1425626906 PMC 4324313 · doi ↗ · pubmed ↗
- 6Persat A, Inclan YF, Engel JN, Stone HA, Gitai Z. 2015. Type IV pili mechanochemically regulate virulence factors in Pseudomonas aeruginosa. Proc Natl Acad Sci USA 112:7563–7568. doi:10.1073/pnas.150202511226041805 PMC 4475988 · doi ↗ · pubmed ↗
- 7O’Neal L, Baraquet C, Suo Z, Dreifus JE, Peng Y, Raivio TL, Wozniak DJ, Harwood CS, Parsek MR. 2022. The Wsp system of Pseudomonas aeruginosa links surface sensing and cell envelope stress. Proc Natl Acad Sci USA 119:e 2117633119. doi:10.1073/pnas.211763311935476526 PMC 9170161 · doi ↗ · pubmed ↗
- 8Zheng X, Gomez-Rivas EJ, Lamont SI, Daneshjoo K, Shieh A, Wozniak DJ, Parsek MR. 2024. The surface interface and swimming motility influence surface-sensing responses in Pseudomonas aeruginosa. Proc Natl Acad Sci USA 121:e 2411981121. doi:10.1073/pnas.241198112139284057 PMC 11441478 · doi ↗ · pubmed ↗