Disruption of putrescine export in experimentally evolved Ralstonia pseudosolanacearum enhances symbiosis with Mimosa pudica
Anne-Claire Cazalé, Marvin Navarro, Ginaini Grazielli Doin de Moura, David Hoarau, Floriant Bellvert, Sophie Valière, Caroline Baroukh, Philippe Remigi, Alice Guidot, Delphine Capela

TL;DR
Disrupting putrescine export in a plant pathogen helps it become a better symbiont for a legume, showing how polyamines affect plant-microbe interactions.
Contribution
The study identifies that inactivating the paeA gene, which encodes a putrescine exporter, is key to enhancing symbiosis in Ralstonia pseudosolanacearum with Mimosa pudica.
Findings
Mutations in paeA abolished putrescine excretion and increased bacterial proliferation in legume nodules.
paeA-mutated bacteria induced functional symbiosis by modulating host gene expression toward nodule development.
Bacterial-derived putrescine acts as a negative signal for the plant, while plant-derived putrescine has distinct roles in symbiosis.
Abstract
Polyamines are essential molecules across all domains of life, but their role as signaling molecules in host–microbe interactions is increasingly recognized. However, because they are produced by both the host and the microbe, their dual origin makes their functional dissection challenging. The plant pathogen Ralstonia pseudosolanacearum GMI1000 secretes large amounts of putrescine both in vitro and in the xylem sap of host plants. In this study, we investigated the genetic changes underlying its experimental evolution into a legume symbiont. We showed that the paeA gene (RSc2277), which was repeatedly mutated during this process, encodes a putrescine exporter. Mutations in paeA completely abolished putrescine excretion in vitro and enhanced bacterial proliferation within nodules during interaction with the legume Mimosa pudica. When these mutations occurred in symbionts already capable…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
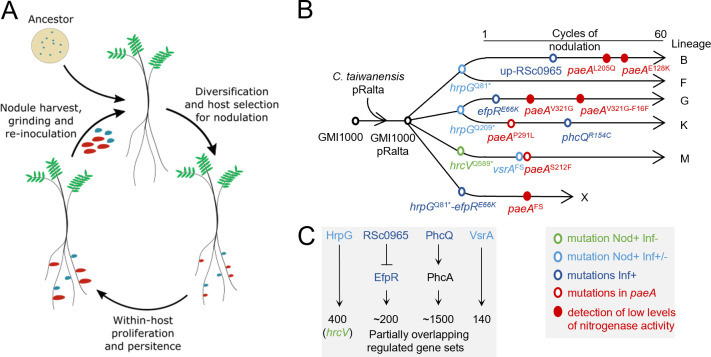 Fig 1
Fig 1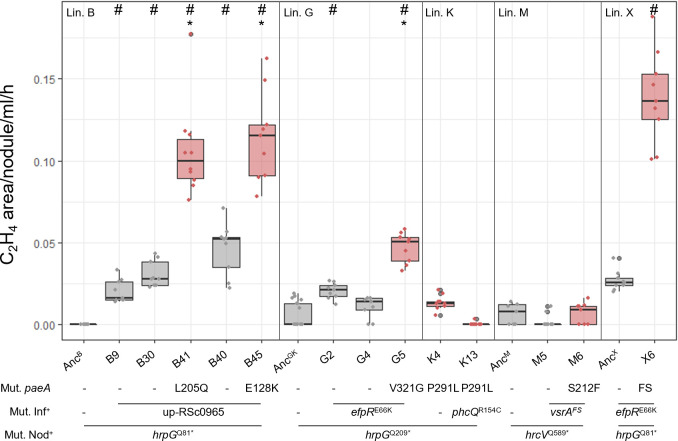 Fig 2
Fig 2 Fig 3
Fig 3 Fig 4
Fig 4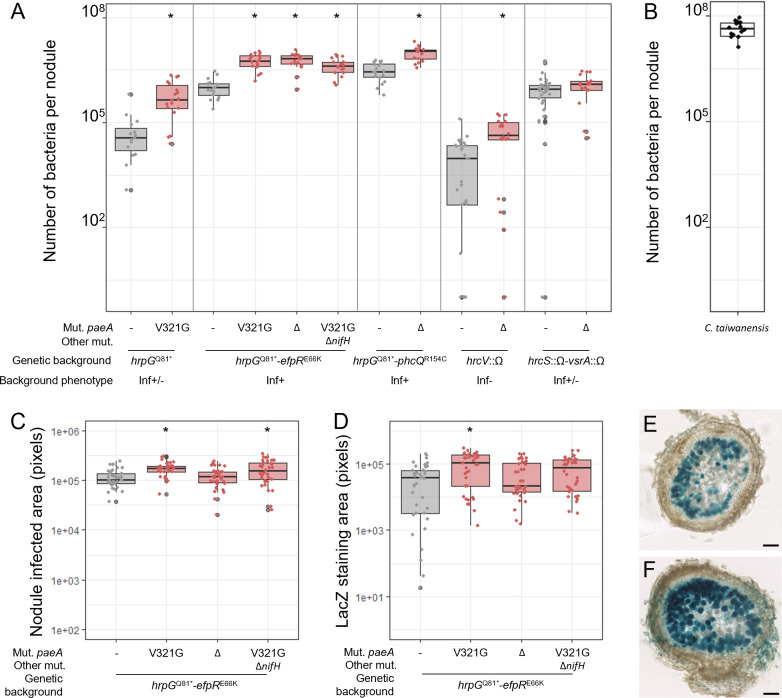 Fig 5
Fig 5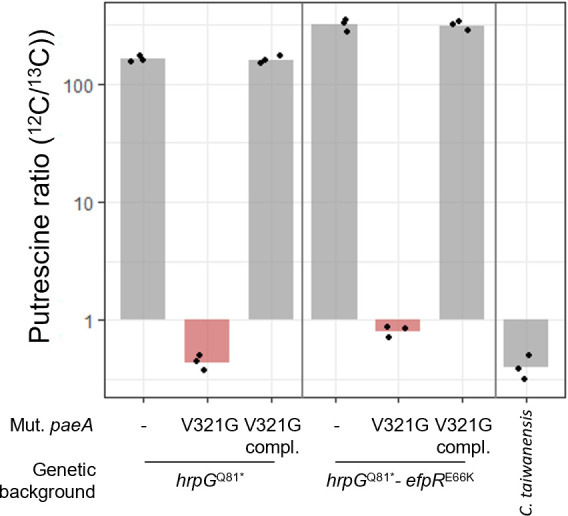 Fig 6
Fig 6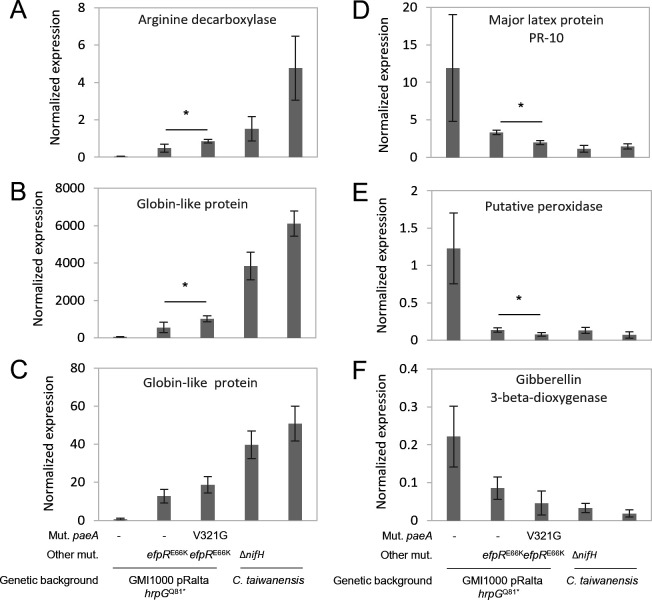 Fig 7
Fig 7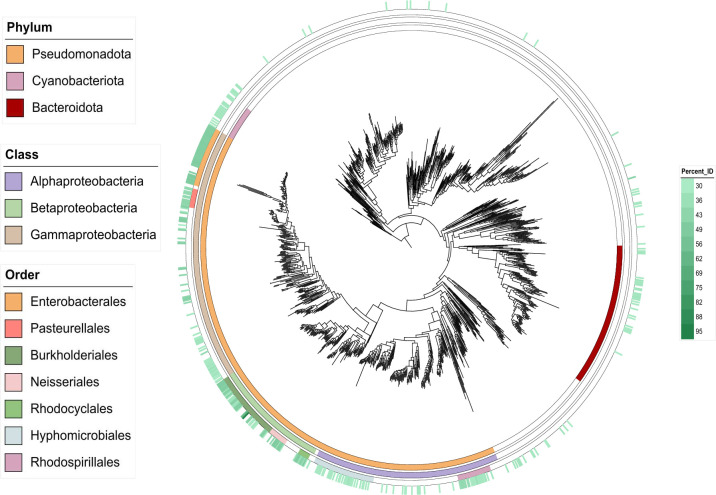 Fig 8
Fig 8| Lineage | Evolved clone | Position on the chromosome | Mutation | Protein modification | Reference |
|---|---|---|---|---|---|
| B | B41 | 2473740 | T/A | L205Q | This study |
| B | B45 | 2473508 | G/A | E128K | This study |
| G | G5 | 2474088 | T/G | V321G | ( |
| G | G16 | 2473174 | C/T | F16F | ( |
| K | K4 | 2473998 | C/T | P291L | ( |
| M | M6 | 2473761 | C/T | S212F | ( |
| X | X6 | 2473453 | G/- | Frameshift | This study |
- —Agence Nationale de la Recherchehttp://dx.doi.org/10.13039/501100001665
- —Agence Nationale de la Recherchehttp://dx.doi.org/10.13039/501100001665
- —Agence Nationale de la Recherchehttp://dx.doi.org/10.13039/501100001665
- —Agence Nationale de la Recherchehttp://dx.doi.org/10.13039/501100001665
- —Agence Nationale de la Recherchehttp://dx.doi.org/10.13039/501100001665
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegume Nitrogen Fixing Symbiosis · Polyamine Metabolism and Applications · Plant Pathogenic Bacteria Studies
INTRODUCTION
Bacteria known as rhizobia establish mutualistic interactions with legumes, resulting in the formation of specialized plant organs called nodules. Within these nodules, hundreds of thousands of bacteria reside intracellularly, fixing atmospheric nitrogen into ammonia to benefit the plant. In return, bacteria receive carbon sources from the plant along with protection from the external environment. To initiate this interaction, most rhizobia synthesize lipochitooligosaccharides, called Nod factors, which are specifically recognized by plant receptors and trigger nodule organogenesis and early infection processes (1–5). Bacteria penetrate the roots either through crack entry (intercellular infection) or by attaching to root hairs, which subsequently invaginate to form infection threads (6–8). These infection threads progress through the root cell layers, guiding the bacteria toward the developing nodule. Then, bacteria are released into the cytoplasm of nodule cells via an endocytosis-like process and differentiate into nitrogen-fixing bacteroids. Intracellular bacteroids are enclosed by a plant-derived membrane, forming structures known as symbiosomes (9, 10). Within symbiosomes, bacteroids face challenging conditions, including highly acidic pH, extremely low oxygen concentrations, the presence of reactive oxygen species, and likely high osmotic pressure. All stages of this symbiotic interaction are tightly regulated by mutual recognition processes (11). While the bacterial genes involved in the early stages of symbiosis have been well studied (12, 13), those required for the later stages, such as the survival and persistence of bacteria within nodule cells, remain far less understood. This is particularly the case outside terminally differentiated systems specific to certain legumes, such as Medicago and Pisum in the inverted repeat-lacking clade (IRLC) or Aeschynomene species in the Dalbergioid clade (14, 15). In these interactions, the proliferation and differentiation of intracellular bacteria is controlled by plant peptides called nodule-specific cysteine-rich (NCR) or NCR-like peptides (15, 16). These peptides interfere with various bacterial cellular processes, inhibiting cell division while promoting nitrogen fixation (17–21). In such cases, symbiotic bacteria must possess the specific transporters, BacA or BclA (22–24), modified lipopolysaccharides (LPS) (25), or peptidoglycan-modifying enzymes (26) to survive within nodule cells.
Rhizobia are polyphyletic bacteria that belong to 21 different genera and hundreds of species among two classes of proteobacteria: alpha and beta (27). The symbiotic capacity of these bacteria emerged following independent and repeated horizontal transfers of symbiotic genes essential for the production of Nod factors (nod genes) and the synthesis and functioning of nitrogenase (nif and fix genes). However, the transfer of nod, nif, and fix genes is not always sufficient to convert a strain that receives these genes into a functional nitrogen-fixing legume symbiont (28–31). In a previous evolution experiment, we transferred the symbiotic plasmid of Cupriavidus taiwanensis LMG19424, a natural symbiont of Mimosa pudica, into the plant pathogenic bacterium Ralstonia pseudosolanacearum GMI1000. The resulting chimeric strain was unable to nodulate M. pudica. However, after multiple large-scale inoculation trials involving hundreds of plants, three nodules appeared, from which we isolated three independent nodulating variants of R. pseudosolanacearum (32). These variants acquired the ability to nodulate M. pudica through mutations that inactivated the major determinant of R. pseudosolanacearum pathogenicity, its type III secretion system (T3SS). Mutations that conferred nodulation affected either the master regulator hrpG (33) or the T3SS structural gene hrcV. The three nodulating strains were then submitted to serial cycles of nodulation on M. pudica. The symbiotic properties of bacteria improved rapidly during the first cycles and then slowed down (34). After 35 evolution cycles, nodulation competitiveness was almost equivalent to that of the natural rhizobium C. taiwanensis, while bacterial proliferation within the nodules, although greatly improved over the cycles, did not reach the level of C. taiwanensis and nitrogen fixation was not achieved. Strong adaptive mutations that improve the symbiotic properties of R. pseudosolanacearum were previously identified. In particular, intracellular infection was enhanced through mutations in the EfpR or PhcA regulatory pathways (32, 35, 36). EfpR and PhcA are master regulators controlling hundreds of genes, either positively or negatively (35, 37–41), with approximately 160 genes being commonly regulated by both. These shared targets include genes involved in EPS synthesis and motility, hemin/siderophore transport, and metabolism genes, as well as genes encoding Hrp and T3 effector proteins. Additionally, mutations in phcA and efpR have been shown to broadly activate bacterial metabolic activities (35, 37, 38, 42). On the plant side, the analysis of M. pudica gene expression profiles in response to progressively adapted R. pseudosolanacearum strains revealed a correlation between bacterial adaptation and a gradual increase in the number of plant genes that are differentially expressed and also differentially expressed during the interaction with the natural symbiont C. taiwanensis strain (43).
In this study, we continued the evolution experiment until 60 cycles in order to further enhance the symbiotic capacities of bacteria. Among the genes that were repeatedly mutated in this experiment, we identified the RSc2277 gene, which was mutated six times independently. Interestingly, low levels of nitrogenase activity could be detected in nodules formed by some evolved clones carrying a mutation in this gene. We showed that the inactivation of RSc2277 is highly adaptive, as it significantly increased bacterial proliferation within nodules and led to the detection of nitrogenase activity. The RSc2277 gene encodes a protein similar to the PaeA proteins of Salmonella typhimurium and Escherichia coli. In these bacteria*,* PaeA has been shown to function as a cadaverine/putrescine exporter, which is critical for bacterial survival under some stress conditions, likely by reducing toxic levels of intracellular polyamines and maintaining cation homeostasis (44, 45). R. pseudosolanacearum GMI1000 is known to synthesize and export large quantities of putrescine in its natural habitat, the xylem of tomato (46, 47), as well as in cultures on various carbon sources (42, 48). However, the mechanisms of putrescine export in R. pseudosolanacearum have not been evidenced. Here, we show that inactivation of a PaeA homolog in R. pseudosolanacearum completely abolished putrescine export in culture and improved symbiosis with M. pudica.
RESULTS
The paeA (RSc2277) gene is repeatedly mutated in four independent parallel lineages of R. pseudosolanacearum experimentally evolved into legume symbionts
In a previous study, we described the dynamics of occurrence of mutations in R. pseudosolanacearum populations from five lineages (B, F, G, K, M) evolved for 35 nodulation cycles on the legume Mimosa pudica. This analysis revealed genes that were mutated at a higher frequency than expected by chance. Among the most frequently mutated genes, RSc2277, which we designated paeA based on its homology to functionally characterized genes in Salmonella Typhimurium and E. coli (44, 45), was found mutated in four evolved clones from three lineages (lineages G, K, and M). These four mutations were fixed in populations, which can be indicative of their adaptive character (Table 1). Indeed, in the G lineage, Doin de Moura et al. (34) previously showed that the V321G mutation in PaeA, occurring in cycle 5, was highly adaptive, increasing bacterial in planta fitness by over 100-fold. When we continued the evolution of the five lineages up to 60 cycles of nodulation, two new mutations in paeA occurred in evolved clones of the B lineage at cycles 41 and 45 (Fig. 1 and Table 1). The R. pseudosolanacearum PaeA protein contains a signal peptide, three transmembrane domains, two tandem cystathionine beta-synthase (CBS) domains, and a CorC-HlyC C-terminal domain (Fig. S1C). This protein shares 48% and 46% sequence identity with the PaeA proteins from Salmonella enterica serovar Typhimurium and E. coli, respectively, for which a polyamine export function has been demonstrated (44, 45). It also displays a similar three-dimensional structure (Fig. S1A and B).
Occurrence of paeA mutations during the experimental evolution of R. solanacearum into Mimosa pudica symbionts. (A) Overview of the evolution experiment. In each cycle, inoculated bacteria diversify due to a transient hypermutagenesis phenomenon that occurs in the rhizosphere (49), and the most competitive variants for host entry are selected by the plants to form nodules (34). Within the nodules, bacteria multiply before the nodules are harvested and ground to inoculate plants of the next cycle. (B) To initiate the evolution experiment, the symbiotic plasmid pRalta of C. taiwanensis was introduced into the GMI1000 strain of R. pseudosolanacearum. Three initial nodulation variants were obtained when the resulting strain GMI1000 pRalta was massively inoculated onto M. pudica (32). Then, five independent lineages (B, F, G, K, M) were derived from these three ancestors and evolved for 60 cycles. The sixth lineage X was derived from the hrpGQ81-efpRE66K reconstructed mutant and evolved for 15 cycles. Red circles indicate the occurrence of paeA mutations along the experiment. In some cases, this mutation was associated with the detection of nitrogenase activity (filled red circles). The main adaptive mutations allowing nodulation (hrpGQ81*, hrpG,Q209* and hrcVQ589* stop mutations) and intracellular infection (hrpGQ81*, hrpGQ209*, up-RSc0965, efpRE66K, phcQ,R154C and vsrAFS) were identified (32, 35, 36, 50). (C) A simplified schematic of the R. pseudosolanacearum regulatory pathways involving these mutations is provided. The HrpG, EfpR, PhcA, and VsrA regulons have been identified and are partially overlapping (33, 35, 37–41). Stop mutation. Up, intergenic mutation upstream of the start codon. FS, frameshift mutation.
Nitrogenase activity was detected in some Ralstonia evolved clones mutated in paeA
Mutualism was not achieved in any of the five lineages even after 60 cycles of evolution on Mimosa pudica. However, low levels of nitrogenase activity measured by acetylene reduction assays (ARA) were detected in nodules formed by some evolved clones. Interestingly, higher levels of nitrogenase activity were specifically detected in evolved clones B41, B45, and G5, all three carrying a mutation in paeA. However, no nitrogenase activity was detected in clones K4 and M6, both also carrying a mutation in paeA. One hypothesis is that the different mutations do not have the same consequence on PaeA protein function. However, the more appealing hypothesis was that paeA could trigger an increase in nitrogenase activity depending on the bacterial genetic background in which these mutations occurred. Indeed, in the nitrogen-fixing clones B41, B45, and G5, the paeA mutations occurred in evolved bacteria that infect nodule cells very well due to mutations in both hrpG (Q81* or Q209* stop mutations) and in the efpR pathway (mutations up-RSc0965 or efpR^E66K^) (35) (Fig. 1). In the non-fixing evolved clones K4 and M6, mutations in paeA occurred in bacteria that infect nodule cells only partially, due to mutations in hrpG (Q81* or Q209* stop mutations) (32) or hrcV (Q589* stop mutation) and vsrA (frameshift) (50), respectively (Fig. 1). In the K lineage, a mutation improving intracellular infection (the mutation phcQ^R154C^) cumulated with the paeA mutation in the clone K13, but no nitrogenase activity was detected in nodules induced by this clone (Fig. 2). Moreover, we started a new evolution lineage (lineage X), this time using as ancestor a reconstructed mutant, GMI1000 pRalta hrpG^Q81*^ efpR^E66K^, that infects nodule cells well. After a few cycles, a frameshift mutation in paeA occurred and was again associated with detectable levels of nitrogenase activity in nodules (clone X6; Fig. 2).
*Nitrogenase activity in nodules formed by Ralstonia-evolved clones. Acetylene reduction assays were performed on plants inoculated with isolated evolved clones 15 days after inoculation. Red box plots correspond to evolved clones carrying a mutation in paeA. Each measurement was taken from a pool of six plants. At least three measurements per experiment and three independent experiments were performed per strain. Lin., lineage. AncB, AncGK, AncM, AncX, nodulating ancestors of the B, G, K, M, or X lineages. Mut. Nod+, mutation conferring the nodulation capacity. Mut. Inf+, main mutation conferring the capacity to infect nodules intracellularly. FS, frameshift mutation. The clone B30 is the closest evolved ancestor of clone B41, and the clone B40 is the closest evolved ancestor of clone B45 (see the phylogeny of evolved clones of the B lineage in Fig. S2). The complete list of mutations present in evolved clones is provided in Table S3. #, significantly different from the nodulating ancestor; , significantly different from the closest evolved ancestor (P < 0.05, pairwise Wilcoxon test).
Inactivation of paeA allows the detection of low levels of nitrogenase activity in Ralstonia clones mutated in the efpR or phc regulatory pathways
Evolved clones have accumulated many mutations during the evolution experiment (34, 49). In order to know whether the paeA mutation is responsible for the detected nitrogenase activity, we reconstructed the paeA^V321G^ mutation and an unmarked deletion of paeA in the strain GMI1000 pRalta carrying the hrpG^Q81*^ and efpR^E66K^ mutations, which confer nodulation and nodule intracellular infection capacity, respectively (32, 35). We measured the nitrogenase activity by ARA in nodules induced by these mutants. At 15 days post-inoculation (dpi), we detected nitrogenase activity in both paeA^V321G^ and paeA deletion mutants, but not in the parental strain (GMI1000 pRalta hrpG^Q81*^ efpR^E66K^). This shows that mutations in paeA in this background allowed the detection of nitrogenase activity in nodules and that the paeA^V321G^ mutation is equivalent to a loss of function (Fig. 3A). No nitrogenase activity was detected in the same strain in which we deleted the nifH gene, meaning that the ethylene measured in ARA is indeed due to the functioning of nitrogenase. Moreover, complementing the paeA^V321G^ mutant with the wild-type allele abolished nitrogenase activity (Fig. S3A).
Acetylene reduction assays of plants inoculated with Ralstonia paeA reconstructed mutants (A) and C. taiwanensis (B), 15 days post-inoculation. The paeAV321G mutation or paeA deletion was reconstructed in different Ralstonia GMI1000 pRalta genetic backgrounds, either hrpGQ81 (partially intracellular infectious) (32), hrpGQ81*-efpRE66K (nicely intracellular infectious) (35), hrpGQ81*-phcQR154C (nicely intracellular infectious) (36), hrcV::Ω (extracellular infectious) (30), or hrcS::Ω-vsrA::Ω (partially intracellular infectious) (50). At 15 dpi, plants inoculated with the mutants (A) or the natural symbiont C. taiwanensis (B) were incubated with an excess of acetylene for four hours. Ethylene produced was measured by gas chromatography. Areas of ethylene peaks were integrated and normalized by the number of nodules, the volume of gas analyzed, and the time of incubation with acetylene. Red box plots correspond to the measures made with the paeA mutants. At least three independent experiments with three measures per experiment were performed for each strain. , statistically different from the parental strain (P < 0.05, pairwise Wilcoxon test).
The nitrogen-fixing capacity of paeA mutants is not specific to the hrpG-efpR background, as we could also measure nitrogenase activity in nodules induced by a hrpG-phcQ-ΔpaeA mutant, another genetic background conferring a good level of intracellular infection in nodules (36). In contrast, no nitrogenase activity was detected in nodules induced by hrcV-ΔpaeA, hrcS-vsrA-ΔpaeA, or hrpG-paeA^V321G^ mutants (Fig. 3A), three genetic backgrounds known to confer only extracellular or partially intracellular infection capacity (32, 50).
Notably, the levels of nitrogenase activity detected in hrpG-efpR or hrpG-phcQ backgrounds are very low, representing less than 1% of the levels measured at 15 dpi with the natural symbiont Cupriavidus taiwanensis LMG19424 (Fig. 3B), and do not support plant growth (Fig. S4). Moreover, this activity is transient as it decreased at 21 dpi (Fig. S5). We also examined the activity of a nifH promoter*-gus* transcriptional fusion introduced in the genomes of the hrpG-efpR and hrpG-efpR-paeA mutant strains in nodules harvested at 10, 15, and 21 dpi (Fig. 4A). We observed that the expression of this fusion was slightly but significantly higher in the paeA mutant than in the parental strain at all time points. In the paeA mutant strain, nifH expression was maximal at 10 dpi and decreased to a very low level at 15 and 21 dpi. At these latter time points, only a few nodule cells showed GUS activity. These results were confirmed at the protein level by Western blot (Fig. 4B).
NifH expression in nodules. (A) The activity of a genomic transcriptional nifH promoter-gus fusion was quantified in nodules formed by the Ralstonia GMI1000 pRalta hrpGQ81-efpRE66K-paeAV321G mutant strain (red box plots) and its parental strain (gray box plots) at 10, 15, and 21 dpi. The percentage of nodule cells expressing nifH (blue-stained area/(blue stained + brawn stained areas)) was measured on 60 µm nodule sections. Data are from three independent experiments, with at least 24 nodules analyzed in each and several sections quantified per nodule. Representative images of nodule sections are shown for each strain at each time point. , values measured with the paeA mutant are statistically different from the parental strain (P < 0.05, pairwise Wilcoxon test). (B) NifH protein detected by Western blot in nodules formed by the hrpGQ81-efpRE66K-paeAV321G mutant strain and its parental strain at 10, 15, and 21 dpi. Equal loading was verified using an α-tubulin antibody. The expected sizes of NifH and tubulin proteins are 32.5 kDa and 52 kDa, respectively.*
Inactivation of paeA increases bacterial proliferation in nodules
To determine whether the observed nitrogenase activity resulted from increased bacterial proliferation in nodules, we quantified the number of viable bacteria recovered from nodules induced by paeA mutants reconstructed in different genetic backgrounds. These backgrounds, hrpG, hrpG-efpR, hrpG-phcQ, hrcV, and hrcS-vsrA, represent the various evolutionary contexts in which paeA mutations arose. We found that both the paeA^V321G^ point mutation and the deletion of paeA significantly enhanced bacterial proliferation within nodules across all tested backgrounds (by a factor of 3 to 8), with the exception of the hrcS-vsrA background (Fig. 5A).
Nodule infection of paeA reconstructed mutants. The paeAV321G mutation and paeA deletion (Δ) were reconstructed in different Ralstonia GMI1000 pRalta genetic backgrounds, hrpGQ81, hrpGQ81*-efpRE66K, hrpGQ81*-efpRE66K-ΔnifH, hrpGQ81*-phcQR154C, hrcV::Ω, or hrcS::Ω-vsrA::Ω. (A) Proliferation of paeA mutants and parental strains in 15-day-old nodules. (B) Proliferation of C. taiwanensis in 15-day-old nodules. (C) Total intracellularly infected areas (blue-stained + brawn-stained plant cells) and (D) blue-stained areas measured on 60 µm sections of 15-day-old nodules induced by paeA mutants and the corresponding parental strain constitutively expressing the lacZ gene. (E, F) LacZ-stained representative nodules formed by the hrpGQ81*-efpRE66K (E) and hrpGQ81*-efpRE66K-paeAV321G (F) mutants, both expressing constitutively the lacZ gene. Bars represent 100 µm length. (A to F) Data are from at least three independent experiments. (A, C, D) Red box plots correspond to measures made with the paeA mutants. , statistically different from the parental strain (P < 0.05, pairwise Wilcoxon test).
The highest numbers of bacteria recovered per nodule were observed when paeA mutations were combined with other highly infective mutations, such as efpR^E66K^ or phcQ^R154C^, in the same genetic backgrounds in which nitrogenase activity was detected. Complementation of the paeA^V321G^ mutant with the wild-type allele in the hrpG-efpR background reduced bacterial proliferation in nodules (Fig. S3B). Furthermore, inactivation of the nifH gene in the hrpG-efpR-paeA mutant did not significantly affect the number of viable bacteria per nodule (Fig. 5A). These results suggest that nitrogenase activity is not driving the enhanced proliferation of paeA mutants. Rather, we propose the opposite: increased bacterial numbers in nodules may enhance the detectability of nitrogenase activity that would otherwise remain below detection thresholds.
We next investigated whether the higher number of viable bacteria recovered per nodule could be attributed to improved bacterial survival within nodule cells. To explore this, we performed live/dead staining on nodule sections. At 15 days post-inoculation (dpi), no significant differences were observed between nodules induced by the paeA^V321G^ mutant and those induced by its parental hrpG-efpR strain. By this time point, bacterial degeneration had already begun in both types of nodules (Fig. S6).
As an alternative approach, we used strains constitutively expressing the lacZ gene to quantify nodule cells containing the LacZ protein, whose persistence may outlast early stages of bacterial degeneration. Nodule sections were stained with X-gal, and we quantified both the area of blue-stained cells (invaded by bacteria still expressing LacZ) and brown-stained cells (invaded by bacteria that had ceased expressing LacZ). At 15 dpi, the blue-stained area was slightly larger in nodules induced by the paeA^V321G^ mutant (Fig. 5D through F), suggesting a modest enhancement in bacterial survival. Additionally, the total infected area per nodule section (blue + brown cells) was marginally larger in nodules induced by the mutant, indicating a slight increase in the number of cells invaded (Fig. 5C). However, these effects were minor and were not observed in the paeA deletion mutant.
Overall, these results suggest that paeA mutations exert several additive effects, including increased bacterial proliferation within nodule cells, a slightly greater number of invaded cells, and slightly improved bacterial survival. Collectively, these factors contribute to a fivefold increase, on average, in the number of viable bacteria recovered per nodule compared to nodules induced by the corresponding parental strains.
PaeA inactivation impairs putrescine export
To determine whether the R. pseudosolanacearum PaeA is involved in polyamine export, such as in Salmonella Typhimurium and E. coli (44), we analyzed culture supernatants from parental strains and paeA mutants reconstructed in two genetic backgrounds, hrpG and hrpG-efpR, using mass spectrometry. The strains were grown in a synthetic medium with glutamine as the sole carbon source. In this medium, no growth difference was observed between the paeA mutants and the corresponding parental strains (Table S1). Consistent with previous findings, we confirmed that R. pseudosolanacearum GMI1000 pRalta, such as GMI1000, produces very high amounts of putrescine in this medium (42, 46, 48). These levels are slightly increased in the hrpG-efpR mutant compared to the simple hrpG mutant. In contrast, we observed that putrescine levels were extremely low in the supernatants of paeA mutants in both the hrpG and hrpG-efpR genetic contexts. Complementation of the paeA mutants with the wild-type allele fully restored putrescine secretion (Fig. 6), confirming the major role of this protein in putrescine export. It is also noteworthy that the level of secreted putrescine in the natural symbiont C. taiwanensis was also very low when cultured in the same medium, despite the presence of a putrescine biosynthesis gene (RALTA_A2412) and a close homolog of paeA (RALTA_A0755) in the C. taiwanensis LMG19424 genome (Fig. S1A).
Quantification of putrescine in culture supernatants. The R. pseudosolanacearum paeA mutants reconstructed in GMI1000 pRalta hrpGQ81 or GMI1000 pRalta hrpGQ81*-efpRE66K backgrounds, the corresponding parental and complemented strains, and C. taiwanensis were grown in minimal medium containing 10 mM glutamine until an optical density at 600 nm equal to 1. The supernatants were separated from the cells by centrifugation and then filtered at 0.2 µm in order to quantify only extracellular putrescine. Quantification of putrescine was carried out by high-resolution mass spectrometry by calculating the ratio between putrescine in the sample and the internal standard (full 13C-labeled putrescine). Compl., complementation of the paeAV321G mutant with the paeA wild-type allele introduced at the native locus. Data are from three independent experiments.*
Inactivation of paeA in Ralstonia symbionts modulates plant gene expression toward a more functional symbiotic state
In a previous study using both R. pseudosolanacearum and C. taiwanensis strains, we identified Mimosa genes whose expression in nodules correlated with the adaptation level of the symbionts (43). Among these genes, the putrescine biosynthesis gene encoding an arginine decarboxylase (ADC) was found to be highly expressed in C. taiwanensis-infected nodules, whereas it showed only weak expression in nodules infected with the R. pseudosolanacearum GMI1000 pRalta hrpG efpR mutant. Similarly, genes essential for functional nodules, such as leghemoglobins, exhibited low expression in nodules infected by R. pseudosolanacearum but high expression in nodules infected by C. taiwanensis. Additionally, we identified Mimosa genes whose expression was negatively correlated with bacterial adaptation to symbiosis. For instance, genes putatively involved in defense responses, such as pathogenesis-related proteins of class 10 (PR10) (51) and peroxidases (52, 53), as well as some genes associated with gibberellin biosynthesis—a phytohormone known to be finely regulated during nodule development (54)—were progressively downregulated.
To further investigate the correlation between the expression of these plant genes and bacterial adaptation, we analyzed their expression by qRT-PCR in nodules induced by the R. pseudosolanacearum hrpG-efpR-paeA mutant. We compared these results to nodules induced by the R. pseudosolanacearum hrpG and hrpG-efpR mutants, as well as to nodules induced by C. taiwanensis, including either the wild-type strain or a nitrogen-fixation-deficient nifH mutant. Nodules were harvested at 10 dpi, an early stage chosen to ensure that Ralstonia bacteria had not yet significantly degenerated and because it corresponded to the maximum expression of nifH detected. The expression of these genes in nodules induced by the hrpG-efpR-paeA mutant differed only slightly from their expression in nodules induced by the hrpG-efpR mutant. However, expression levels consistently fell between those observed in nodules induced by the hrpG-efpR mutant and those in nodules induced by the C. taiwanensis non-fixing mutant (Fig. 7). Specifically, the putrescine biosynthesis gene ADC and two globin-encoding genes exhibited increased expression, whereas PR10, a peroxidase, and a gibberellin 3-beta dioxygenase encoding gene were down-regulated in nodules induced by the hrpG-efpR-paeA mutant compared to nodules induced by the hrpG-efpR mutant. These data indicate that R. pseudosolanacearum symbionts carrying mutations in paeA triggered plant gene expression changes, shifting nodules toward a more functional symbiotic state.
Gene expression of Mimosa pudica in response to progressively adapted R. pseudosolanacaerum symbionts and C. taiwanensis strains. Expression of M. pudica genes encoding an arginine decarboxylase (MpudA1P6v1r1_Scf09g0369161) (A), two globin-like proteins (MpudA1P6v1r1_Scf07g0334541 (B) and MpudA1P6v1r1_Scf29g0605241 (C)), a major latex protein PR-10 (MpudA1P6v1r1_Scf15g0080361) (D), a putative peroxidase (MpudA1P6v1r1_Scf04g0255631) (E), and a gibberellin 3-beta-dioxygenase (MpudA1P6v1r1_Scf32g0613491) (F) was measured in 10-day-old nodules by qRT-PCR. Gene expressions were normalized by three reference genes encoding a putative ubiquitin protein (MpudA1P6v1r1_Scf06g0302791) and two putative DNA helicases (MpudA1P6v1r1_Scf39g0686881 and MpudA1P6v1r1_Scf10g0050861). wt, wild-type strain of C. taiwanensis. ΔnifH, non-fixing strain of C. taiwanensis deleted in the nifH gene. Data represent mean values and standard deviations from four independent experiments. , significant difference between nodules induced by hrpGQ81-efpRE66K and hrpGQ81-efpRE66K-paeAV321G R. pseudosolanacaerum mutants (P < 0.05, Student t-test).*
The PaeA proteins are widespread in Burkholderiales and Enterobacterales
To examine the distribution of PaeA proteins in bacterial genomes, we searched for homologs of the PaeA protein from R. pseudosolanacearum showing at least 30% identity over 80% of the protein length in the data set assembled by Smith et al. (55), consisting of 1,255 bacterial genomes, each representing a distinct genus and covering the whole bacterial diversity available in the NCBI database. We identified 340 PaeA homologs, which are widespread in α-, β-, and γ-proteobacteria of Pseudomonadota, as well as in Cyanobacteriota and to a lesser extent Bacteroidota (Fig. 8; Table S2). The closest homologs (>45% identity) were found in the orders Burkholderiales, Neisseriales, and Rhodocyclales among β-proteobacteria, and in Enterobacterales and Pasteurellales among γ-proteobacteria. Consistently, many of these bacteria have been reported to synthesize putrescine as their predominant polyamine (56–59). However, in most cases, their capacity to export putrescine has not been assessed.
Phylogenetic distribution of PaeA proteins in the domain Bacteria. R. pseudosolanacearum PaeA homologs were searched using BlastP among 1,255 proteome representatives from the domain bacteria described by Smith et al. (55). An unrooted maximum-likelihood phylogeny of the 1,255 bacteria (55) is shown. The rings represent the following, starting from the inner ring: (i) the phylum to which each strain belongs, limited to phyla in which most strains encode PaeA homologs; (ii) the class to which each strain belongs, limited to classes in which most strains encode PaeA homologs and are mentioned in the text; (iii) the class to which each strain belongs, limited to classes in which most strains encode PaeA homologs and are mentioned in the text; and (iv) the presence of PaeA homologs in the strains, represented by a green gradient according to the percentage of identity with the R. pseudosolanacearum PaeA protein. The complete list of strains encoding PaeA homologs is provided in Table S2.
PaeA homologs were detected in numerous species living in association with eukaryotic hosts, particularly within Enterobacterales and Burkholderiales, but close homologs were likewise found in environmental bacteria, such as the denitrifying bacteria of the Azoarcus, Thauera, and Oryzomicrobium genera (order Rhodocyclales). In Burkholderiales, PaeA proteins are highly conserved in both plant-associated and environmental species of Ralstonia, Cupriavidus, and Pandoraea, where they belong to the core genome of these three genera (Table S3). By contrast, they are absent from some closely related genera, including Burkholderia/Paraburkholderia, Caballeronia, and Mycetohabitans.
DISCUSSION
In this study, we identified in Ralstonia pseudosolanacearum GMI1000 a putrescine exporter, PaeA (RSc2277), whose inactivation enhances the symbiotic interaction between this bacterium and the legume Mimosa pudica.
Putrescine, along with spermine and spermidine, belongs to a class of small polycationic molecules known as polyamines, which are found across all three domains of life (60). Through their electrostatic interactions with various anionic macromolecules, polyamines play a crucial role in fundamental cellular processes, including protein synthesis by facilitating ribosomal subunit assembly and activity, DNA replication, chromosomal stability, and membrane and cell wall integrity (61–63). Beyond these essential physiological functions, polyamines have increasingly been recognized as key players in bacterial virulence. Their role in host-pathogen interactions is complex, as both hosts and microbes can synthesize polyamines and exploit them for their own benefit (64, 65). Hosts produce polyamines as part of their defense response, primarily through the generation of reactive oxygen species (ROS) from polyamine oxidation (66). Conversely, pathogens accumulate polyamines to counteract oxidative stress and enhance their survival within the host (67, 68). Additionally, pathogens utilize polyamines to regulate the expression of virulence factors. For example, putrescine enhances the production of plant cell wall-degrading enzymes and upregulates genes involved in chemotaxis and flagellar biogenesis in the plant pathogen Dickeya fangzhongdai (69). In other plant pathogens (Dickeya zeae) and human pathogens (Proteus mirabilis, Yersinia pestis), putrescine acts as an extracellular signal to modulate bacterial motility and biofilm formation, two functions important for host cell invasion (70–73). Interestingly, some pathogens also exploit host-derived polyamines for infection. For instance, Salmonella typhimurium and Pseudomonas aeruginosa uptake host-produced spermidine, which enables the expression and assembly of type III secretion systems (T3SS) essential for bacterial infection (74, 75).
In R. pseudosolanacearum, putrescine biosynthesis is essential, as a mutant lacking speC (RSc2365), which encodes an ornithine decarboxylase, is unable to grow in the absence of exogenous putrescine (46). Moreover, this bacterium excretes large amounts of putrescine in the xylem sap of host plants as well as in synthetic culture media, whereas only trace amounts of cadaverine and no detectable spermidine have been observed (46, 48). Additionally, R. pseudosolanacearum was shown to boost the synthesis of host putrescine during its interaction with tomato through the secretion of a type III transcription-activator-like effector (TALE) called Bgr11, which activates the expression of the tomato ADC gene encoding an arginine decarboxylase involved in putrescine biosynthesis. Effector-mediated activation of this gene leads to elevated agmatine and putrescine levels in tomato root and leaf tissues (76). The function of this host- and microbe-produced putrescine in planta remains unclear. Wu et al. (76) showed that the increased ADC activity on the plant side did not affect the growth of R. pseudosolanacearum in leaves.
PaeA homologs are widely distributed across Proteobacteria, particularly in Enterobacterales and Burkholderiales, which include both host-associated, such as plant and animal pathogens and symbionts, and free-living species. This broad distribution suggests that PaeA is not specifically linked to eukaryotic interactions. Polyamines, and putrescine in particular, are recognized as important metabolites produced by the gut microbiome, an environment that includes Enterobacterales (77, 78), but their functions remain unclear. In Cupriavidus and Ralstonia (Burkholderiales), PaeA homologs are present in the core genomes regardless of bacterial lifestyle, whether environmental, pathogenic, or symbiotic. Importantly, the presence of paeA does not predict the production of high amounts of putrescine. For instance, the Ralstonia solanacearum strain CMR15, unlike the GMI1000 strain, produces only modest amounts of putrescine in tomato xylem but accumulates substantial levels in culture (46), indicating that putrescine export is differentially regulated between strains. Similarly, although a close PaeA homolog is present in the Mimosa pudica symbiont Cupriavidus taiwanensis LMG19424, this bacterium does not produce detectable putrescine in culture (Fig. 6). Interestingly, Paraburkholderia phymatum STM815, a more ancient M. pudica symbiont (79), lacks paeA homolog, suggesting that this gene is not essential for symbiosis with M. pudica.
In a previous study, we experimentally evolved R. pseudosolanacearum into a M. pudica symbiont capable of forming nodules and infecting these nodules intracellularly. Nodulation and intracellular infection were mainly achieved through a combination of mutations that inactivated the regulators hrpG and efpR (35) or hrpG and components of the PhcA regulatory pathway (36). Here, we found that mutations in the putrescine exporter paeA occurred independently six times during this evolution experiment. Repeated mutations in the same gene are usually a strong indication that these mutations confer a fitness advantage to bacteria. We demonstrated that the V321G mutation in paeA completely abolished putrescine export in synthetic medium and significantly enhanced bacterial proliferation in nodules, resulting in an approximately five-fold increase in viable bacteria recovered per nodule. This increase in bacterial proliferation was observed in most of the genetic backgrounds tested (GMI1000 pRalta hrcV, hrpG, hrpG-efpR, and hrpG-phcQ), indicating that the infection phenotype does not depend on the presence of specific mutations other than those conferring nodulation, either hrcV or hrpG. Furthermore, the additive effect of the paeA mutation on nodule infection, when combined with another highly adaptive mutation for intracellular infection (such as efpR^E66K^ or phcQ^R154C^), allowed the detection of nitrogenase activity in nodules that would otherwise have been too low to be observed. No higher levels of nitrogenase activity were detected in nodules formed by evolved clones of this evolution experiment. Moreover, nif gene expression and nitrogenase activity in hrpG-efpR-paeA-induced nodules were only transiently detected at 10–15 dpi and then decreased to very low levels at 21 dpi. This transient phenotype likely reflects the limited intracellular persistence of evolved symbionts. Although the paeA mutation seems to slightly improve intracellular persistence, this trait is far from being optimized in experimentally evolved R. pseudosolanacearum symbionts compared to natural rhizobia. Even after 60 serial evolution cycles on M. pudica, R. pseudosolanacearum did not acquire the ability to survive for long in the cytoplasm of nodule cells, a trait found almost exclusively in rhizobia among plant-associated bacteria (10) and which may require specific functions (80).
One hypothesis to explain the improved adaptation of the paeA mutants to symbiosis with legumes is that the absence of polyamine secretion makes the strain intrinsically more fit. However, no growth differences were observed between the mutants and their respective parental strains in synthetic medium containing glutamine as carbon source, a medium in which R. pseudosolanacearum secretes large amounts of putrescine. An alternative hypothesis is that the putrescine produced by bacteria negatively interferes with the symbiotic process. Consistent with this idea, the natural symbiont C. taiwanensis also does not secrete putrescine in a synthetic medium containing glutamine as carbon source. Several similarities between natural and experimental evolutionary processes have already been highlighted in this experiment (81). This finding provides another striking example of the parallel between the two processes, demonstrating the relevance of experimental evolution approaches for understanding natural evolution (82).
Another interesting finding is that the expression profile of Mimosa genes gradually changes as bacteria progressively adapt to symbiosis (43). In nodules induced by the hrpG-efpR-paeA mutant, the expression of certain genes approaches the levels measured in nodules induced by the natural symbiont C. taiwanensis. Specifically, several defense-related genes are downregulated, reinforcing the idea that polyamines excreted by the bacterium are perceived negatively by the plant. In contrast, the expression of other plant genes essential for functional symbiosis, such as genes encoding leghemoglobins, is slightly increased in nodules induced by the hrpG-efpR-paeA mutant. Although the expression of these genes remains much lower than in C. taiwanensis-induced nodules, it indicates that bacterial evolution is progressing toward a more effective symbiosis. An intriguing observation is the progressive upregulation of the plant ADC gene as the bacteria adapt to the symbiosis. This gene encodes a key enzyme of the putrescine biosynthetic pathway in plants. Its upregulation is not due to the action of the type III TALE effector Bgr11 (76) in nodules since the T3SS of R. pseudosolancearum is inactivated in nodulating strains (32). Moreover, this upregulation is not specifically correlated with the presence of paeA mutated symbionts. An increase in its expression was also observed between hrpG-induced and hrpG-efpR-induced nodules, with further upregulation in nodules formed by the hrpG-efpR-paeA mutant and even more by C. taiwanensis symbionts. Instead, plant putrescine biosynthesis may be correlated with nodule development. Polyamine levels in legume nodules have been reported to be five to ten times higher than in other plant organs, although their composition varies considerably among legume species and appears to be an intrinsic characteristic of each species (83). The physiological role of putrescine in nodules is not entirely clear, but in Vigna nodules, polyamine levels, particularly putrescine, correlate linearly with both nitrogenase activity and leghemoglobin levels (84). Additionally, a study in Lotus japonicus suggested that polyamines primarily contribute to cell division and expansion during nodule development (85). Once again, the changes in Mimosa ADC gene expression in nodules induced by the hrpG-efpR-paeA mutant support the idea of an evolutionary trajectory toward better-developed nodules. These results also suggest that polyamines play a complex role in plant–microbe interactions and that bacterial- and plant-derived putrescine contribute differently to nodule symbiosis.
In conclusion, our study highlights the negative role of extracellular putrescine in the adaptation of Ralstonia pseudosolanacearum to symbiosis with Mimosa pudica. The inactivation of the putrescine exporter paeA contributes to a shift toward a more advanced symbiotic state. While the mutation does not confer full symbiotic competence comparable to natural rhizobia, it represents a step toward bacterial adaptation by modulating host responses and improving bacterial proliferation within nodules, giving the first signs of nitrogen fixation. Despite these advances, R. pseudosolanacearum remains limited in its ability to persist intracellularly and sustain nitrogen fixation over time. This highlights the challenges of evolving a non-rhizobial species into a mutualistic nitrogen-fixing symbiont and suggests that additional genetic changes are required. Future studies should focus on identifying the key rhizobial determinants that mediate this evolutionary transition.
MATERIALS AND METHODS
Bacterial strains and growth conditions
Bacterial strains used in this study are listed in Table S4. Ralstonia pseudosolanacearum-derived strains were grown at 28°C either on rich Phi medium containing 10 g·L^−1^ bacto-peptone, 1 g·L^−1^ yeast extract, and 1 g·L^−1^ casamino acids (86) or on synthetic medium (25 mM KH_2_PO_4_, 3.8 mM (NH_4_)2_SO_4, 0.203 mM MgSO_4_·7H_2_O, 40 µM Na_2_EDTA·2H_2_O, 15.6 µM ZnSO_4_·7H_2_O, 1.26 µM CoCl_2_·6H_2_O, 5 µM MnCl_2_·4H_2_O, 16.1 µM H_3_BO_3_, 1.6 µM Na_2_MoO_4_·2H_2_O, 10.8 µM FeSO_4_·7H_2_O, 1.2 µM CuSO_4_·5H_2_O) supplemented with 2% glycerol for natural transformation or 10 mM glutamine for putrescine quantification. The Cupriavidus taiwanensis strain was grown at 28°C on rich TY medium (tryptone 5 g·L^−1^, yeast extract 3 g·L^−1^). E. coli strains were grown in LB medium at 37°C. Antibiotics were used at the following concentrations: trimethoprim 100 µg·mL^−1^, spectinomycin 40 µg·mL^−1^, kanamycin 50 µg·mL^−1^ for R. pseudosolanacearum and 25 µg·mL^−1^ for E. coli, tetracycline 10 µg·mL^−1^, and gentamicin 10 µg·mL^−1^.
Plant material
Mimosa pudica seeds (LIPME production obtained from one commercial seed [B&T World Seed, Paguignan, France] of Australian origin) were sterilized as described (34). Then, seedlings were transferred in glass tubes (two seedlings per tube) in N-free conditions, containing a Fahraeus slant agar and liquid Jensen ¼ medium (87, 88). Plants were grown at 28°C in a growth chamber under the following conditions: 16 h light and 8 h dark with 70% humidity.
Experimental evolution
The evolution experiment was conducted as previously described (34, 89). Five lineages—two (B and F) derived from the CBM212 ancestor, two (G and K) derived from the CBM349 ancestor, and one (M) derived from the CBM356 ancestor, previously evolved for 35 cycles (34)—were further evolved until cycle 60 using 21-day cycles of nodulation. A new lineage (X) was derived from the reconstructed mutant GMI1000 pRalta hrpG^Q81*^ efpR^E66K^ and evolved for 15 nodulation cycles of 21 days each.
Bacterial proliferation in nodules
After 15 days post-inoculation, 5 to 10 nodules per plant from six different plants per strain analyzed were collected independently, sterilized in 2.4% hypochlorite solution for 15 min, rinsed three times with sterile H_2_O, and ground in 1 mL sterile H_2_O. Serial dilutions were plated on Phi medium containing trimethoprim and grown for 48 h at 28°C.
Acetylene reduction assays
At 15 or 21 days post-inoculation, three sets of 6 plants were collected for each inoculated strain. The number of nodules was counted, and the six plants were transferred together to 60 mL test tubes sealed with a Suba-seal septa. A syringe was used to remove 1 mL of air and replace it with 1 mL of acetylene. Plants were incubated for 4 h at 28°C in the light. The ethylene produced was measured by analyzing 0.4 mL of gas using an Agilent 7820A gas chromatograph system with GS-Alumina column (ref. 115-3552). The area of the ethylene peak was integrated and normalized by the number of nodules, the volume of gas analyzed, and the number of hours of incubation with acetylene.
Cytological analyses
Nodules were harvested at 10, 15, or 21 days post-inoculation. For LacZ staining, nodules were fixed in Z′ buffer (potassium phosphate buffer 0.1 M, KCl 10 mM, MgSO_4_ 1 mM, pH 7.4) with 2.5% glutaraldehyde and 0.1% triton for 1 h under vacuum. Nodules were embedded in 4% agarose and sectioned at 60 µm using a Leica VT1000S vibratome. Sections were incubated in a staining solution (potassium ferricyanide and ferrocyanide 5 mM each, 0.008% X-Gal in Z′ buffer for LacZ staining, and potassium ferricyanide and ferrocyanide 5 mM each; 1 mM X-Gluc in phosphate buffer pH 7 for GUS staining) for 1 h at 37°C. Sections were observed using an axioplan microscope (Zeiss) or a NanoZoomer scanner (Hamamatsu). The blue areas revealed by LacZ or GUS staining and brown areas corresponding to nodule invaded cells were measured in pixels using the Fiji software with the HSB color model. Statistical significance was analyzed using the pairwise Wilcoxon test. Live/dead staining of nodule sections was performed using Live/dead Baclight Bacterial Viability Kit (Invitrogen L7012). Sections were incubated 15 min at room temperature in the dark and observed by an inverted fluorescence microscope (Nikon Eclipse Ti).
Quantification of extracellular putrescine
Bacterial strains were grown in a synthetic medium containing 10 mM glutamine as sole carbon source at 28°C until an optical density at 600 nm (OD_600_) of 1 (~5 × 10^8^ CFU mL^−1^). Two milliliters of bacterial cultures was centrifuged for 5 min at maximum speed. Then, the supernatants were filtered using a syringe equipped with a 0.2 µm filter. The filtered supernatants were diluted at 1:10 with ultra-pure water and then diluted at 1:2 with isotopic dilution mass spectrometry (IDMS) internal standard. Finally, samples were transferred in HPLC vials and analyzed by liquid chromatography-mass spectrometry using an UHPLC vanquish (Thermo Fisher), with an HS F5 DISCOVERY 150 × 2.1 mm i.d., particle size 5 µm (Supelco) column with guard column SUPELGUARD KIT HS F5 5 µm 20 × 2.1 mm (Supelco) at 30°C. The mobile phases were water (A) and acetonitrile (B) both containing 0.1% of formic acid. The LC gradient program was as follows: 0–15 min, 0% B; 30 min, 35% B; 35–40 min, 40% B; and 40–45 min, 0% B. The injection volume was 5 µL. The HPLC was coupled to a detection by high-resolution ion mass spectrometry with an Orbitrap Exploris 120 Orbitrap (Thermo Fisher). Full scan HRMS analyses were performed in FTMS mode at a resolution of 60,000 (at 200 m/z), with the following source parameters: spray voltage, 3400 V; ion transfer tube temperature, 320°C; vaporization temperature, 75°C; sheath gas, 25 arb; and auxiliary gas, 5 arb. Data processing was performed by Skyline (version 24.1).
Other materials and methods concerning genome resequencing and detection of mutations in evolved clones, constructions of mutants and plasmids, phylogeny of evolved clones of the B lineage, visualization of NifH proteins by Western immunoblot, M. pudica gene expression analyses by quantitative reverse transcription-PCRs and the phylogenetic distribution of PaeA protein homologs are provided in Text S1 in the supplemental material.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Limpens E, Franken C, Smit P, Willemse J, Bisseling T, Geurts R. 2003. Lys M domain receptor kinases regulating rhizobial Nod factor-induced infection. Science 302:630–633. doi:10.1126/science.109007412947035 · doi ↗ · pubmed ↗
- 2Madsen EB, Madsen LH, Radutoiu S, Olbryt M, Rakwalska M, Szczyglowski K, Sato S, Kaneko T, Tabata S, Sandal N, Stougaard J. 2003. A receptor kinase gene of the Lys M type is involved in legumeperception of rhizobial signals. Nature 425:637–640. doi:10.1038/nature 0204514534591 · doi ↗ · pubmed ↗
- 3Radutoiu S, Madsen LH, Madsen EB, Felle HH, Umehara Y, Grønlund M, Sato S, Nakamura Y, Tabata S, Sandal N, Stougaard J. 2003. Plant recognition of symbiotic bacteria requires two Lys M receptor-like kinases. Nature 425:585–592. doi:10.1038/nature 0203914534578 · doi ↗ · pubmed ↗
- 4Arrighi J-F, Barre A, Ben Amor B, Bersoult A, Soriano LC, Mirabella R, de Carvalho-Niebel F, Journet E-P, Ghérardi M, Huguet T, Geurts R, Dénarié J, Rougé P, Gough C. 2006. The Medicago truncatula lysin [corrected] motif-receptor-like kinase gene family includes NFP and new nodule-expressed genes. Plant Physiol 142:265–279. doi:10.1104/pp.106.08465716844829 PMC 1557615 · doi ↗ · pubmed ↗
- 5Smit P, Limpens E, Geurts R, Fedorova E, Dolgikh E, Gough C, Bisseling T. 2007. Medicago LYK 3, an entry receptor in rhizobial nodulation factor signaling. Plant Physiol 145:183–191. doi:10.1104/pp.107.10049517586690 PMC 1976573 · doi ↗ · pubmed ↗
- 6Gage DJ. 2002. Analysis of infection thread development using Gfp- and Ds Red-expressing Sinorhizobium meliloti. J Bacteriol 184:7042–7046. doi:10.1128/JB.184.24.7042-7046.200212446653 PMC 135452 · doi ↗ · pubmed ↗
- 7Sprent JI. 2007. Evolving ideas of legume evolution and diversity: a taxonomic perspective on the occurrence of nodulation. New Phytol 174:11–25. doi:10.1111/j.1469-8137.2007.02015.x 17335493 · doi ↗ · pubmed ↗
- 8Montiel J, Reid D, Grønbæk TH, Benfeldt CM, James EK, Ott T, Ditengou FA, Nadzieja M, Kelly S, Stougaard J. 2021. Distinct signaling routes mediate intercellular and intracellular rhizobial infection in Lotus japonicus. Plant Physiol 185:1131–1147. doi:10.1093/plphys/kiaa 04933793909 PMC 8133683 · doi ↗ · pubmed ↗