Morphological and phylogenetic analyses reveal a new genus and two new species of Hymenochaetales (Basidiomycota) from southeast China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
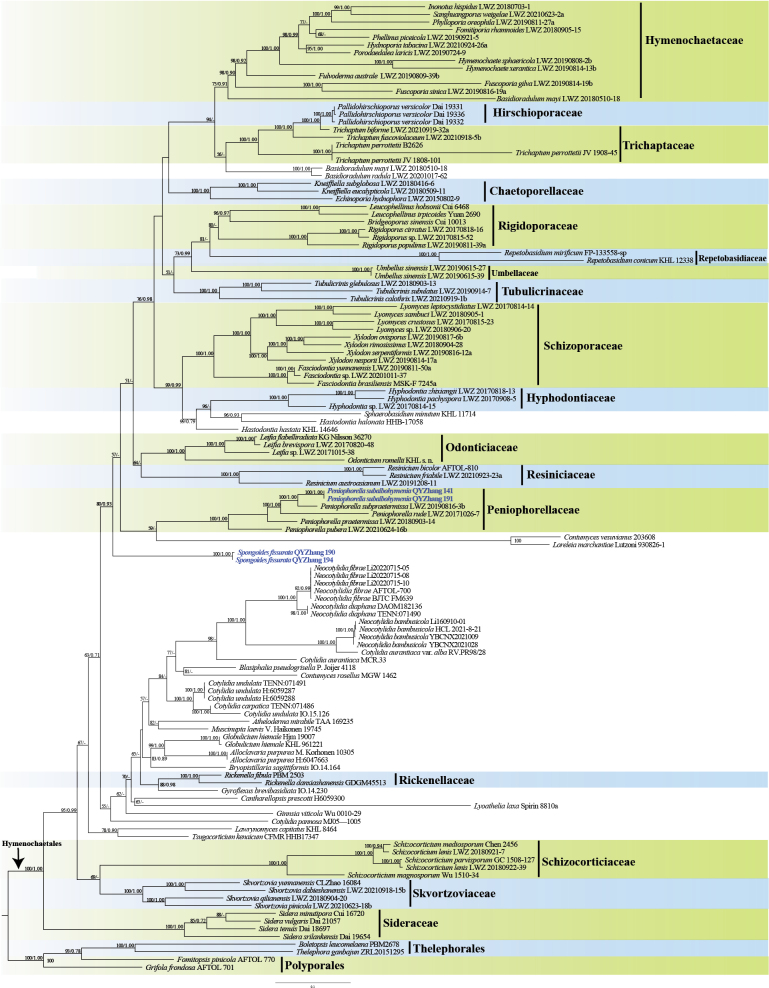 Figure 1
Figure 1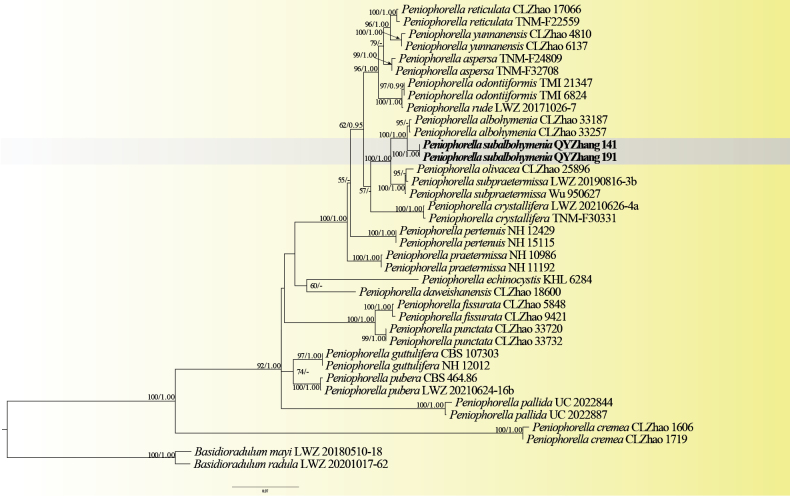 Figure 2
Figure 2 Figure 3
Figure 3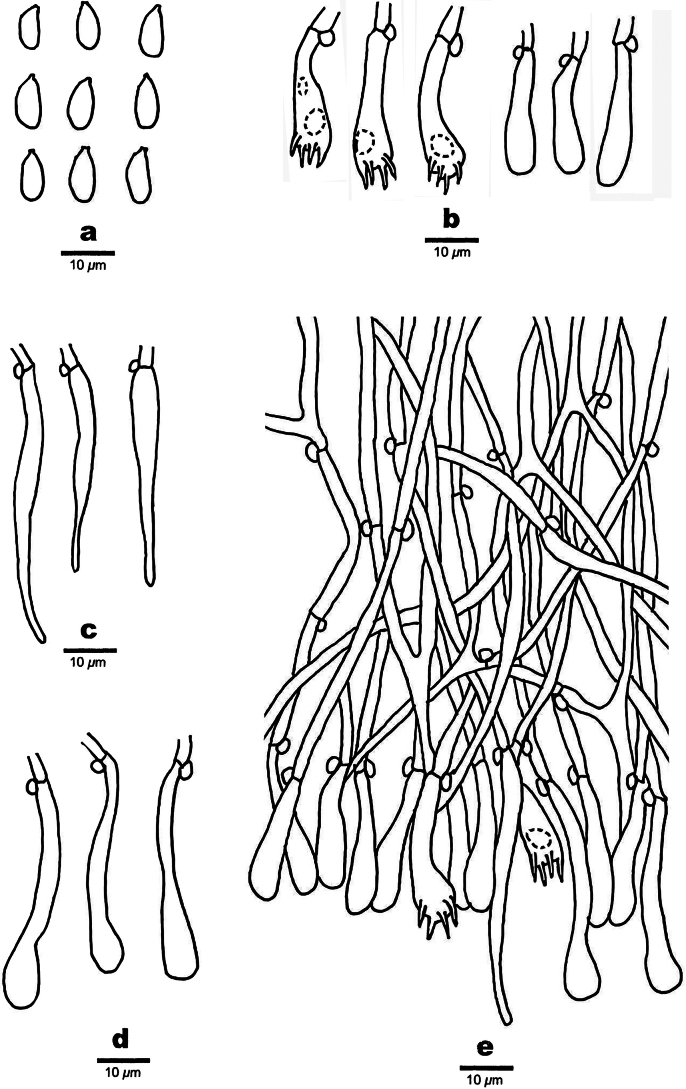 Figure 4
Figure 4 Figure 5
Figure 5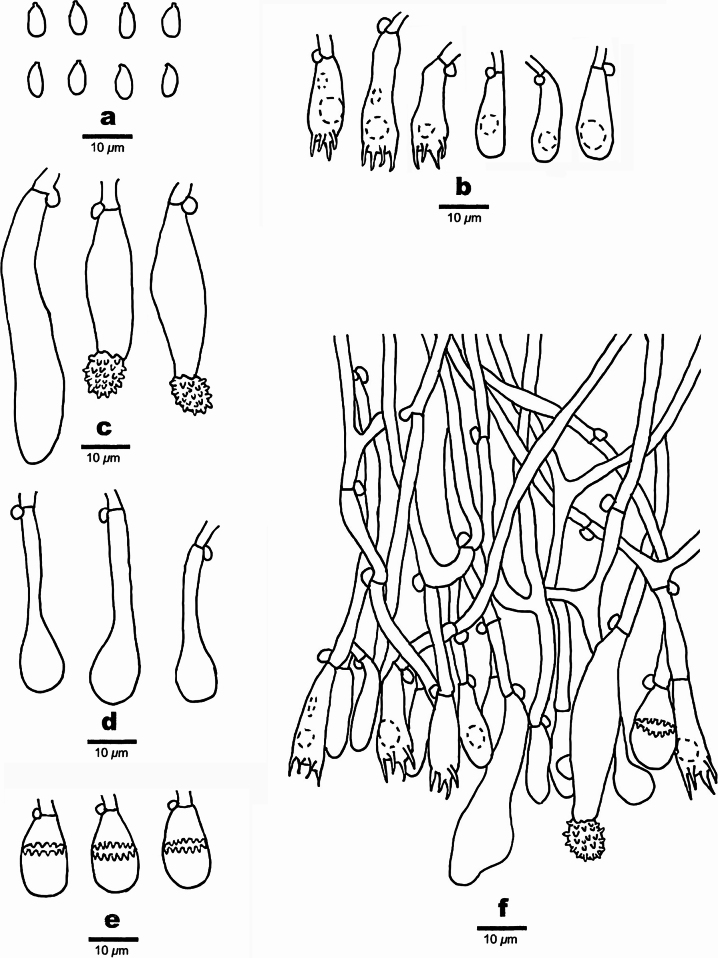 Figure 6
Figure 6| Order/Family | Species | Voucher | ITS | nrLSU |
|
|
|---|---|---|---|---|---|---|
|
| LWZ 20150802-9 |
|
|
|
| |
|
| LWZ 20180509-11 (Type) |
|
| — |
| |
|
| LWZ 20180416-6 |
|
| — |
| |
| — / |
| Dai 19331 |
|
|
| — |
|
| Dai 19332 |
|
|
|
| |
|
| Dai 19336 |
| — |
|
| |
| — / |
| LWZ 20180510-18 |
|
|
|
|
|
| LWZ 20201017-62 |
|
|
|
| |
|
| Cui 10321 |
|
|
|
| |
|
| LWZ 20180905-15 |
|
| — |
| |
|
| LWZ 20190809-39b |
|
|
|
| |
|
| LWZ 20190814-19b |
|
|
|
| |
|
| LWZ 20190816-19a |
|
|
|
| |
|
| LWZ 20210924-26a |
|
|
|
| |
|
| LWZ 20190808-2b |
|
|
|
| |
|
| LWZ 20190814-13b |
|
|
|
| |
|
| LWZ 20180703-1 |
|
|
|
| |
|
| LWZ 20190921-5 |
|
|
|
| |
|
| LWZ 20190811-27a |
|
|
|
| |
|
| LWZ 20190724-9 |
|
|
|
| |
|
| LWZ 20210623-2a |
|
|
|
| |
| — / |
| LWZ 20170908-5 |
|
| — |
|
|
| LWZ 20170818-13 |
|
| — |
| |
| LWZ 20170814-15 |
|
| — |
| ||
| — / |
| LWZ 20170820-48 |
|
|
|
|
|
| KG Nilsson 36270 |
|
| — | — | |
| LWZ 20171015-38 |
|
|
|
| ||
|
| KHL s. n. |
|
| — | — | |
| — / |
| CLZhao 33187 |
|
| — | — |
|
| CLZhao 33257 |
| — | — | — | |
|
| TNM |
|
| — | — | |
|
| TNM |
|
| — | — | |
|
| CLZhao 1606 |
| — | — | — | |
|
| CLZhao 1719 |
| — | — | — | |
|
| LWZ 20210626-4a |
|
| — | — | |
|
| TNM |
|
| — | — | |
|
| CLZhao 18600 |
|
| — | — | |
|
| KHL 6284 |
|
| — | — | |
|
| CLZhao 5848 |
|
| — | — | |
|
| CLZhao 9421 |
|
| — | — | |
|
| CBS 107303 |
|
| — | — | |
| — / |
| NH 12012 (GB) |
| — | — | — |
|
| TMI 21347 |
| — | — | — | |
|
| TMI 6824 |
| — | — | — | |
|
| CLZhao 25896 |
|
| — | — | |
|
| UC 2022844 |
| — | — | — | |
|
| UC 2022887 |
| — | — | — | |
|
| NH 12429 (GB) |
| — | — | — | |
|
| NH 15115 (GB) |
| — | — | — | |
|
| NH 10986 (GB) |
| — | — | — | |
|
| NH 11192 (GB) |
| — | — | — | |
|
| LWZ 20180903-14 |
|
|
|
| |
|
| CBS:464.86 |
|
| — | — | |
|
| LWZ 20210624-16b |
|
|
|
| |
|
| CLZhao 33720 |
|
| — | — | |
|
| CLZhao 33732 * |
|
| — | — | |
|
| CLZhao 17066 |
|
| — | — | |
|
| TNM |
|
| — | — | |
|
| LWZ 20171026-7 |
|
|
|
| |
|
| LWZ 20190816-3b |
|
|
|
| |
|
| Wu 950627 |
| — | — | — | |
|
|
|
|
| — | — | |
|
|
|
|
| — | — | |
|
| CLZhao 4810 |
|
| — | — | |
|
| CLZhao 6137 |
| — | — | — | |
| — / |
| KHL 12338 |
|
|
| — |
|
| FP-133558-sp | — |
|
|
| |
| — / |
| LWZ 20191208-11 |
|
|
|
|
|
| AFTOL-810 |
| — | — | — | |
|
| LWZ 20210923-23a |
|
|
|
| |
| — / |
| GDGM45513 |
| — |
|
|
|
| PBM 2503 (AFTOL 486) |
|
|
| — | |
| — / |
| Cui 10013 |
|
| — | — |
|
| Cui 6468 |
|
| — |
| |
|
| Yuan 2690 |
|
| — |
| |
|
| LWZ 20170818-16 |
|
|
|
| |
|
| LWZ 20190811-39a |
|
|
|
| |
| LWZ 20170815-52 |
|
|
|
| ||
| — / |
| LWZ 20180921-7 |
|
|
|
|
|
| LWZ 20180922-39 |
|
|
|
| |
|
| Wu 1510-34 |
|
| — | — | |
|
| Chen 2456 |
|
| — | — | |
|
| GC 1508-127 |
|
| — | — | |
| — / |
| MSK-F 7245a |
|
| — | — |
|
| LWZ 20190811-50a |
|
|
|
| |
| LWZ 20201011-37 |
|
|
|
| ||
|
| LWZ 20170815-23 |
|
| — |
| |
| — / |
| LWZ 20170814-14 |
|
| — |
|
|
| LWZ 20180905-1 |
|
|
|
| |
| LWZ 20180906-20 |
|
|
|
| ||
|
| LWZ 20190814-17a |
|
|
| — | |
|
| LWZ 20190817-6b |
|
|
|
| |
|
| LWZ 20180904-28 |
|
|
|
| |
|
| LWZ 20190816-12a |
|
|
|
| |
| — / |
| Cui 16720 |
|
|
|
|
|
| Dai 19654 |
|
|
|
| |
|
| Dai 18697 |
|
|
|
| |
|
| Dai 21057 |
|
|
|
| |
| — / |
| LWZ 20210918-15b |
|
|
|
|
|
| LWZ 20210623-18b |
|
|
|
| |
|
| LWZ 20180904-20 |
|
|
|
| |
|
| CLZhao 16084 |
|
| — |
| |
| — / |
| LWZ 20210919-1b |
|
|
|
|
|
| LWZ 20180903-13 |
|
|
| — | |
|
| LWZ 20190914-7 |
|
|
|
| |
| —/ |
| LWZ 20210919-32a |
|
|
|
|
|
| LWZ 20210918-5b |
|
|
|
| |
|
| JV 1808/ 101 |
|
|
|
| |
|
| JV 1908/ 45 |
|
|
|
| |
|
| B2626 |
|
|
| — | |
| —/ |
| LWZ 20190615-27 |
|
|
|
|
|
| LWZ 20190615-39 |
|
|
| — | |
| — / Incertae sedis |
| H:6047663 |
|
|
| — |
|
| M. Korhonen 10305 |
|
|
| — | |
|
| TAA 169235 |
|
| — | — | |
|
| P. Joijer 4118 |
|
|
| — | |
|
| IO.14.164 |
|
| — | — | |
|
| H6059300 |
|
|
| — | |
|
| MGW 1462 |
|
|
| — | |
|
| 203608 | — |
|
| — | |
|
| RV.PR98/28 | — |
| — | — | |
|
| MCR.33 | — |
| — | — | |
|
| TENN:071486 |
|
| — | — | |
|
| MJ05—1005 |
|
| — | — | |
|
| H:6059287 |
|
| — | — | |
|
| H:6059288 |
|
|
| — | |
|
| IO.15.126 |
|
| — | — | |
|
| TENN:071491 |
|
| — | — | |
|
|
|
|
|
|
| |
|
|
|
|
|
|
| |
|
| Wu 0010-29 |
|
| — | — | |
| — / Incertae sedis |
| Hjm 19007 |
|
| — | — |
|
| KHL 961221 |
|
| — | — | |
|
| IO.14.230 |
|
| — | — | |
|
| HHB-17058 |
|
| — | — | |
|
| KHL 14646 |
|
| — | — | |
|
| KHL 8464 |
|
| — | — | |
|
| Lutzoni 930826-1 |
|
| — | — | |
|
| Spirin 8810a |
|
| — | — | |
|
| V. Haikonen 19745 |
|
|
| — | |
|
| Li160910-01 (Type) |
|
| — | — | |
|
| HCL 2021-8-21 |
|
| — | — | |
|
| YBCNX2021009 |
|
| — | — | |
|
| YBCNX2021028 |
| — | — | — | |
|
| DAOM182136 | — |
| — | — | |
|
| TENN:071490 | — |
| — | — | |
|
| AFTOL-700 |
|
|
|
| |
|
| BJTC FM639 (Type) |
|
| — | — | |
|
| Li20220715-05 |
|
| — | — | |
|
| Li20220715-08 |
|
| — | — | |
|
| Li20220715-10 |
|
| — | — | |
|
| KHL 11714 |
|
| — | — | |
|
| CFMR HHB17347 | — |
|
|
| |
|
| AFTOL 770 |
|
|
|
| |
| — / |
| AFTOL 701 |
|
|
| — |
|
| PBM2678 |
|
|
| — | |
| — / |
| ZRL20151295 |
|
|
| — |
| Species name | Hymenial surface | Generative Hyphae | Cystidia | Basidia | Basidiospores | References |
|---|---|---|---|---|---|---|
|
| pellicular, smooth, margin whitish, fimbriate or with thin rhizomorphs | monomitic, clamped, thin-walled | cylindrical, basally widened, with obtuse to subcapitate apex | clavate, often constricted | narrowly ellipsoid, colorless, thin-walled, smooth, |
|
|
| Pellicular, membranaceous, smooth | Monomitic, simple-septate | present | clavate with stalked bases | ellipsoid, thin-walled, guttulate, |
|
|
| adnate, ceraceous, margin indeterminate | monomitic, clamps, thin-walled, richly branched | absent, paraphysoid hyphae present, some encrusted | clavate to cylindrical, constricted | globose, thin-walled, smooth, |
|
|
| resupinate, effused, adnate, thin, even, margin indeterminate, without rhizomorphs | monomitic, simple-septa, thin to thick-walled | hyphidia sometimes present | suburniform (utriform) to subcylindrical and constricted, more or less pedunculate | broadly ellipsoid to subglobose, slightly thickened walls and distinct apiculus, smooth, c, |
|
|
| resupinate, loosely attached, pellicular or membranous, smooth, with a thin, whitish subiculum and sparse or distinct hyphae | monomitic, clamped, thin-walled or thick-walled, more or less encrusted | capitata, moderately encrusted | thin-walled, relatively large | thin-walled or moderately thick-walled, smooth |
|
|
| resupinate, adnate, effused, very thin, smooth, margin indeterminate. | monomitic, clamped, thin-walled | leptocystidia present | subglobose to obconical | thin-walled, smooth, |
|
|
| resupinate, effused, closely adnate, thick, spongy, smooth, with wrinkles or cracks | monomitic, clamped, colorless, thin-walled | two kinds of cystidia, 1) tapering; 2) bottled, thin-walled | cylindrical | cylindrical or ellipsoid with an apiculus, colorless, thin-walled, smooth, | In present study |
|
| resupinate, effused, adnate, smooth, pilose by projecting cystidia | monomitic, simple-septa, distinct and with thin- to thickened walls | two kinds of cystidia, 1) subulate, lateral, smooth or encrusted, thick-walled; 2) gloeocystidia present or absent, | clavate or subcylindrical | globose to subglobose, with thin- to slightly thickened walls, |
|
|
| effuse, adnate, soft, smooth, with distinct, abrupt margin | monomitic, clamped, smooth, subhymenium thickening, with dendrohyphidia | present | suburniform, elongate | globose to subglobose, thin-walled, smooth, |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Lichen and fungal ecology · Plant Pathogens and Fungal Diseases
Introduction
Hymenochaetales Oberw. is one of the fungal orders mainly composed of wood-inhabiting macrofungi within the class Agaricomycetes, Basidiomycota, that play crucial ecological roles in forest ecosystems (Dai 2010; Dai et al. 2021; Wu et al. 2022; Yuan et al. 2023; Zhao et al. 2024; Dai et al. 2025). The order Hymenochaetales exhibits significant variations in morphological characteristics, primarily comprising hydnoid, corticioid, and poroid (Larsson et al. 2006; Dai 2010; Korotkin et al. 2018; Zhou et al. 2022; Zhou et al. 2023; Liu et al. 2025; Zhao et al. 2025). The majority of species in the order Hymenochaetales are wood-decaying fungi that cause white rot, as well as some important plant pathogens, symbiotic fungi, and medicinal species (Wu et al. 2022; Wang and Zhou 2023; Zhao et al. 2023, 2024). Consequently, Hymenochaetales fungi represent significant biological resources, holding importance for forest ecosystem function and economic development.
Currently, more than 1,400 species have been placed in Hymenochaetales, with ongoing discoveries highlighting substantial unexplored diversity, particularly in biodiverse regions such as East Asia (Zhou et al. 2018; Wu et al. 2022; Wang et al. 2023; Zhao et al. 2024). While the species diversity has been well explored all over the world, the systematics of Hymenochaetales at the family/genus level were poorly established. At the family level, the classification of Hymenochaetales has been subject to ongoing revisions, particularly over the past fifteen years, and a total of 14 family names have been successively applied. But at the genus level, about 19 genera have never been placed in any family in Hymenochaetales (Wu et al. 2022; Wang et al. 2023; Wang and Zhou 2023).
A notable component of this order is its considerable diversity of corticioid fungi, a polyphyletic morphological group (Liu et al. 2025; Zhao et al. 2025). Historically, many corticioid genera were classified based on morphology, leading to artificial groupings. The molecular phylogenetic studies have greatly clarified the systematics of these corticioid taxa (Wu et al. 2022; Yuan et al. 2023; Zhou et al. 2023). For instance, several newly proposed families, including Hyphodontiaceae, Peniophorellaceae L.W. Zhou et al., and Schizoporaceae Jülich, primarily encompass corticioid genera, highlighting the considerable phylogenetic diversification that has occurred within this morphological group (Wang et al. 2021, 2023). Therefore, integrated morphological and phylogenetic approaches remain crucial for accurately delineating taxa among these often cryptic fungi.
The family Peniophorellaceae was established within Hymenochaetales by Wang et al. (2023), to include the genus Peniophorella P. Karst, typed with P. pubera (Fr.) P. Karst. (Karst 1889), characterized by resupinate basidiomata with ceraceous to corneus consistency, smooth to tuberculate, and odontioid hymenophore with white to brownish-yellow hymenial surface, monomitic hyphae with generative hyphae bearing clamp connections (Guan et al. 2020). It is a cosmopolitan fungal genus with 39 species level names in Index Fungorum (http://www.indexfungorum.org, accessed on 1 July 2025). With the development of molecular systematics, the positioning of Peniophorella in Hymenochaetales has been further strengthened. Through molecular phylogeny of Hyphoderma Wallr., Larsson (2007) reinstated 19 new combinations within Peniophorella, which phylogenetically nested within the hymenochaetoid clade, confirming its placement outside traditional classifications. Subsequent studies by Miettinen and Larsson (2011) and Telleria et al. (2012) further resolved that Peniophorella species formed a distinct monophyletic group, clearly separated from Hyphoderma sensu stricto. Justo et al. (2017) revised the family-level classification of Peniophorella, which classified under Hyphodermataceae in the order Hymenochaetales, solidifying its taxonomic realignment. Recently, building on these phylogenetic frameworks, Guan et al. (2020) and Deng et al. (2025) described four new species of Peniophorella from southwestern China using integrated morphological and molecular analyses.
An examination of four corticioid specimens collected in Fujian, China, confirmed that all represent undescribed species within Hymenochaetales. To identify these two specimens at a species level and determine their taxonomic position at higher ranks, careful morphological examinations and phylogenetic analyses were performed. Ultimately, this study described a new Peniophorella species and a new genus within incertae sedis of Hymenochaetales.
Materials and methods
Morphological studies
The specimens were collected from Lai Zhou Forest Farm Experimental Center, Fujian Province in South China. The obtained specimens were stored in the Forest Pathology Laboratory at Nanjing Forestry University (NJFC), Nanjing, China. Macromorphological characteristics were investigated based on field notes and color photos of basidiomata. Color codes were verified as proposed by Petersen (1996). Microscopy studies were conducted as described by Wang and Zhou (2023) and Deng et al. (2025). Morphological observations of reproductive structures were conducted using the Zeiss fluorescence microscope. The Nikon Digital Sight DS-L3 camera was used to photograph microscopic structures. Drawings were made with the aid of a drawing tube. Microscopic features, measurements and drawings were made from slide preparations stained with Cotton Blue and Melzer’s Reagent. Basidiospores were measured from sections cut from the hymenophore. To present the variation of basidiospores size, 5% of measurements were excluded from each end of the range and are given in parentheses. The following abbreviations are used: IKI = Melzer’s Reagent, IKI− = neither amyloid nor dextrinoid, KOH = 5% potassium hydroxide, CB = Cotton Blue, CB− = acyanophilous, L = arithmetic average of all basidiospores length, W = arithmetic average of all basidiospores width, Q = variation in the L/W ratios between the specimens studied, (n = x/y) = the number of basidiospores (x) measured from a given number of specimens (y).
DNA extraction and sequencing
A cetyl trimethylammonium bromide (CTAB) rapid plant genome extraction kit (Aidlab Biotechnologies, Co., Ltd., Beijing, China) was used to extract DNA (Wu et al. 2020). The ITS region (nuclear ribosomal internal transcribed spacer, ITS), nuclear ribosomal large subunit (nrLSU), nuclear ribosomal small subunit (nrSSU), mitochondrial small subunit (mtSSU)) regions were amplified using the selected primer pairs: ITS5/ITS4 (White et al. 1990), LR0R/LR5 (Vilgalys and Hester 1990), NS1/NS41 (Hibbett 1996), MS1/MS2 (White et al. 1990), respectively. The PCR cycling schedule for ITS and mtSSU included an initial denaturation at 95 °C for 3 min or 94 °C for 2 min, followed by 35 cycles at 94 °C for 40 s, 54 °C for 45 s and 72 °C for 1 min, and a final extension of 72 °C for 10 min. The PCR cycling schedule for nrLSU and nrSSU included an initial denaturation at 94 °C for 1 min, followed by 35 cycles at 94 °C for 30 s, 50 °C (nrLSU), 53 °C (nrSSU) for 1 min and 72 °C for 1.5 min, and a final extension of 72 °C for 10 min (Wang et al. 2023; Zhou et al. 2023; Ma et al. 2025). All newly generated sequences in this study were deposited in GenBank and are listed in bold in Table 1.
Phylogenetic analyses
New sequences, deposited in GenBank (http://www.ncbi.nlm.nih.gov/genbank/) (Table 1), were aligned with additional sequences retrieved from GenBank (Table 1) using BioEdit 7.0.5.3 (Hall 1999) and MAFFT v.74 (http://mafft.cbrc.jp/alignment/server/, Katoh et al. 2017). Sequences of Hymenochaetales were adopted mainly from ITS + nrLSU + nrSSU + mtSSU + rpb2 + tef1α tree topologies established by Wang and Zhou (2023) and Ma et al. (2025). Sequences of Peniophorella were adopted mainly from ITS + nrLSU tree topologies established by Deng et al. (2025).
Maximum Likelihood (ML) and Bayesian Inference (BI) methods were used for the phylogenetic analyses. The ML analysis was carried out with RAxML version 8.2.12 (Stamatakis 2014), which statistical support values were obtained by using rapid bootstrapping with 1000 replicates, with default settings for other parameters and the best-fit models. The BI tree reconstruction was carried out with MrBayes v. 3.2.5 (Ronquist and Huelsenbeck 2003), which the best-fit partitioning scheme and substitution model were determined by using ModelFinder (Kalyaanamoorthy et al. 2017; Zhang et al. 2020) via the “greedy” algorithm, and branch lengths were estimated as “linked” and AICc. Four Markov chains were run for two runs from random starting trees for 10 million generations and trees were sampled every 1000 generations. The burn-in was set to discard 25% of the trees. A majority rule consensus tree of all remaining trees was calculated. Branches that received bootstrap support for Maximum Likelihood (ML) and Bayesian Posterior Probabilities (BPP) greater than or equal to 75% (ML) and 0.95 (BPP) were considered as significantly supported.
Results
Phylogeny
In this study, the combined ITS + nrLSU + nrSSU + mtSSU dataset (Fig. 1) included sequences from 118 specimens, representing 103 species of Hymenochaetales and Boletopsis leucomelaena (Pers.) Fayod and Thelephora ganbajun M. Zang from the order Thelephorales, and Fomitopsis pinicola (Sw.) P. Karst. and Grifola frondosa (Dicks.) Gray from order Polyporales as the outgroups. The dataset had an aligned length of 4,102 characters including gaps, consisting of 1,165 characters from ITS, 927 characters from nrLSU, 1,101 characters from nrSSU, and 909 characters from mtSSU (Table 1). ModelFinder suggested models were GTR + F + I + G4 for ITS, nrLSU and nrSSU, GTR + F + G4 for mtSSU, for the Bayesian analysis. The BI analysis resulted in a concordant topology with an average standard deviation of split frequencies of 0.009808. The ML and BI analyses resulted in nearly identical topologies and only the ML tree is presented with the bootstrap supports for ML and BPP not less than 50% and 0.70, respectively. In Fig. 1, our analysis recognizes 15 families and some genera have no definite position at the family level were recognized in the Hymenochaetales, confirming the results presented by Wang and Zhou (2023) and Ma et al. (2025). The two undescribed specimens (QYZhang 190 and QYZhang 194) formed a distinct, well-supported lineage, phylogenetically distant from known Hymenochaetales families and positioned within incertae sedis. This lineage close to Odonticiaceae L.W. Zhou & X.Wei Wang and Repetobasidiaceae Jülich, but it is not stable and has a low support values (54/-). In addition, two undescribed specimens (QYZhang 141 and QYZhang 191) consistently clustered within the Peniophorella clade, forming a highly supported lineage (100/1.00).
Maximum Likelihood (ML) tree illustrating the phylogeny of the order Hymenochaetales, based on a combined ITS + nrLSU + nrSSU + mtSSU dataset. Branches are labelled with parsimony bootstrap values (ML) higher than 50% and Bayesian Posterior Probabilities (BPPs) more than 0.70.
The combined ITS + nrLSU dataset (Fig. 2) included sequences from 38 specimens, representing 20 species of Peniophorella and two species of Basidioradulum mayi X.Wei Wang & L.W. Zhouand and B. radula (Fr.) Nobles as the outgroups. The dataset had an aligned length of 1,517 characters, including 655 characters from ITS and 862 characters from nrLSU (Table 1). For the Bayesian analysis, ModelFinder suggested GTR + F + I + G4 as the optimal substitution model for both ITS and nrLSU. The BI analysis resulted in a concordant topology with an average standard deviation of split frequencies of 0.003108. The ML and BI analyses resulted in nearly identical topologies and only the ML tree is presented with the bootstrap supports for ML and BPP not less than 50% and 0.70, respectively. In Fig. 2, the phylogram inferred from ITS + nrLSU sequences within Peniophorella (Fig. 2) highlighted two undescribed specimens (QYZhang 141 and QYZhang 191) formed an independent lineage with a robust support (100/1.00), and closely related to P. albohymenia Y.L. Deng & C.L. Zhao.
Maximum Likelihood (ML) tree illustrating the phylogeny of Peniophorella, based on a combined ITS + nrLSU dataset. Branches are labelled with parsimony bootstrap values (ML) higher than 50% and Bayesian Posterior Probabilities (BPPs) more than 0.70.
Taxonomy
Spongoides
Taxon classificationFungiHymenochaetalesRickenellaceae
Q.Y. Zhang gen. nov.
BD736A62-D064-50EA-A50C-9EBC1440D42F
861199
Etymology.
Spongoides (Lat.): derived from the Latin spongos (sponge) and the suffix-eides (resembling), referring to the spongy texture of the basidiomata.
Type species.
Spongoides fissurata Q.Y. Zhang, sp. nov.
Description.
Basidiomata annual, resupinate, effused, closely adnate, inseparable from substrate, thick, spongy. Hymenophore smooth, with wrinkles or cracks, white to cream; margin thinning out or abrupt, adnate. Hyphal system monomitic; generative hyphae clamped, colorless, thin walled, frequently branched, septate. Hymenium two kinds of cystidia, tapering or bottled, colorless, thin-walled. Basidia cylindrical, with a basal clamp connection and four sterigmata, filled with refractive oil-like matter. Basidiospores cylindrical or ellipsoid with an apiculus, colorless, thin-walled, smooth, IKI–, CB–, with oily contents.
Notes.
Building upon the well-established taxonomic framework of Hymenochaetales (Liu et al. 2024; Ma et al. 2025), our phylogeny confirms that Spongoides belongs to the order but is distinct from all known families and genera (Fig. 1). The family position of this genus needs to be further clarified. Spongoides seems to be morphologically related to members of Schizocorticiaceae L.W. Zhou & Xue W. Wang; however, Spongoides is distinct by the spongy basidiomata and the absence of tubular with obtuse apex leptocystidia. Given the phylogenetic distance and absence of key morphological characteristics possessed by the other genera in Hymenochaetales, we propose the establishment of Spongoides as a novel genus within incertae sedis of Hymenochaetales, with Spongoides fissurata as its generic type.
Spongoides
fissurata
Taxon classificationFungiHymenochaetalesRickenellaceae
Q.Y. Zhang sp. nov.
153D0601-0696-56DB-9D09-99F4BB708F3D
861200
Etymology.
fissurata (Lat.): Refers to the cracking hymenial surface of the type specimens.
Holotype.
China • Fujian Province, Nanping, Laizhou Town, Lai Zhou Forestry Experiment Station, on living Chamaecyparis formosensis, leg. Q.Y. Zhang, 20 June 2025, QYZhang 190 (NJFC).
Description.
Basidiomata annual, resupinate, effused, closely adnate, inseparable from substrate, thick, spongy when fresh and dry, without odor or taste when fresh, up to 8 cm long, 4 cm wide, 0.5 cm thick, and with extremely thin layer. Hymenial surface smooth, with wrinkles or cracks, white to light gray when fresh and dry. margin thinning out or abrupt, adnate. Hyphal system monomitic; generative hyphae with clamp connections, thin-walled, colorless, occasionally branched, 2.5–4 μm in diameter, IKI–, CB–, tissues unchanged in KOH. Cystidia of two types: (1) tapering cystidia, thin-walled, 30–49 × 2–4 μm; (2) bottled cystidia, with a relatively long neck, smooth, thin-walled, 23–35 × 3–7 μm. Basidia clavate, with four sterig-mata and a basal clamp connection, colorless, thin-walled, 16–25 × 6–8 μm; basidioles in shape similar to basidia, but slightly smaller. Basidiospores ellipsoid, colorless, thin-walled, IKI–, CB–, (6.5–)7–10(–11) × 3–6.4(–7) μm, L = 8.16 μm, W = 4.05 μm, Q = 1.96–2.05 (n = 60/2).
Basidiomata of Spongoides fissurata (holotype, QY Zhang 190). Scale bars: 1 cm (A); 2 mm (B).
Additional specimen (paratype) examined.
China • Fujian Province, Nanping, Laizhou Town, Lai Zhou Forestry Experiment Station, on living Chamaecyparis formosensis, leg. Q.Y. Zhang, 20 June 2025, QYZhang 194 (NJFC).
Microscopic structures of Spongoides fissurata (holotype, QY Zhang 190). a. Basidiospores; b. Basidia and basidioles; c. Tapering cystidia; d. Bottled cystidia; e. A section of hymenium.
Peniophorella
subalbohymenia
Taxon classificationFungiHymenochaetalesRickenellaceae
Q.Y. Zhang sp. nov.
798F176C-FCD5-5A39-A6AF-3C357912D715
861201
Holotype.
China • Fujian Province, Nanping, Laizhou Town, Lai Zhou Forestry Experiment Station, on fallen angiosperm branch, leg. Q.Y. Zhang, 20 June 2025, QY Zhang 141 (NJFC).
Etymology.
Subalbohymenia (Lat.): Refers to the morphological similarity and close phylogenetic relationship with P. albohymenia.
Diagnosis.
Peniophorella subalbohymenia is characterized by the membrana-ceous basidiomata with white hymenial surface, three types cystidia as stephanocyst, fusiform cystidia and lageniform cystidia, and ellipsoid basidiospores measuring 5–8 × 3.8–4.5 μm.
Basidiomata of Peniophorella subalbohymenia (holotype, QY Zhang 141). Scale bars: 1 cm (A); 2 mm (B).
Description.
Basidiomata annual, resupinate, adnate, membranaceous, with-out odor or taste when fresh, up to 12 cm long, 5 cm wide, and with extremely thin layer. Hymenial surface smooth, white when fresh and dry. Sterile margin distinctly, thin, white, up to 5 mm long. Hyphal system monomitic; generative hyphae with clamp connections, thin-walled, colorless, occasionally branched, 2.5–5 μm in diameter, IKI–, CB–, tissues unchanged in KOH. Cystidia of four types: (1) stephanocyst, thin-walled, with a ring-like wart protrusion on the upper-middle part, 13–17 × 9–10 μm; (2) fusiform cystidia, the apical part encrusted with asteroid, smooth, thin-walled, 25–55 × 6–15 μm; (3) lageniform cystidia, thin-walled, with a relatively long neck, 30–60 × 8–10 μm. Basidia clavate, with four sterig-mata and a basal clamp connection, colorless, thin-walled, 18–25 × 6.5–8 μm; basidioles in shape similar to basidia, but slightly smaller. Basidiospores ellipsoid, colorless, thin-walled, IKI–, CB–, (4.8–)5–8 × (3.5–)3.8–4.5(–4.8) μm, L = 6.14 μm, W = 4.12 μm, Q = 1.45–1.53 (n = 60/2).
Microscopic structures of Peniophorella subalbohymenia (holotype, QY Zhang 141). a. Basidiospores; b. Basidia and basidioles; c. Fusiform cystidia; d. Lageniform cystidia e. Stephanocyst; f. A section of hymenium.
Another specimen (paratype) examined.
China • Fujian Province, Nanping, Laizhou Town, Lai Zhou Forestry Experiment Station, on fallen angiosperm branch, leg. Q.Y. Zhang, 20 June 2025, QYZhang 191 (NJFC).
Discussion
The order Hymenochaetales represents a well-studied group of wood-inhabiting fungi within the Agaricomycetes (Basidiomycota), characterized by diverse basidiomata including poroid, hydnoid, corticioid, and agaricoid forms. Among these, corticioid fungi represent a significant and morphologically well-defined group within the order. Although the phylogenetic relationships of the order Hymenochaetales have been extensively studied, the results are different depending on the specimens and several gene sequences (Wang et al. 2021, 2023; Wu et al. 2022; Zhou et al. 2023).
At the family level, the classification of Hymenochaetales has been continually emended, especially in the past fifteen years. For example, Peniophorella was initially placed within the Rickenellaceae sensu lato based on morphological observation (Larsson 2007; He et al. 2019). Multilocus phylogenetic analyses have since shown that Peniophorella constitutes a distinct, well-supported lineage independent from other families in Hymenochaetales, leading to the establishment of the family Peniophorellaceae (Wang et al. 2023). Additionally, Wang et al. (2023) identified 14 families using seven genetic loci. In contrast, Zhou et al. (2023) accepted 11 families based on five genetic loci along with morphological evidence. Meanwhile, Zhao et al. (2025) proposed the recognition of 10 families and the rejection of 2 families based on the phylogenomics analysis. Furthermore, the delimitations of certain families are not fully resolved, and they are therefore regarded as incertae sedis, which encompass the following 19 genera, viz. Alloclavaria Dentinger & D.J. McLaughlin, Atheloderma Parmasto, Blasiphalia Redhead, Bryopistillaria Olariaga, Huhtinen, Læssøe, J.H. Petersen & K. Hansen, Cantharellopsis Kuype, Contumyces Redhead, Moncalvo, Vilgalys & Lutzoni, Cotylidia P. Karst., Ginnsia Sheng H. Wu & Hallenb., Globulicium Hjortstam, Gyroflexus Raithelh., Hastodontia (Parmasto) Hjortstam & Ryvarden, Kurtia Karasiński, Lawrynomyces Karasiński, Loreleia Redhead, Moncalvo, Vilgalys & Lutzoni, Lyoathelia Hjortstam & Ryvarden, Muscinupta Redhead, Lücking & Lawrey, Sphaerobasidium Oberw., Subulicium Hjortstam & Ryvarden, and Tsugacorticium Nakasone & Burds.
Phylogenetically, the phylogram based on the combined ITS + nrLSU + nrSSU + mtSSU sequences (Fig. 1) revealed that the new genus Spongoides (represented by its type species Spongoides fissurata) forms a distinct monophyletic lineage within Hymenochaetales, with uncertain familial placement (incertae sedis). Although this lineage appears phylogenetically close to Odonticiaceae and Repetobasidiaceae, the relationship remains unstable and is supported by low statistical support values. Morphologically, Odonticiaceae species differs from Spongoides by its grandinioid, odontioid to hydnoid basidiomata and cylindrical cystidia, while Spongoides has thick, spongy basidiomata and two kinds of cystidia, tapering or bottled; and Repetobasidiaceae species differs from Spongoides by its thin, ceraceous basidiomata and generative hyphae with clamp connections (Wang et al. 2023). Additionally, eight genera within the incertae sedis of Hymenochaetales exhibit resupinate, membranaceous or corticioid basidiomata with white to yellowish appearances, making them difficult to distinguish based on macromorphological characteristics. A morphological comparison between the new genus Spongoides and the other eight genera is presented in Table 2.
Furthermore, the corticioid fungi represents a highly polyphyletic status, they are not confined to the order Hymenochaetales but are prevalent in several other major orders of Agaricomycetes, such as Polyporales and Russulales. Similarly, some other genera, such as Hyphoderma Wallr (Polyporales) and Peniophora Cooke (Russulales) may share superficial morphological similarities with Spongoides. Hyphoderma niveomarginatum Y. Yang & C.L. Zhao is similar to Spongoides fissurata by its resupinate, white, cracking hymenial surface and basidiospores of similar size, but it differs from Spongoides fissurata by its ceraceous basidiomata and cystidia with contractions of varying degrees (Yang et al. 2023). Peniophora is an old corticioid genus comprising a large number of species. Similar to Spongoides fissurata, they have pale, resupinate basidiomata, and cause white rot. In contrast, a key distinguishing feature of Peniophora is the production of both encrusted cystidia and gloeocystidia (Xu et al. 2023).
In the present study, a new species, Peniophorella subalbohymenia is described based on phylogenetic and morphological characters. Phylogenetically, Peniophorella subalbohymenia is closely related to P. albohymenia. But morphologically P. subalbohymenia differs from P. albohymenia by having lageniform cystidia measuring 30–60 × 8–10 μm, and smaller basidiospores (5–8 × 3.8–4.5 μm vs. 9–10.9 × 4.5–5 μm, Deng et al. 2025). Also, there are 20 base pairs differences between Peniophorella subalbohymenia and P. albohymenia, which amounts to > 2% nucleotide differences in the ITS regions. Morphologically, P. praetermissa, P. yunnanensis C.L. Zhao, are similar to P. subalbohymenia based on the smooth hymenophore, allantoid basidiospores, and fusiform cystidia of the apical part encrusted with asteroid. However, P. praetermissa differs P. subalbohymenia in its longer basidiospores (8–11 μm in length vs. 5–8 μm in length), and the absence of large gloeocystidia measuring 30–120 × 8–18 μm (Hallenberg et al. 2007). Peniophorella yunnanensis differs P. subalbohymenia in smaller cystidia of the apical part encrusted with asteroid (9–28 × 3–8.5 μm vs. 25–55 × 6–15 μm) and the absence of stephanocyst (Guan et al. 2020).
Macrofungi, particularly wood-rotting fungi, constitute a vital component of forest ecosystems (Wu et al. 2022; Zhao et al. 2024). They produce a wide array of bioactive compounds and enzymes that break down organic matter, facilitating the decomposition of dead plant and animal tissues and enabling nutrient recycling (Wei and Dai 2004; Cui et al. 2018; Hyde 2022; Zhou et al. 2025). As such, they play an essential role in sustaining the biosphere, and play a core role in ecosystem processes and functioning. In recent years, with the rapid development of genome sequencing technologies at more affordable costs, the study of the phylogenetic relationships has attracted widespread attention due to its provision of higher species resolution. Studies on the taxonomy and phylogeny of wood-decomposing fungi have made significant progress. While numerous new species and genera within the order Hymenochaetales have been reported and described (Wu et al. 2022; Zhou et al. 2023; Liu et al. 2025), many novel taxa remain undiscovered, particularly in subtropical and tropical regions. The fungal species diversity in Hymenochaetales still has considerable potential for development. In addition, more undescribed Hymenochaetales records will be discovered throughout China after extensive collection combined with morphological and molecular analyses.
Supplementary Material
XML Treatment for Spongoides
XML Treatment for Spongoides fissurata
XML Treatment for Peniophorella subalbohymenia
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cui YY Cai Q Tang LP Liu JW Yang ZL (2018) The family Amanitaceae: Molecular phylogeny, higher-rank taxonomy and the species in China.Fungal Diversity 91: 5–230. 10.1007/s 13225-018-0405-9 · doi ↗
- 2Dai YC (2010) Hymenochaetaceae (Basidiomycota) in China.Fungal Diversity 45(1): 131–343. 10.1007/s 13225-010-0066-9 · doi ↗
- 3Dai YC Yang ZL Cui BK Wu G Yuan HS Zhou LW He SH Ge ZW Wu F Wei YL Yuan Y Si J (2021) Diversity and systematics of the important macrofungi in Chinese forests.Mycosystema 40: 770–805. 10.13346/j.mycosystema.210036 · doi ↗
- 4Dai YF Yuan Q Yang X Liu R Liu DF Yuan HS Zhao CL (2025) Morphological characteristics and phylogenetic analyses reveal five new species of Hymenochaetales (Agaricomycetes, Basidiomycota) from southwestern China.Myco Keys 114: 133–175. 10.3897/mycokeys.114.14385140051985 PMC 11883501 · doi ↗ · pubmed ↗
- 5Deng Y Chen M Liu L (2025) Morphological and molecular analyses revealed four new wood-inhabiting fungal species (Hymenochaetales, Basidiomycota) from Yunnan. Myco Keys 117: 29. 10.3897/mycokeys.117.146236 PMC 1205957940351353 · doi ↗ · pubmed ↗
- 6Eriksson J Ryvarden L (1973) The Corticiaceae of North Europe. 2. Fungifora, Oslo.
- 7Guan QX Zhao TJ Zhao CL (2020) Morphological characters and phylogenetic analyses reveal two new species of Peniophorella from southern China.Mycological Progress 19(4): 397–404. 10.1007/s 11557-020-01568-6 · doi ↗
- 8Hall TA (1999) Bio Edit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.Nucleic Acids Symposium Series 41(41): 95–98.