A homozygous synonymous NOP58 variant causes a neurodevelopmental disorder by impairing maturation of pre-ribosomal RNAs
Loisa D. Bonde, Tess Holling, Malik Alawi, Ahmed A. El Beheiry, Zabih Mir Hassani, François Bachand, Ibrahim M. Abdelrazek, Kerstin Kutsche

TL;DR
A genetic variant in NOP58 causes a severe neurodevelopmental disorder by disrupting ribosomal RNA processing in cells.
Contribution
NOP58 is identified as a novel disease gene for a neurodevelopmental disorder linked to ribosomal RNA maturation defects.
Findings
A homozygous NOP58 variant leads to reduced NOP58 and fibrillarin protein levels in patient fibroblasts.
The variant impairs pre-rRNA maturation and alters nucleolar morphology and snoRNA levels.
The NOP58 variant is classified as hypomorphic, contributing to a neurodevelopmental disorder.
Abstract
Ribosomes are ribonucleoproteins that are responsible for protein synthesis. They consist of ribosomal proteins and ribosomal RNAs (rRNAs). Pre-rRNAs are co-transcriptionally processed and chemically modified. The 2′-O-methylation of rRNAs is guided by box C/D small nucleolar ribonucleoprotein particles (snoRNPs), which are composed of a box C/D snoRNA and the core proteins NOP56, NOP58, SNU13, and the methyltransferase fibrillarin. Catalytically active box C/D snoRNPs function in nucleoli. We performed trio whole-exome sequencing in a proband with a severe neurodevelopmental disorder including global developmental delay, microcephaly, seizures, and ophthalmological and brain abnormalities and his healthy parents and identified the homozygous synonymous variant c.516G>A; p.Leu172= in NOP58. In fibroblasts of the proband, we demonstrated skipping of exon 7 in most NOP58 mRNAs, while ∼20%…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
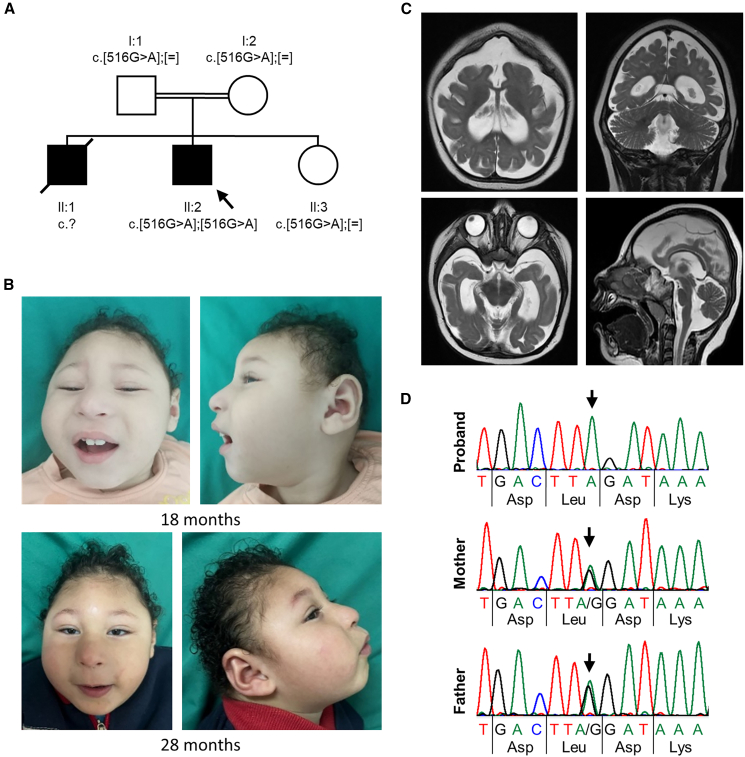 Figure 1
Figure 1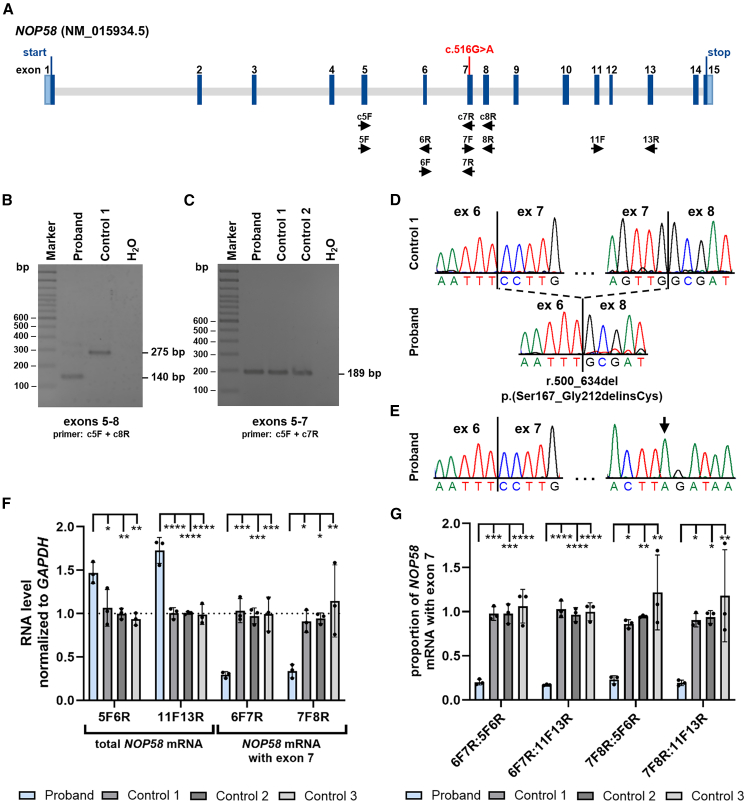 Figure 2
Figure 2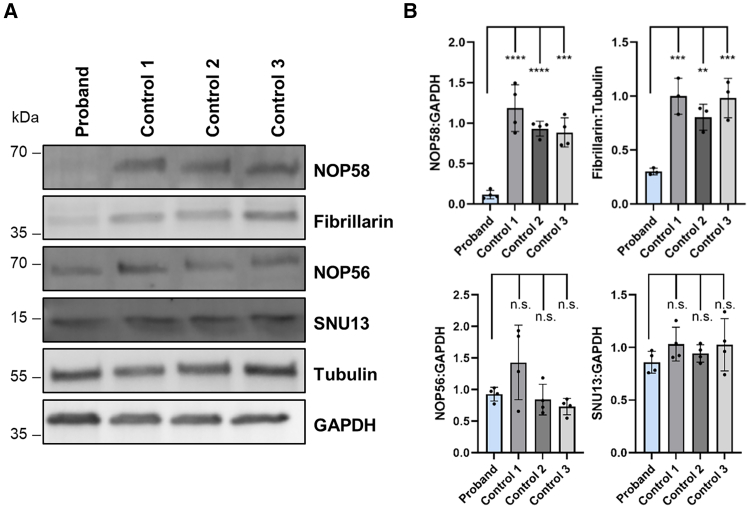 Figure 3
Figure 3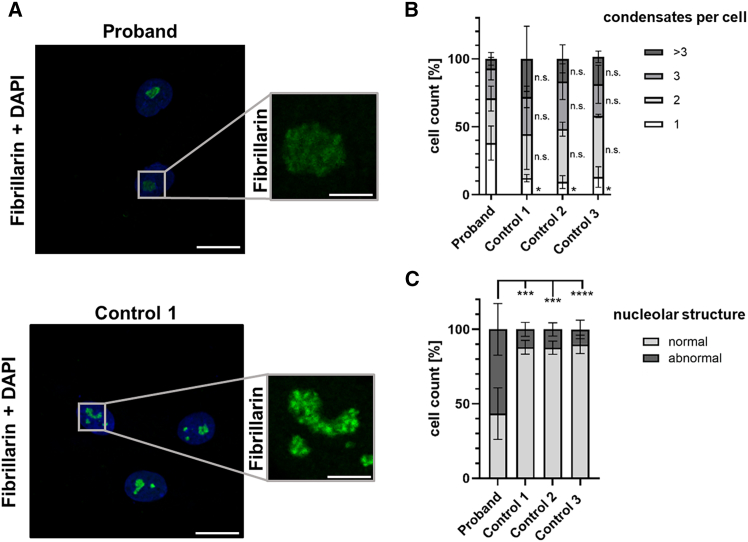 Figure 4
Figure 4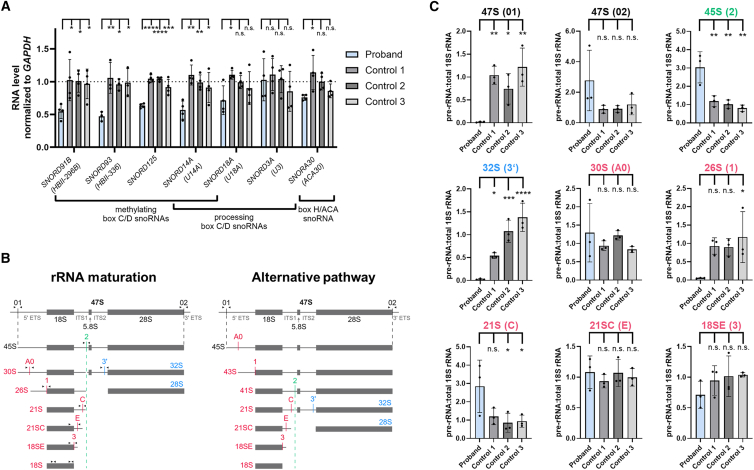 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA modifications and cancer · RNA and protein synthesis mechanisms · Cancer-related gene regulation
Main text
Ribosome biogenesis is an essential cellular process for embryonic development and cell survival.1 The assembly of small and large ribosomal subunits that are the heart of mRNA translation requires the regulated action of over 250 ribosome biogenesis factors, including proteins and non-coding RNAs.2^,^3 Ribosomal RNAs (rRNAs) are important for the catalytic activity of ribosomes. Pre-rRNA is transcribed by polymerase I and co-transcriptionally folded, chemically modified, and processed.3 rRNA modifications, such as 2′-O-ribose methylation and pseudouridylation, serve to stabilize the ribosome core and are important for ribosome heterogeneity and the fine-tuning of ribosome function.4^,^5 Small nucleolar RNAs (snoRNAs) are a highly abundant class of RNAs that function in the covalent modification and processing of pre-rRNAs in the nucleolus. Based on conserved sequence elements, snoRNAs are categorized into box H/ACA snoRNAs (SNORAs) and box C/D snoRNAs (SNORDs). For box H/ACA and box C/D snoRNAs, a specific set of core proteins associates with the snoRNA to form small nucleolar ribonucleoprotein particles (snoRNPs). The box H/ACA snoRNPs are required to isomerize uridine to pseudouridine in the rRNA, while the box C/D snoRNPs are responsible for transferring a methyl group to the 2′-hydroxyl of the ribose moiety of the rRNA. A subset of box H/ACA and C/D snoRNPs are required for pre-rRNA cleavage and folding events.6^,^7^,^8
The box C/D snoRNP forms by binding of a box C/D snoRNA to a set of core proteins including the methyltransferase fibrillarin, the related proteins NOP56 and NOP58, and SNU13.9^,^10^,^11 For the assembly of box C/D snoRNPs, several proteins are required including the HSP90/R2TP complex, C12orf45, NUFIP, ZNHIT3, and ZNHIT6. The synthesis of box C/D snoRNAs and the maturation of the box C/D snoRNP occurs in the nucleus. Pre-snoRNP particles are then transported to Cajal bodies where the final remodeling and processing occurs. Catalytically active box C/D snoRNPs are targeted to nucleoli to function in ribose methylation of rRNA.7 Base pairing between the snoRNA and the substrate RNA is required for fibrillarin-mediated 2′-O-methylation of rRNAs.12^,^13
Pathogenic variants in ZNHIT3 (MIM: 604500) and NOP56 (MIM: 614154) cause the progressive encephalopathy with edema, hypsarrhythmia, and optic atrophy (PEHO) syndrome (MIM: 260565) and spinocerebellar ataxia 36 (SCA36 [MIM: 614153]), respectively.14^,^15 A heterozygous GGCCTG repeat expansion in intron 1 of NOP56 underlies SCA36.15 Bi-allelic ZNHIT3 missense variants have been reported in subjects with PEHO syndrome, while a missense in trans with a frameshift variant has recently been identified in two fetuses with hydrops from one family, resulting in pregnancy loss.14^,^16^,^17 Functional studies in human cell lines and yeast have shown that ZNHIT3 pathogenic variants reduce steady-state levels of ZNHIT3 proteins and of box C/D snoRNAs, impair rRNA processing or decrease rRNA levels, change rRNA modification, and reduce cellular translation.14^,^17^,^18
For this study, the proband’s parents provided written informed consent for participation, clinical data and specimen collection, genetic analysis, and publication of relevant findings, including facial photographs and brain MRI scans, under a protocol approved by the Ethics Committee of the Hamburg Medical Chamber (PV7038-4438-BO-ff; Hamburg, Germany). Detailed methodologies are given in the supplemental information.
A 28-month-old male was born full-term as the second child of healthy consanguineous parents via Cesarean section following an uncomplicated pregnancy. His birth weight was 3.5 kg. Shortly after birth, he was admitted to the NICU for 7 days due to neonatal jaundice. He had a healthy younger sister and an older brother with a similar condition, who exhibited global developmental delay and severe microcephaly, with prenatal ultrasound showing alobar holoprosencephaly (Figure 1A). His brother passed away at the age of 7 months due to a respiratory infection. The proband developed focal to bilateral seizures at the age of 4 months.Figure 1. Pedigree, variant segregation, photographs, and brain MRI scans of the proband with the homozygous synonymous NOP58 c.516G>A variant(A) Pedigree of the family. The healthy father (I:1) and the healthy mother (I:2) are first-degree cousins and heterozygous carriers of the NM_015934.5:NOP58 c.516G>A; p.Leu172= variant. The proband (II:2, marked with an arrow) carries the NOP58 c.516G>A variant in the homozygous state. His similarly affected older brother (II:1) died at the age of 7 months, but could not be genetically tested due to lack of material. The proband has one healthy sister (II:3) who carries the NOP58 c.516G>A variant in the heterozygous state.(B) Facial photographs of the proband at the age of 18 months (top) and 28 months (bottom) show brachycephaly, prominent metopic ridge, receding anterior hairline, prominent antihelix, thin eyebrows, hypotelorism, narrow and upslanted palpebral fissures, epicanthus, strabismus, microphthalmia, microcornea, blue sclera, wide nasal ridge, long and deep philtrum, thin upper lip vermilion, and full cheeks.(C) Brain MRI scans of the proband at age 18 months. Axial and coronal T2-weighted images (top) show significant reduction in volume of the supratentorial brain with evidence of simplified gyral pattern, dilated bodies of the lateral ventricles tapering anteriorly, and slanted frontal convexity, more evident in the axial view. Axial T2-weighted image at lower level (bottom left) shows dilated temporal horns of the lateral ventricles along with thinning of the cisternal prechiasmatic segments of the optic nerves. Sagittal T2-weighted image (bottom right) shows a hypoplastic corpus callosum.(D) Partial sequence electropherograms showing the NOP58 c.516G>A variant in the homozygous state in the proband, and in the heterozygous state in the healthy parents (mother and father). Arrows point to the G-to-A change.
By the age of 28 months, the proband had global developmental delay, characterized by an inability to sit independently, limited vocalizations, and poor visual attention. At examination, he had severe microcephaly, brachycephaly, receding anterior hairline, prominent metopic ridge, thin eyebrows, hypotelorism, narrow and upslanted palpebral fissures, epicanthus, strabismus, blue sclera, wide nasal ridge, long and deep philtrum, thin upper lip vermilion, full cheeks, and prominent antihelix (Figure 1B). His growth parameters at the age of 28 months were as follows: an occipitofrontal head circumference of 37.5 cm (−6.39 z), along with a decreased length of 81.5 cm (−2.25 z) and weight of 11.5 kg (−0.92 z).
Brain imaging at age 18 months revealed a severe microcephalic configuration of the skull, accompanied by a marked reduction in the entire brain volume, particularly in the supratentorial region. The imaging also showed a simplified gyral pattern, and hypoplasia of the corpus callosum, optic nerve, and optic chiasm (Figure 1C). His fundoscopic examination identified bilateral microphthalmia, microcornea, optic disc pallor, and optic atrophy. Echocardiography, hearing assessment, and abdominal and pelvic ultrasound were unremarkable. His karyotype was normal (46,XY).
We performed trio whole-exome sequencing in the proband and healthy parents and did not detect any rare, likely pathogenic variant in a known disease gene. In the proband, we identified a homozygous synonymous variant in exon 7 of the candidate gene NOP58 (MIM: 616742), GenBank: NM_015934.5:c.516G>A; p.Leu172= (Figure 2A), which was confirmed by Sanger sequencing in the proband in the homozygous state and in his healthy sister and both parents in the heterozygous state (Figure 1D; Table S1). Material from the similarly affected older brother was not available for segregation analysis. The NOP58 variant is absent in the gnomAD database (v.4.1.0)19 and in the Regeneron Genetics Center Million Exome data (Table S2).20 With a loss-of-function observed/expected upper bound fraction of 0.995 and a Z score of 0.93, NOP58 is not intolerant to loss-of-function and missense variants, respectively (Database: gnomAD v.4.1.0).19 Splice site prediction programs did not predict any change for the detection of the canonical splice acceptor in intron 6 of NOP58 (Figure S1A; Table S2). Creation of a new splice site in exon 7 due to the G-to-A change was also not predicted. However, when we looked at exonic splicing enhancers using ESEFinder,21 we found that the binding site for the two splicing factors SRSF2 and SRSF5 was lost in exon 7 due to the c.516G>A change (Figure S1). We received a single match from GeneMatcher22 that was an individual with intellectual disability, autism, status epilepticus, hippocampal atrophy, and a de novo missense variant in NOP58. As there was only partial overlap between the phenotype of our proband and that of the subject from GeneMatcher, and the zygosity of the variants differed, we decided to investigate the functional consequences of the homozygous synonymous NOP58 variant. We obtained primary skin fibroblasts from the proband and performed NOP58 transcript analysis and various biochemical and cell biology experiments that collectively demonstrate clinical relevance of the bi-allelic NOP58 variant.Figure 2. The homozygous NOP58 c.516G>A variant leads to aberrant splicing of NOP58 pre-mRNA(A) Exon-intron structure of the NOP58 gene based on the mRNA reference sequence NM_015934.5, showing the c.516G>A variant located in exon 7. Exons are represented as boxes and introns as gray lines. Untranslated regions are depicted in light blue, while the coding region is shown in dark blue. Start and stop codons are indicated. The primers used for qualitative and quantitative RT-PCR experiments are shown below the exon-intron structure.(B and C) Agarose gels (2%) showing RT-PCR amplicons from fibroblast-derived cDNA of proband and control fibroblasts. (B) Using primers located in exons 5 (c5F) and 8 (c8R), the expected RT-PCR product of 275 bp was amplified in control 1. In contrast, a major amplicon of ∼140 bp was obtained from cDNA of proband-derived cells. (C) Using primers located in exons 5 (c5F) and 7 (c7R), the expected RT-PCR product of 189 bp was amplified from cDNA of control 1 and 2 and proband cells.(D) Direct sequencing of the RT-PCR amplicons obtained with primers located in exons 5 and 8 shown in (B). Partial sequence electropherograms show the canonically spliced NOP58 transcript with exon 6 spliced to exons 7 and 8 in control 1 (top) and aberrantly spliced NOP58 transcripts in the proband (bottom). Skipping of exon 7 (r.500_634del) is predicted to lead to an in-frame loss of 46 amino acid residues and insertion of a cysteine (p.Ser167_Gly212delinsCys) at the protein level.(E) Direct sequencing of the RT-PCR amplicons obtained with primers located in exons 5 and 7 shown in (C). The partial sequence electropherograms of the proband show canonically spliced NOP58 transcripts containing exon 7. The arrow points to the r.516G>A variant in exon 7.(F and G) Relative quantification of NOP58 mRNA levels by RT-qPCR using fibroblast-derived cDNA from cells of the proband and controls 1–3. A total of 300,000 primary fibroblasts from the proband and controls was seeded. The bars and errors show the mean ± SD of three independent experiments, each performed in triplicate. Individual data points are shown. One-way ANOVA followed by Dunnett’s post hoc test was used for statistical analysis. (F) For quantification of total NOP58 mRNA, primers located in exons 5 and 6 (F5R6) or exons 11 and 13 (11F13R) were used (left panel). To amplify only canonically spliced NOP58 mRNAs with exon 7, primers located in exons 6 and 7 (6F7R) or exons 7 and 8 (7F8R) were used (right panel). The amount of NOP58 mRNA relative to GAPDH mRNA is presented. (G) The proportion of canonically spliced NOP58 mRNAs with exon 7 is shown by calculating the ratio of NOP58 mRNA with exon 7 to total NOP58 mRNA. Ratios were calculated from relative NOP58 mRNA levels using the primer combinations shown in (F) and as indicated below the graph. ∗p ≤ 0.05, ∗∗p ≤ 0.01, ∗∗∗p ≤ 0.001, ∗∗∗∗p ≤ 0.0001. bp, base pairs; ex, exon; F, forward primer; R, reverse primer.
We investigated the effect of the NOP58 variant c.516G>A in exon 7 on NOP58 pre-mRNA splicing using RNA (cDNA) isolated from fibroblasts of the proband and controls. We used a forward primer in exon 5 (c5F) and a reverse primer in exon 8 (c8R) in RT-PCR experiments (Figure 2A; Table S1). In control 1 cells, we observed a strong RT-PCR band of the expected wild-type size (275 bp) in the agarose gel, while there was only a very faint 275-bp band in proband cells. In addition, a prominent smaller band of ∼140 bp was observed in proband cells (Figure 2B). Sequencing of the 275-bp RT-PCR product from control 1 revealed the reference sequence, while direct sequencing of the smaller RT-PCR amplicon from the proband identified aberrantly spliced NOP58 transcripts with exon 6 directly spliced to exon 8 (Figure 2D). Skipping of exon 7 causes loss of 135 nucleotides in NOP58 transcripts and is predicted to lead to loss of 46 amino acid residues and insertion of a cysteine at the protein level (NM_015934.5:r.500_634del; p.Ser167_Gly212delinsCys). To analyze whether proband cells still express NOP58 transcripts with exon 7, we performed RT-PCR using a forward primer in exon 5 (c5F) and a reverse primer in exon 7 (c7R) (Figure 2A). We obtained the expected RT-PCR product of 189 bp in control 1 and 2 and proband cells (Figure 2C). Sequencing of the amplicon identified canonically spliced NOP58 transcripts with the r.516G>A change in proband cells (Figure 2E). The data suggest that the homozygous NOP58 c.516G>A variant causes preferential skipping of exon 7 in NOP58 pre-mRNAs, while leaving some transcripts intact.
Next, we studied total NOP58 mRNA levels by RT-qPCR using two different primer combinations that are located outside the aberrantly spliced region. With both primer pairs (5F and 6R and 11F and 13R; Figure 2A), we detected total NOP58 mRNA levels that were 1.5- to 1.7-fold increased in proband compared with control cells (Figure 2F, left panel). To determine levels of canonically spliced NOP58 mRNAs in proband cells, we used two primer combinations, one with a forward primer in exon 6 (6F) and a reverse primer in exon 7 (7R) and another with a forward primer in exon 7 (7F) and a reverse primer in exon 8 (8R) (Figure 2A). For both primer combinations, we found that levels of NOP58 mRNAs with exon 7 were significantly decreased by ∼3.3-fold in proband compared with control cells (Figure 2F, right panel). We next wanted to determine the proportion of canonically spliced NOP58 mRNAs in proband cells. For this, we calculated the ratio of NOP58 mRNA levels with exon 7 to total NOP58 mRNA levels in proband and control cells, using values obtained from RT-qPCR experiments with four different primer pairs (see Figure 2F). As shown in Figure 2G, the proportion of NOP58 mRNAs with exon 7 was between 86% and 122% in control cells versus 17% and 23% in proband cells. Together, these data show that total NOP58 mRNA levels were significantly increased in proband cells, likely due to enhanced transcription compensating for potential NOP58 deficiency. Nonetheless, the proportion of correctly spliced NOP58 transcripts was drastically reduced in fibroblasts of the proband, potentially leading to the production of a small amount of NOP58 wild-type protein.
Studies in human cell lines and yeast showed that ZNHIT3 missense variants compromise the protein stability of ZNHIT3 and reduce the steady-state levels of ZNHIT3’s binding partner NUFIP1.14^,^17^,^18 We therefore assessed if the homozygous NOP58 variant impacts steady-state levels of NOP58 and of all other components of the C/D box snoRNP complex in proband and control cells. Fibroblasts of the proband showed statistically significant reductions in the amount of NOP58 and fibrillarin, to ∼12% and ∼30%, respectively, compared with control cells (Figures 3A and 3B). However, levels of NOP56 and SNU13 were similar in proband and control cells (Figures 3A and 3B). The data show that fibroblasts of the proband have a residual amount of NOP58, which likely represents wild-type protein produced from the canonically spliced NOP58 mRNAs. While the drastically decreased NOP58 amount concomitantly reduces steady-state levels of the methyltransferase fibrillarin in proband cells, levels of NOP56 and SNU13 do not seem to be affected.Figure 3. Levels of some components of the box C/D snoRNP complex are decreased in fibroblasts of the proband(A) Representative immunoblots of whole-cell lysates from proband and control fibroblasts. A total of 150,000 fibroblasts from the proband and controls was seeded. Equal amounts of whole-cell lysates were loaded. Endogenous NOP58, fibrillarin, NOP56, and SNU13 were monitored with the indicated antibodies. Anti-tubulin and anti-GAPDH antibodies were used to control for equal loading.(B) Quantification of protein levels from immunoblots shown in (A). Band intensities of fluorescence signals were quantified using the ChemiDoc imaging system. Levels of target proteins were normalized to tubulin or GAPDH. The bars and errors show the mean ± SD of three or four independent experiments. One-way ANOVA followed by Dunnett’s post hoc test was used for statistical analysis. ∗∗p ≤ 0.01, ∗∗∗p ≤ 0.001, ∗∗∗∗p ≤ 0.0001. kDa, kilodalton; n.s., not significant.
The function of the box C/D snoRNP complex is important for efficient assembly of ribosomes in the nucleolus, a multilayered biomolecular condensate.7^,^23 The nucleolar localization of the box C/D snoRNP requires all four core box C/D proteins.24 We therefore analyzed the nucleolar morphology of proband and control cells by staining of fibrillarin, a nucleoli marker, followed by immunofluorescence analysis and confocal microscopy. As shown in Figures 4A and S2, the distribution of the fibrillarin signal was different in cells of the proband compared with control cells, with nucleoli showing a textured structure and bright spots in the condensates of control cells, whereas a more uniform signal distribution was found in nucleoli of proband cells. We first counted the number of nucleolar condensates per cell and identified a statistically significantly higher proportion of cells with a single condensate in proband (∼38%) compared with control cells (9%–13%) (Figure 4B). In contrast, the percentage of cells with two and more condensates was similar in proband and control cells (Figure 4B). We next analyzed the nucleolar structure and categorized the cells as follows: (1) cells with a normal nucleolar structure, showing at least one very bright spot within the condensates and (2) cells with an abnormal nucleolar structure, showing evenly distributed fibrillarin signals throughout the condensates. We identified ∼57% of proband fibroblasts with an abnormal nucleolar structure compared with 10%–12% in control cells (Figure 4C). Together, the data indicate significant changes in nucleolar condensates and morphology in fibroblasts of the proband.Figure 4. Nucleolar morphology is altered in proband-derived fibroblasts(A) Immunofluorescence analysis using an anti-fibrillarin antibody followed by anti-mouse Alexa Fluor 488-conjugated secondary antibody (green) to stain nucleoli in proband and control fibroblasts. Nuclear DNA was stained with DAPI (blue). Fibroblasts were seeded on coverslips, cultivated under basal conditions, and imaged by confocal fluorescence microscopy. Representative image of proband (top) and control cells (bottom) is shown. Scale bars, 10 μm (left) and 5 μm (right).(B and C) Quantification of nucleolar features in fibroblasts of the proband and three controls. The bars and errors show the mean ± SD of three independent experiments. Samples were blinded for analyses. A minimum of 20 cells per cell line was analyzed in each experiment. Two-way ANOVA followed by Dunnett’s post hoc test was used for statistical analysis.(B) Cells were classified based on the number of nucleolar condensates per cell, ranging from one to seven. Cells with more than three condensates were grouped together.(C) Cells were categorized into two groups: (1) cells with normal nucleolar structure (i.e., cells with a textured fluorescence signal, showing at least one very bright dot within the condensate) and (2) cells with abnormal nucleolar structure (i.e., cells with evenly distributed fluorescence signals throughout the condensate). ∗p ≤ 0.05, ∗∗∗p ≤ 0.001, ∗∗∗∗p ≤ 0.0001; n.s., not significant.
Depletion of Nop58p, the NOP58 ortholog in yeast, causes a reduction of all five tested box C/D snoRNAs, while snoRNAs of the box H/ACA snoRNP complex were not affected.9 Similarly, ectopic expression of disease-associated ZNHIT3 variant proteins in HEK293T cells decreased the steady-state levels of some methylating box C/D snoRNAs, while levels of box C/D snoRNAs involved in rRNA processing were not changed.17 We next analyzed levels of six box C/D snoRNAs in proband and control fibroblasts. This included SNORD91B, SNORD93, and SNORD125 that are important for 2′-O-methylation of rRNAs and SNORD3A and SNORD18A required for pre-rRNA processing.17^,^25^,^26 The box C/D snoRNA SNORD14A has a dual role in production and 2′-O-methylation of rRNAs.27^,^28 Levels of SNORD14A, SNORD91B, SNORD93, and SNORD125 were significantly decreased to ∼57%, ∼55%, ∼47%, and ∼63%, respectively, in proband compared with all control fibroblasts (Figure 5A). For SNORD18A, we found a decrease to ∼72% in cells of the proband compared with controls that was only statistically significant compared with control 1 cells (Figure 5A). Levels of SNORD3A were similar in proband and control cells (Figure 5A). We also analyzed levels of SNORA30, a snoRNA of the box H/ACA snoRNP complex,30 and identified a decrease in the levels of this snoRNA to ∼76% in the proband that was only statistically significant when compared with control 1 cells (Figure 5A). Our data show that the steady-state levels of all tested box C/D snoRNAs guiding 2′-O-methylation and of SNORD14A, which is involved in both production and 2′-O-methylation of rRNAs, were consistently reduced in proband cells.Figure 5. Reduced levels of certain snoRNAs and impaired pre-rRNA processing in fibroblasts of the proband(A) Quantification of steady-state snoRNA levels in proband and control fibroblasts. A total of 300,000 primary fibroblasts from the proband and controls was seeded. For quantification, RNA levels were normalized to GAPDH levels. The bars and errors show the mean ± SD of three or four independent experiments, each performed in triplicate. Individual data points are shown. One-way ANOVA followed by Dunnett’s post hoc test was used for statistical analysis. The gene symbol and the canonical name (shown in brackets below the gene symbol) of the analyzed snoRNAs are displayed beneath the graph. The type and function of the snoRNAs are also indicated.(B) Pre-rRNA processing in human cells.29 The earliest species of rRNA is the 47S pre-rRNA, which is cleaved at sites 01 and 02 (in black) to generate the 45S precursor. At this point, the maturation process can take place in one of the two possible pathways. In one pathway (rRNA maturation, left panel), 45S pre-rRNA processing continues by cleavage at site 2 (in green) within ITS1, generating the 30S and 32S species. Once cleavage at site 2 takes place, the resulting 30S species is further trimmed at A0 and 1 (in red), generating the 21S rRNA. In an alternative rRNA maturation pathway (right panel), concomitant cleavage at site A0 and 1 in the 45S precursor may precede cleavage at site 2. This can result in the generation of 43S and 41S species. The 21S is then further trimmed at sites C and E (in red), generating the 18SE, which is exported to the cytoplasm where it is cleaved at site 3 (in red), generating the mature 18S rRNA. In parallel, cleavage at site 3ʹ (blue) is required for the production of mature 28S rRNA.(C) Relative quantification of pre-rRNA levels by RT-qPCR using fibroblast-derived cDNA from cells of the proband and controls 1–3. For each primer pair, the forward and reverse primers are positioned directly upstream and downstream of the respective cleavage site shown in (B) (arrowheads) and were designed to specifically amplify the corresponding rRNA precursor indicated above each panel (nomenclature refers to the left panel in (B) [rRNA maturation pathway]). The corresponding cleavage site is given in parentheses. For quantification, pre-rRNA levels were normalized to total 18S rRNA levels. The bars and errors show the mean ± SD of three independent experiments, each performed in triplicate. Individual data points are shown. One-way ANOVA followed by Dunnett’s post hoc test was used for statistical analysis. ∗p ≤ 0.05, ∗∗p ≤ 0.01, ∗∗∗p ≤ 0.001, ∗∗∗∗p ≤ 0.0001. 3′ETS, 3′ external transcribed spacer; 5′ETS, 5′ external transcribed spacer; ITS1, internal transcribed spacer 1; ITS2, internal transcribed spacer 2; n.s., not significant.
Reduced levels of certain box C/D snoRNAs, particularly SNORD14A, may suggest that pre-rRNA processing is altered in proband cells. To explore this possibility, we performed RT-qPCR using nine primer pairs that specifically target rRNA precursors and thereby monitor pre-rRNA processing in proband and control fibroblasts (Figure 5B). Similar levels of the 47S (primer pair spanning cleavage site 02), 30S, 21SC, and 18SE pre-rRNAs were observed in proband and control cells (Figure 5C). In contrast, the 45S and 21S rRNA precursors showed significantly increased levels in the proband cells (Figure 5C). Notably, the amplification signal for the 47S (primer pair spanning cleavage site 01), 32S, and 26S pre-rRNAs was substantially decreased in the proband compared with control cells (Figure 5C). This result was particularly unexpected, as the two primer pairs targeting different cleavage sites (01 and 02) of the same 47S pre-rRNA yielded contradictory results. Together, the increased levels of the 45S and 21S rRNA precursors in the proband’s fibroblasts support an impairment in pre-rRNA processing; yet, the near-complete failure to amplify an RT-PCR product with three primer pairs remains to be elucidated.
The 28-month-old male proband with the homozygous synonymous variant c.516G>A; p.Leu172= in NOP58 had a severe neurodevelopmental disorder, including severe global developmental delay, microcephaly, epilepsy, facial dysmorphism, microphthalmia, and other ophthalmological abnormalities. The proband had abnormal cerebral morphology, such as reduced brain volume, simplified gyral pattern, and hypoplastic corpus callosum. Our functional studies using proband-derived fibroblasts discovered skipping of exon 7 in most of the NOP58 mRNAs. However, ∼20% canonically spliced NOP58 mRNAs were identified in proband cells that likely account for the production of a small amount of normal NOP58 protein in the fibroblasts. The residual amount of NOP58 is likely needed for the biogenesis and function of the box C/D snoRNP and may be compatible with life, suggesting that the NOP58 variant c.516G>A; p.Leu172= is a hypomorphic rather than a complete loss-of-function allele. In line with this, deletion of the NOP58 ortholog in yeast (Nop58p) causes lethality, while its depletion impairs growth.9^,^31 The three known PEHO syndrome-associated ZNHIT3 missense variants p.Cys14Arg, p.Cys14Phe, and p.Ser31Leu allow embryonic development by destabilizing the ZNHIT3 protein leading to decreased ZNHIT3 steady-state levels in yeast and human cell culture.14^,^17^,^18 In contrast, a ZNHIT3 loss-of-function allele, such as the frameshift variant c.251_254del; p.Glu84Alafs∗8, in trans with the p.Cys14Arg variant cause hydrops fetalis followed by early pregnancy loss.17 Together, the data suggest that some residual amount of (less) functional box C/D snoRNPs is required for embryonic and cellular survival. Nonetheless, bi-allelic pathogenic variants in ZNHIT3 and NOP58 likely affecting the biogenesis of box C/D snoRNPs have severe consequences for neuronal and brain development in humans.
In fibroblasts of the proband with the homozygous NOP58 variant, levels of the two core box C/D snoRNP proteins SNU13 and NOP56 were similar to control cells, while those of the methyltransferase fibrillarin were significantly reduced. This is in contrast to normal fibrillarin (Nop1p) levels in a yeast strain depleted of Nop58p.9 The ZNHIT3-Cys14Phe variant protein concomitantly reduces endogenous NUFIP1 levels in HEK293T cells; however, steady-state levels of SNU13 and NOP58 are not affected.17 In addition, ZNHIT3-Cys14Phe and -Ser31Leu variant proteins form complexes with NUFIP1 similar to wild-type ZNHIT3.14^,^17 The data provide further evidence for an impaired, but not abolished, snoRNP biogenesis due to hypomorphic variants in ZNHIT3 and NOP58. Furthermore, the data suggest that different human cells, as well as yeast, have distinct compensatory mechanisms to respond to the depletion of key box C/D snoRNP components or assembly factors.
Our data show that the steady-state levels of some box C/D snoRNAs are significantly reduced in proband-derived fibroblasts with the homozygous NOP58 variant, whereas levels of other snoRNAs are only slightly affected, if at all. A consistent reduction was found for the three analyzed methylating box C/D snoRNAs SNORD91B, SNORD93, and SNORD125, while variable or no reduction was detected for the pre-rRNA processing box C/D snoRNAs and a box H/ACA snoRNA. Interestingly, the steady-state levels of the same three snoRNAs SNORD91B, SNORD93, and SNORD125 are also reduced in HEK293T cells expressing PEHO-associated ZNHIT3 variant proteins. Other box C/D snoRNAs involved in methylation and pre-rRNA processing, as well as orphan box C/D snoRNAs, are not changed in this cellular system.17 In yeast, Nop58p depletion and the introduction of the PEHO syndrome-causing ZNHIT3 variants C11F and S29L (corresponding to p.Cys14Phe and p.Ser31Leu in human) result in lower levels of both rRNA modifying and processing box C/D snoRNAs.9^,^18 The data suggest that some snoRNAs are more sensitive to the depletion of an important assembly or core protein of the box C/D snoRNP than others.
The consequences of the pathogenic ZNHIT3 missense variants on rRNA modification, rRNA processing, and cellular translation have been investigated in various cellular and model systems. In yeast mutants and lung tissue of an affected fetus, rRNA hypomethylation of specific 2′-O-methylation sites was detected, rather than a general decrease in 2′-O-methylation of rRNAs. The defect in box C/D snoRNP assembly caused by ZNHIT3 pathogenic variants results in a reduction of mature rRNA levels in a human cell line, rRNA processing impairments in yeast, and a decrease in global translation in both yeast and human cell culture.17^,^18 Nop58p depletion in yeast causes severe pre-rRNA processing defects.9 A first hint that rRNA homeostasis and/or the translational program may be impaired in proband-derived fibroblasts with the homozygous NOP58 variant is that the number and morphology of nucleolar condensates are altered compared with control fibroblasts. The nucleolus serves an important biological role as a site of ribonucleoprotein particle and early ribosome assembly.3^,^23 A significant increase in the nucleolar size and additional nucleolar dysfunctions have been reported in motor neurons of a spinal muscular atrophy mouse model that were associated with disturbances in snoRNP biogenesis and rRNA processing.32 Further evidence for an impaired pre-rRNA processing in the fibroblasts with the homozygous NOP58 variant was obtained from our RNA analyses, as the levels of the 45S and 21S rRNA precursors were significantly increased. An unexpected finding was the almost complete failure to amplify an RT-PCR product with three primer pairs using cDNA derived from proband but not from control fibroblasts, suggesting a specific effect in the proband cells. For cDNA synthesis, we used a retroviral reverse transcriptase, which is known to pause in the presence of secondary structure and/or modified nucleotides in non-coding RNAs, including rRNA. Modifications of rRNA can either permit reverse transcription, induce polymerase pausing, or completely block nucleotide incorporation. A block in polymerization is likely caused by an inability to form a base pair with any canonical nucleotide or by steric hindrance that impedes recognition of the modified residue by the reverse transcriptase.33^,^34 Based on the known limitations of retroviral reverse transcriptases, the inability to amplify RT-PCR products with some primer pairs suggests that pre-rRNAs in proband cells may be hypomethylated, accompanied by secondary structural stabilization, and/or show non-canonical modifications at specific sites that induce polymerase stalling and dissociation from the transcript. Interestingly, the fragile X mental retardation protein (FMRP), loss of which leads to fragile X syndrome in humans, interacts with snoRNAs and contributes to differential rRNA methylation.35 A trend of rRNA hypermethylation was observed in a human embryonic stem cell line deficient of FMRP,36 suggesting that alterations in the 2′-O-methylation pattern can involve both hypo- and hypermethylation, which collectively contribute to dysregulated protein synthesis and disease.36^,^37^,^38 Together, data from the literature and our functional data obtained from proband-derived fibroblasts suggest the existence of defects in pre-rRNA methylation and maturation in cells and tissues of the proband with the homozygous synonymous NOP58 variant.
PEHO syndrome, caused by bi-allelic ZNHIT3 variants, and the phenotype observed in the proband with the homozygous NOP58 variant share several clinical features, including microcephaly, global developmental delay, early-onset seizures, and optic atrophy.14 These findings suggest that defects in box C/D snoRNP biogenesis may particularly impact ribosome function and translation during neuronal development. Recent studies have highlighted the importance of 2′-O-methylation in developmental processes.4 In zebrafish, reduction or loss of rRNA methylation causes profound developmental defects, particularly of the head and brain, and leads to embryonic lethality.39 Studies in mouse, frog, and human identified differentially modified 2′-O-methylation sites in the rRNA during development and across tissues.4 Specifically, differential rRNA 2′-O-methylation defines early stages of development and rRNA 2′-O-methylation patterns differ between brain regions during mouse development and between human embryonic stem cells and neural stem cells.36^,^40 Taken together, the data show that dynamic rRNA modifications are functionally relevant during development, particularly in the brain.
In conclusion, we show that the homozygous NOP58 variant c.516G>A; p.Leu172= causes skipping of exon 7 in the majority of NOP58 pre-mRNAs, but leaves a small amount of transcripts intact. The hypomorphic nature of the NOP58 variant allows production of a residual amount of NOP58 wild-type protein. However, steady-state levels of fibrillarin and certain box C/D snoRNAs are significantly reduced in proband fibroblasts. Our data provide evidence of impaired pre-rRNA maturation and indirect evidence for an altered 2′-O-methylation pattern of rRNAs in the proband’s fibroblasts. These findings, together with changes in the morphology and condensate number of nucleoli in proband cells, suggest impairment of the box C/D snoRNP biogenesis that particularly affected brain and neuronal development. Both the NOP58*-*associated neurodevelopmental disorder and PEHO syndrome likely belong to the group of ribosome biogenesis disorders, also known as ribosomopathies, that includes a large spectrum of phenotypes, such as Diamond Blackfan anemia, Treacher-Collins syndrome, dyskeratosis congenita, and hydrops fetalis.41^,^42
Data and code availability
- •The published article includes all data generated or analyzed during this study.
- •Consent restrictions preclude sharing of full datasets, and the consents do not cover the deposition of the exome sequencing data in a public database. NOP58 variant and phenotypic information were submitted to the LOVD database (https://databases.lovd.nl/shared/genes/NOP58), with the LOVD Variant ID: 0001045374 and Phenotype ID: 0000351322.
Acknowledgments
We are grateful to the proband’s parents who agreed to participate in this project. We thank Sina Ramcke for skillful technical assistance and the UKE Microscopy Imaging Facility (UMIF) at the University Medical Center Hamburg-Eppendorf for technical support. This work was supported by the Deutsche Forschungsgemeinschaft (KU 1240/17-1 to K.K.) and the “Close the Gap” project from the Gender Equality Unit of the University Medical Center Hamburg-Eppendorf.
Declaration of interests
The authors declare no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Teixeira F.K.Lehmann R.Translational Control during Developmental Transitions Cold Spring Harb. Perspect. Biol.112019 a 03298710.1101/cshperspect.a 032987 PMC 654604330082467 · doi ↗ · pubmed ↗
- 2Dorner K.Ruggeri C.Zemp I.Kutay U.Ribosome biogenesis factors-from names to functions EMBO J.422023 e 11269910.15252/embj.2022112699 PMC 1006833736762427 · doi ↗ · pubmed ↗
- 3Vanden Broeck A.Klinge S.Eukaryotic Ribosome Assembly Annu. Rev. Biochem.93202418921010.1146/annurev-biochem-030222-11361138768392 · doi ↗ · pubmed ↗
- 4Milenkovic I.Novoa E.M.Dynamic r RNA modifications as a source of ribosome heterogeneity Trends Cell Biol.35202560461410.1016/j.tcb.2024.10.00139500673 · doi ↗ · pubmed ↗
- 5Sloan K.E.Warda A.S.Sharma S.Entian K.D.Lafontaine D.L.J.Bohnsack M.T.Tuning the ribosome: The influence of r RNA modification on eukaryotic ribosome biogenesis and function RNA Biol.1420171138115210.1080/15476286.2016.125978127911188 PMC 5699541 · doi ↗ · pubmed ↗
- 6Falaleeva M.Welden J.R.Duncan M.J.Stamm S.C/D-box sno RN As form methylating and non-methylating ribonucleoprotein complexes: Old dogs show new tricks Bioessays 392017160026410.1002/bies.201600264 PMC 558653828505386 · doi ↗ · pubmed ↗
- 7Massenet S.Bertrand E.Verheggen C.Assembly and trafficking of box C/D and H/ACA sno RN Ps RNA Biol.14201768069210.1080/15476286.2016.124364627715451 PMC 5519232 · doi ↗ · pubmed ↗
- 8Webster S.F.Ghalei H.Maturation of small nucleolar RN As: from production to function RNA Biol.20202371573610.1080/15476286.2023.225454037796118 PMC 10557570 · doi ↗ · pubmed ↗