In vitro and in vivo studies on the activity and selectivity of butoconazole in experimental infection by Trypanosoma cruzi
Gabriela Rodrigues Leite, Denise da Gama Jaen Batista, Marcos Meuser Batista, Krislayne Nunes da Costa, Tomás Mac Loughlin, Emilia M Barrionuevo, Alan Talevi, Lucas N Aberca, Otacilio C Moreira, Amanda Faier-Pereira, Beatriz Iandra da Silva Ferreira

TL;DR
This study explores butoconazole as a potential treatment for Chagas disease, finding it effective in lab settings and when combined with another drug.
Contribution
The study identifies butoconazole as a potent and low-toxic compound against Trypanosoma cruzi, with insights into combination therapy efficacy.
Findings
Butoconazole showed high potency against T. cruzi amastigotes with low cytotoxicity.
Combining butoconazole with benznidazole showed additive effects in reducing parasitaemia and improving survival in mice.
Despite combination therapy benefits, no parasitological cure was achieved as parasite DNA remained in the heart tissue.
Abstract
The protozoan Trypanosoma cruzi causes Chagas disease (CD). There are two drugs available for the treatment with limited efficacy, especially in the later stage. Focusing on drug repurposing by virtual screening of chemical databases, butoconazole (BTZ) was identified as promising hit. Our aim was to explore the trypanosomicidal effect of BTZ alone or in combination with benznidazole (BZ) against T. cruzi. Our in vitro assays validated the low cytotoxicity of BTZ and high potency on amastigotes (EC50 = 0.07 μM), being 24-fold more potent than BZ. Washout assays demonstrated the sterilisation capacity of BTZ, whereas its combination with BZ gave an additive interaction (xƩFICI = 0.66). In a mouse model of acute T. cruzi infection, BTZ was unable to suppress parasitaemia but ensured the animal survival. BTZ plus BZ reduced parasitaemia and provided higher survival rates than…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1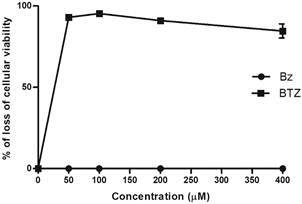 Figure 2
Figure 2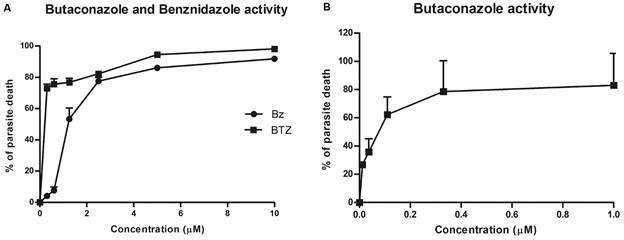 Figure 3
Figure 3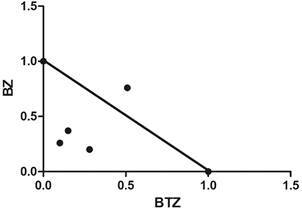 Figure 4
Figure 4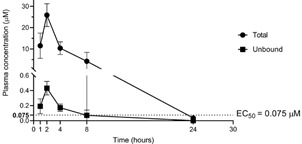 Figure 5
Figure 5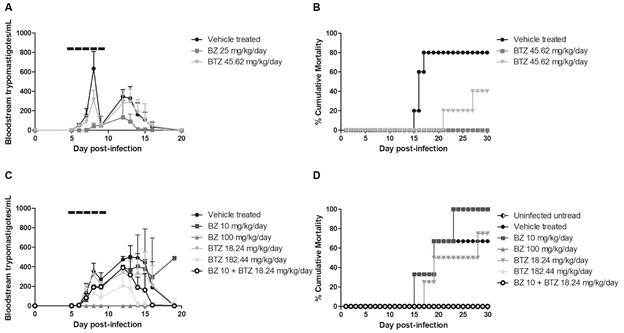 Figure 6
Figure 6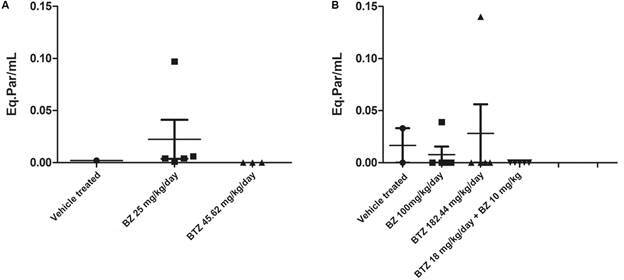 Figure 7
Figure 7- —CNPq
- —FIOCRUZ
- —FAPERJ
- —Agencia I+D+I
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTrypanosoma species research and implications · Research on Leishmaniasis Studies
American trypanosomiasis, also called Chagas disease (CD), is caused by the haemoflagellate protozoan Trypanosoma cruzi transmitted through infected triatomine vector insects to several species of mammals.1 It is considered endemic in 21 Latin American countries, especially in regions with lower human development index, but the identification of CD carriers in non-endemic countries is a concern due to the globalisation and increased migratory flow.2 According to the World Health Organisation (WHO), more than six million people are infected, with about 30,000 new cases and 12,000 deaths annually due to the development and complications of the disease, representing a relevant global public health problem.1 ^,^ 2
CD has an acute phase of short duration (four-eight weeks), usually asymptomatic or oligosymptomatic and with patent parasitaemia. Although parasite proliferation is controlled by a competent host immune response, there is no cure and parasites remain hidden in tissues and organs for years or decades, without developing clinical symptoms. However, this asymptomatic chronic stage may evolve to the symptomatic clinical form, usually cardiac and/or digestive, in about 30-40% of the carriers, which can be debilitating and fatal.3
It is well reported that early diagnosis is crucial to increasing the chances of cure, during T. cruzi infection. However, most of those affected people only become aware of their diagnosis in the late phase of CD.4 ^,^ 5
In addition, the therapeutic arsenal against CD is unsatisfactory and restricted to the use of two old nitroderivatives: benznidazole (BZ) and nifurtimox (NF). Both require an extended period of administration, display limited efficacy in chronic stages, induce severe side effects, and have limited efficacy against strains of T. cruzi that are naturally resistant to these nitroderivatives, factors that favour low adherence and/or discontinuity of the therapy.6 Based on intensive research on transmission, treatment, and control, the Pan American Health Organisation (PAHO) estimates that among the people affected by CD living in the Americas, only around < 10% can access diagnosis, and of those, only about 1% have access to appropriate care and treatment.7 ^,^ 8
Drug repurposing is a promising approach that may combine bioinformatics tools with the availability of drug libraries/chemical databases comprising marketed, discontinued and/or archived drugs, to find a new therapeutic indication, different from the original one.9 Compared to the process of discovering and developing a de novo drug, repurposing has several advantages such as significantly reducing the time and investment costs required in research, bypassing the main barriers that make new advances aimed at neglected tropical diseases, such as CD, unfeasible.10 ^,^ 11
Combined therapy has gained more prominence in screenings with successful results in clinical treatment for various diseases.12 The co-administration of drugs with different mechanisms of action modulates more than one target simultaneously, increasing the chances of improved efficacy with lower doses and reduced time, decreasing adverse effects and the chances of drug resistance.12 ^,^ 13 ^,^ 14
The 14α-demethylase (CYP51) inhibitors, indicated to treat fungal infections, have been investigated in preclinical and clinical assays for CD.15 They act by inhibiting sterol biosynthesis, which is considered as an essential metabolic pathway for membrane remodelling, metabolism, and cell division process of T. cruzi.12 Due to the promising findings in mouse and dog models of T. cruzi experimental infection, two azole antifungals advanced into clinical studies in CD chronic carriers.16 ^,^ 17 ^,^ 18 The CHAGASAZOLE trial aimed to evaluate the efficacy of posaconazole (POSA) in chronic patients with CD. Although the treatment achieved parasitaemia suppression, its efficacy was not sustained after the drug withdrawal.17 ^,^ 18 STOP-CHAGAS trial (Study of Oral Posaconazole in the Treatment of Chagas Disease) also evaluated POSA in monotherapy regimens and in combination with BZ in asymptomatic chronic patients with CD. The antiparasitic effect of the POSA + BZ combination showed no advantage over BZ monotherapy.18
Fosravuconazole (E1224) is a prodrug of ravuconazole (RAV), a CYP51 inhibitor with promising anti-T. cruzi results in vitro and in vivo experimental models.12 However, E1224 also failed to sustain negative quantitative polymerase chain reaction (qPCR) in a randomised clinical trial with different oral dosing regimens administered to chronic patients.16 The lack of translation of E1224 and POSA has been attributed to many factors including underdosing and the short treatment period.6
On the other hand, the well-known trypanosomicidal activity of CYP51 inhibitors and the metabolic differences on trypanosomatidae and fungi justify the search for novel drugs more selective for the parasite enzymes, as has been reported in a in vivo mouse model, reaching high parasitological cure rates especially when combined with BZ,19 arguing in favour of continuing with studies testing CYP51 inhibitors.6
Focusing on drug discovery, Alberca and colleagues conducted virtual screening campaigns based on ligands and target structures and identified butoconazole (BTZ) as a promising anti-T. cruzi agent.20 Then, our aim was to further explore the therapeutic potential of the BTZ in vitro and in vivo assays of experimental T. cruzi infection, in monotherapy or in combination with BZ, both on bloodstream trypomastigotes and intracellular amastigotes, by testing also distinct parasite strains.
MATERIALS AND METHODS
Compounds - BZ (Fig. 1A) obtained from LAFEPE (Pharmaceutical Laboratory of the State of Pernambuco Governador Miguel Arraes). BTZ (Fig. 1B) provided in the form of its nitrate salt by Drs Alan Talevi and Lucas Alberca (Laboratorio de Investigación y Desarrollo de Bioactivos - LIDeB/AR, Buenos Aires, Argentina) based on a previous virtual and in vitro preliminary screenings.20 For our in vitro studies, the stock (50 mM) solutions were prepared in dimethyl sulfoxide (DMSO) (Sigma-Aldrich, St. Louis, MO, USA) and used with final working concentrations never exceeding 0.6%, meaning that there is no toxic effect on mammalian cells or parasites in vitro.21 For in vivo evaluations, BTZ was dissolved in 10% DMSO while BZ in 3% Tween 80 and then both were diluted in sterile and deionised water.
Fig. 1:chemical structure of the compounds: (A) benznidazole (BZ) and (B) butoconazole (BTZ).
Mammalian cell cultures - L929 fibroblast lineage were cultured in Roswell Park Memorial Institute 1640 medium without phenol red (RPMI - Sigma-Aldrich, St. Louis, MO, USA), supplemented with 10% foetal bovine serum (FBS - Cultilab - SP - Brazil) and 2 mM glutamine (Sigma-Aldrich, St. Louis, MO, USA) with pH 7.2 - 7.4, as previously reported.21 H9C2, cardiomyoblast cultures, were maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) high glucose (4500 mg/L) medium supplemented (Sigma-Aldrich, St. Louis, MO, USA) with 10% FBS (Cultilab - SP -Brazil), 2 mM glutamine (Sigma-Aldrich, St. Louis, MO, USA) and 100 μg/mL penicillin/streptomycin (Sigma-Aldrich, St. Louis, MO, USA).22 Both cell cultures maintained at 37ºC under an atmosphere of 5% CO_2_, with weekly dissociation protocol using 0.01% trypsin.
Parasites - Trypomastigotes of the Y strain of T. cruzi [discrete typing unit (DTU) II] were acquired by cardiac puncture of Swiss Webster mice at the peak of parasitaemia.23 Trypomastigotes of the Tulahuen strain (DTU VI), expressing the β-galactosidase gene of Escherichia coli, were collected from the supernatant of previously infected L929 cultures.21
Cytotoxicity assay - The toxicity of the compounds was evaluated on two mammalian cell lines (L929 and H9C2), as reported.21 The cells were seeded in 96-well plates (4 x 10^3^ cells/well - for L929; 25x10^3^ cells/well - for H9C2) and incubated at 37ºC and 5% CO_2_. The L929 cultures were kept under treatment for 96 and 168 h with a serial dilution (1:2) of the compounds (up to 400 μM). After the treatment periods, the AlamarBlue^®^ reagent (Thermo Fisher Scientific Inc) was added and incubated for 24 h. The H9C2 cultures were also treated in serial dilution up to 200 μM for 24 h. PrestoBlue^®^ reagent (Thermo Fisher Scientific Inc) was added to the cultures and incubated for 3 h. All plates were analysed in the spectrophotometer SpectraMaxM3 (570 and 600 nm) and LC_50_ values (concentration capable of inducing loss of viability in 50% of the cells) determined.21 All assays were done at least twice, in triplicates.
In vitro activity on intracellular forms of T. cruzi - L929 cells previously seeded in 96-well plates (4 x 10^3^ cells/well) were infected (ratio of 10 parasites:1 host cell) for 2 h with trypomastigote forms (Tulahuen strain transfected with the β-galactosidase gene, DTU VI). After 48 h, the cultures were incubated with the compounds (0 - 10 μM) for 96 h at 37ºC/ 5% CO_2_ and the enzymatic activity measured at 570 nm after the addition of chlorophenol red (CPRG) to determine EC_50_ and EC_90_ values (minimum concentration able of eliminating 50 and 90% of the infection, respectively).21
In vitro activity on bloodstream trypomastigotes - Swiss male mice were inoculated with 5 x 10^4^ bloodstream trypomastigotes (BT, Y strain, DTU II) and at the parasitaemia peak [8th post-infection (dpi)], euthanised using 3% isoflurane and blood samples obtained by cardiac puncture (CEUA FIOCRUZ L038-2017). Then, 100 μL of parasite suspension (10^7^/mL in RPMI medium + 5% FBS) was added to the same volume of RPMI + 5% FBS containing treatment with BTZ (serially diluted - 1:2) at twice the desired final concentration. After 2 and 24 h at 37ºC, the number of live parasites was determined by light microscopy quantification using a Neubauer chamber.6 Untreated controls were carried out with parasites kept under the same conditions and BZ was run in parallel. The activity of the compounds was expressed by the EC_50_ and EC_90_ values. The assays were done at least twice, in duplicate.
Drug combination on intracellular forms of T. cruzi - L929 were seeded in 96-well plates (4 x 10^3^ cells/well) and infected with *T. cruzi (*Tulahuen strain). Predetermined EC_50_ values were used to determine the top concentrations of the test compounds under fixed ratios ensuring that EC_50_ fell in the midpoint of a seven-point two-fold dilution series. The fixed ratios were 5:0, 4:1, 3:2, 2:3, 1:4 and 0:5.24 ^,^ 25 According to Odds (2003), the Fractional Inhibitory Concentration Indexes (FICI) was calculated by the ratio of the EC_50_ values of each ratio of the combinations/EC_50_ of each compound alone. Finally, the sum of the FICIs (ƩFICI) is calculated to classify the interaction nature of the combinations.26 ΣFICI ≤ 0.5 = Synergism; ΣFICI between 0.5 and 4 = additive (No interaction); ΣFICI > 4 = antagonism. At least two independent experiments were performed in triplicate.
Washout assays in L929 cultures infected with T. cruzi - L929 cultures were infected with Tulahuen strain as above described and incubated or not for 168 h with 10 µM of BZ and BTZ (37ºC/5% CO_2_). RPMI culture medium (containing the tested compounds) was replaced every 72 h. Then, the cultures were rinsed with 0.1M PBS, and the drug-free culture medium added for another 168 h, replacing it every 72 h. The number of parasites released into the supernatant and of intracellular forms were analysed by light microscopy and by spectrophotometry (570 nm) after adding CPRG, respectively. The results were expressed as the percentage of reduction in parasite growth compared to untreated cells. BZ was used as a positive treatment control.25 The data results from two independent experiments done in at least triplicate.
Ethics - The pharmacokinetic study that was conducted under supervision of Dr Talevi according to protocols approved by the Institutional Committee for the Care and Use of Laboratory Animals (CICUAL) of the Faculty of Exact Sciences of the University of La Plata, Argentina (FCE-UNLP) (protocol number 004-00-23). In parallel, all animal studies were carried out in strict accordance with the guidelines established by the FIOCRUZ Committee of Ethics for the Use of Animals (CEUA L038-2017), and were approved by CTNBio and CIBIO/IOC/FIOCRUZ for a Biosafety Quality Certificate (CQB 105/99) for the use of GMOs (T. cruzi strain Tulahuen transfected with β-galactosidase gene), in addition to registration in the SisGen (A5825BF).
In vivo pharmacokinetic study - Adult male specific-pathogen-free BALB/c mice, provided by the Faculty of Veterinary Sciences, National University of La Plata (FCV-UNLP) were used. Animals were housed in cages with four animals per cage, under a 12 h light/dark cycle and controlled temperature, provided with water and food ad libitum and environmental enrichment. A minimum period of six days was given after transport for physiological acclimatisation before any procedure. Five groups of mice (n = 3 per group) were used, one per sampling time. A fresh solution of BTZ at 50 mg/mL in DMSO (BIOPACK) was administered intraperitoneally (i.p.) at an injection volume of 2 mL/kg, achieving a final dose of 100 mg/kg. Each mouse was sampled only once, collecting a maximum of 150 µL of submandibular blood at 1, 2, 4, 8 and 24 h post-administration, and transferred into EDTA-treated tubes. Plasma was obtained immediately after blood collection by centrifugation for 10 min at 4,000 rpm and conserved at -17ºC until quantification. After the experiment, all animals were euthanised using CO_2_. The i.p. route of BTZ was chosen due to its low oral bioavailability.
Plasma sample preparation and LC-MS/MS analysis - The plasma samples were treated by keeping them in an ice-water bath. An equal volume of cold acetonitrile (pre-cooled in a freezer) was added to induce protein precipitation. The mixture was vortexed for 10 s and then allowed to stand for 15 min in the cold bath. Subsequently, the samples were centrifuged at 10,000 rpm at 4ºC for 15 min. After, the supernatant was collected, diluted with water/acetonitrile (1:1), and filtered through a 0.22 µm nylon membrane filter before being transferred to an insert for injection into the LC-MS/MS system.
The equipment used was a Waters Alliance 2695 HPLC coupled to a tandem quadrupole mass spectrometer, Premier XE (Waters Corp.), equipped with an electrospray ionisation source operating in positive mode (ESI+). Chromatographic separation was performed using an X-SELECT™ C_18_ column (75 mm × 4.6 mm, 3 µm pore size; Waters Corp.) with a gradient of methanol and ultrapure water, with formic acid and ammonium acetate as ionisation additives. The mass spectrometer was configured to operate in multiple reaction monitoring (MRM) mode, employing two mass transitions for the compound, one for quantification (411.0 > 164.9) and the other for confirmation (413.2 > 164.9).
Unbound drug concentrations in plasma were estimated by multiplying the analytically determined total drug concentrations by the unbound fraction predicted by Deep-PK.27
In vivo analysis in a mouse model of acute T. cruzi infection - Swiss male mice (18 - 20 g) were obtained from the Institute of Sciences and Technologies in Biomodels (ICTB-FIOCRUZ), housed with a maximum of 5 animals per cage and kept in a specific pathogen-free (SPF) room at 20 - 24ºC under a 12 h light/dark cycle. All animals received sterilised water and food ad libitum. The animals were acclimatised for seven days before the experiments. For a proof-of-concept study, animals were infected by i.p (1x10^4) with bloodstream trypomastigotes (Y strain). The drugs administration started at the onset of parasitaemia (5th day dpi), only using mice with detectable parasitaemia.19 ^,^ 28 Control mice groups were age-matched and housed under identical conditions. The following experimental groups were used ≥ 3 mice per group: untreated (infected vehicle-treated control) and treated orally (p.o) with BZ (10, 25 and 100 mg/kg/day), corresponding to suboptimal and optimal doses of the reference drug; infected and treated with BTZ (i.p) using concentrations adjusted to be equimolar to BZ doses (18.24, 45.61 and 182.44 mg/kg/day). Also, the following combo was administered: BZ (10 mg/kg/day/p.o) + BTZ (18.24 mg/kg/day/i.p). The animals received the drugs once a day for five consecutive days. A 10-fold lower optimal dose of BZ (10 mg/kg/day) was chosen to clearly observe any benefit of its association with BTZ. The i.p. route for BTZ was chosen due to its low aqueous solubility.
Parasitaemia and mortality rate - Parasitaemia was performed by collecting 5 μL of the blood from the tail’s vein, disposing it on a slide and covering with coverslips (18 x 18 mm) for direct counting of parasites observed in 50 fields under light microscope (40X magnification).19 ^,^ 28 The mortality was followed daily up to 30 days after the end of treatment and expressed as a percentage of cumulative mortality (% CM).19 ^,^ 27 In vivo assays were conducted with n = 5 per group.
In vivo efficacy assessments by qPCR evaluations - At the end point, the animals were weighed; euthanised and submitted to cardiac puncture. 500 μL of fresh blood was collected and added to 1 mL of guanidine. The DNA was extracted from 300 µL of guanidine-EDTA blood, using the kit High Pure PCR Template Preparation (Roche diagnostics, Mannhein, Germany), according to the manufacturer’s instructions. At the last step of the protocol, DNA was eluted in 100 µL of elution buffer. Also, cardiac tissues were collected, weighed and fragmented using a scalpel, 200 μL of Tissue Lysis Buffer was added and samples immediately placed in Styrofoam with dry ice. Blood and heart tissues were stored in the freezer at -80ºC until analysis by qPCR. Heart tissues were homogenised using a Tissue Homogeneizer (Qiagen). DNA was extracted using the kit High Pure PCR Template Preparation (Roche diagnostics, Mannhein, Germany), according to the manufacturer’s instructions for tissue samples. qPCR was done using a TaqMan multiplex assay, targeting T. cruzi satDNA and mouse GAPDH, as an endogenous control, and to normalise the parasite load. To obtain standard curves, serial dilutions of the DNA were prepared in appropriate plates for the assay. After normalising the results obtained on the samples, the parasite load was expressed as parasite equivalents/mL when evaluated in blood, or parasite equivalents/mg when in mouse tissue. In all trials, positive and negative controls were also tested in parallel.
Data analysis and statistics - The data were plotted and evaluated using the GraphPad Prism version 5.0 program (GraphPad Software, San Diego, CA, USA). The Deep-PK platform (available for free at https://biosig.lab.uq.edu.au/deeppk) was applied for prediction of PK and toxicity data of BTZ.27
RESULTS
The first step was evaluating the cytotoxic profile of BTZ on L929 cells treated for 96 h. BTZ displayed a LC_50_ value of 23.9 μM ± 1.1. while BZ was non-toxic up to 400 μM (Table I, Fig. 2). Next, the trypanosomicidal activity was investigated using L929 cultures infected with the Tulahuen strain (Fig. 3A-B, Table I). BTZ was highly potent with EC_50_ = 0.07 ± 0.02 μM while BZ reached 1.73 ± 0.16 μM. Regarding selectivity, BTZ displayed an excellent SI of 341, being about 24-fold more potent than the reference drug (BZ) (Fig. 3, Table I).
TABLE IThe activity (mean value and standard deviation - SD) of benznidazole (BZ) and butoconazole (BTZ) against intracellular forms (Tulahuen strain) of Trypanosoma cruzi, the mammalian cytotoxicity and respective selectivity indexes (SI)CompoundsActivity against T. cruzi intracellular forms - μM - 96 h (pEC_50_)Cytotoxicity on L929 cells - μM - 96 h (pEC_50_)Cytotoxicity on L929 cells -μM - 168 h (pEC_50_)SI^ c ^ EC_50_ ^ a ^ EC_90_ ^ a ^ LC_50_ ^ b ^ LC_50_ ^ b ^ BZ1.73 ± 0.16 (pEC_50_ 5.76)7.65 ± 1.69> 400> 200 (pEC_50_ 3.69)> 231BTZ0.070 ± 0.02 (pEC_50_7.15)3.88 ± 0.4523.90 ± 1.117.53 ± 3.24 (pEC_50_ 4.62)341 a: EC_50_/EC_90_ concentration capable of reducing 50% and 90% of infection, respectively; b: LC_50_ concentration capable of reducing 50% of viable cells; c: SI ratio between LC_50_/EC_50_ values considering 96 h of incubation.
Fig. 2:cytotoxicity profile of benznidazole (BZ) and butoconazole (BTZ) on L929 mammalian cell lines after 96 h of incubation at 37ºC
Fig. 3:the trypanosomicidal effect of butaconazole (BTZ) and benznidazole (BZ) against intracellular forms of Trypanosoma cruzi (Tulahuen strain) after 96 h of incubation at 37ºC. (A) Dose-dependent activity of BZ and BTZ up to 10 µM. (B) Dose-dependent activity of BTZ up to 1 µM.
To further explore toxicity of BTZ in vitro now upon longer exposure, L929 were treated for 168 h. Our findings showed that BTZ and BZ displayed LC_50_ values of 17.53 μM ± 3.24 μM and > 200 μM, respectively (Table I).
The potential cardiotoxic effect was also assessed using H9C2 cell lines. Our findings demonstrated LC_50_ values of 36 ± 0.25 and >200 μM after 24 h of incubation with BTZ and BZ, respectively (Table II).
TABLE IIThe activity (mean value and standard deviation - SD) of benznidazole (BZ) and butoconazole (BTZ) against bloodstream trypomastigotes (Y strain) of Trypanosoma cruzi, their cardiotoxicity and respective selectivity indexesCompoundsActivity on T. cruzi (Y strain) blood trypomastigotes - µM (pEC_50_)Toxicity on H9C2 mammalian cells in µM (pEC_50_)EC_50_ ^ a ^ - 2 hEC_90_ ^ a ^ - 2 hEC_50_ ^ a ^ - 24 hEC_90_ ^ a ^ - 24 hLC_50_ ^ b ^ - 24 hBZ> 20> 205.93 ± 0.15 (pEC_50_ 5.22)17.54 ± 0.65> 200 (pEC_50_ 3.69)BTZ> 20> 2013.83 ± 0.70 (pEC_50_ 4.85)18.64 ± 0.1136.36 ± 0.25 (pEC_50_ 4.43) a: EC_50_/EC_90_ concentration capable of reducing 50% and 90% of parasites, respectively; b: LC_50_ concentration capable of reducing 50% of viable cells.
The effect against bloodstream trypomastigotes (BT) revealed that BTZ was 2.3-fold less potent than BZ after 24 h of incubation displaying EC_50_ values of 13.83 ± 0.70 and 5.93 ± 0.15 µM, respectively (Table II).
Washout assays showed that BTZ and BZ suppressed the parasite release into the supernatant of L929. Also, BTZ and BZ incubation resulted in > 96% reduction of the intracellular parasitism (Table III).
TABLE IIIThe washout analysis performed against intracellular forms of Trypanosoma cruzi (Tulahuen strain) in L929 cell lines after incubation with benznidazole (BZ) and butoconazole (BTZ). The data displays the percentage (%) of reduction in the number of released and internalized parasites. Results represent mean and standard deviation (SD)CompoundsReleased parasitesIntracellular formsBZ100 ± 098.6 ± 1.4BTZ100 ± 096 ± 4
The BTZ+BZ combo effect against intracellular forms demonstrated that the FICI ranged from 0.35 to 1.28 revealing an additive drug interaction, with a value xΣFICI = 0.66 (Table IV, Fig. 4).
The next step was to evaluate the concentration vs. time profile of the compound in mice after administration in a single dose of 100 mg/kg of BTZ. The plasma drug concentration levels were monitored at 1, 2, 4, 8 and 24 h after administration (Fig. 5). The mean peak concentration after 2 h of administration showed a total plasma concentration of 25.9 µM. Interestingly, when correcting the total concentration by the estimated unbound fraction calculated with Deep-PK (1.67%), even a single administration with this dose would provide an unbound concentration above the EC_50_ against amastigotes for around 7 h. Noteworthy, since the estimated EC_50_ considers both bound and unbound drug in the culture medium, the unbound EC_50_ value is expected to be below the observed EC_50_ if binding to proteins in the culture medium has occurred.
TABLE IVThe analysis of the combinatory effect of butoconazole (BTZ) and benznidazole (BZ) on intracellular forms of Trypanosoma cruzi (Tulahuen strain) in L929 cell linesProportion of each drugDrug concentrationFICI^ a ^ ∑FICI^ b ^ x∑FICI^ c ^ BTZ | BZBTZBZBTZBZ5 | 00.031-10 0.664 | 10.0240.7380.760.511.283 | 20.0060.4020.200.280.482 | 30.0120.2190.370.150.521 | 40.0080.1380.260.100.360 | 5-1.44501
a: fractional inhibitory concentration index; b: sum of the FICIs; c: mean value of ƩFICIs.
Fig. 4:isobologram of benznidazole (BZ)and butoconazole (BTZ) used in combination. The result shows an additive effect on intracellular forms of Trypanosoma cruzi (Tulahuen strain).
Fig. 5:the pharmacokinetic profile of butoconazole (BTZ) plasma levels measured up to 24 h after in vivo administration of a single dose of the compound in a total of 15 mice (three mice per sampling time): (A) Mean total plasma concentration - where the peak recorded was 25.9 μM in the first 2 h after administration; (B) Estimated unbound drug concentration vs. time. The unbound drug concentration was estimated using Deep-PK tool to predict the unbound fraction (1.67%) and revealed that BTZ, even in a single dose, has a high potential to provide unbound plasma concentrations above EC_50_ for at least 7 h.
Based on the compilation of our findings, we proceeded with the antiparasitic activity of BTZ tested in vivo in a mouse model of acute infection. The animals were treated alone or in co-administration with BZ. Male Swiss mice were infected with T. cruzi and treated for five consecutive days, starting at the parasitaemia onset (5 dpi). Two different assays were performed. In the first one, animals were treated with 25 and 45.61 mg/kg/day of BZ and BTZ, respectively. Our findings demonstrated that BZ and BTZ gave 93 and 50% of peak decline at the 8 dpi (Fig. 6A). Regarding mortality, while untreated mice reached 80% of death, BZ and BTZ gave 0 and 40%, respectively (Fig. 6B).
Fig. 6: in vivo effect of benznidazole (BZ) and butoconazole (BTZ) alone (A and B) or in combination (C and D) on experimental mouse model of Trypanosoma cruzi infection. (A and C) Parasitaemia curves from assays 1 and 2 respectively; (B and D) Percentage of cumulative death (%) relative of assays 1 and 2, respectively. Five mice (n = 5) were used in each experimental group. Dashed lines (-----) mean the period of drug administration (five consecutive days - 5-9 dpi).
Next, sub-optimal and optimal doses of both drugs (10 and 100 mg/kg/day for BZ and 18.24 and 182.44 for BTZ, respectively) were tested. BTZ at its highest concentration (182.44 mg/kg/day) reduced by 63% the parasitaemia (p = 0.45), while BZ (100 mg/kg/day) suppressed the peak (p = 0.000047) (Fig. 6C).
The lowest concentrations of both BTZ and BZ only slightly (p > 0.05) reduced the parasitaemia (Fig. 6B). Regarding animal mortality, at the lower doses, BTZ and BZ reached 75% and 100% mortality (Fig. 6D). As depicted in the Fig. 6C, the combo treatment (BZ + BTZ) reduced by 47% (p = 0.026) the parasitaemia with higher effect as compared to the drugs given alone (Fig. 6C). The combo and the higher doses of BTZ and BZ resulted in 100% animal survival, contrasting to 33% of mice survival in vehicle group (Fig. 6D).
The qPCR analysis on blood samples from the first trial showed that all surviving mice treated with BTZ (45.61 mg/kg/day) were negative (three of three animals), while for the BZ group, parasite DNA was amplified in all animals (Fig. 7A). In the second trial, regarding blood samples, only one animal treated with BZ and BTZ, at higher doses, amplified parasite DNA. All surviving animals in the COMBO group were negative for parasite DNA amplification (Fig. 7B). When cardiac samples were evaluated, BZ 100 mg/kg/day gave 100% negative qPCR, while only two of five mice treated with BTZ 184 mg/kg/day were negative. For COMBO (BTZ + BZ, at 18.24 and 10 mg/kg/day, respectively), all surviving mice showed positive amplification for parasite DNA, demonstrating drug failure (data not shown).
Fig. 7:the determination of blood parasite load by quantitative polymerase chain reaction (qPCR) in benznidazole (BZ) and butoconazole (BTZ) treated mice alone (A) or in combination (B) during Trypanosoma cruzi infection. (A) vehicle, BZ (25 mg/kg/day) and BTZ (45.62 mg/kg/day) treated animals (assay 1) and (B) vehicle, BZ (100 mg/kg/day) and BTZ (182.44 mg/kg/day), and combo (BZ + BTZ (assay 2).
DISCUSSION
The current available drugs to treat CD have unsatisfactory efficacy and several limitations, reinforcing the need for new low-cost treatments that have higher safety and efficacy.6 ^,^ 12 ^,^ 29
The clinical success of repurposing drugs has stimulated the systematic approach for drug screening, providing the potential to reduce costs and time between preclinical and clinical steps, which are especially valuable characteristics to address the development of new therapeutics solutions for neglected tropical diseases.30 Among the treatment strategies, the drug combinations show promising results in different therapeutic fields resulting in increased efficacy, reduced dose regimens and time of drug administration that minimises the chances of adverse effects and drug resistance, enhancing patient adherence.6 ^,^ 31
Therefore, the objective of our study was to evaluate the therapeutic efficacy of BTZ alone or in combination with BZ on experimental T. cruzi infection. BTZ was identified as a promising drug candidate for CD by Alberca and colleagues using a computational strategy to explore repurposing-oriented chemical databases.20
Our present data corroborates these studies regarding the trypanosomicidal effect of BTZ against T. cruzi in vitro as it was highly effective against intracellular amastigotes at sub micromolar doses, being about 24-fold more potent than BZ. However, BTZ was less potent against bloodstream trypomastigotes as compared to the reference drug. Our data are aligned to the current literature related to the trypanosomicidal effect of sterol biosynthesis inhibitors, especially against the multiplicative amastigote forms, which are much more metabolically active than trypomastigotes.31 ^,^ 32 ^,^ 33 In parallel, cytotoxicity tests were conducted on different mammalian cell types, and a mild time-dependent toxicity was observed, as reported besides a mild cardiotoxicity.20
One of the main obstacles to be overcome for the successful treatment of CD is to find an alternative that is effective and sustained in both phases of the disease avoiding parasite relapses.6 Therefore, we performed washout assays and monitored the parasite release into the cell culture supernatant besides evaluating the percentage of intracellular parasitism to inspect for quiescent parasites. BZ and BTZ eliminated parasite release into the supernatant but were unable to sterilise the infection, achieving about 96 - 98% of decline in the intracellular parasite load due to presence of latent forms.
Another approach was evaluating the effect of BTZ + BZ association in vitro. Our findings demonstrate an additive effect (xΣFICI = 0.66), remarkably close to a synergistic interaction, which is a very promising aspect.
The PK study demonstrated that a single 100 mg/kg dose of BTZ, i.p, was readily bioavailable, with peak plasma concentrations two hours after administration and, more important, estimated unbound concentrations above the EC_50_ against amastigotes over several hours. Reasonably, the exposure is expected to be higher in the context of repeated dosing schemes or upon the administration of higher doses.
In view of these results, we checked a proof-of-concept using a murine model of experimental acute infection. BTZ administered alone gave moderate trypanosomicidal activity, achieving a maximum reduction of 63% at the peak when the highest concentration was administered (182.44 mg/kg/day). BZ (100 mg/kg/day) fully suppressed parasitaemia and gave 100% animal survival.
The lowest dose of BTZ (18.24 mg/kg/day) guaranteed mice survival ≥ 50%, while in the lowest dose of BZ (10 mg/kg/day) 100% animal mortality was observed. Also, the combination of the lowest dose of BZ and BTZ reaching 47% of parasitaemia decline, improved the effect in vivo as compared to monotherapies providing 100% of mice survival. This data corroborates the promising in vitro additive activity profile.
The qPCR findings of mouse blood and heart samples demonstrated that the cardiac tissues provide a more sensitive detection of T. cruzi infection. Although 75% of the BTZ (182.44 mg/kg/day) gave negative amplification in all blood samples, the corresponding cardiac analysis demonstrated cure failure in 50% of the surviving animals. Also, in the combo group composed by lower doses of both drugs, despite all treated mice survived, no parasitological cure was achieved since the parasite satDNA was amplified in the heart of all surviving animals.
The bulk of our findings encourage future studies performing different combo protocols with these as well as other repurposing drugs to contribute for new therapeutic options for treating neglected tropical diseases.
In conclusion - Our findings corroborated and expanded previous findings regarding drug repurpose and combination approaches for experimental analysis of novel therapies for CD. Further studies using additional protocols as well as BTZ analogues may contribute to the identification of novel therapeutic strategies to combat this silent and neglected disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1de Sousa AS Vermeij D Ramos AN Jr Luquetti AO Chagas disease Lancet 2024403104222032183807198510.1016/S 0140-6736(23)01787-7 · doi ↗ · pubmed ↗
- 2Pérez-Molina JA Molina I Chagas disease Lancet 20183911011582942867342310.1016/S 0140-6736(17)31612-4 · doi ↗ · pubmed ↗
- 3Velasco A Morillo CA Chagas heart disease a contemporary review J Nucl Cardiol 20202724454513002240410.1007/s 12350-018-1361-1 · doi ↗ · pubmed ↗
- 4De La Rosa E.Paglini-Oliva P.Prato LB.Benizio E.Triquell MF.Muñoz SE Early detection of chronic asymptomatic Chagas infection Med Sci Monit 201824456745712996595610.12659/MSM.907735 PMC 6058734 · doi ↗ · pubmed ↗
- 5Schijman AG Alonso-Padilla J Britto C Bernal CPH Retrospect, advances and challenges in Chagas disease diagnosis a comprehensive review Lancet Reg Health Am 2024361008211008213900612610.1016/j.lana.2024.100821 PMC 11246061 · doi ↗ · pubmed ↗
- 6Soeiro MNC Sales-Junior PA Pereira VRA Vannier-Santos MA Murta SMF Sousa AS Drug screening and development cascade for Chagas disease an update of in vitro and in vivo experimental models Mem Inst Oswaldo Cruz 2024119 e 2400573895834110.1590/0074-02760240057 PMC 11218046 · doi ↗ · pubmed ↗
- 7Garg NJ Current global situation of Chagas disease Rev Biomed 20213226668
- 8PAHO Chagas disease 2019 https://www.paho.org/en/topics/chagas-disease