O-acyltransferase genes involved in the production of volatile sex pheromones in Caenorhabditis elegans
Xuan Wan, Sarah M. Cohen, Yan Yu, Henry Hoan Le, Heenam Park, Alessandro Groaz, Rachel Moreno, Minyi Tan, Jessica Schneider, Matthew R. Gronquist, Ryoji Shinya, Frank C. Schroeder, Paul W. Sternberg

TL;DR
Researchers identified four O-acyltransferase genes in C. elegans that are essential for producing volatile sex pheromones, which attract males.
Contribution
The study reveals functional specialization among OAC gene paralogs in pheromone biosynthesis and signaling.
Findings
Four OAC genes (oac-13, oac-16, oac-25, oac-28) are required for volatile sex pheromone production.
OAC-13 and OAC-16 are necessary for both major pheromone components, while OAC-25 and OAC-28 regulate one component.
OAC genes also modulate nonvolatile ascaroside pheromone synthesis and secretion.
Abstract
As a step toward identification of the volatile sex pheromones (VSPs) that attract males as well as to understand where and under what conditions they are produced, we sought to identify genes necessary for VSP production. We generated loss-of-function mutants of 56 oac genes, finding that four are necessary for production of the (VSPs). VSPs are made up of at least two components, identified by their differential behavior in GC-MS. oac-13 and oac-16 are necessary for both major VSP components, while the tandem paralogs oac-25 and oac-28 are necessary for only one component. We found that oac-16, oac-25, and oac-28 have specific expression in the epidermal “seam” cells, implicating these lateral midline cells as the principal sites of VSP biosynthesis. Gene family expansions are critical for functional diversification, yet the contributions of paralogs to metabolic pathways are often…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
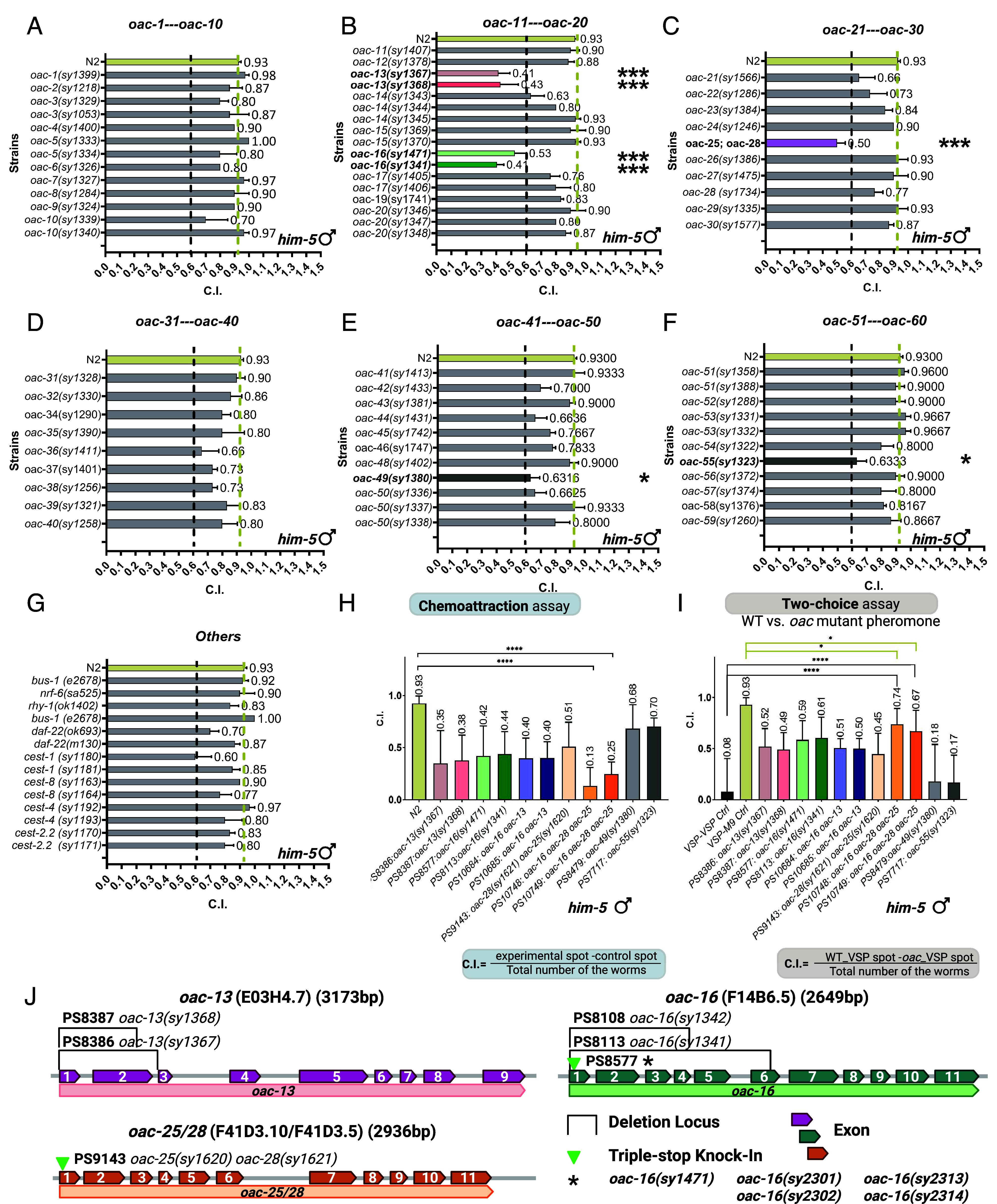 Fig. 2
Fig. 2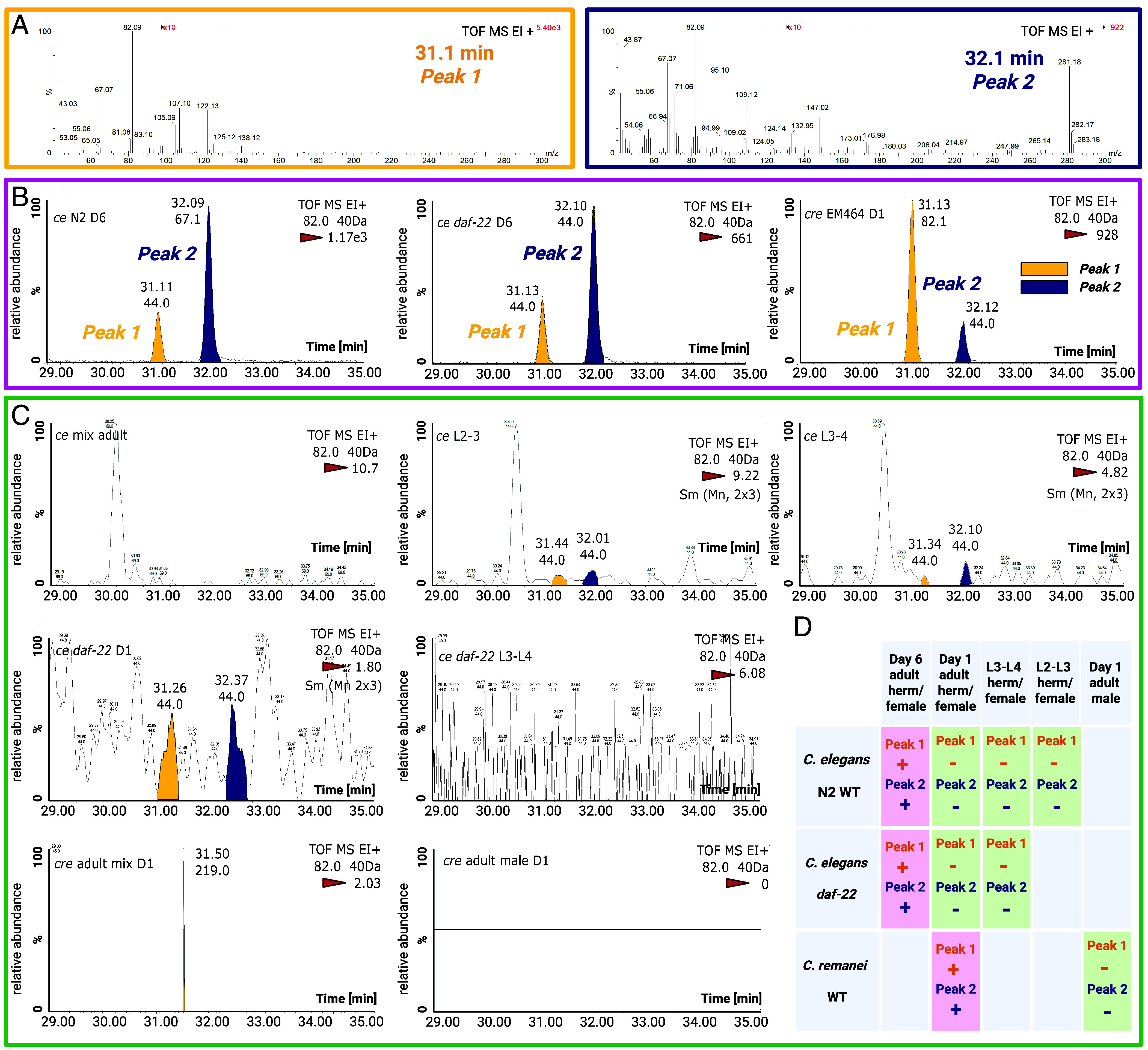 Fig. 3
Fig. 3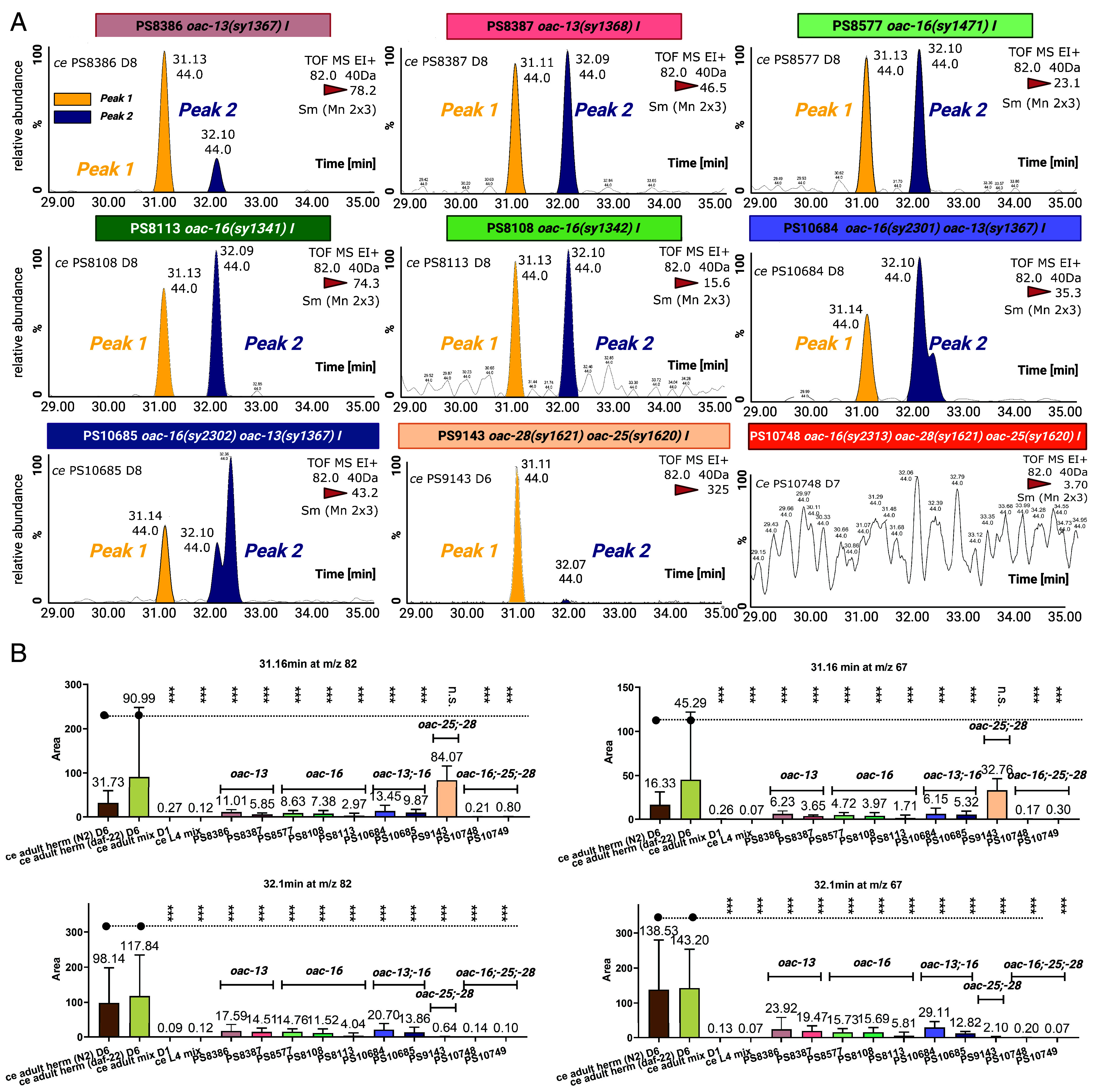 Fig. 4
Fig. 4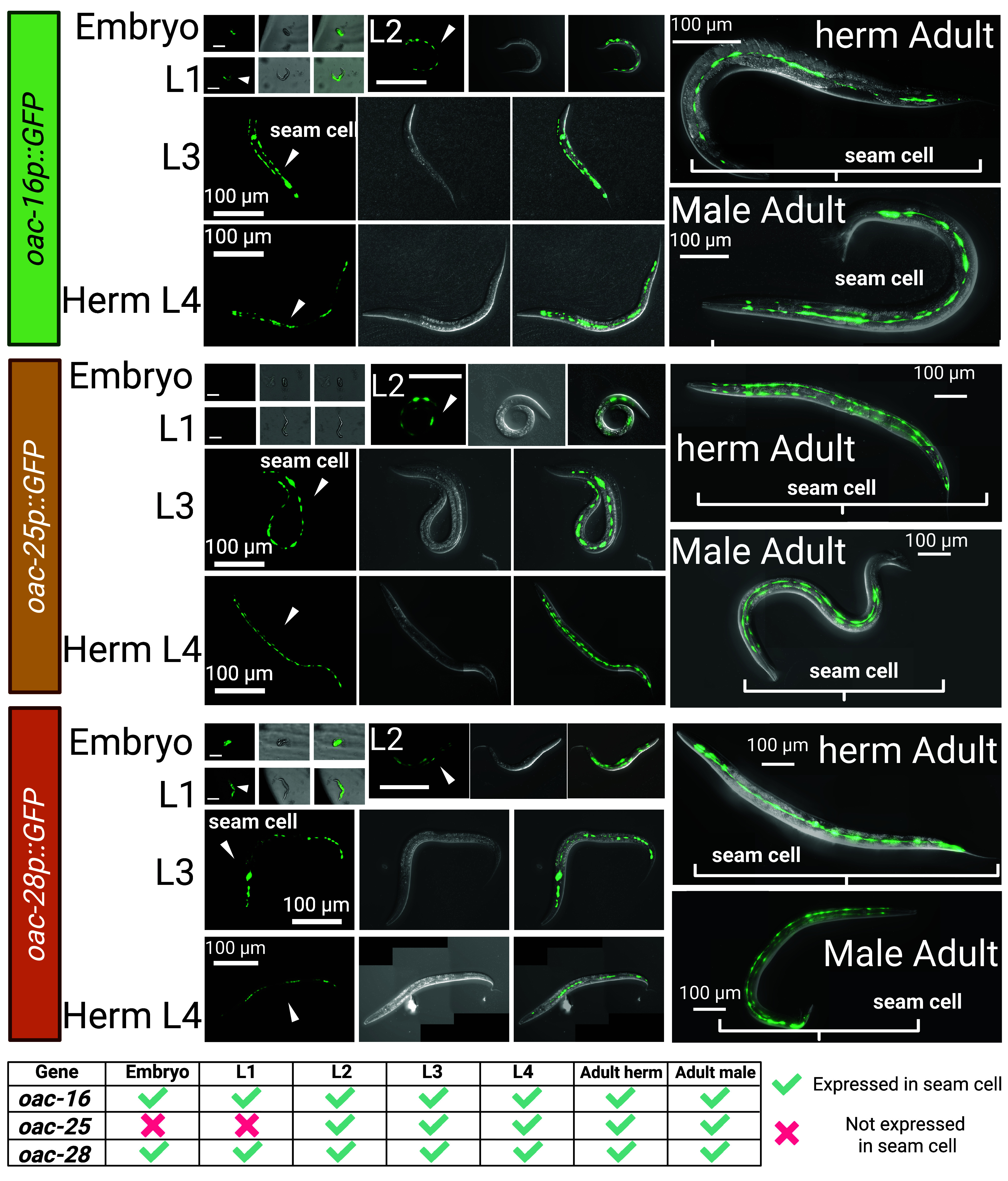 Fig. 5
Fig. 5- —HHS | NIH (NIH)100000002
- —NSF | NSF Graduate Research Fellowship Program (GRFP)100023581
- —Tianqiao and Chrissy Chen Institute for Neuroscience
- —Chuck Lorre Research Scholars Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Neurobiology and Insect Physiology Research · Insect-Plant Interactions and Control
The nematode Caenorhabditis elegans has served as an important model organism for decades, owing to its transparency, compact genome, and fully mapped neural connectomes, features that enable unparalleled genetic and neurobiological studies (1?–3). C. elegans has approximately 20,000 genes (2, 4). Only about 4,000 have any described phenotype or other information derived from nematode-based in vivo or in vitro experiments, excluding bulk gene expression measurements and RNAi information (5). About half the genes have names, indicating either due to phenotypes directly studied in C. elegans, for homology with researched genes in other model organisms or to established protein domains (6). By comparison, about 40% of human genes have some known functions, and two-thirds of this information comes from model organisms (7). Thus, systematic analyses are important for understanding C. elegans, especially given its role as a model for nematodes in general and for human genes.
O-acyltransferases are a subset of the acyltransferase class of enzymes that catalyze the transfer of acyl groups onto hydroxyl groups. This contrasts them with N-acyltransferases, which transfer acyl groups onto amines. O-acyltransferases are ubiquitous throughout the cellular environment and are used in pathways ranging from bacterial growth of biofilms to Wnt signaling in flies, to the conversion of cholesterol to cholesteryl ester in humans (8?–10). The most commonly studied forms of O-acyltransferases are the membrane-bound O-acyltransferases, which include ACAT1 and PORCN and their orthologs in Drosophila and other model organisms (11, 12). C. elegans has several families of membrane-bound O-acyltransferases, such as mboa (membrane-bound O-acyltransferase) family, mom (more of MS), and the oac (O-acyltransferase) family (5). The mboa gene family (10 genes), and the mom gene family (5 genes) all contain a membrane-bound O-acyltransferase domain (MBOAT) (InterPro IPR004299; Pfam PF03062); and are related to genes in the Wnt and Hedgehog pathways (9, 13). However, the largest yet least-studied class of O-acyltransferases in C. elegans is named OAC gene family, which contains a different protein domain, the acyltransferase 3 domain (InterPro IPR002656; Pfam PF01757) (Fig. 1B). The OAC class of enzymes was named due to its predicted ability to transfer acyl groups other than amino-acyl groups, but it has otherwise been unstudied in C. elegans.
In addition, the acyltransferase 3 domain is highly conserved across species. In bacteria, this domain facilitates cell surface modifications that confer resistance to lysosomes and bacteriophages (14, 15). OAC is an evolutionary ancient and conserved gene family, spanning prokaryotes (archaea, bacteria) and eukaryotes (fungi, nematodes, insects, mammals). Notably, insects and nematodes exhibit multicopy retention of OAC genes in their genomes, and mammals retain only a single orthologous copy (16). A nonsense mutation in acyltransferase 3 domain caused a loss of function of ACYL3 in humans and chimpanzees during the evolution of the great apes (16). C. elegans genome harbors 59 protein-coding oac genes (5).
Chemical communication is vital for C. elegans reproduction. The nematode employs two pheromone classes: nonvolatile ascaroside pheromones, which regulate development and social behaviors (17????–22), and VSPs that guide long-range mate navigation (23???–27). Ascarosides comprise a chemical language within nematodes, regulating many aspects of development and social behaviors (17?–19, 21, 22, 28, 29). The VSPs are produced exclusively by self-sperm/sperm-depleted hermaphrodites/females to attract males. Their structures and biosynthetic pathway remains elusive. Many studies have shown that OACs are essential to volatile insect pheromone production (30?–32). The conservation of OAC-driven pheromone biosynthesis in other species raises compelling questions: Could C. elegans’s OAC family genes similarly regulate pheromone signaling? Why has the OAC family undergone expansion in C. elegans? Does this expansion reflect functional redundancy, subfunctionalization, or novel adaptations unique to nematode ecology?
To fully allow us to understand the C. elegans organism, we need to systematically create mutants and examine gene families rather than relying on a shotgun approach. In this paper, we chose the understudied OAC gene family to create strains and study their potential functions. In this study, we generated 56 mutants spanning the entire family and performed behavioral assays, GC-MS analysis, and comparative metabolomic profiling to assess their roles in pheromone production.
Results
Phylogenetic Analysis of the OAC Gene/Protein Family.
We compared the protein sequences and created a protein phylogenetic tree (Fig. 1A) to understand how the genes/proteins were related. We also found that four additional genes should be included in the OAC gene family (Fig. 1B). Although there are several strains available that contain an oac mutation as well as numerous other mutations, these mutant strains were less useful in identifying the specific functions of OAC genes (33). Therefore, we created at least one isogenic mutant strain for each protein-coding oac for which there was not already an existing complete loss-of-function mutant (34, 35). We conducted a systematic phenotypic characterization of these mutants by metabolomic profiling of volatile sex-pheromone production across the OAC gene class.
The OAC family is defined by the acyltransferase 3 domain, a conserved catalytic motif essential for acyl group transfer. In addition to the named oac genes, we identified three additional genes including this OAC-specific acyltransferase-3 domain: bus-1, rhy-1, and nrf-6. These genes were named prior to the designation of the OAC gene class. These genes were previously characterized for roles unrelated to acyltransferase activity. bus-1 is a gene that is expressed in the rectal epithelial cells, and bus-1 mutants are unaffected by the nematode rectal pathogen Microbacterium nematophilum, which causes tail-swelling, hence the name bacterially unswollen (36). rhy-1 is a regulator of the hypoxia-inducible factor HIF-1 through the egl-9 pathway; mutants of rhy-1 exhibit some egg-laying defects (37). Fluoxetine (Prozac) causes C. elegans nose-muscle contractions, yet nose resistant-to-fluoxetine (nrf) mutants are immune to this phenomenon (38). nrf-6 mutants also display a pale-egg phenotype caused by a lack of yolk granules; this causes subsequent retarded development and partial embryonic lethality (38). Based on their shared domain architecture, we include bus-1, rhy-1, and nrf-6 in the OAC gene family. Following the inclusion of these genes, the OAC family comprises 63 genes, with four pseudogenes (oac-18, oac-33, oac-47, and oac-60) identified among them.
All 59 OAC protein-coding genes have an acyltransferase 3 domain. OACs are generally categorized into two groups based on their nonacyltransferase 3 domain architecture (Fig. 1C): 42 OACs have an acyltransferase 3 domain coupled with a C-terminal SGNH hydrolase domain (InterPro IPR043968), while 15 OACs have an acyltransferase 3 domain with an N-terminal nose resistant-to-fluoxetine (NRF) domain (InterPro IPR006621) (Fig. 1B). The SGNH hydrolase domain, often associated with carbohydrate modification (15), and the NRF domain suggest diverse functional roles for these enzymes. In general, membrane-bound O-acyltransferases have conserved histidine residues required for functional enzymatic activity (39, 40). We find that all the OACs that have an acyltransferase 3 domain likewise have a conserved catalytic histidine residue and the combination of Ser (S)-Tyr (Y)-His (H) residues in the acyltransferase 3 domain as shown in the motif logo (SYH catalytic motif, Fig. 1D and SI Appendix, Fig. S1). Tandemly duplicated genes, such as oac-21/oac-22/oac-23; oac-13/oac-16; oac-25/oac-28 tend to cluster together, suggesting recent gene duplication events. However, the overall phylogenetic structure does not correlate strongly with genomic loci, indicating functional divergence among paralogs.
Strain Construction for Functional Analysis.
To systematically investigate the OAC gene family, we obtained or generated knockout strains for all 59 protein-coding oac genes. Existing mutants for bus-1(e2678), nrf-6(sa525), and rhy-1(ok1402) were obtained from the Caenorhabditis Genetics Center (CGC). For the remaining 56 genes, we employed CRISPR/Cas9-mediated genome editing to create 95 mutant alleles (SI Appendix, Tables S1–S3). We used two strategies: 1) nonhomologous end joining to generate deletions of several hundred base pairs, and 2) homology-directed repair to introduce multiple early stop codons or frameshifts. These strains provide a comprehensive resource for probing the functional roles of OACs in C. elegans.
None of the oac mutant strains exhibited observable behavioral phenotypes, except oac-16(sy1342). Worms homozygous for this allele displayed a locomotion defect, characterized by restricted movement away from the hatching site. However, the other two oac-16 alleles (sy1341, sy1471) lacked this mobility defect, suggesting the sy1342 phenotype stems from an off-target mutation, rather than oac-16 loss of function. As this defect is unrelated to pheromone signals, we included oac-16(sy1342) strain in the pheromone extraction for GC-MS analysis.
OAC-13, OAC-16, OAC-25, and OAC-28 Are Required to Maintain the Male-Attracting Feature of Hermaphrodite-Derived Extracts.
Given the established roles of OACs in lipid modification and pheromone biosynthesis in other species (30?–32), we hypothesized that C. elegans OACs might similarly regulate VSP production. Using behavioral assays (Fig. 2) and GC-MS analyses (Figs. 3 and 4), we identified four OACs required for VSP biosynthesis.
*Comparative analysis of chemotactic responses to C. elegans VSPs. (A–G) Chemoattraction index of volatile pheromone extracts from wild type (N2) strain and 59 oac mutants, assayed with him-5 males (Materials and Methods). Most mutants retained wild type bioactivity, but oac-13, oac-16, oac-28 oac-25 (double mutant of identical paralogs), oac-49, and oac-55 exhibited significant reductions (below 0.6. Sample sizes: n = 60 males per trial, three trials. (H) Validation of VSPs signal defects in oac-13, oac-16, oac-28 oac-25 (double mutant of identical paralogs), oac-49, and oac-55 mutants in large-scale trials (n = 400 males per trial, 20 trials). (I) Two-choice preference assays showing male preference for wild type hermaphrodite extracts over mutant extracts (n = 400 males per trial, 20 trials). Mutants of tandem paralog pair oac-13, oac-16, oac-16 oac-13, and oac-28 oac-25 mutant exhibited reduced attractiveness compared to wild type extracts in both (H and I). The oac-16 oac-13 double mutant showed no additive defects, indicating functional redundancy. Error bars: S.D.; P < 0.005; P < 0.05 (two-tailed unpaired t test); ns., not significant (P > 0.05). The yellow dashed line represents the wild type N2 strain chemoattraction index. The black dashed line represents the screening threshold C.I. < 0.6. (J) Allele schematics for oac-13, oac-16, oac-25, and oac-28. Exon–intron maps show deletion spans (solid bars) and CRISPR edit sites (arrowheads); allele IDs and strain numbers are indicated. Complete junction sequences and genotyping primers see SI Appendix, Tables S1–S3. Mutation configurations: oac-13(sy1367), 708-bp deletion; oac-13(sy1368), 481-bp deletion; oac-16(sy1341), 1,255-bp deletion; oac-16(sy1342), 746-bp deletion; oac-16(sy1471) CRISPR triple-stop knock-in; oac-25(sy1620); oac-28(sy1621) CRISPR triple-stop knock-ins. All oac-16 alleles used in the single-, double-, and triple-mutant analyses—oac-16(sy1471), oac-16(sy2301), oac-16(sy2302), oac-16(sy2313), oac-16(sy2314)—harbor the same CRISPR triple “stop-in” cassette insertion.
GC-MS analysis identifies two conserved chromatographic peaks associated with reproductive-stage-regulated VSPs production. (A) Representative mass spectra of Peak 1 (tR = 31.1 min) and peak 2 (tR = 32.1 min) detected in pheromone-positive samples. Retention times and mass spectra were conserved across samples. (B) Representative GC-MS extracted ion chromatograms (EIC) for m/z 82.0 of a pheromone-positive sample (6-d-old self-sperm-depleted C. elegans N2 and daf-22 mutant hermaphrodites and 1-d-old not mated Caenorhabditis remanei female) showing Peaks 1 (yellow arrows) and Peak 2 (blue arrows). (C) Representative GC-MS EIC for m/z 82.0 of a pheromone-negative control lacking both peaks. Peaks 1/2 were exclusively detected in pheromone-positive samples. (B and C) Relative abundance (y-axis) is normalized to the highest signal intensity (100%). The intensity of the highest peak is labeled on each panel’s top-right (highlighted with red triangle). Retention time (x-axis, min) and annotated peaks reflect data acquired with ±0.40 Da mass accuracy in electron ionization positive mode (EI+). Peaks were smoothed using a 2 × 3 average filter [denoted as Sm (Mn 2 × 3)]. The m/z 82.0 ion corresponds to the target analyte, with the integrated peak area used for quantification. Background signals (e.g., m/z 44.0) are labeled under that peak’s retention times. TOF MS stands for Time-of-Flight Mass Spectrometry. (D) Stage- and status-dependent production of Peaks 1/2 across C. elegans and C. remanei: pink indicates pheromone production (post-sperm-depletion adults in C. elegans; virgin females in C. remanei), green indicates lack of pheromone production (larvae, self-sperm nondepleted C. elegans, mated C. remanei or males), and gray signifies untested conditions.
OAC-13, OAC-16, and identical paralogs OAC-25 and OAC-28 are required for the production of volatile pheromone components. (A) Representative GC-MS EIC for m/z 82.0 of (oac-13, oac-16, oac-16 oac-13 double mutant, and oac-28 oac-25 double mutant). Peaks 1/2 (tR = 31.1 and 32.1 min), identified in wild type pheromone-positive samples (Fig. 3), were absent or significantly reduced in all test mutants. Peaks 1 (yellow arrows) and Peak 2 (blue arrows). (B) Quantification of integrated peak areas for VSPs-related Peaks 1/2 (on m/z 82.0 ion and m/z 67.0 ion). Single mutants (oac-13 and oac-16) reduced abundances of both peaks similarly. Double mutants (oac-16 oac-13) showed no additive reduction compared with single mutants. oac-28 oac-25 double mutant abolished Peak 2 entirely, while Peak 1 remained unaffected. n = 5 to 8 biological replicates for each strain. Error bars: S.D.; P < 0.005; P < 0.05 (two-tailed unpaired t test); ns., not significant (P > 0.05).
Specifically, we extracted pheromones from hermaphrodites lacking self-sperm (to avoid sperm-derived signals that inhibit VSP production) (25) and evaluated their ability to attract wild type males. In particular, we evaluated their activity using a standardized chemoattraction assay with him-5(e1490) males (wild-type for oac genes). We adopted the pheromone extraction and chemoattraction assay protocol from Wan et al. with minor modification (26). Of 59 oac mutants screened (56 oac named genes and bus-1, nrf-6, rhy-1) most retained normal pheromone activity (Fig. 2 A–G). Strikingly, mutants of four genes on chromosome I—two tandem paralogs (oac-13 and oac-16) and two nearly identical paralogs (oac-25 and oac-28)—displayed defective volatile sex pheromone (VSP) production, suggesting impairments in biosynthesis, secretion, or in the availability of a precursor molecule or essential nutrient required for VSP production. oac-13 and oac-16 are 179 kilobases apart on the “forward” strand, while oac-25 and oac-28 are 6.4 kilobases apart on the reverse strand (Fig. 1E). Due to their nearly identical genomic sequences (99% similarity) and close proximity on chromosome I, oac-25 and oac-28 are difficult to target individually using standard genetic modification approaches. Furthermore, their coding sequences (CDS) are 100% identical, prompting us to analyze an oac-28 and oac-25 double mutant. During the generation of this double mutant, we obtained a single oac-28 mutant strain and performed pheromone extraction and assays on this strain in parallel. Allele configurations, including exon schematics with deletion spans and CRISPR stop-in/guide sites, are summarized in Fig. 2, with junction sequences and genotyping primers see SI Appendix, Tables S2 and S3.
From the screening, we found that pheromone extracts from oac-13 and oac-16 single mutants and oac-28 oac-25 double mutants elicited significantly reduced male attraction compared to wild type extracts, whereas the oac-28 single mutant did not (Fig. 3 B, C, and H). Larger-scale trials confirmed these defects (n = 20 trials, 400 males; Fig. 3H). We then evaluated an oac-16 oac-13 double mutant, which exhibited a similar level of male attraction defect as the cognate single mutants. Since oac-13 and oac-16 show similar male attraction to their double mutant, we tested oac-16 oac-28 oac-25 triple mutants. Strikingly, this triple mutant almost completely lost male attractiveness (C.I. = 0.19 from two strains, 14 VSP samples and n = 800 worms, Fig. 2H). Two additional mutants, oac-49 and oac-55, exhibited minor reductions in initial small-scale screening, but these reductions were not statistically significant in large-scale trials (Fig. 2H). We then adopted two-choice assays (Materials and Methods) and compared wild-type VSP extracts from 6-d-old adult N2 hermaphrodites against those from oac mutants. Males (him-5) strongly preferred wild type extracts over those from oac-13 and oac-16 single mutants, oac-16 oac-13 and oac-28 oac-25 double mutants, and oac-16 oac-28 oac-25 triple mutant (C.I. = 0.705, two strain). These assay results further confirm their pheromone deficiencies (Fig. 2I).
OAC-13 and OAC-16 share 60.18% amino acid sequence identity, while OAC-25 and OAC-28 are identical proteins. Further comparisons reveal 70.45% identity between OAC-25/OAC-28 and OAC-16, and 57.78% identity with OAC-13, highlighting significant sequence conservation among these structurally homologous proteins. In addition, all four genes cluster within a conserved genomic region on Chromosome I (13.07 to 13.20 cM).
GC-MS Analysis Identifies Two Conserved Peaks in Volatile Sex Pheromone Samples, Highly Regulated by Reproductive Status.
Caenorhabditis hermaphrodites produce VSPs only after exhausting their self-sperm, which occurs 5 to 6 d postadulthood in wild type individuals or 6 to 8 d in oac mutants. By contrast, virgin adult females of dioecious species continuously secrete these VSPs until mated (23???–27, 41). To identify candidate VSPs, we performed headspace solid-phase microextraction (SPME) coupled with GC-MS (Materials and Methods) on cultures of C. elegans and Caenorhabditis remanei strains at stages associated with pheromone production (Fig. 3 and SI Appendix, Fig. S2) (42).
Across oac mutant strains, egg laying persisted at low levels through Days 5-7, while wild type largely ceased by Day 5, indicating a modest but reproducible extension of the reproductive period (SI Appendix, Fig. S6 A and B). The brood-size reductions were allele specific: PS8113: oac-16(sy1341) (oac-16Δ) yielded fewer progeny on Day 2-3 and a smaller total brood than N2 (SI Appendix, Fig. S6 A and C). However, other CRISPR stop-in mutants, namely an oac-16 single mutant, oac-13; oac-16 double mutant, and oac-16; oac-25; oac-28 triple mutant, did not show this decrease, suggesting that the effect reflects a consequence of the deletion strategy or an off-target effect rather than oac-16 gene function. Somatic growth from 24 to 68 h after hatch—length, width, and area—did not differ from N2 (SI Appendix, Fig. S6 D–F). In summary, oac mutants exhibit a modest extension of reproduction with delayed self-sperm depletion, without detectable somatic growth defects. We collected pheromone after self-sperm depletion.
Pheromone-positive samples have that were tested included 1) 6-d-old adult hermaphrodites of wild type C. elegans N2 strain, which were self-sperm depleted; 2) 6-d-old adults of C. elegans daf-22(m130) mutants, defective in ascaroside biosynthesis, to control for contributions from ascaroside signals in GC-MS; and 3) 1-d-old virgin adult females of wild type C. remanei, a dioecious species (Fig. 3B and SI Appendix, Fig. S3). Pheromone-negative controls included 1) 1-d-old adult C. elegans hermaphrodites, retaining self-sperm; 2) C. elegans larvae (L2–L4 stages), representing nonreproductive developmental stages; 3) C. remanei adult males; and 4) mixed-sex 1-d-old adults of C. remanei, where mating should suppress the VSP production (Fig. 3C and SI Appendix, Fig. S4). GC-MS analyses revealed two conserved chromatographic peaks (Peaks 1/2) present exclusively in all pheromone-positive samples and absent in all negative controls. Despite the unresolved chemical identities of Peaks 1/2, the retention time (t_R_) alignment (Fig. 3B) and identical mass spectral profiles (Fig. 3A and SI Appendix, Fig. S3A) confirmed their identity across species and strains, suggesting these compounds represent conserved volatile chemicals associated with previously reported male attraction stages (Fig. 3D) (23???–27, 41).
OAC Gene Mutants Exhibit Loss of Volatile Sex Pheromone Signals in C. elegans.
To determine whether oac-13, oac-16, oac-25, and oac-28 influence VSPs production or secretion, we analyzed pheromone extracts from behaviorally deficient oac mutants using SPME-GC-MS. To quantify the results, we analyzed GC elution times, MS profiles, and integrated peak areas derived from extracted ion chromatograms (EICs) for analyte ions (m/z 67.0 and 82.0; Fig. 4 and SI Appendix, Fig. S5) (42).
Single mutants of oac-13 or oac-16 showed significant reductions in the abundance of Peak 1/2 compared to wild-type controls (Fig. 4). The oac-16 oac-13 double mutant exhibited no further reduction, consistent with behavioral assays, whereas the double mutants impaired male attraction to a similar degree as the single mutant (Fig. 2H). By contrast, the oac-28 oac-25 double mutant retained Peak 1 but nearly lost Peak 2 (Fig. 4B). While the remaining male attractiveness in oac-28 oac-25 double mutants suggest that both peaks retain male attractiveness (Fig. 2H). The oac-16 oac-28 oac-25 triple mutant lost both Peak 1/2, matching the pheromone-negative controls GC-MS data such as C. elegans L4 and 1-d-old adults (Fig. 4B).
Behavioral assays and biochemical evidence suggest distinct roles of those four OAC genes: oac-13/oac-16 act upstream, influencing a precursor required for both Peak 1/2, while oac-25/oac-28 function downstream, specifically regulating Peak 2. The lack of additive effects in oac-16 oac-13 double mutants implies they act in a common pathway or complex, with both paralogs essential for synthesizing Peaks 1/2. On the other hand, oac-25/oac-28 have identical protein sequences and are in close chromosomal proximity, which likely share redundant roles in Peak 2 production.
oac-16, oac-25, and oac-28 Expression Pattern and Stage.
Transcriptional reporters for oac-16, oac-25, and oac-28 were generated to characterize their spatiotemporal expression. All three genes were expressed in hermaphrodite seam cells (specialized lateral line epidermal cells), with oac-16 and oac-28 showing constitutive expression from embryogenesis to adulthood. In contrast, oac-25 expression was restricted to postembryonic stages, suggesting stage-specific regulation (Fig. 5). The images of the embryo, L1, and L2 stages shown here were captured from the same individual worm throughout its early development (Fig. 5). No differences in expression were detected between hermaphrodites and males, implicating either additional biosynthetic enzymes or sex-specific substrates in VSP production. Technical challenges precluded the generation of a transcriptional reporter for oac-13, potentially due to sequence assembly issues in its promoter region sequence. Despite this limitation, the robust seam cell expression of oac-16, oac-25, and oac-28 supports a model in which VSPs are synthesized in seam cells.
Volatile sex pheromone related OAC stage specific expression pattern. We constructed transcriptional reporters by cloning the oac-16 (PS10804), oac-25 (PS10805), and oac-28 (PS10806) promoters upstream of gfp in the pPD95.75 vector (Materials and Methods). All three OAC genes are expressed in seam cells. The embryo, L1, and L2 images shown here are from the same individual worm. Stable extrachromosomal lines were established.
Discussion
Genetic Redundancy and Functional Roles of oac-13/16 and oac-25/28 in Volatile Sex Pheromone Production.
Our findings demonstrate that four OAC family genes—oac-13, oac-16, oac-25, and oac-28—are required for the production of volatile sex pheromones. These genes may act by influencing secretion pathways, limiting the availability of essential biosynthetic precursors, or altering the metabolic supply of key nutrients necessary for pheromone generation.
The first model proposes that OAC-13 and OAC-16 mediate production of a shared or distinct precursor utilized in both Peak 1/2 pathways, while OAC-25 and OAC-28 act downstream to specifically convert this precursor into Peak 2. There are two possible explanations for the needs of both OAC-13 and OAC-16. First, the phenotypic similarity between single and double mutants suggests these genes may act sequentially in a shared biochemical pathway. For example, OAC-16 might depend on a substrate produced by OAC-13, or vice versa, therefore, disrupting either pauses the pathway. Alternatively, OAC-13 and OAC-16 may need to form a heteromultimer to enable precursor synthesis. Our second model suggests that OAC-25/OAC-28 might help maintain minimal Peak 1 production, as losing OAC-13/OAC-16 alone reduces but does not eliminate Peak 1, whereas the triple mutant (lacking OAC-16, OAC-25, and OAC-28) loses Peak 1 entirely. OAC-25 and OAC-28 are nearly identical paralogs, likely arising from a recent tandem duplication event in C. elegans. Such sequence identity (100%) and genomic proximity—only 6.4 kb apart on the reverse strand of Chromosome I (Fig. 1E)—strongly suggest that these genes are functionally redundant. Genes created by recent duplications often serve as genetic “backups,” providing robustness to essential pathways and buffering against deleterious mutations (43, 44). In principal, they can safeguard an essential process (45), consistent with broader principles of genetic robustness (46?–48).
Gene Duplication in Caenorhabditis Nematodes.
Using BLAST starting with the OAC-25 and OAC-28 protein sequence, we identified homologs across Caenorhabditis species and identified hits in C. remanei, Caenorhabditis briggsae, Caenorhabditis brenneri, and Caenorhabditis nigoni. No identical proteins were found in closely related species; the closest match was the C. briggsae protein CBG10742 (54.47% sequence identity). We also used BLAST of OAC-13 and OAC-16 but found no identical proteins and no protein similarity over 60%. While C. remanei produces VSPs indistinguishable from those of C. elegans by GC-MS retention time and mass spectrometry, we found no highly conserved orthologs of oac-13, oac-16, oac-25, or oac-28 in its genome. This genetic divergence suggests that conserved pheromone components are generated via functionally convergent pathways or a shared catalytic motif.
To characterize the evolutionary duplication event, we performed sequence analysis of 18 orthologous proteins showing >92% query coverage and >50% amino acid similarity to OAC-25/OAC-28. As demonstrated in SI Appendix, Fig. S7, these proteins display conserved duplication patterns across multiple species. In C. remanei, we identified two highly conserved paralog pairs (96.2% and 98% identical). C. briggsae showed even greater conservation, with one paralog pair exhibiting 99.6% similarity and three additional paralogs demonstrating near-perfect identity (100% and 99.6%). All mapped genes are located near their paralogs in the genome. There are three genes that have not been mapped yet. These levels of sequence conservation strongly suggest recent, lineage-specific gene duplication events in both species. In our analysis, only the double mutant of oac-25 and oac-28 affects the VSPs synthesis, consistent with the dual role of gene duplication in evolution: maintaining stability while enabling innovation (44, 49, 50). Future investigations in nematodes across the evolutionary tree-derived species (both ancestral to and descendant from the C. elegans lineage) could clarify whether OAC-25 and OAC-28 function as redundant backups or subfunctionalized intermediates, which would elucidate their evolutionary trajectory and distinguish between conserved redundancy (shared ancestral function) and lineage-specific specialization (51).
Volatile Sex Pheromone Biosynthesis Location.
Prior work established that laser ablation of Z1 and Z4—precursor cells of the somatic gonad (52)—abolishes hermaphrodite male attractiveness, while ablation of germline precursors Z2 and Z3 has no effect, implicating the somatic gonad in volatile pheromone (VSP) biosynthesis (24). Consistent with this finding, vulvaless hermaphrodites retain wild-type attractiveness, ruling out the vulva as a secretion opening for VSPs (25). Our expression analysis of oac-16, oac-25, and oac-28 localizes these genes to seam cells (Fig. 5). We thus infer that the seam cells are a crucial site of VSP biosynthesis and might be the source of secreted VSP.
The somatic gonad of C. elegans hermaphrodites comprises eight distinct structural regions, including connections to the vulva and seam cells of the lateral epidermis, the uterus, spermathecal valve, spermatheca, oviduct, ovary sheath, and the distal tip cell (53, 54). Notably, the uterine-seam (utse) syncytium forms a critical physical linkage between the somatic gonad and hypodermal seam cells during the mid-to-late L4 stage (53, 55). These anatomical and developmental insights contextualize our findings on VSPs biosynthesis. The L4-stage establishment of utse cell connectivity coincides with the onset of VSP production (24), raising the possibility that somatic gonadal signaling to seam cells—mediated through this physical linkage—may regulate pheromone synthesis. While VSPs are not produced in the gonad itself, the gonad’s structural integration with seam cells (via the utse cell) may provide developmental cues to modulate OAC gene activity in seam cells. For instance, the gonad could relay other metabolic or temporal signals during late larval stages to initiate or terminate VSPs production. Future work could assess whether disrupting utse-seam cell connectivity (e.g., via mutations in adhesion genes) ablates VSPs synthesis, linking morphogenesis to pheromone regulation.
The lateral position and established secretory functions of the seam cells make them exceptionally well-suited for synthesizing and secreting volatile pheromones. Embedded within the hypodermis but exposed to the external environment via their apical surface directly underlying the cuticle (including the alae) (56?–58), seam cells possess the physical location necessary for efficient volatile compound release. Furthermore, their documented capacity for synthesizing and secreting stage-specific cuticular components, such as collagens and the specialized alae structures (59), demonstrates robust biosynthetic and secretory machinery. This existing molecular infrastructure for producing extracellular molecules aligns with a role in VSP biosynthesis and direct secretion into the surrounding environment.
Volatile Sex Pheromone Chemical Identity.
We propose that oac genes catalyze acyl transfer reactions to modify hydrocarbon precursors. The products of this reaction are likely short-chain esters (e.g., acetate or propionate derivatives). This hypothesis aligns with the observed properties of Peaks 1/2, which exhibit chromatographic behaviors and mass spectra consistent with low-molecular-weight acylated hydrocarbons. However, the identities of these peaks remain unresolved, preventing precise pathway reconstruction. This challenge arises from their volatility, low abundance, structural isomerism, and potential branching of hydrocarbon chains. A key limitation of our study is the reliance on GC-MS alone, which lacks the resolution to distinguish between closely related isomers. Similar challenges in insect pheromone studies were overcome using isotopic tracing and heterologous expression in E. coli, yeast, or cell systems, which may resolve the biosynthetic steps (60??–63). Synthesis of candidate chemicals and bioassays could then validate their bioactivity as pheromones, bridging the gap between enzymatic activity and biosynthesis pathway reconstruction.
Conservation of Volatile Sex Pheromones across Nematode Lineages with Divergent Reproductive Strategies.
Peaks 1/2 were detected in pheromone-positive samples of both C. elegans (androdioecious, hermaphrodites/males) and C. remanei (dioecious, females/males), despite their phylogenetic divergence (~100 My) and divergent sexual systems. The presence of Peaks 1/2 in both suggests these VSP signals evolved early before their sexual systems diverged. A key question is how species avoid cross-attraction. Spatial separation of populations, differences in pheromone concentration and components ratio, or other species-specific ascarosides may help maintain reproductive isolation (64??–67). Further comparing Peaks 1/2 across nematode species could reveal whether their ratios or combinations drive speciation.
Evolutionary Implications of Gene Family Expansion.
Complete loss-of-function alleles are the gold standard for genetic studies. O-acyltransferases are used in many different pathways throughout the cells of eukaryotic animals, and we now have the tools to elucidate their functions in C. elegans. Our creation of null mutants for the entire OAC gene family allows for future studies of individual genes or the family as a whole. It is particularly interesting to study why this gene family is largely expanded in Caenorhabditis and how they contribute to species-specific pheromone signaling and lead to species differentiation.
All identified oac genes associated with VSPs synthesis are clustered closely on chromosome I and exhibit close phylogenetic relationships. oac-4 is evolutionarily related to those four VSPs-synthesis-associated OAC proteins with high sequence similarity (Fig. 1A). In contrast, oac-4 is the sole gene not required for VSPs synthesis and is located on a distinct chromosome. Sequence analysis of the oac-4 shows a unique insertion that contains two repeat HWPIYA[F/H] fragment insertions in the SYH catalytic motif (See SI Appendix, Fig. S1; highlighted with pink), a feature absent in all other 58 C. elegans OAC family proteins. These insertions may disrupt oac-4’ s ability to synthesize VSPs, suggesting potential repurpose of this gene for novel functional roles.
The Caenorhabditis genus’s OAC expansion contrasts with other nematodes, suggesting lineage-specific adaptations. The conserved SYH catalytic motif across OACs indicates a shared enzymatic mechanism, while domain architectures (e.g., SGNH hydrolase or NRF domains) suggest divergent functions. For instance, oac-13 and oac-16 both essential for Peak 1/2 may reflect neofunctionalization after duplication, and two identical genes oac-25 and oac-28 ensure the peak 1 production may reflect gene function redundancy. All four OAC genes involved in the same VSPs synthesis pathway are evolutionally closed evidenced by phylogenetic clustering and sequence alignment; also their genomic loci are remarkably close.
Materials and Methods
Protein Alignment.
Protein alignments were performed using the ClustalOmega multiple sequence alignment tool with default settings. Protein sequences were obtained from WormBase (WS295; https://wormbase.org).
Phylogenetic Analysis of OAC Proteins.
Protein sequences of C. elegans OAC family members were retrieved from WormBase WS295. The multiple sequence alignment was performed using the MUSCLE algorithm in MEGA (Molecular Evolutionary Genetics Analysis, version 12) with default parameters. The alignment was trimmed to exclude positions containing gaps or missing data, retaining conserved domains to ensure phylogenetic robustness. A maximum likelihood (ML) phylogeny was inferred using the Tamura-Nei model (68). Tree topology was refined using the Nearest-Neighbor-Interchange (NNI) heuristic method, with initial trees generated automatically via default neighbor-joining (NJ) and maximum parsimony (MP) algorithms. Branch lengths represent substitutions per site, and the final tree was visualized and annotated in MEGA.
C. elegans Strain Construction and Maintenance.
The Bristol N2 strain was used as a wildtype for C. elegans. Ascaroside biosynthetic defective mutant strain, DR476: daf-22(m130) was used as a control in this study. The rhy-1, bus-1, and nrf-6 strains were obtained from the Caenorhabditis Genetics Center (CGC). SI Appendix, Table S4 shows a list of all strains used in this paper. All nematode strains were kept on NGM agar plates seeded with E. coli (OP50) at 20 °C.
The creation of deletion mutants by nonhomologous end joining was done using a modification of the method described in Köhler et al. (2017) (69) in which only one guide RNA was used. SI Appendix, Table S2 gives the flanking sequences around the deletions and the primers used in the creation of strains using the single-primer deletion method.
Creation of triple-stop knock-in mutants was done using the universal STOP-IN cassette method as described in Wang et al. (70). SI Appendix, Table S3 gives the flanking sequences around the insertions and the primers used in the creation of strains using the triple-stop knock-in method.
All oac-16 alleles analyzed in this study—sy1471, sy2301, sy2302, sy2313, sy2314—are independent insertions of the same CRISPR “stop-in” loss-of-function cassette as PS8577 (sy1471). Because oac-16 is tightly linked to oac-13 and to oac-25/28, conventional crossing cannot reliably recover recombinant chromosomes. To ensure matched comparators, we therefore reintroduced the identical stop-in cassette into the oac-13 and oac-25/28 mutant backgrounds, yielding independent insertion lines. The updated allele-prefix designations indicate independent origins (and thus potential background differences), rather than distinct alleles. All insertions were sequence-verified to be identical at the nucleotide level. Consequently, genetic interaction analyses are benchmarked against the same oac-16 loss-of-function allele throughout.
Volatile Sex Pheromone Extraction.
VSPs were extracted from synchronized C. elegans hermaphrodites and C. remanei females as described in Wan et al. 2024 with minor modifications (26). For the chemoattraction assay, use 100 µL of solution containing 100 C. elegans. For each GC–MS analysis, 2 mL of VSP extract was used per sample. Extracts were prepared from 100 µL of C. elegans worm pellet soaked in 2 mL of M9 buffer. For C. remanei GC–MS analyses, samples were prepared from 120 individuals soaked in 2 mL of solution.
Worms were synchronized via bleach lysis, washed in M9 buffer. The extraction day was determined by the exhaustion of self-sperm, with no more new-born progeny presenting on the NGM plate. On the extraction day, worms were incubated for 6 h in M9 to allow pheromone accumulation. Following centrifugation (15,000× g, 30 to 60 s), the supernatant containing pheromones was stored at −80 °C.
Chemoattraction Assay and Two-Choice Assay.
Male chemotaxis responses were quantified using a chemoattraction assay as described in Wan et al. (26). Briefly, 2 µL of pheromone extract or M9 control was applied to designated spots alongside 1 M sodium aside to immobilize arriving males. 20 one-day-old adult males were released at the starting point, and chemotaxis indices (C.I.) were calculated after 30 min using C.I. = [(E − C)/(E + C + N)]. E, C, and N represent worms at experimental, control, or neither spot, respectively. The two-choice assay was performed using the same setup as the chemoattraction assay. The M9 control buffer was replaced with wild type N2 C. elegans sex pheromone extract. Chemotaxis indices (C.I.) were calculated using C.I. = [(WT_VSP – OAC_VSP Mutant)/(WT_VSP + OAC_VSP Mutant +N)]. WT_VSP, OAC_VSP Mutant, and N represent worms at wildtype pheromone, oac mutant pheromone, or neither spot, respectively.
Solid-Phase MicroExtraction (SPME).
Volatile compounds were collected from hermaphrodite cultures using headspace solid-phase microextraction (SPME). A divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS, 50/30 µm) fiber (Supelco, Bellefonte, PA) was preconditioned for 10 min at 265 °C in the GC injector before each use. For sampling, the fiber was exposed to the headspace of sealed culture vials for 10 min at 60 °C under continuous agitation (250 rpm, 5 s-on /20 s-off cycles) using a CTC Analytics autosampler. After extraction, fibers were immediately desorbed in the GC injector for 6 min.
Gas Chromatography-Mass Spectrometry (GC-MS).
Analyses were performed on an Agilent 6890 N gas chromatograph coupled to a Waters GCT Premier orthogonal acceleration time-of-flight mass spectrometer (oaTOF-MS). Separation was achieved using a DB-624 capillary column (30 m × 250 µm × 1.40 µm; Agilent). Helium carrier gas was maintained at a constant flow rate of 0.80 mL/min. The oven temperature program was initiated at 25 °C (0 min hold), ramped at 5 °C/min to 85 °C (0 min hold), then ramped at 1 °C/min to 150 °C (0 min hold), followed by a 30 °C/min ramp to 250 °C (1.5 min hold). Electron ionization positive (EI+) was performed at 70 eV, with mass spectra acquired in positive ion mode over a range of m/z 35 to 500. Blank runs (SPME fiber exposed to miliQ H_2_O sample vials) were interspersed with samples to monitor carryover. System performance was validated daily using alkane standard mixtures (C7–C30). All biological replicates (n = 5 to 8 per condition) were analyzed in randomized order to minimize batch effects.
GC-MS Data Acquisition and Quantification.
Raw data were processed using Waters MassLynx software. Peaks 1/2 (putative pheromone components) were quantified by integrating ion chromatograms (m/z 67.0 ± 0.3 and m/z 82.1 ± 0.3) within a retention time window of ±0.4 min around predicted values (t_R_ = 31.1 min and 32.1 min, respectively). External calibration curves were generated using absolute peak areas, with baseline correction and noise thresholds applied uniformly across samples Peaks 1 and 2 were identified across samples based on retention time alignment (±0.1% tolerance) and spectral matching.
GC–MS data are presented in two parts to link identification with quantification. Representative chromatograms with matched EI mass spectra identify Peak 1 and Peak 2; extracted-ion chromatograms (EICs) for the major diagnostic ions m/z 82.0, color-coded (yellow, Peak 1; blue, Peak 2), and annotated with retention times (Fig. 4A). Quantification was performed by integrating EIC peak areas within the corresponding retention windows for each sample, and summary statistics across 5 to 8 independent samples per condition are reported as mean ± SEM. (Fig. 4B). This presentation directly connects qualitative peak assignments to the quantitative measurements.
Brood Size Assay.
We synchronized worms by picking hermaphrodites at the early L4 stage. For each strain, five L4 animals were placed on a seeded NGM plate (OP50) and maintained 24 h at 20 °C to reach young adulthood. Six plates were set up in parallel (six biological replicates; five worms/plate, 30 worms in total). Beginning on Day 1, the five adults on each plate were transferred daily to a fresh NGM/OP50 plate at the same time of day. Brood size for each five-worm sample plate was scored as the number of progeny (embryos and larvae) present on the plate left behind from the previous 24 h interval; per-worm values were calculated by dividing by five and plotted accordingly. Daily counts continued for 7 d (or until egg-laying ceased). The total brood size per sample was computed as the sum of daily counts across Days 1 to 7; per-worm values were calculated by dividing by five and plotted accordingly. All plates were incubated at 20 °C, and transfers/counting were performed at consistent times to minimize handling effects.
Developmental Growth Measurements.
Synchronized worms were generated by a 1-h egg-lay window. Briefly, ~100 early L4 stage hermaphrodites were transferred to a seeded NGM plate and maintained for 24 h at 20 °C to obtain young adults. From these, 20 young adult hermaphrodites were placed onto each of three fresh NGM plates (OP50), allowed to lay eggs for 1 h, and then removed. The resulting progeny were considered 0 h after egg deposition. Worms were imaged at 24 h and 36 h after egg deposition, and subsequently from 40 to 68 h after egg deposition at 4-h intervals. Time annotations on all plots refer to hours after egg deposition on the assay plates. Length, width, and body area were quantified from bright-field images for each time point; measurements were aggregated across the three plates (biological replicates). 70 worms are measured for each data point.
Image acquisition and morphometric analysis were performed using the WormLab system (hardware and WormLab software, MBF Bioscience). For each time point, plates were imaged under identical illumination and magnification; pixel size was calibrated with a stage micrometer. Animals were segmented in WormLab with a fixed threshold and size filter across all datasets. Body length (skeletonized midline length), body width (orthogonal maximum width), and body area (planar 2D area in µm^2^) were exported directly from WormLab.
Expression Pattern Imaging.
Transcriptional reporters were constructed in vector pPD95.75, driving GFP expression from the oac-16p, oac-25p, or oac-28p promoters (PS10804: him-5; syEx2004[pPD95.75::oac-16p::gfp], PS10805: him-5; syEx2005[pPD95.75::oac-25p::gfp], PS10806: him-5; syEx2006[pPD95.75::oac-28p::gfp]). Plasmid DNA (80 ng/µL) was coinjected with the coelomocyte marker unc-122p::RFP (20 ng/µL) into him-5(e1490) mutants to obtain males for imaging. Transgenic lines expressing these extrachromosomal arrays were isolated, maintained, and imaged.
We cultured C. elegans in a 398-well plate format, with one embryo per well, to enable imaging at the embryo, L1, and L2 stages. Worms were immobilized using Levamisole, imaged within the wells, and then washed out, which prevented recovery of the same individuals for subsequent time points. Consequently, worms imaged at later stages (L3 to adult) were cultured separately using a different method optimized for those stages. Although this limits continuous tracking of the same individual throughout development, the use of stage-matched populations and consistent imaging conditions allows for reliable comparisons across stages. Consequently, worms imaged at later stages (L3 to adult) were cultured separately using a different method optimized for those stages. For these stages, animals were immobilized on a 4% ultrapure agarose pad in 5 mM Levamisole (in H_2_O) and imaged using a Zeiss AxioImager2 microscope equipped with a Colibri 7 LED fluorescence illumination system and an Axiocam 506 Mono camera (Carl Zeiss Inc.).
Supplementary Material
Appendix 01 (PDF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1S. J. Cook , Whole-animal connectomes of both Caenorhabditis elegans sexes. Nature 571, 63–71 (2019).31270481 10.1038/s 41586-019-1352-7PMC 6889226 · doi ↗ · pubmed ↗
- 2C. elegans Sequencing Consortium, Genome sequence of the nematode C. elegans: A platform for investigating biology. Science 282, 2012–2018 (1998).9851916 10.1126/science.282.5396.2012 · doi ↗ · pubmed ↗
- 3S. Brenner, The genetics of Caenorhabditis elegans. Genetics 77, 71–94 (1974).4366476 10.1093/genetics/77.1.71PMC 1213120 · doi ↗ · pubmed ↗
- 4J. Yoshimura , Recompleting the Caenorhabditis elegans genome. Genome Res. 29, 1009–1022 (2019).31123080 10.1101/gr.244830.118PMC 6581061 · doi ↗ · pubmed ↗
- 5P. W. Sternberg , Worm Base 2024: Status and transitioning to Alliance infrastructure. Genetics 227, iyae 050 (2024).38573366 10.1093/genetics/iyae 050PMC 11075546 · doi ↗ · pubmed ↗
- 6M. A. Tuli, A. Daul, T. Schedl, “Caenorhabditis nomenclature” in Worm Book: The Online Review of C. elegans Biology (The C. elegans Research Community, Pasadena, CA, 2018), pp. 2–14.10.1895/wormbook.1.183.1PMC 636958029722207 · doi ↗ · pubmed ↗
- 7M. Feuermann , A compendium of human gene functions derived from evolutionary modelling. Nature 640, 146–154 (2025).40011791 10.1038/s 41586-025-08592-0PMC 11964926 · doi ↗ · pubmed ↗
- 8A. A. Spector, S. N. Mathur, T. L. Kaduce, Role of acylcoenzyme A: Cholesterol o-acyltransferase in cholesterol metabolism. Prog. Lipid Res. 18, 31–53 (1979).42927 10.1016/0163-7827(79)90003-1 · doi ↗ · pubmed ↗